Introduction
Preeclampsia (PE) is characterized by hypertension
with or without proteinuria after the 20th week of pregnancy, and
is associated with endothelial dysfunction, oxidative stress,
systemic vasoconstriction, inflammation, intrauterine growth
restriction and multiorgan dysfunction (1,2). The
World Health Organization reports that >60,000 pregnant women
worldwide die of preeclampsia each year (3). Women with PE and their offspring are
also at increased risk of developing cardiovascular disease later
in life (4).
Although the pathogenesis of PE remains unclear,
increased oxidative stress in conjunction with antioxidative
defence mechanisms may contribute to the progression of PE.
Pregnancy itself increases susceptibility to oxidative stress,
which could result in tissue damage (5). However, higher production of
pro-oxidants and reactive oxygen species (ROS) is observed at the
later stages of normal uncomplicated pregnancies and maintained
through the accumulation of antioxidants, such as superoxide
dismutase (SOD), glutathione (GSH), tocopherols, carotenoids and
ascorbic acid (6). Nevertheless,
the balance between pro-oxidants, such as malondialdehyde (MDA),
and antioxidants, such as GSH and SOD, is disturbed in pregnancies
with PE (7); this can lead to
cellular dysfunction and death (8). Moreover, previous studies (9,10)
have found that abnormal placentation and shallow trophoblast
invasion stimulate placental oxidative stress, which leads to an
intravascular inflammatory response and endothelial dysfunction
(11). Although multiple sources
of ROS have been identified in the placenta (12), such as the reduction of antioxidant
activity and the lack of blood flow, few studies have examined the
role of mitochondrial dysfunction and ROS in the pathology of PE.
Mitochondrial dysfunction induces a reduction in oxygen consumption
and an increase in superoxide production (13).
Mitochondria play an important role in ATP synthesis
during aerobic respiration. During this process, ROS, such as the
superoxide anion, hydroxyl radical, hydroperoxyl and hydrogen
peroxide (H2O2), are formed as metabolites of
mitochondrial oxidative phosphorylation (14). Under conditions of oxidative
stress, mitochondria can synthesize ROS at a level that extensively
disrupts mitochondrial homeostasis, and alters the composition of
lipids, proteins and nucleic acids (15). The resulting disruption in the
composition of the mitochondrial membrane impairs the
electrochemical potential of the membrane, which leads to
mitochondrial dysfunction and apoptosis (16).
As mitochondrial damage can lead to apoptosis, the
timely elimination of excess ROS is important for cell survival.
Elevated levels of ROS trigger mitophagy, a cellular process in
which lysosomes selectively scavenge for and eliminate damaged
mitochondria (17). Under
conditions of oxidative stress, mitophagy is upregulated to prevent
the accumulation of dysfunctional mitochondria (18,19).
Thus, it was hypothesized that mitochondrial dysfunction regulated
by mitophagy plays an important role in PE.
DNA damage-regulated autophagy modulator 1 (DRAM1)
is an evolutionarily conserved transmembrane protein (20). This protein was previously
identified as the first direct molecular link between the tumor
suppressor p53 and autophagy (21). Since then, various roles for DRAM1
in several processes, including autophagy, cell death, immunity and
cellular differentiation, have been described (21,22).
As stated above, elevated oxidative stress caused by mitochondrial
dysfunction leads to apoptosis, and mitophagy plays an important
role in the maintenance of mitochondrial function. However, neither
mitochondrial dysfunction nor mitophagy have been examined in PE,
and there is limited understanding regarding the contributions of
mitochondrial dysfunction and mitophagy to the development of PE.
Moreover, the causes of mitochondrial dysfunction and mitophagy in
PE are unknown. Therefore, the aim of the present study was to
evaluate the roles of mitochondrial dysfunction and ROS in the
development of PE, and to determine the role of mitophagy in
mitochondrial dysfunction in PE, in order to investigate whether
DRAM1 plays a role in mitophagy and mitochondrial dysfunction in
PE.
In the present study, it was demonstrated that
oxidative stress was induced by mitochondrial dysfunction and
decreased DRAM1 expression, and that increased mitophagy could
cause mitochondrial dysfunction in the PE placenta. Additionally,
it was demonstrated that the mitophagy dysfunction in PE might
result from disturbances in mitochondrial fusion and fission due to
a deficit in DRAM1. Altogether, the results of the present study
indicated a key role for DRAM1 in mitophagy that contributed to PE.
To the best of the authors' knowledge, this study provided the
first evidence that DRAM1 plays a role in PE, and offers a new
target for the treatment of PE and new insights into the
pathophysiological mechanisms of PE.
Materials and methods
Animals
A total of 80 C57BL/6J wild-type (WT) mice (age, 2
months; weight, 30–40 g) were purchased from Guangdong Medical
Laboratory Animal Center, and all the experiments were approved by
the Ethics Committee of Shenzhen Hospital of Peking University
(permit no. SYXK-2015-0106, 2019-078) and conducted in accordance
with the ARRIVE guidelines on the Care and Use of Experimental
Animals (23). Male and female
animals were fed separately and housed in groups of four to five.
All the mice were maintained under standard laboratory conditions
with a temperature of 22±2°C, 50±10% relative humidity and a 12-h
light/12-h dark cycle and provided with food and water ad
libitum. The fertility cycle of the mice was controlled, and
the animals were allowed to mate overnight when the females were in
a pro-estrus state. On the following morning at 8:00 a.m., vaginal
smears on glass slides were examined, and if spermatozoa were
found, that day was designated the first day of pregnancy,
embryonic day (E)0.5. Pregnant mice at E8 were randomly divided
into three groups (n=8/group) and injected systemically via the
tail vein with 100 µl Ad-CMV-HIF (8×1010 viral
particles) encoding hypoxia-inducible factor 1α (Hif-1α),
Ad-CMV-GFP (8×1010 viral particles) as a green
fluorescent protein (GFP)-encoding vehicle (Veh-GFP) or saline
solution [Shanghai OBiO Technology (Shanghai) Corp., Ltd.].
Blood pressure measurements
Systolic blood pressure of conscious mice was
measured on E0.5, E8.5, E12.5, E16.5 and E19.5 of gestation using
the tail cuff technique (24)
(Shanghai Yuyan Instruments Co., Ltd.) The mice were prewarmed at
37°C for 30 min before the measurements were taken. The mean from
five consecutive readings was recorded as the systolic blood
pressure.
Urinary albumin and serum lipid
measurements
Urine was collected on E16.5 and E19.5 of gestation
for albumin measurements, and the urinary albumin was quantified
using a bicinchoninic acid (BCA) protein assay with an Easy II
Protein Quantitative kit (cat. no. DQ111-01; Beijing Transgen
Biotech Co., Ltd.) according to the manufacturer's recommended
protocol. In brief, urine was collected from each mouse, and the
protein concentration was determined using a BCA protein assay. The
absorbance at 595 nm was measured using a microplate reader, and
BSA (Sigma-Aldrich; Merck KGaA) was used as the standard.
Blood was collected on E19.5 of gestation for serum
lipid measurements, and the level of serum lipids was quantified by
ultraviolet-visible spectrophotometry using the following
biochemical detection kits for blood lipids: Total cholesterol (TC;
cat. no. BC1985; Beijing Solarbio Science & Technology Co.,
Ltd.), triglyceride (TG; cat. no. BH017Z; Tellgen Corporation),
low-density lipoprotein (LDL; cat. no. BH019Z; Tellgen Corporation)
and high-density lipoprotein (HDL; cat. no. BH018Z; Tellgen
Corporation). All assays were performed according to the
manufacturer's protocols.
Tissue processing
For histological and biochemical analyses, the mice
were euthanized, and their placentas and kidneys were rapidly
removed. Some placentas were maintained at 80°C for biochemical
analysis, and the remaining placentas and kidneys were fixed with
4% (vol/vol) paraformaldehyde overnight at 4°C, dehydrated with
gradient sucrose (20 and 30%) for 24 h and embedded in optimal
cutting temperature compound. The embedded tissue was sagittally
cut into 30-µm-thick sections using a freezing-sliding microtome
(Leica Microsystems GmbH).
Histology
The obtained placental kidney tissue sections were
washed with PBS and used for hematoxylin and eosin staining, which
was performed according to the manufacturer's protocol (cat. no.
G1120; Beijing Solarbio Science & Technology Co., Ltd.). The
sections were visualized using a light microscope (Leica DM4 M;
magnification, ×20).
Oxidative stress measurements
To test the level of oxidative stress in the
placenta, different kits were used. The activity of SOD in
placental tissue was detected by nitro-blue tetrazolium (cat. no.
S0107; Beyotime Institute of Biotechnology). The levels of MDA were
tested by thiobarbituric acid (cat. no. BC0025; Beijing Solarbio
Science & Technology Co., Ltd.), and the levels of
H2O2 were tested by titanium sulfate (cat.
no. BC3595; Beijing Solarbio Science & Technology Co., Ltd.).
All assays were performed according to the manufacturer's
protocol.
Western blotting
Placental samples were homogenized in RIPA lysis
buffer (cat. no. P0013B; Beyotime Institute of Biotechnology),
containing 1 mM PMSF (Beyotime Institute of Biotechnology), a
protease inhibitor cocktail and phosphatase inhibitors (Roche
Diagnostics). The samples were centrifuged at 13,000 × g and 4°C
for 30 min, and the supernatants were collected. The protein
concentration was measured with a BCA kit (cat. no. DQ111-01;
Beijing Transgen Biotech Co., Ltd.). The protein samples (20 µg)
were loaded onto a 10% SDS-PAGE gel and then transferred onto
0.45-mm polyvinylidene difluoride membranes (EMD Millipore) at 90
mA for 1–2 h. After blocking with 5% nonfat milk for 1 h at 37°C,
the membrane was incubated with primary antibody (Table I) overnight at 4°C and horseradish
peroxidase-conjugated secondary antibodies (anti-mouse and
anti-rabbit; 1:500; cat. nos. 6919–100 and 6925-100; NeoBioscience)
for 1 h at 37°C. The immunoreactive bands were visualized with an
enhanced chemiluminescence kit (cat. no. A38555; Thermo Fisher
Scientific, Inc.) and scanned for densitometric analysis using an
imaging system (Image Station 4000 M; Kodak) and Quantity One
software v4.6.6 (Bio-Rad Laboratories, Inc.). GAPDH or β-actin were
used as the loading control.
 | Table I.Antibody information. |
Table I.
Antibody information.
| Antibody | Host species | Cat. no. | Dilution | Supplier |
|---|
| Bax | Rabbit | ab32503 | 1:2,000 | Abcam |
| Bcl-2 | Rabbit | ab185002 | 1:2,000 | Abcam |
| COX IV | Rabbit | 4850 | 1:1,000 | Cell Signaling
Technology, Inc. |
| PINK1 | Rabbit | ab216144 | 1:1,000 | Abcam |
| Parkin | Mouse | 4211 | 1:1,000 | Cell Signaling
Technology, Inc. |
| DRAM1 | Rabbit | PA5-69473 | 1:500 | Invitrogen; Thermo
Fisher Scientific, Inc. |
| Mfn1 | Rabbit | 14739 | 1:1,000 | Cell Signaling
Technology, Inc. |
| Mfn2 | Rabbit | 11925 | 1:1,000 | Cell Signaling
Technology, Inc. |
| OPA1 | Rabbit | ab157457 | 1:1,000 | Abcam |
| DRP1 | Rabbit | ab184247 | 1:1,000 | Abcam |
| GAPDH | Rabbit | ab181602 | 1:2,000 | Abcam |
| β-actin | Mouse | 3700 | 1:2,000 | Cell Signaling
Technology, Inc. |
In utero electroporation (IUE)
IUE was performed as previously described (25). Briefly, pregnant female PE mice
(induced by Hif1-α) at E14.5 were anesthetized by diluting ketamine
(100 mg/kg) and xylazine (10 mg/kg) with 0.9% saline. The abdomen
was cut, and the uterine horns were then carefully removed. Next,
~3 µg plasmid DNA (V5-DRAM1-pCAGEN-GFP or V5-pCAGEN-GFP; Biovector
Science Lab, Inc.) was mixed with 1.5 µl 0.025% Fast Green (cat.
no. F7252; Sigma-Aldrich; Merck KGaA), which was pressure-injected
into the junctional zone of the placenta by pulling the glass
capillaries. Five pulses of current (40 mV for 40 msec) were
injected into the placenta using an electroporator (BTX T830; BTX
Molecular Delivery Systems). The uterus was moved into the
peritoneal cavity, and the abdominal wall and skin were sutured.
The transfection efficiency was determined using immunofluorescence
and a confocal microscope (Nikon AR1; magnification, ×20).
Statistical analysis
Data were analyzed using GraphPad Prism v 7.04
software (GraphPad Software, Inc.). In animal experiments, pregnant
mice at E8 were randomly divided into three groups (n=8/group) to
determine the blood pressure and total urinary protein levels. To
investigate TG, TC, LDL and HDL levels, 4 samples were chosen. All
the data are expressed as mean ± SEM. One-way ANOVA followed by
Bonferroni's post hoc test was used for multiple comparisons among
the WT (injected with saline only), Veh-GFP and Hif-1α groups.
Unpaired Student's t-test was used when comparing the control and
DRAM1 groups. P<0.05 was considered to indicate a statistically
significant difference.
Results
Hif-1α induces PE in mice
To effectively evaluate whether DRAM1 plays an
important role in PE, a PE mouse model induced by Hif-1α was
established (26). The symptoms of
the PE mouse model were then evaluated by examining pathological
and physiological indicators associated with PE.
Hypertension is a defining feature of PE (27), and thus dynamic changes in blood
pressure were examined in PE mice during the progression of
pregnancy. The first measurement was taken at E0.5, and blood
pressure was found to increase during pregnancy. On both E16.5 and
E19.5, the blood pressure of the Hif-1α group was significantly
higher than that of the Veh-GFP and WT groups, and no significant
difference was found between the Veh-GFP and WT groups (Fig. 1A). The blood pressure of the mice
was also measured after delivery, which revealed that the blood
pressure began to decrease following delivery, with no significant
difference found between the three groups (Fig. 1A).
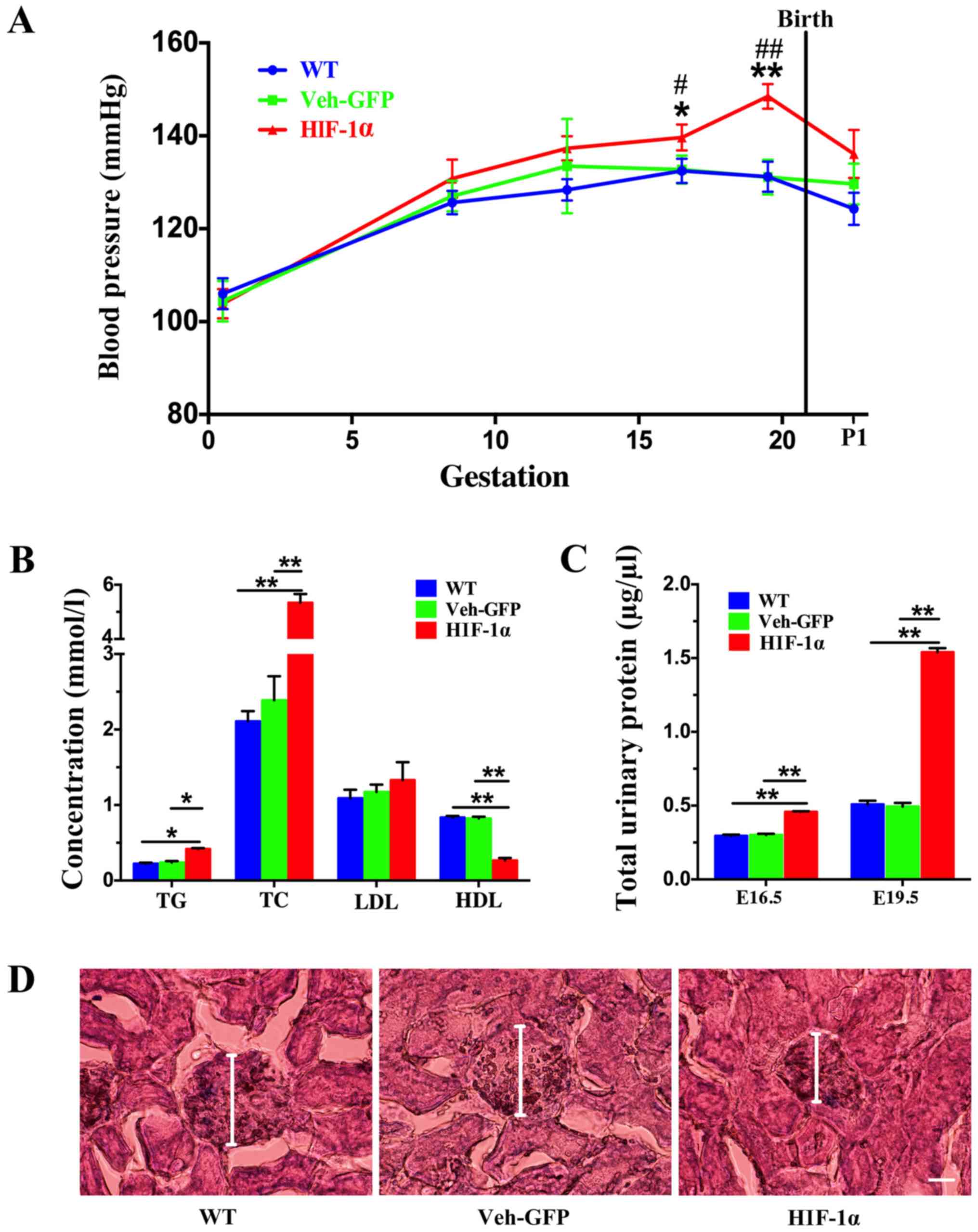 | Figure 1.Construction of a PE mouse model by
Hif-1α virus. PE mice exhibited elevated blood pressure, increased
urinary protein levels and impaired renal function. (A) Blood
pressure of the WT, Veh-GFP and Hif-1α mice was measured throughout
gestation at the indicated time points (n=8 per group). Data are
presented as the mean ± SEM from several mice used in each
experiment. *P<0.05 and **P<0.01 vs. Veh-GFP group;
#P<0.05 and ##P<0.01 vs. WT group.(B)
TG, TC, LDL and HDL levels in the serum of WT, Veh-GFP and Hif-1α
mice at E19.5 were measured (n=4 per group). Data are presented as
the mean ± SEM from several mice used in each experiment.
*P<0.05, **P<0.01. (C) Total urinary protein level in WT,
Veh-GFP and Hif-1α mice at E16.5 and E19.5 was determined using
bicinchoninic acid protein assays (n=8). **P<0.01. (D) Renal
histology of WT, Veh-GFP and Hif-1α mice at E19.5 demonstrated by
hematoxylin and eosin staining. Scale bar, 50 µm. PE, preeclampsia;
Hif-1α, hypoxia-inducible factor 1α; WT, wild-type; Veh-GFP,
vehicle-green fluorescent protein; E, embryonic day; TG,
triglyceride; TC, total cholesterol; LDL, low-density lipoprotein;
HDL, high-density lipoprotein. |
Dyslipidemia is closely related to the occurrence
and development of PE during pregnancy (1). TG, TC, LDL and HDL levels were
measured on E19.5. The results showed that the levels of TG and TC
in the Hif-1α group were significantly increased compared with
those in the WT and Veh-GFP groups (Fig. 1B), with no significant difference
detected between the WT and Veh-GFP groups. However, no significant
change in the LDL content was found between the Hif-1α group, and
the WT and Veh-GFP groups (Fig.
1B). In addition, the HDL content of the Hif-1α group was
significantly decreased compared with that of the WT and Veh-GFP
groups (Fig. 1B), and no
significant difference was detected between the WT and Veh-GFP
groups.
The presence of proteinuria, another defining
feature of PE, was measured on E16.5 and E19.5. Urine protein
content of the Hif-1α group was significantly increased compared
with that of the WT and Veh-GFP groups on both E16.5 and E19.5
(Fig. 1C). No significant
differences were found between the WT and Veh-GFP groups on either
E16.5 or E19.5 (Fig. 1C).
Kidney histology was assessed on E19.5, and it was
found that the WT and Veh-GFP mice showed no glomerular changes
compared with each other. Glomerular shrinkage was observed in the
Hif-1α mice relative to the WT and Veh-GFP mice (Fig. 1D). In addition, the Hif-1α mice
developed moderate swelling of glomerular endothelial cells and
mesangial cells with limited occlusion of the glomerular capillary
lumen (Fig. 1D).
The symptoms of hypertension, hyperlipidemia, high
urine protein and renal dysfunction observed in the Hif-1α mice
were consistent with pathological observations in patients with PE.
These results indicated successful establishment of a PE mouse
model that could be used in subsequent experiments.
Excessive oxidative stress in the
placentas of PE mice
The placenta is an important organ for the exchange
of substances between the fetus and the mother. To assess whether
the placental function of mice with PE was normal, the morphology
and oxidative stress levels of these mice were evaluated. During
the progression of gestation, the relative area of the junctional
zone (JZ) decreases as the labyrinth expands in size (28). Firstly, morphological changes in
the placentas of the mice were examined, and it was found that the
JZ region of the Hif-1α mice was increased compared with in the WT
and Veh-GFP mice on E19.5 (Fig.
2A). To determine the level of oxidative stress in the
placenta, the enzymatic activity of SOD, an important antioxidant
enzyme, was measured. The results showed that the activity of SOD
in the Hif-1α group was significantly reduced compared with that of
the WT and Veh-GFP groups, and no significant difference was found
between the WT and Veh-GFP groups (Fig. 2B). The levels of MDA were also
examined. MDA is a natural product of lipid oxidation in organisms
and reflects levels of oxidative stress (24). It was found that the MDA content in
the Hif-1α group was significantly increased compared with in the
WT and Veh-GFP groups, and no significant difference was detected
between the WT and Veh-GFP groups (Fig. 2C). In addition, levels of
H2O2, the most common active oxygen molecule
in organisms, were measured. It was shown that
H2O2 levels were increased significantly in
the Hif-1α group compared with the WT and Veh-GFP groups, and that
no significant difference was found between the WT and Veh-GFP
groups (Fig. 2C). These findings
suggested the presence of excessive oxidative stress in the
placentas of PE mice.
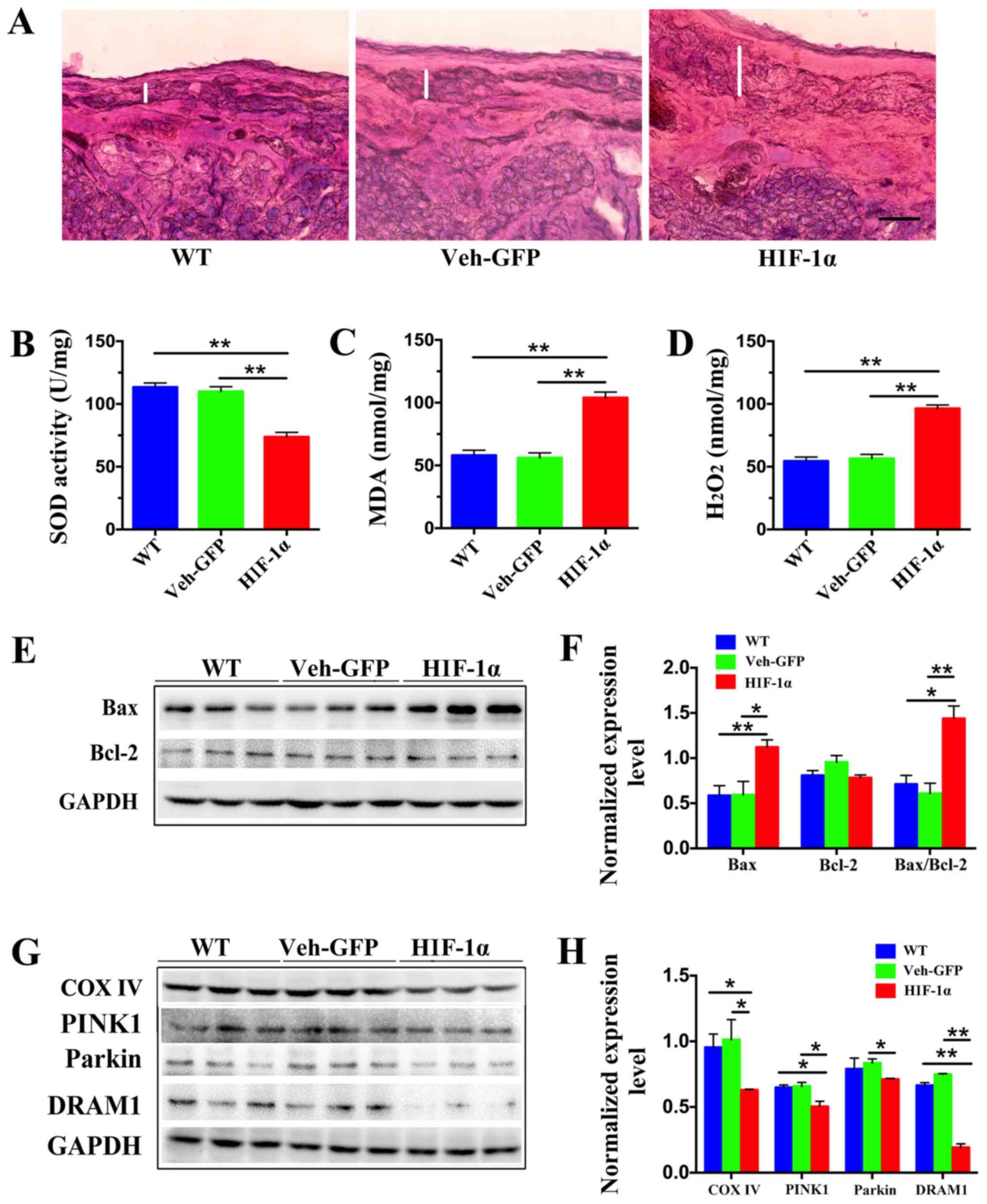 | Figure 2.Levels of oxidative stress, apoptosis
and mitochondrial autophagy are increased in the placenta of
preeclampsia mice. (A) Morphological examination of mouse placenta
at embryonic day 19.5 by hematoxylin and eosin staining revealed
that the junctional zone (represented by the white line) of Hif-1α
mice was larger than those of WT and Veh-GFP mice. Scale bar, 100
µm. (B) Activity of SOD in placental tissue was detected by
nitro-blue tetrazolium, (C) levels of MDA were measured using
thiobarbituric acid and (D) levels of H2O2
were measured using titanium sulfate. **P<0.01 (n=4). (E)
Representative western blot images and (F) semi-quantification of
Bax and Bcl-2 in the placenta of WT, Veh-GFP and Hif-1α mice.
Densitometry results were normalized against the levels of GAPDH.
(G) Representative western blot images and (H) semi-quantification
of COX IV, PINK1, Parkin and DRAM1 in the placentas of WT, Veh-GFP
and Hif-1α mice. Densitometry results were normalized against the
levels of GAPDH. Values are expressed as the mean ± SEM (n=3).
*P<0.05, **P<0.01. Hif-1α, hypoxia-inducible factor 1α; WT,
wild-type; Veh-GFP, vehicle-green fluorescent protein; SOD,
superoxide dismutase; MDA, malondialdehyde; COX IV, cytochrome
c oxidase IV; PINK1, PTEN-induced putative kinase 1; DRAM1,
DNA damage-regulated autophagy modulator 1. |
Mitochondrial dysfunction in the
placenta of PE mice leads to apoptosis
High levels of oxidative stress usually trigger
inflammation and apoptosis (29).
There is a close relationship between oxidative stress and
mitochondrial dysfunction, which can lead to the aggravation of
oxidative stress (30). The levels
of apoptosis and mitochondrial function were therefore
examined.
Firstly, to investigate whether placental cells were
growing normally, the expression of proteins associated with
apoptosis was examined. Bax is one of the early-acting proapoptotic
factors, and mainly exists in the cytoplasm and on the surface of
mitochondria (31,32). Bcl-2 is a classical gene that
inhibits apoptosis (33);
specifically, Bcl-2 is increased during the antioxidant status of
cells, thereby protecting them from oxidative stress, which plays a
certain role in reducing apoptosis, and it is mainly distributed in
the mitochondrial membrane (31).
Therefore, the levels of Bax and Bcl-2 and the ratio of Bax to
Bcl-2 can be detected as a representation of the level of
apoptosis.
Western blotting was used to assess the levels of
Bax and Bcl-2. Bax protein expression in the Hif-1α group was
significantly increased compared with those in the WT and Veh-GFP
groups, and no significant difference was found between the WT and
Veh-GFP groups (Fig. 2E and F). In
addition, no significant difference in the protein expression of
Bcl-2 was detected between the WT, Veh-GFP or Hif-1α groups
(Fig. 2E and F). The Bax/Bcl-2
ratio found to be significantly increased in the Hif-1α group
compared with the WT and Veh-GFP groups, whilst there was no
significant difference between the WT and Veh-GFP groups (Fig. 2E and F).
Mitochondria are the core of energy metabolism, and
the growth of a fetus requires a large amount of ATP from the
placenta (34). Therefore,
mitochondrial dysfunction is an important factor that could affect
PE. The maintenance of energy metabolism, the regulation of changes
in mitochondrial function or adaptation to environmental
stimulation, the generation of new mitochondria and the clearance
of defective mitochondria are critical for the maintenance of
mitochondrial health in eukaryotic cells (35,36).
In mitochondria, energy is mainly synthesized by the electron
transport chain, and cytochrome c oxidase IV (COX IV) plays
an important role in this (37).
Therefore, protein expression levels of COX IV in placental tissue
were measured by western blotting. This showed that COX IV protein
expression in the Hif-1α group was significantly lower compared
with in the WT and Veh-GFP groups (Fig. 2E and F), and that there was no
significant difference between the WT and Veh-GFP groups. These
results suggested that apoptosis induced by oxidative stress in the
placentas of PE mice may be caused by mitochondrial
dysfunction.
Decreased DRAM1 expression in the
placentas of PE mice leads to mitophagy dysfunction
Mitophagy is the most important pathway for the
clearance of damaged mitochondria. An important mitophagy pathway
is the PTEN-induced putative kinase 1 (PINK1)/E3 ubiquitin ligase
(Parkin) regulatory pathway (17).
Protein aggregates and damaged organelles are ubiquitinated to
initiate selective autophagy. For the initiation of mitophagy,
phosphorylated PINK1 is ubiquitinated to activate Parkin; this
process establishes a ubiquitin chain on outer mitochondrial
membrane proteins, recruits autophagy receptors and triggers
mitophagy (38,39). Therefore, western blotting was
performed to detect the protein expression of PINK1 and Parkin.
This showed that the level of PINK1 in the Hif-1α group was
significantly lower compared with in the WT and Veh-GFP groups, and
that there was no significant difference between the WT and Veh-GFP
groups (Fig. 2G and H). Parkin
protein expression in the Hif-1α group was significantly decreased
compared with that of the Veh-GFP group, and no significant
difference was found between the WT and Veh-GFP groups (Fig. 2G and H).
The DRAM1 protein, which plays an important role in
autophagy, was examined and it was discovered that DRAM1 protein
expression in the Hif-1α group was significantly decreased compared
with in the WT and Veh-GFP groups, and no significant difference
was detected between the WT and Veh-GFP groups (Fig. 2G and H).
These findings suggested that mitochondrial
dysfunction may be due to the inability of damaged mitochondria to
be cleared, and that the loss of DRAM1 may be related to mitophagy
dysfunction.
Overexpression of DRAM1 significantly
improves the symptoms of PE mice
To further investigate whether PE is affected by
DRAM1, a DRAM1 overexpression plasmid was transferred into the
trophoblasts of the placentas of PE mice on E14.5 via embryonic
electroporation (Fig. 3A). Protein
expression of DRAM1 was then tested by western blotting to verify
the overexpression of DRAM1. It was demonstrated that the level of
DRAM1 in the DRAM1 group was significantly increased compared with
that in the control group (Fig. 3C and
D). This result indicated that a model of DRAM1 overexpression
was successfully established.
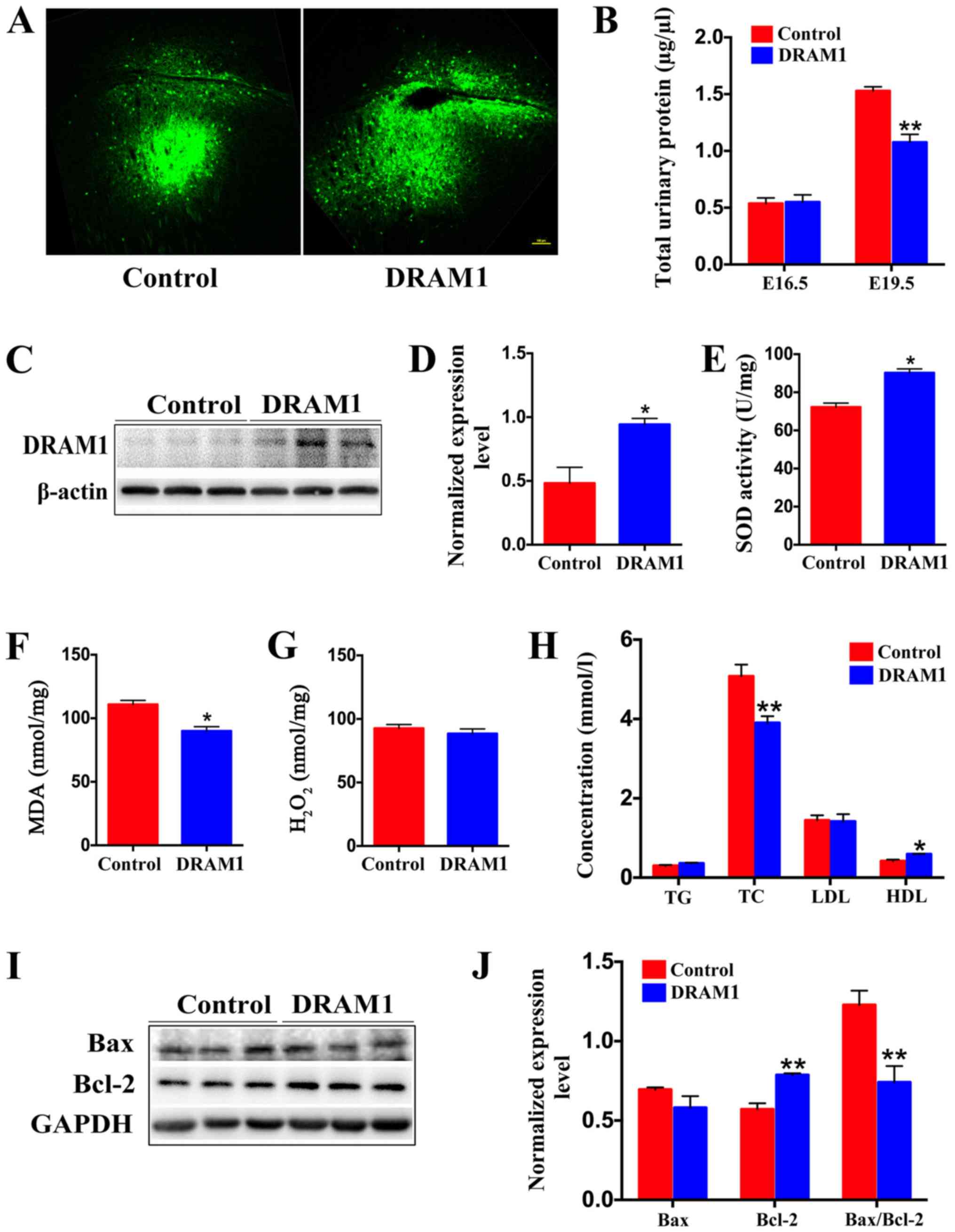 | Figure 3.Overexpression of DRAM1 can
effectively reduce apoptosis induced by oxidative stress and
significantly reduce the urine protein and blood lipid levels in
preeclampsia. (A) Representative images showing the overexpression
of DRAM1 or control in the placenta of mice at E19.5. Scale bar,
100 µm. (B) Bicinchoninic acid protein assays were used to
determine the total urinary protein level in the control and DRAM1
mice at E16.5 and E19.5 (n=6). (C) Representative western blot
images and (D) semi-quantification of DRAM1 levels in the placenta
of DRAM1-overexpressing or control mice at E19.5 (n=3).
Densitometry results were normalized against the levels of GAPDH.
Levels of placental oxidative stress in DRAM1 mice was
significantly decreased. (E) SOD in placental tissue was detected
by nitro-blue tetrazolium, (F) levels of MDA were measured using
thiobarbituric acid and (G) levels of H2O2
were measured using titanium sulfate (n=3). (H) TG, TC, LDL and HDL
levels in the serum of control and DRAM1 mice at E19.5 (n=6). (I)
Representative western blot images and (J) semi-quantification of
Bax and Bcl-2 in the placenta of control and DRAM1 mice.
Densitometry results were normalized against the levels of GAPDH
(n=3). Data are presented as the mean ± SEM. *P<0.05,
**P<0.01. DRAM1, DNA damage-regulated autophagy modulator 1; E,
embryonic day; SOD, superoxide dismutase; MDA, malondialdehyde; TG,
triglyceride; TC, total cholesterol; LDL, low-density lipoprotein;
HDL, high-density lipoprotein. |
To evaluate the effect of DRAM1 overexpression on
PE, the total amount of urine protein was measured in the mice. The
results showed that there was no significant difference between the
DRAM1 and control groups on E16.5, but that the level of urinary
protein in the DRAM1 group was significantly decreased on E19.5
(Fig. 3B).
Moreover, the serum lipid levels of E19.5 mice were
detected, which found no significant difference in the TG or LDL
levels between the DRAM1 group and the control group (Fig. 3H). The TC content decreased
significantly after DRAM1 overexpression, while the HDL content of
the DRAM1 group was significantly increased compared with in the
control group (Fig. 3H). Together,
these results demonstrated that overexpression of DRAM1 effectively
improved the symptoms of hyperlipidemia and high urine protein in
PE mice.
DRAM1 overexpression effectively
reduces the apoptosis of placental cells in PE mice
In addition, the level of oxidative stress was
measured in placental tissues on E19.5; this showed that the level
of SOD in the DRAM group was significantly higher compared with
that in the control group (Fig.
3E). DRAM1 overexpression significantly decreased the level of
MDA (Fig. 3F) and did not
significantly change the level of H2O2
(Fig. 3G).
Apoptosis was subsequently studied in placental
cells on E19.5. The DRAM1 group did not show a significant change
in the level of Bax, while Bcl-2 protein expression was
significantly increased compared with the control group (Fig. 3I and J). The Bax/Bcl-2 ratio was
significantly decreased after DRAM1 overexpression (Fig. 3J). These results showed that the
overexpression of DRAM1 effectively reduced oxidative
stress-induced apoptosis.
The overexpression of DRAM1 promotes
mitochondrial fusion, fission and mitophagy
To explore how DRAM1 attenuates the levels of
oxidative stress and apoptosis in the placentas of PE mice,
mitophagy and energy synthesis in the placenta were assessed after
DRAM1 overexpression. Protein expression of PINK1 and Parkin in the
DRAM1 group was significantly increased compared with in the
control group (Fig. 4A and B).
These results indicated that the overexpression of DRAM1
effectively promoted the occurrence of mitophagy. Overexpression of
DRAM1 also significantly promoted the expression of COX IV
(Fig. 4A and B). Together, these
results indicate that the overexpression of DRAM1 can promote
mitophagy, clear damaged mitochondria and improve mitochondrial
function.
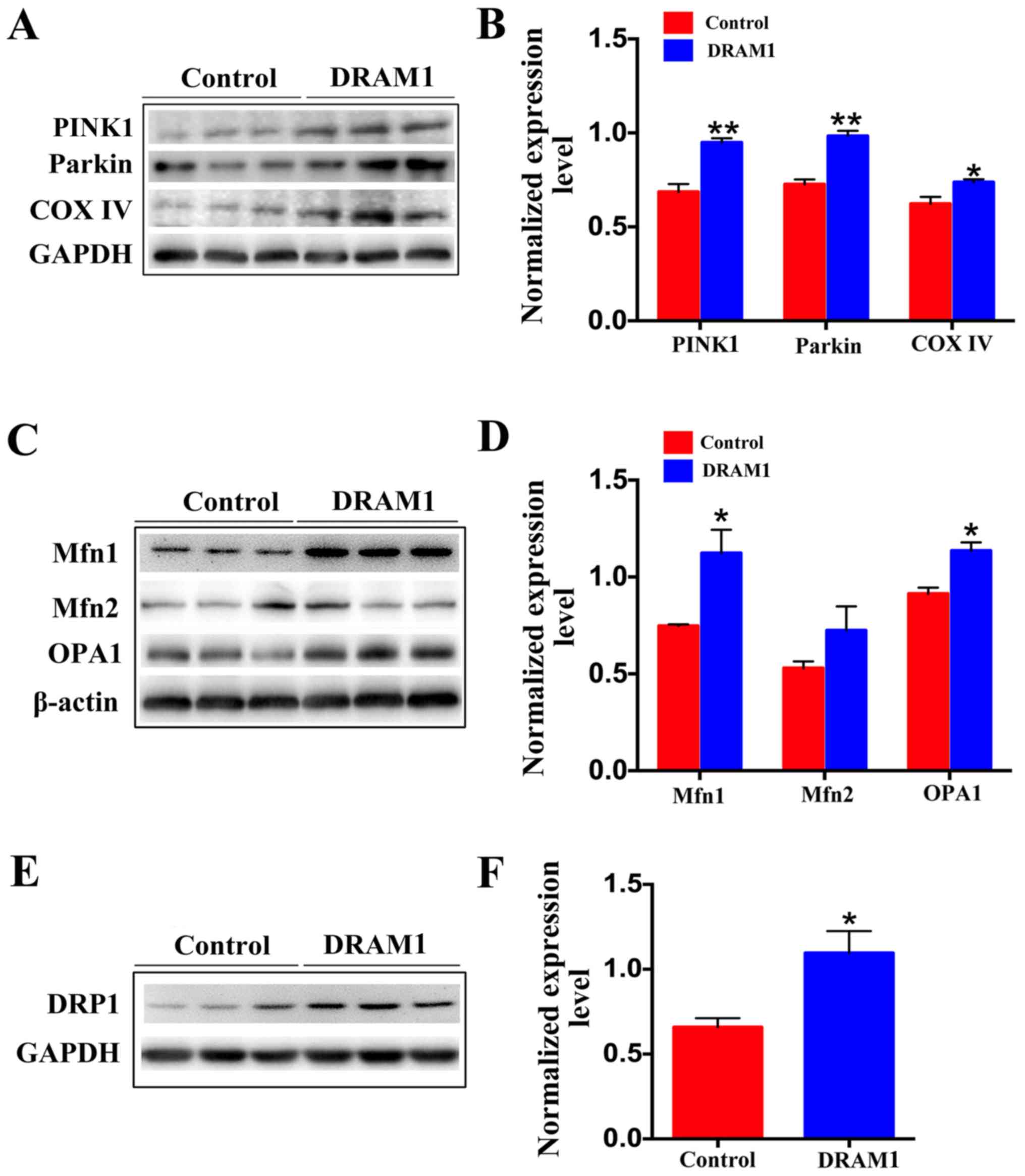 | Figure 4.Overexpression of DRAM1 induces
mitochondrial fusion and fission, and thus promotes mitophagy. (A)
Representative western blot images and (B) semi-quantification of
PINK1, Parkin and COX IV in the placenta of control and DRAM1 mice
suggested that DRAM1 overexpression promotes mitophagy. (C)
Representative western blot images and (D) semi-quantification of
Mfn1, Mfn2 and OPA1 in the placenta of control and DRAM1 mice
suggested that DRAM1 overexpression promotes mitochondrial fusion.
(E) Representative western blot images and (F) semi-quantification
of DRP1 in the placenta of control and DRAM1 mice suggested that
DRAM1 overexpression promotes mitochondrial fission. Densitometry
results were normalized against the levels of GAPDH or β-actin.
Values are expressed as the mean ± SEM (n=3). *P<0.05,
**P<0.01. DRAM1, DNA damage-regulated autophagy modulator 1;
PINK1, phosphatase and tensin homolog-induced putative kinase 1;
COX IV, cytochrome c oxidase IV; Mfn1, mitofusin 1; Mfn2,
mitofusin 2; OPA1, optic atrophy 1; DRP1, dynamin-related protein
1. |
The mitochondrial kinetic balance is critical for
the regulation of mitochondrial function, and the maintenance of
both energy metabolism and normal function through continuous
fission and fusion (40).
Therefore, changes in proteins associated with mitochondrial
fission and fusion were examined. Mitochondria have a double-layer
membrane structure, and the fusion of mitochondria is performed by
optic atrophy 1 (OPA1) and mitofusin (Mfn)1/2 (41). The fusion of the outer
mitochondrial membrane is mediated by Mfn1/2, and OPA1 mediates
that of the inner membrane. Protein expression of Mfn1/2 in the
placenta was measured using western blotting. This showed that Mfn1
expression was significantly increased (Fig. 4C and D), and that Mfn2 expression
was not significantly altered (Fig. 4C
and D), in the DRAM1 group compared with that of the control
group. OPA1 protein levels in the mouse placenta was significantly
increased in the DRAM1 group compared with the control group
(Fig. 4C and D). The expression
levels of DRP1 were also significantly increased in the DRAM1 group
compared with the control group (Fig.
4E and F). These results indicated that DRAM1 can promote both
the fusion and fission of mitochondria.
Discussion
Mitochondrial dysfunction and oxidative stress are
associated with PE, but thus far their levels have not been
measured in a mouse model of PE. However, such an investigation is
important for obtaining an improved understanding of the
contribution of mitochondrial dysfunction and oxidative stress to
the function of the placenta in PE. The present study revealed
several novel findings: i) The blood pressure of PE mice increased
gradually during gestation and recovered gradually after the end of
gestation; ii) PE mice exhibited high levels of oxidative stress in
the placenta, leading to apoptosis; iii) the placentas of PE mice
exhibited mitochondrial dysfunction, weaker mitophagy and low
levels of DRAM1; iv) DRAM1 overexpression effectively reduced both
the urine protein content and serum lipid levels in the placentas
of PE mice; v) DRAM1 overexpression significantly reduced the level
of oxidative stress, improved mitochondrial function and enhanced
the level of mitophagy in the placentas of PE mice; and vi) DRAM1
improved mitophagy by promoting mitochondrial fusion and fission,
thereby improving mitochondrial function. These results indicated
that increased DRAM1 in the placenta contributed to improved
mitochondrial dysfunction and decreased levels of oxidative stress
in PE.
Although ROS production is known to be induced by
the ischemic placenta in PE, ROS are also produced by systemic
blood vessels during the second phase of PE onset (12). Certain studies have revealed that
placental homogenates derived from patients with PE show 39% higher
H2O2 production than those derived from
healthy pregnant women (3,42). The present study also found a
significant increase in the level of H2O2 in
a mouse model of PE induced by Hif-1α. However, the content of
H2O2 in the placenta decreased after DRAM1
overexpression in PE mice. Furthermore, SOD is an enzyme that
catalyzes the scavenging of superoxide radicals, which are
constitutively expressed in the mitochondria and cytoplasm
(43). SOD is increased during
normal pregnancy, but the activity and mRNA expression of SOD in
placentas derived from patients with PE are decreased, which might
result in increased oxidative stress in the placentas of patients
with PE (44,45). It has been reported that SOD is
decreased in erythrocytes derived from patients with PE. The
present study also found a significant decrease in the activity of
SOD in the placentas of PE mice, and the activity of SOD was
increased significantly after overexpression of DRAM1.
Moreover, increased ROS concentrations in patients
with PE have been confirmed by the measurement of increased levels
of MDA, which is an index of lipid peroxidation (46). Consistently, the present study also
found that the expression of MDA in the placentas of PE mice was
significantly increased, and that the MDA content decreased
significantly after DRAM1 overexpression. In addition, lipid
peroxidation is closely related to heat-shock protein 70 (HSP70)
(44). Previously, the HSP70 level
in the peripheral blood was shown to be significantly higher in
both the fetal and maternal circulation in PE (47). The present study also found that
the serum TC and TG levels in the placentas of PE mice were
significantly increased, and that these levels gradually decreased
after DRAM1 overexpression.
Oxidative stress is the result of ROS content
exceeding the available antioxidant activity, and is one of the
risk factors for the development of PE through vascular dysfunction
(45). Poor invasion of
cytotrophoblasts into the uterine myometrium and disturbed spiral
artery remodeling play important roles in the pathophysiology of
PE, which results in placental hypoxia (48,49).
At present, termination of pregnancy is the only method for
avoiding maternal eclampsia and fetal distress in PE (50). Supplementation with numerous
antioxidants might prevent or repair the occurrence of PE (51). The results of the present study
indicated that DRAM1 overexpression can effectively reduce
oxidative stress levels in the placentas of PE mice, which could be
useful in developing a treatment for PE.
Over the past 20 years, >800 peer-reviewed
publications have validated the hypothesis that oxidative damage is
involved in the pathophysiology of PE; however, current antioxidant
interventions are not clinically effective (52). One possible explanation is that
these antioxidant regimens fail to reach the mitochondria in the
cell and thus fail to improve the pathology of oxidative damage.
Great advances have been made in mitochondrial pharmacology due to
the development of a number of different pharmacological strategies
for addressing mitochondrial dysfunction (53). Mitochondrial dysfunction is a
pathogenic mediator of oxidative stress in PE, as demonstrated by
the observation that the placentas of women with pregnancies
complicated by PE exhibit increased mitochondrial lipid
peroxidation and enhanced susceptibility of mitochondria to
oxidation (54,55). The present study also found that
COX IV levels were significantly decreased in the placentas of PE
mice.
There are a number of different indicators of
disrupted mitochondrial function, such as altered oxygen
consumption, decreased ATP production, increased ROS production and
mtDNA damage (56). Mitochondrial
damage is one of the most important factors that leads to
mitochondrial dysfunction (43).
Therefore, the prevention of mitochondrial damage is very
important, and several quality control mechanisms have evolved to
prevent mitochondrial damage and preserve a population of healthy
mitochondria. The selective autophagy of mitochondria, which is
known as mitophagy, is an important mitochondrial quality control
mechanism that eliminates damaged mitochondria (43). The elimination of damaged
mitochondria in mammals is mediated by a pathway involving PINK1
and Parkin (18). PINK1 and Parkin
accumulate on damaged mitochondria, promote their segregation from
the mitochondrial network and target these organelles for
autophagic degradation through a process that requires the
Parkin-dependent ubiquitination of mitochondrial proteins (57,58).
In the present study, significant decreases in the levels of PINK1
and Parkin in the placental tissues of PE mice were observed,
suggesting that the mitochondrial dysfunction in PE is likely due
to a decreased level of mitophagy. DRAM1 overexpression
significantly increased the levels of PINK1 and Parkin in the
placentas of PE mice, which indicated that mitophagy was
effectively activated. COX IV protein expression was also
increased, which indicated an effective improvement in
mitochondrial function.
How DRAM1 could improve mitophagy and thus promote
recovery of mitochondrial function is important to explore.
Mitochondrial dynamics are regulated via mitochondrial biogenesis,
and continuous cycles of fission and fusion (40). These processes are hypothesized to
target damaged/depolarized mitophagy (38). Mitochondrial fission and fusion are
regulated by members of a family of conserved large GTPases that
were initially identified in yeast (41). The dynamin-like GTPase
dynamin-related protein 1 (DRP1) mediates fission by forming a
multimeric complex that wraps around the outer membrane of
mitochondrial tubules and exerts a mechanical force to produce
membrane scission (17). In
contrast to fission, during which mitochondria are divided using
only an outer membrane apparatus, two distinct machineries, Mfn1/2
and OPA1, are required for fusion of the outer and inner membranes
(41). In the present study, OPA1
and Mfn1/2 protein expression was measured, and it was found that
mitochondrial fusion was significantly increased after DRAM1
overexpression. These findings are consistent with the findings
reported by Zhang et al (59), who found that Mfn2 is downregulated
in PE placentas. In addition, Zhang et al (59) demonstrated that ATP is
significantly reduced after knockout of the Mfn2 gene in
trophoblast cells. Therefore, the findings of the present study
suggested that DRAM1 can improve mitochondrial dysfunction by
promoting mitochondrial fusion in PE. Alternatively, during
fission, unhealthy, nonfunctional mitochondrial fragments that lack
transmembrane potentials are discarded and targeted for degradation
via mitophagy (60). Wakabayashi
et al (61) demonstrated
that DRP1−/− mice exhibit embryonic lethality due to a
deficiency in the formation of trophoblast giant cells and
consequent placental dysfunction, which underscores the requirement
of mitochondrial fission for proper placental and embryonic
development. Hence, DRAM1 promotes mitochondrial fission to
accelerate mitophagy for the clearance of damaged mitochondria,
which is significant for the maintenance of mitochondrial
function.
As previously reported, the expression of DRAM1
could induce apoptosis in an autophagy dependent or independent
manner (21,62). However, the mechanism via which
DRAM1 induces apoptosis or autophagic apoptosis remains unclear. In
addition, it has been reported that inflammation-associated
apoptosis is regulated by Bcl-2 and Bax (63,64),
and that decreased invasion and migration ability of trophoblast
cells induced by cell apoptosis in the placenta may be the major
causes of PE (65,66). Therefore, the specific mechanism of
DRAM1 and the relationship of between inflammation and Bcl-2 and
Bax requires further investigation.
In conclusion, to the authors' best knowledge, the
present study provided the first demonstration that DRAM1 reduces
oxidative stress-induced apoptosis by improving mitochondrial
dysfunction, enhancing mitophagy clearance through mitochondrial
fission and fusion, and rescuing urinary protein and serum lipid
levels in the placentas of PE mice. These data shed light on a
novel mechanism underlying the therapeutic effect of DRAM1 in
PE.
Acknowledgements
The authors are grateful to Professor Yao-ting Gui
and Professor Guang-hui Cui of Shenzhen Hospital of Peking
University for their assistance with the technical and animal
experiments.
Funding
The present study was supported by the Medical
Science and Technology Research Fund Project of Guangdong (grant
no. A2019363), the National Science Foundation of China (grant nos.
81260385 and 81360383), the Clinical Research Project of Shenzhen
Municipal Health Commission (grant no. SZLY2017017) and the
Doctoral Project of Shenzhen Maternal and Child Health Hospital
(grant no. FYA2018005).
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
GQC contributed to the acquisition and
interpretation of the data, performed the experiments and
statistical analyses, and participated in the drafting of the
manuscript. YL, LC and FZ analyzed and interpreted the data, and
LZ, YH, PPH and LL performed the statistical analyses. YL and LL
participated in the drafting of the manuscript. YLY designed the
study, performed the experiments, analyzed and interpreted the
data, drafted the manuscript, procured funding and supervised the
study. All authors read and approved the final manuscript.
Ethics approval and consent to
participate
All animal experiments and procedures were approved
by the Ethics Committee of Shenzhen Hospital of Peking University
(permit no. SYXK-2015-0106, 2019-078).
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Reijnders D, Olson KN, Liu CC, Beckers KF,
Ghosh S, Redman LM and Sones JL: Dyslipidemia and the role of
adipose tissue in early pregnancy in the BPH/5 mouse model for
preeclampsia. Am J Physiol Regul Integr Comp Physiol. 317:R49–R58.
2019. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Kay VR, Ratsep MT, Figueiro-Filho EA and
Croy BA: Preeclampsia may influence offspring neuroanatomy and
cognitive function: A role for placental growth factor? Biol
Reprod. 101:271–283. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Maged AM, Shoab AY and Dieb AS: Antepartum
and postpartum uterine artery impedance in women with
pre-eclampsia: A case control study. J Obstet Gynaecol. 39:633–638.
2019. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Benkő Z, Chaveeva P, de Paco Matallana C,
Zingler E, Wright A, Wright D and Nicolaides KH: Validation of
competing-risks model in screening for pre-eclampsia in twin
pregnancy by maternal factors. Ultrasound Obstet Gynecol.
53:649–654. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Shin EK, Kang HY, Yang H, Jung EM and
Jeung EB: The regulation of fatty acid oxidation in human
preeclampsia. Reprod Sci. 23:1422–1433. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Sun MN, Yang Z and Ma RQ: Interaction of
fatty acid oxidation with oxidative stress in preeclampsia-like
mouse model at multiple stages of gestation. Zhonghua Yi Xue Za
Zhi. 91:2343–2347. 2011.(In Chinese). PubMed/NCBI
|
|
7
|
Huai J, Yang Z, Yi YH and Wang GJ: Role of
mammalian target of rapamycin signaling pathway in regulation of
fatty acid oxidation in a preeclampsia-like mouse model treated
with pravastatin. Chin Med J (Engl). 132:671–679. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Zárate A, Saucedo R, Valencia J, Manuel L
and Hernández M: Early disturbed placental ischemia and hypoxia
creates immune alteration and vascular disorder causing
preeclampsia. Arch Med Res. 45:519–524. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Schoots MH, Gordijn SJ, Scherjon SA, van
Goor H and Hillebrands JL: Oxidative stress in placental pathology.
Placenta. 69:153–161. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Aouache R, Biquard L, Vaiman D and
Miralles F: Oxidative Stress in Preeclampsia and Placental
Diseases. Int J Mol Sci. 19:14962018. View Article : Google Scholar
|
|
11
|
Lecarpentier E and Tsatsaris V: Angiogenic
balance (sFlt-1/PlGF) and preeclampsia. Ann Endocrinol (Paris).
77:97–100. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Taysi S, Tascan AS, Ugur MG and Demir M:
Radicals, Oxidative/Nitrosative stress and preeclampsia. Mini Rev
Med Chem. 19:178–193. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Saito S and Nakashima A: A review of the
mechanism for poor placentation in early-onset preeclampsia: The
role of autophagy in trophoblast invasion and vascular remodeling.
J Reprod Immunol 101–102. 80–88. 2014. View Article : Google Scholar
|
|
14
|
Klingenberg M: The ADP-ATP translocation
in mitochondria, a membrane potential controlled transport. J Membr
Biol. 56:97–105. 1980. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Kalyanaraman B, Cheng G, Hardy M, Ouari O,
Bennett B and Zielonka J: Teaching the basics of reactive oxygen
species and their relevance to cancer biology: Mitochondrial
reactive oxygen species detection, redox signaling, and targeted
therapies. Redox Biol. 15:347–362. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Samartsev VN, Vedernikov AA, Khoroshavina
EI and Dubinin MV: Comparative study of free oxidation and ATP
synthesis in mitochondria in the liver of different animal species.
J Evol Biochem Phys. 53:245–247. 2017. View Article : Google Scholar
|
|
17
|
Kubli DA, Zhang X, Lee Y, Hanna RA,
Quinsay MN, Nguyen CK, Jimenez R, Petrosyan S, Murphy AN and
Gustafsson AB: Parkin protein deficiency exacerbates cardiac injury
and reduces survival following myocardial infarction. J Biol Chem.
288:915–926. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Tusi SK, Khodagholi F and Sanati MH:
Mitophagy pathway is induced by alginate oligosaccharide in PC12
cells exposed to oxidative stress. Alzheimers Dement. 9:3072013.
View Article : Google Scholar
|
|
19
|
Oyewole AO and Birch-Machin MA:
Mitochondria-targeted antioxidants. FASEB J. 29:4766–4771. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Zhang XD, Qi L, Wu JC and Qin ZH: DRAM1
regulates autophagy flux through lysosomes. PLoS One. 8:e632452013.
View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Liu D, Li R, Guo X, Pang L, Zang Y, Liu K
and Chen D: DNA damage regulated autophagy modulator 1 recovers the
function of apoptosis-stimulating of p53 protein 2 on inducing
apoptotic cell death in Huh7.5 cells. Oncol Lett. 15:9333–9338.
2018.PubMed/NCBI
|
|
22
|
Nagata M, Arakawa S, Yamaguchi H, Torii S,
Endo H, Tsujioka M, Honda S, Nishida Y, Konishi A and Shimizu S:
Dram1 regulates DNA damage-induced alternative autophagy. Cell
Stress. 2:55–65. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Cho A and Seung-Hyeok S: Ethical
Guidelines for Use of Experimental Animals in Biomedical Research.
J Bacteriol Virol. 43:18–26. 2013. View Article : Google Scholar
|
|
24
|
Wilde E, Aubdool AA, Thakore P, Baldissera
L Jr, Alawi KM, Keeble J, Nandi M and Brain SD: Tail-Cuff technique
and its influence on central blood pressure in the mouse. J Am
Heart Assoc. 6:e0052042017. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Saito T: In vivo electroporation in the
embryonic mouse central nervous system. Nat Protoc. 1:1552–1558.
2006. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Albers RE, Kaufman MR, Natale BV, Keoni C,
Kulkarni-Datar K, Min S, Williams CR, Natale DRC and Brown TL:
Trophoblast-Specific expression of Hif-1α results in
Preeclampsia-Like symptoms and fetal growth restriction. Sci Rep.
9:27422019. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Iriyama T, Wang W, Parchim NF, Song A,
Blackwell SC, Sibai BM, Kellems RE and Xia Y: Hypoxia-independent
upregulation of placental hypoxia inducible factor-1α gene
expression contributes to the pathogenesis of preeclampsia.
Hypertension. 65:1307–1315. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Zadora J, Singh M, Herse F, Przybyl L,
Haase N, Golic M, Yung HW, Huppertz B, Cartwright JE, Whitley G, et
al: Disturbed placental imprinting in preeclampsia leads to altered
expression of DLX5, a Human-Specific early trophoblast marker.
Circulation. 136:1824–1839. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Samimi M, Pourhanifeh MH, Mehdizadehkashi
A, Eftekhar T and Asemi Z: The role of inflammation, oxidative
stress, angiogenesis, and apoptosis in the pathophysiology of
endometriosis: Basic science and new insights based on gene
expression. J Cell Physiol. 234:19384–19392. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Wu MY, Yiang GT, Lai TT and Li CJ: The
oxidative stress and mitochondrial dysfunction during the
pathogenesis of diabetic retinopathy. Oxid Med Cell Longev.
2018:34201872018. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Hassan M, Watari H, AbuAlmaaty A, Ohba Y
and Sakuragi N: Apoptosis and molecular targeting therapy in
cancer. Biomed Res Int. 2014:1508452014. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Demendi C, Börzsönyi B, Végh V, Nagy ZB,
Rigó J Jr, Pajor A and Joó JG: Gene expression patterns of the
Bcl-2 and Bax genes in preterm birth. Acta Obstet Gynecol Scand.
91:1212–1217. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Irani RA, Zhang Y, Blackwell SC, Zhou CC,
Ramin SM, Kellems RE and Xia Y: The detrimental role of angiotensin
receptor agonistic autoantibodies in intrauterine growth
restriction seen in preeclampsia. J Exp Med. 206:2809–2822. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Han Y, Yang Z, Ding X, Yu H and Yi Y:
Expression of long chain fatty acid oxidase in maternal and fetal
tissues in preeclampsia-like mouse model in mid-gestation. Zhonghua
Yi Xue Za Zhi. 95:26–29. 2015.(In Chinese). PubMed/NCBI
|
|
35
|
Whitley BN, Engelhart EA and Hoppins S:
Mitochondrial dynamics and their potential as a therapeutic target.
Mitochondrion. 49:269–283. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Koch RE, Josefson CC and Hill GE:
Mitochondrial function, ornamentation, and immunocompetence. Biol
Rev Camb Philos Soc. 92:1459–1474. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Adam-Vizi V: Production of reactive oxygen
species in brain mitochondria: Contribution by electron transport
chain and non-electron transport chain sources. Antioxid Redox
Signal. 7:1140–1149. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Yoo SM and Jung YK: A molecular approach
to mitophagy and mitochondrial dynamics. Mol Cells. 41:18–26.
2018.PubMed/NCBI
|
|
39
|
Villa E, Marchetti S and Ricci JE: No
Parkin Zone: Mitophagy without Parkin. Trends Cell Biol.
28:882–895. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Guzmán-Lastra F, Kaiser A and Löwen H:
Fission and fusion scenarios for magnetic microswimmer clusters.
Nat Commun. 7:135192016. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Kanki T, Klionsky DJ and Okamoto K:
Mitochondria autophagy in yeast. Antioxid Redox Signal.
14:1989–2001. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Bartha JL, Visiedo F, Fernández-Deudero A,
Bugatto F and Perdomo G: Decreased mitochondrial fatty acid
oxidation in placentas from women with preeclampsia. Placenta.
33:132–134. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Beretta S, Sala G, Mattavelli L, Ceresa C,
Casciati A, Ferri A, Carrì MT and Ferrarese C: Mitochondrial
dysfunction due to mutant copper/zinc superoxide dismutase
associated with amyotrophic lateral sclerosis is reversed by
N-acetylcysteine. Neurobiol Dis. 13:213–221. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Ilhan N, Ilhan N and Simsek M: The changes
of trace elements, malondialdehyde levels and superoxide dismutase
activities in pregnancy with or without preeclampsia. Clin Biochem.
35:393–397. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Xuan RR, Niu TT and Chen HM: Astaxanthin
blocks preeclampsia progression by suppressing oxidative stress and
inflammation. Mol Med Rep. 14:2697–2704. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Feng YL, Yin YX, Ding J, Yuan H, Yang L,
Xu JJ and Hu LQ: Alpha-1-antitrypsin suppresses oxidative stress in
preeclampsia by inhibiting the p38MAPK signaling pathway: An in
vivo and in vitro study. PLoS One. 12:e01737112017. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Fekete A, Vér A, Bögi K, Treszl A and Rigó
J Jr: Is preeclampsia associated with higher frequency of HSP70
gene polymorphisms? Eur J Obstet Gynecol Reprod Biol. 126:197–200.
2006. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Yan JY and Xu X: Relationships between
concentrations of free fatty acid in serum and oxidative-damage
levels in placental mitochondria and preeclampsia. Zhonghua Fu Chan
Ke Za Zhi. 47:412–417. 2012.(In Chinese). PubMed/NCBI
|
|
49
|
Hilali N, Kocyigit A, Demir M, Camuzcuoglu
A, Incebiyik A, Camuzcuoglu H, Vural M and Taskin A: DNA damage and
oxidative stress in patients with mild preeclampsia and offspring.
Eur J Obstet Gynecol Reprod Biol. 170:377–380. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Salomon C, Yee SW, Mitchell MD and Rice
GE: The possible role of extravillous trophoblast-derived exosomes
on the uterine spiral arterial remodeling under both normal and
pathological conditions. Biomed Res Int. 2014:6931572014.
View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Salles AM, Galvao TF, Silva MT, Motta LCD
and Pereira MG: Antioxidants for preventing preeclampsia: A
systematic review. ScientificWorldJournal. 2012:2434762012.
View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Thomas T, Jophy R, Mhaskar A and Misquith
D: Are we increasing serious maternal morbidity by postponing
termination of pregnancy in severe pre-eclampsia/eclampsia? J
Obstet Gynaecol. 25:347–351. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
53
|
Vishnyakova PA, Volodina MA, Tarasova NV,
Marey MV, Tsvirkun DV, Vavina OV, Khodzhaeva ZS, Kan NE, Menon R,
Vysokikh MY and Sukhikh GT: Mitochondrial role in adaptive response
to stress conditions in preeclampsia. Sci Rep. 6:324102016.
View Article : Google Scholar : PubMed/NCBI
|
|
54
|
Zsengellér ZK, Rajakumar A, Hunter JT,
Salahuddin S, Rana S, Stillman IE and Ananth Karumanchi S:
Trophoblast mitochondrial function is impaired in preeclampsia and
correlates negatively with the expression of soluble fms-like
tyrosine kinase 1. Pregnancy Hypertens. 6:313–319. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
55
|
Abad C, Vargas FR, Zoltan T, Proverbio T,
Piñero S, Proverbio F and Marín R: Magnesium sulfate affords
protection against oxidative damage during severe preeclampsia.
Placenta. 36:179–185. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
56
|
Kumar A, Davuluri G, Welch N, Kim A,
Gangadhariah M, Allawy A, Priyadarshini A, McMullen MR, Sandlers Y,
Willard B, et al: Oxidative stress mediates ethanol-induced
skeletal muscle mitochondrial dysfunction and dysregulated protein
synthesis and autophagy. Free Radic Biol Med. 145:284–299. 2019.
View Article : Google Scholar : PubMed/NCBI
|
|
57
|
Nakashima A, Tsuda S, Kusabiraki T, Aoki
A, Ushijima A, Shima T, Cheng SB, Sharma S and Saito S: Current
understanding of autophagy in pregnancy. Int J Mol Sci.
20:23422019. View Article : Google Scholar
|
|
58
|
Ausman J, Abbade J, Ermini L, Farrell A,
Tagliaferro A, Post M and Caniggia I: Ceramide-induced BOK promotes
mitochondrial fission in preeclampsia. Cell Death Dis. 9:2982018.
View Article : Google Scholar : PubMed/NCBI
|
|
59
|
Zhang J, Qu B, Yu W, Zhu Y, Yan X, Shen H
and Zhao J: Role of surface ectoderm-specific mitofusin 2 in the
corneal morphologic development of mice. Am J Transl Res.
11:3620–3628. 2019.PubMed/NCBI
|
|
60
|
Chapple SJ, Cheng X and Mann GE: Effects
of 4-hydroxynonenal on vascular endothelial and smooth muscle cell
redox signaling and function in health and disease. Redox Biol.
1:319–331. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
61
|
Wakabayashi J, Zhang Z, Wakabayashi N,
Tamura Y, Fukaya M, Kensler TW, Iijima M and Sesaki H: The
dynamin-related GTPase Drp1 is required for embryonic and brain
development in mice. J Cell Biol. 186:805–816. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
62
|
Lorin S, Borges A, Ribeiro Dos Santos L,
Souquère S, Pierron G, Ryan KM, Codogno P and Djavaheri-Mergny M:
c-Jun NH2-terminal kinase activation is essential for
DRAM-dependent induction of autophagy and apoptosis in
2-methoxyestradiol-treated ewing sarcoma cells. Cancer Res.
69:6924–6931. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
63
|
Nguyen HB, Loomba M, Yang JJ, Jacobsen G,
Shah K, Otero RM, Suarez A, Parekh H, Jaehne A and Rivers EP: Early
lactate clearance is associated with biomarkers of inflammation,
coagulation, apoptosis, organ dysfunction and mortality in severe
sepsis and septic shock. J Inflamm (Lond). 7:62010. View Article : Google Scholar : PubMed/NCBI
|
|
64
|
Marriott HM, Hellewell PG, Cross SS, Ince
PG, Whyte MK and Dockrell DH: Decreased alveolar macrophage
apoptosis is associated with increased pulmonary inflammation in a
murine model of pneumococcal pneumonia. J Immunol. 177:6480–6488.
2006. View Article : Google Scholar : PubMed/NCBI
|
|
65
|
Jiang Y and Chen Y and Chen Y: Knockdown
of JARID2 inhibits the viability and migration of placenta
trophoblast cells in preeclampsia. Mol Med Rep. 16:3594–3599,
20174. View Article : Google Scholar : PubMed/NCBI
|
|
66
|
Arroyo J, Price M, Straszewski-Chavez S,
Torry RJ, Mor G and Torry DS: XIAP protein is induced by placenta
growth factor (PLGF) and decreased during preeclampsia in
trophoblast cells. Syst Biol Reprod Med. 60:263–273. 2014.
View Article : Google Scholar : PubMed/NCBI
|














