Introduction
Bisphosphonate (BP) is an effective treatment for
osteoporosis, multiple myeloma, Paget's disease and bone-metastatic
cancer (1). BP decreases the rate
of complications of bone metastasis and minimizes
skeletal-associated events in malignancies. However, unwanted side
effects, such as osteonecrosis of the jaw and atypical femur
fractures, still occur (2). Lo
et al (3) used thorough
screening to show an increased number of cases of BP-related
osteonecrosis of the jaw (BRONJ; prevalence in America in 2006,
0.10%; 95% CI, 0.05–0.20) due to chronic oral BP usage. The highest
reported incidence rate of BRONJ is 5–10%, occurring in patients
using high doses of BP, which leads to decreased drug prescription
and increased occurrence of osteoporosis-associated complications
(4,5).
The majority of BRONJ cases occur following dental
extraction; incidence rates depend on the severity of dental
disease or adjacent active periodontal disease. Marx (6) reported that 85/152 BRONJ cases were
associated with dental extraction, and suggested prevention
strategies, such as interrupting BP treatment before such
procedures. As periodontal disease is present in the majority of
patients with osteonecrosis, oral infections and surgical dental
treatments are considered to be a triggering event for BRONJ
(7). Previous case reports and
animal studies have identified periodontal disease as a key risk
factor for the development of BRONJ (8,9).
Clinically, a number of reports have noted that BRONJ occurs
following tooth extraction due to the severity of dental disease or
the presence of active periodontal or periapical disease in
surrounding tissues (10,11). In 2014, Oteri et al
(8) conducted a case-control study
and revealed that patients with BRONJ have fewer teeth, greater
clinical attachment level and less alveolar bone support than
control subjects. Our previous study demonstrated that patients
with BRONJ had deep periodontal pockets and severe periodontal bone
defects adjacent to the exposed necrotic bone (12). However, at present, the exact
molecular signaling pathway of BRONJ pathogenesis, as well as its
association with infections or other stress, remains poorly
understood.
In order to explore the underlying mechanism of
BRONJ, the effect of the most widely used BP (zoledronic acid) on
periodontal ligament stem cells (PDLSCs) was analyzed along with
intracellular molecular changes. Human PDLSCs isolated from human
periodontal ligament (PDL) tissue exhibited a capacity for
self-renewal and multipotency, thereby serving a key role in
regeneration of periodontal tissue. The aim of the present study
was to analyze the impact of BP treatment on the transcriptome of
PDLSCs and to identify key processes and genes involved in BRONJ
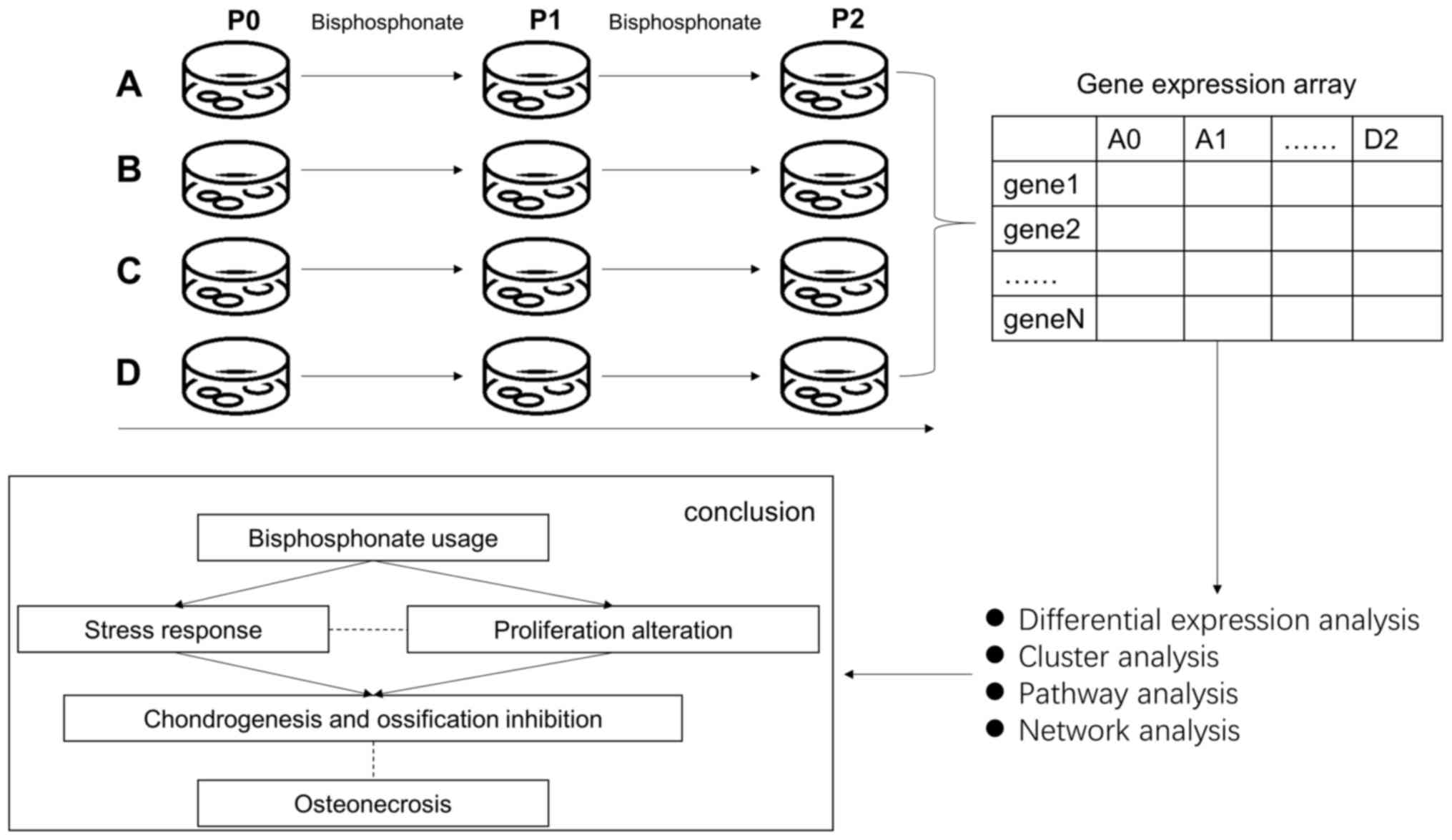
that may serve as potential therapeutic targets. A schematic
outlining the current experiments is presented in Fig. 1.
Materials and methods
Cell culture
Human PDLSCs were isolated from healthy premolars or
third molars extracted from eight young adults (five males and
three females, aged 15–23 years) under orthodontic treatment with
no history of periodontal disease. Sample collection was conducted
at Shanghai Ninth People's Hospital between January, 2018 and
April, 2018. The experimental protocols were approved by the Ethics
Committee of the Ninth People's Hospital Affiliated with Shanghai
Jiao Tong University, School of Medicine (Shanghai, China). All
donors provided written informed consent prior to participating in
the present study. PDLSCs were isolated and cultured as previously
reported, with a slight modification (12). Following extraction, teeth were
immediately placed into a solution of PBS containing 100 U/ml
penicillin and 100 U/ml streptomycin. After washing thoroughly in
PBS (3–5 times) to remove blood components, the PDL tissues were
gently scraped from the surface of the middle of the root, minced
into 1–3 cubes (2 mm3), and placed into 6-well culture
dishes. Subsequently, glass cover slips were placed over the
tissues to prevent floating, and PDL tissues were incubated in
culture medium (DMEM, HyClone; Cytiva) with 10% FBS (Gibco; Thermo
Fisher Scientific, Inc.), 100 U/ml penicillin and 100 µg/ml
streptomycin) in a humidified atmosphere of 95% air and 5%
CO2 at 37°C. The tissues were subsequently maintained by
replacing the medium every 3–4 days until cell density reached 90%
confluence. Subsequently, the cells were detached using 0.25%
trypsin/EDTA and sub-cultured at a density of 1×105
cells/cm2 in 100-mm dishes. Cells at passage 3–5 were
used for subsequent experiments.
Self-renewal and multiple
differentiation of PDLSCs in vitro
In order to assess the self-renewal capacity of
PDLSCs, cells (200/well) were plated in a 6-well plate. The 14-day
cultures were fixed with 4% formalin for 15 min at 37°C and stained
with 0.1% crystal violet (Beyotime Institute of Biotechnology) for
15 min at 37°C. Aggregates of ≥50 cells were scored as colonies. In
order to investigate the osteogenic ability, PDLSCs at passage 3
were seeded at a density of 5×104 cells/well in 12-well
plates and incubated in growth medium (DMEM, HyClone; Cytiva) with
10% FBS (Gibco; Thermo Fisher Scientific, Inc.), 100 U/ml
penicillin and 100 µg/ml streptomycin) containing 10 nM
dexamethasone (Sigma-Aldrich; Merck KGaA), 5 mM β-glycerophosphate
(Sigma-Aldrich; Merck KGaA) and 500 µM ascorbic acid
(Sigma-Aldrich; Merck KGaA) for 14 days to induce mineralization.
The cells were then stained with 1% Alizarin Red (Sigma-Aldrich;
Merck KGaA) for 1 h at 37°C. In order to investigate adipogenic
ability, PDLSCs at passage 3 were seeded at a density of
5×104 cells/well in 12-well plates and incubated in
basal medium (Gibco; Thermo Fisher Scientific, Inc.) (DMEM with 10%
FBS, 100 U/ml penicillin and 100 µg/ml streptomycin) at 37°C
supplemented with 0.5 mM methylisobutylxantine, 0.5 mM
hydrocortisone, 200 µM indomethacin and 10 µg/ml insulin for 4
weeks and stained with Oil Red O (5 mg/ml; Sigma-Aldrich; Merck
KGaA) for 1 h at 37°C. All images of the cells were captured using
an inverted contrast-phase light microscope (Nikon
Corporation).
Zoledronate treatments
Zoledronate was purchased from Novartis
Pharmaceuticals UK Ltd. and diluted in 0.9% NaCl infusion solution.
PDLSCs were treated with an oncologic dose of zoledronate (50 µM)
at days 3 and 5 at 37°C. Control cells were incubated with the same
procedure, except that they were treated with vehicle only (0.9%
NaCl). The oncologic dose of zoledronate was selected based on
published pharmacokinetic data (13). Calculations of the concentrations
of zoledronate exposure equivalents on cells were performed as
previously described (14).
RNA extraction, purification and
quality control
Total RNA from each cell sample was extracted using
TRIzol® (cat. no. 15596-018; Thermo Fisher Scientific,
Inc.) according to the manufacturer's protocol. RNA integrity was
evaluated using an Agilent Bioanalyzer 2100 (Agilent Technologies,
Inc.). Qualified total RNA was further purified using an RNeasy
Micro kit (cat. no. 74004; Qiagen GmbH) and RNase-Free DNase Set
(cat. no. 79254; Qiagen GmbH). Extracted RNA was analyzed using a
NanoDrop ND-2000 spectrophotometer (NanoDrop Technologies; Thermo
Fisher Scientific, Inc.) and Agilent Bioanalyzer 2100 (Agilent
Technologies, Inc.). Quality thresholds were set as RNA integrity
number >6 and S28/S18 >0.7.
Array hybridization
Prior to hybridization, total RNA was first
amplified, labeled and purified using a GeneChip 3′IVT PLUS Reagent
kit (cat. no. 902416; Affymetrix; Thermo Fisher Scientific, Inc.)
according to the manufacturer's protocol to obtain biotin-labeled
cRNA. Array hybridization and washing were performed using a
GeneChip® Hybridization, Wash and Stain kit (cat. no.
900720; Affymetrix; Thermo Fisher Scientific, Inc.) in a
Hybridization Oven 645 (cat. no. 00-0331-220V; Affymetrix; Thermo
Fisher Scientific, Inc.) and Fluidics Station 450 (cat. no.
00-0079; Affymetrix; Thermo Fisher Scientific, Inc.) according to
the manufacturer's protocols.
Data acquisition and
pre-processing
Slides were scanned using a GeneChip®
Scanner 3000 (Affymetrix; Thermo Fisher Scientific, Inc.) and
Command Console software (version 4.0; Affymetrix; Thermo Fisher
Scientific, Inc.) using the default settings. Raw data for each
sample were filtered using the following thresholds: i) Average
background value <100; and ii) for at least one housekeeping
gene, 3′/5′signals <3. All samples passed these thresholds. Raw
data were quantile-normalized and log2-transformed using
the robust multi-array average algorithm and Gene Spring software
(Agilent Technologies, Inc., version 12.6.1). A total of 49,282
detected probes were mapped to the GRCh37/hg19 human genome. For
each gene, the probe with the maximum average value was selected to
represent its expression level. Only probes that were detected in
all samples were included in subsequent analysis. Expression level
data for 18,384 genes were obtained for downstream analysis.
Differential expression analysis
Differential expression analysis was performed using
limma version 3.38.3 (15) on R
3.4.1 (16). The limma pipeline
for time course experiments was used to compare P2 (5 days after
treatment) vs. P0 (day 0), P2 vs. P1 (3 days after treatment) and
P1 vs. P0. A linear model was fitted with the linear formula
exp-series + timepoint where exp denoted gene expression, series
denoted cell line batch and timepoint corresponded to P0, P1 and
P2.
The empirical Bayes method (15) was used to estimate the fold change
between time points. For P2 vs. P0 and P1 vs. P0, the reference
level for timepoint was set as P0, and results were extracted for
each comparison with the parameter coef. For P2 vs. P1,
another linear model was fitted using only eight samples
(discarding four samples of P0) of P2 and P1 and the reference
level for timepoint was set as P1. The threshold of differentially
expressed genes (DEGs) was set at: i) Benjamini-Hochberg-adjusted
P-value <0.05; and ii) |log2 fold change (FC)|>1.
Control cells were analyzed in the same way. DEGs that exhibited
the same direction in both treatment and control cells were
excluded from downstream analysis.
Cluster analysis
Hierarchical cluster analysis based on Euclidean
distance was performed on all 12 samples of three timepoints using
two sets of features: i) All DEGs from P2 vs. P0; and ii) union of
DEGs from P2 vs. P1 and P1 vs. P0. The expression level value of
each gene was scaled and centered to 0 before analysis. Cluster
analysis and construction of the heatmap were achieved using the
heatmap function in R (17).
Gene Ontology (GO) analysis
GO-biological process (GO-BP) pathway analysis
(18) was performed using the
clusterProfiler R package version 3.16.0 (19). A hypergeometric test was used to
investigate whether the DEG lists were enriched in any GO-BP
pathways. Gene background was defined as all genes with GO
annotation. Only pathways with ≥10 genes were included in the
analysis. P-values of hypergeometric tests were adjusted for
multiple testing via the Benjamini-Hochberg method. For all
pathways with adjusted P-value ≤0.05, non-redundant pathways were
selected as results (i.e. no pathway was the parent term of any
other pathway).
Protein-protein interaction (PPI)
network analysis
Human PPI data was obtained from the Biogrid
database (20) (access date, June
1, 2018). A PPI network of all DEGs was constructed using Cytoscape
version 3.6.1 (21). Node degree
was calculated using the network analyzer in Cytoscape. In order to
investigate co-expression patterns in the PPI network, Pearson
correlation coefficients (PC) were calculated for all identified
PPI pairs using the ‘cor’ function in R.
Results
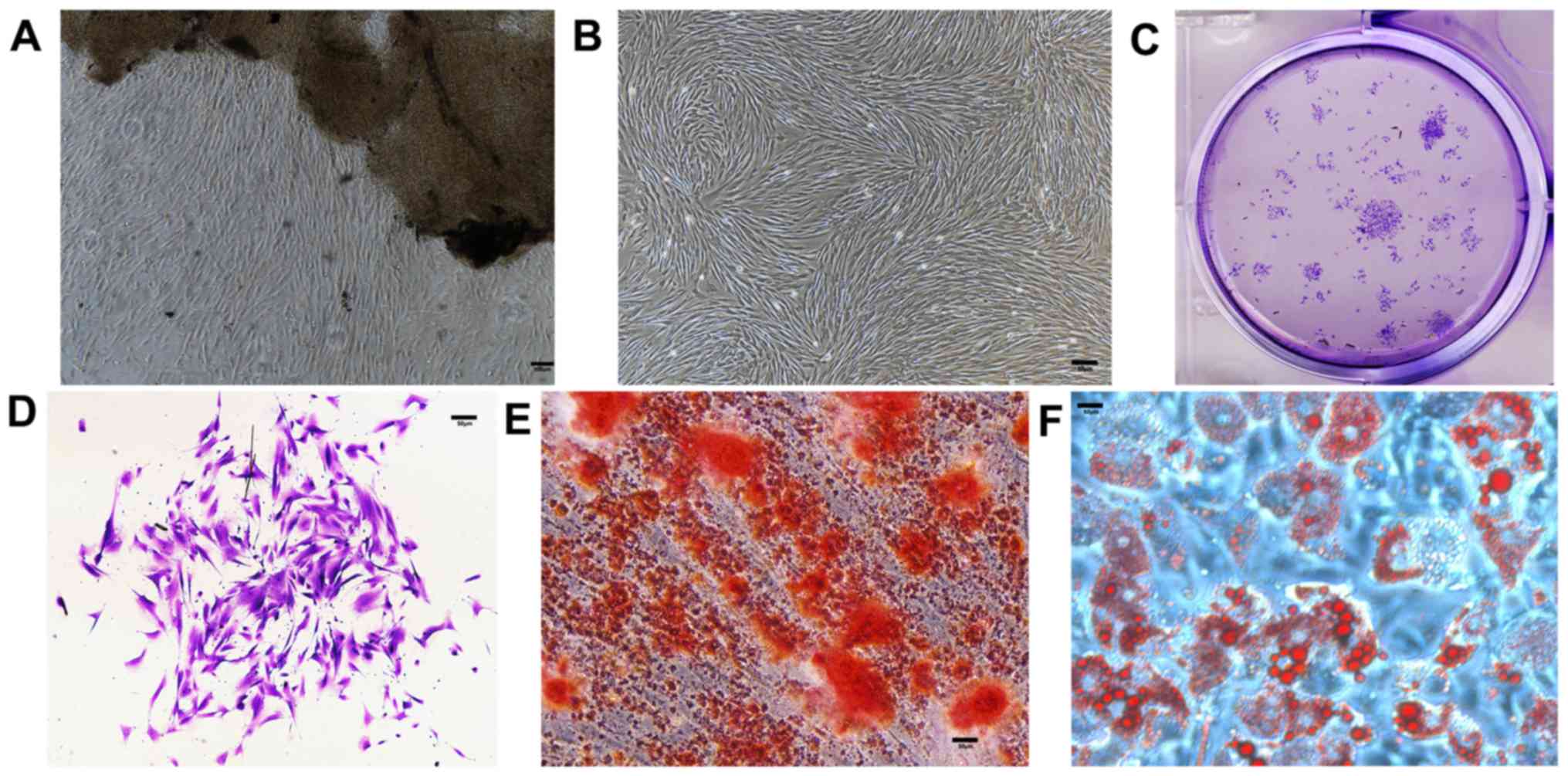
In vitro self-renewal and multipotent
capacities of PDLSCs
The first step of the present study was to verify
whether the in vitro model of PDLSCs could correctly imitate
the in vivo physiology of PDL tissues. Cells were found
growing around the PDL tissues 5–10 days after initial incubation
(Fig. 2A) and proliferated to
reach 90% confluence in 7–10 days (Fig. 2B). Similar to other mesenchymal
stem cells, PDLSCs formed adherent clonogenic cell clusters of
fibroblast-like cells (Fig. 2C and
D). The multipotent capacity of PDLSCs was verified by
induction in the osteogenic and adipogenic media in vitro.
Alizarin Red staining (Fig. 2E)
revealed a number of calcified nodules in the cultures after 14
days of induction. After 4 weeks of culture in adipogenic medium,
intracellular lipid vacuoles appeared in PDLSCs, the presence of
which was confirmed by Oil Red O staining (Fig. 2F).
Quantification of the PDLSC
transcriptome via gene expression level array
In order to evaluate the impact of BP on PDLSCs,
genome-wide transcription levels of all samples were measured at
three time points (days 0, 3 and 5) using the Affymetrix PrimeView
Human Gene Expression Array. A total of 49,282 probes were detected
and mapped to 20,608 genes. Following normalization and filtration,
the expression levels of 18,384 genes for all samples were
obtained. These expression level data enabled further analysis of
the biological effects of BP usage.
Genes exhibit different patterns of
expression in response to BP
Having quantified the gene expression levels of
PDLSCs at different time points (P0, 1 and 2), gene expression
levels which were significantly altered following BP exposure were
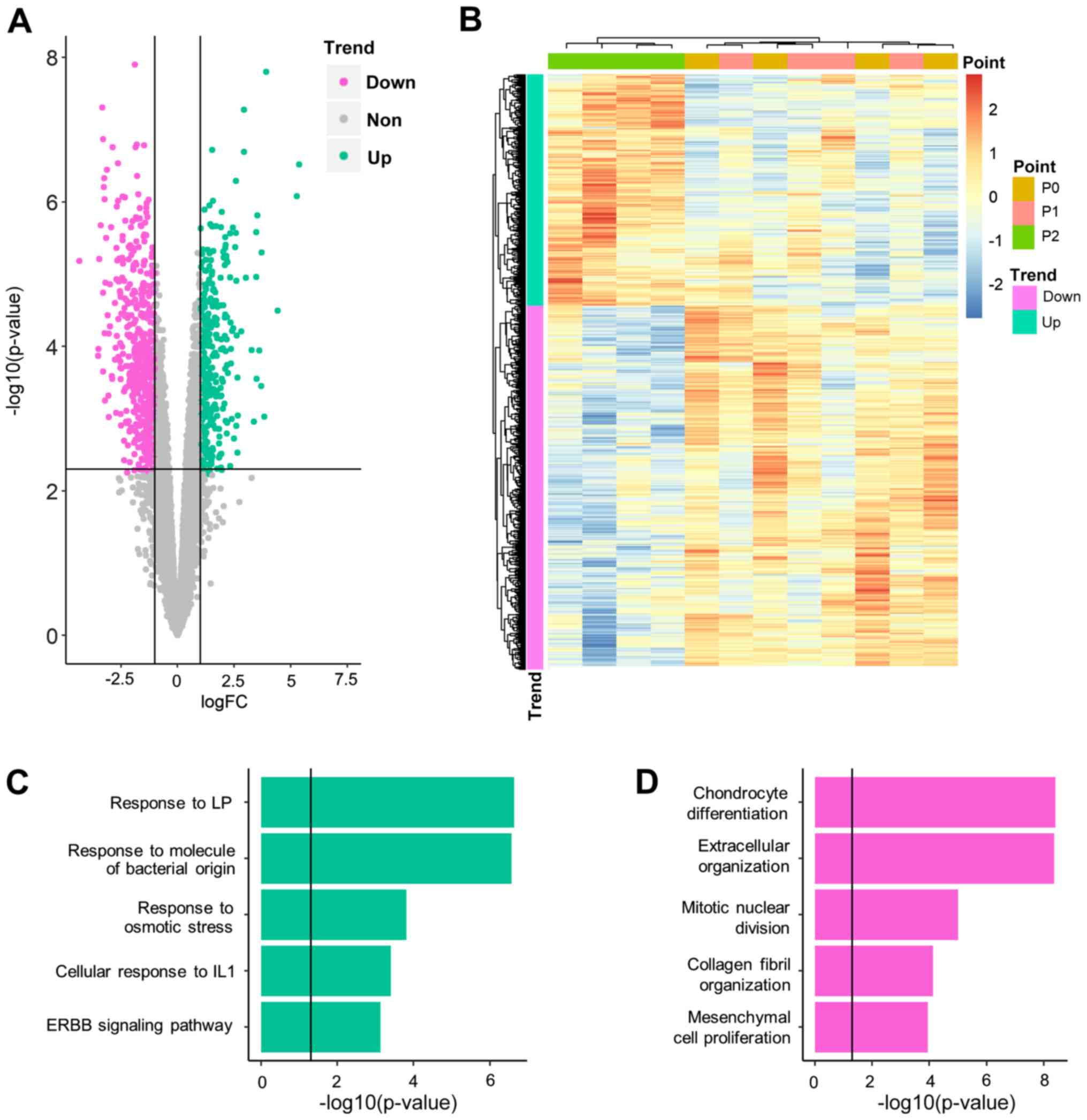
identified. First, gene expression levels were compared at P2 and
P0, which represented the longest exposure to BP. A total of 906
genes (352 upregulated and 554 downregulated) were demonstrated to
be significantly differentially expressed between P2 and P0
(Fig. 3A; Table SI). Among these genes, two were
also differentially expressed in control cell lines (Table SII) and were, therefore, excluded
from downstream analysis. The remaining DEGs could distinguish P2
from P1 and P0 samples but could not distinguish between P1 and P0
samples (Fig. 3B-D). It was
hypothesized that these genes required a long period of BP exposure
to induce significant alteration. The DEGs between P2 and P0 were
referred to as slow-response DEGs. One of these DEGs, COL1A1
(logFC, −2.89; P=0.007), was highlighted in a previous genetic
association study, which reported that COL1A1 polymorphism
increased the risk of BRONJ (22).
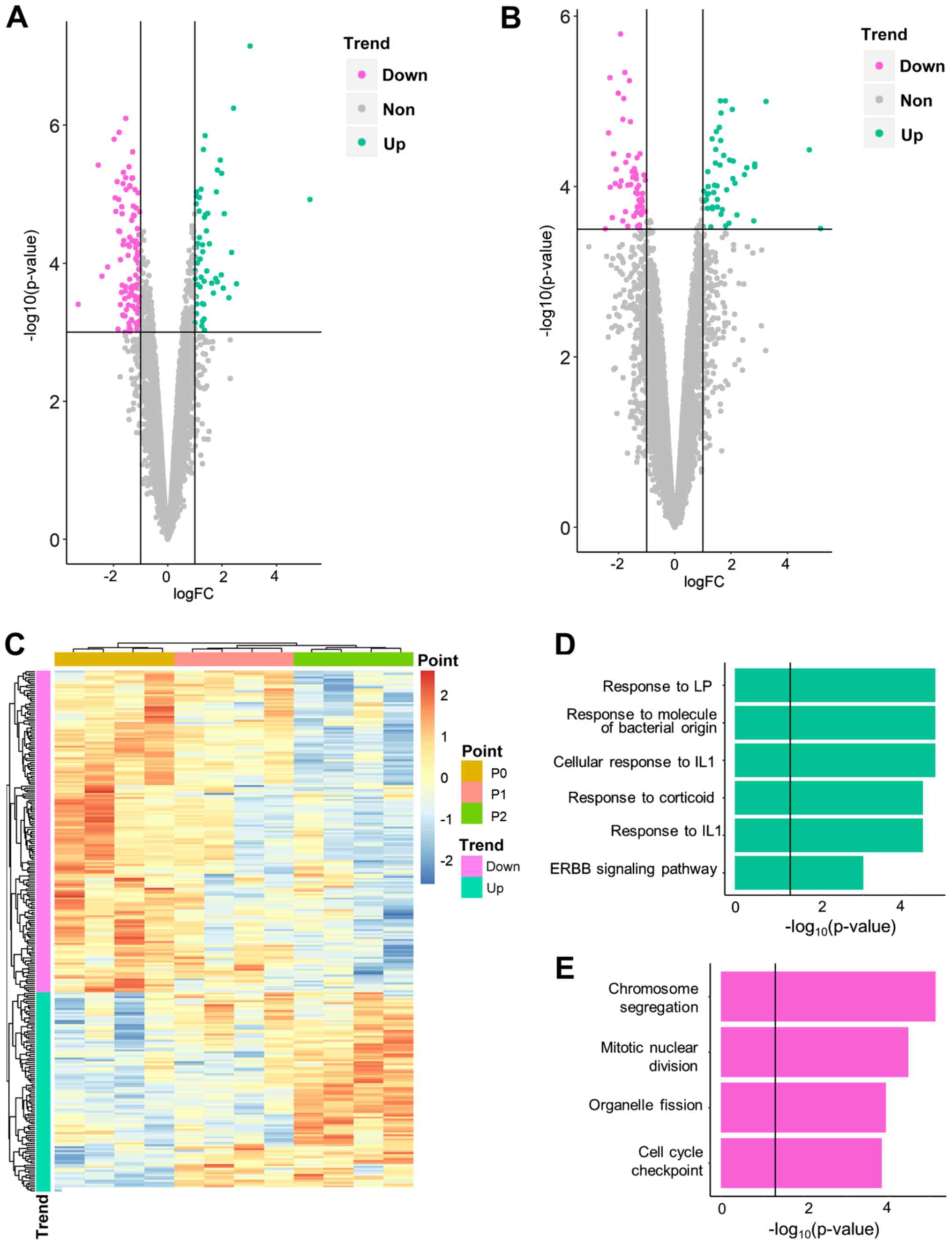
Similarly, expression level data was compared
between P2 and P1, and P1 and P0. It was hypothesized that such
comparisons would identify genes that were altered rapidly
following BP exposure; hence, these DEGs between adjacent time
points were referred to as fast-response DEGs. A total of 264
fast-response DEGs, including 168 DEGs for P1 vs. P0 (Fig. 4A; Table SIII) and 105 DEGs for P2 vs. P1 (9
overlapping genes; Fig. 4B;
Table SIV), were identified. The
majority (229/264) were also slow-response DEGs (i.e.
differentially expressed between P0/1 and 2). Among all
fast-response DEGs, 163 genes were downregulated and 101 were
upregulated over time (Fig. 4C).
No DEGs were identified in adjacent comparisons of control cells
(Table SII), which indicates that
the fast-response DEGs were not driven by experimental procedures.
Fast-response DEGs successfully separated three time points in
hierarchical clustering analysis (Fig.
4C).
BP exposure influences stress response
and cartilage generation
Having identified genes influenced by BP treatment,
the biological consequences of BP usage were investigated. GO-BP
analysis was applied separately to upregulated and downregulated
DEG lists.
For slow-response DEGs, a notable cellular stress
reaction was observed (Fig. 3C;
Table SV). Upregulated
slow-response DEGs were significantly enriched in response to
inflammatory factors, such as lipopolysaccharide
(P-adjusted=2.27×10−7), bacterial molecules
(P-adjusted=2.69×10−7) and interleukin-1
(P-adjusted=3.93×10−4). Another inflammation-associated
signaling pathway, ‘ERBB signaling pathway’
(P-adjusted=7.40×10−4), was also moderately enriched.
Downregulated genes (Fig. 3D;
Table SVI) inhibited cartilage
generation and were notably enriched in ‘chondrocyte
differentiation’ (P-adjusted=3.92×10−9) and
‘extracellular organization’ (P-adjusted=4.38×10−9).
‘Ossification’ (P-adjusted=1.36×10−5) was also
significantly enriched by downregulated genes, which suggested that
it was inhibited during BP treatment. Cell proliferation was also
inhibited, demonstrated by enrichment of ‘mitotic nuclear division’
(P-adjusted=9.98×10−6) and ‘mesenchymal cell
proliferation’ (P-adjusted=1.12×10−4).
For fast-response DEGs, different enrichment
patterns were observed. All enriched pathways were associated with
proliferation and cell cycle regulation (e.g. ‘chromosome
segregation’; P-adjusted=7.62×10−6; Table VII). For
upregulated fast-response DEGs, the induction of cellular stress
reaction was similar to that of slow-response DEGs. These results
indicated that stress reaction was induced more rapidly than
inhibition of cartilage generation and ossification.
CYLD lysine 63 deubiquitinase (CYLD)
and poly(ADP-ribose) polymerase 1 (PARP1) function as hub genes in
BP pathology
Having identified the key biological processes in BP
pathology, it was investigated whether any key genes (hubs) served
a central role in these processes. Such hubs may serve as potential
interventional targets to prevent osteonecrosis.
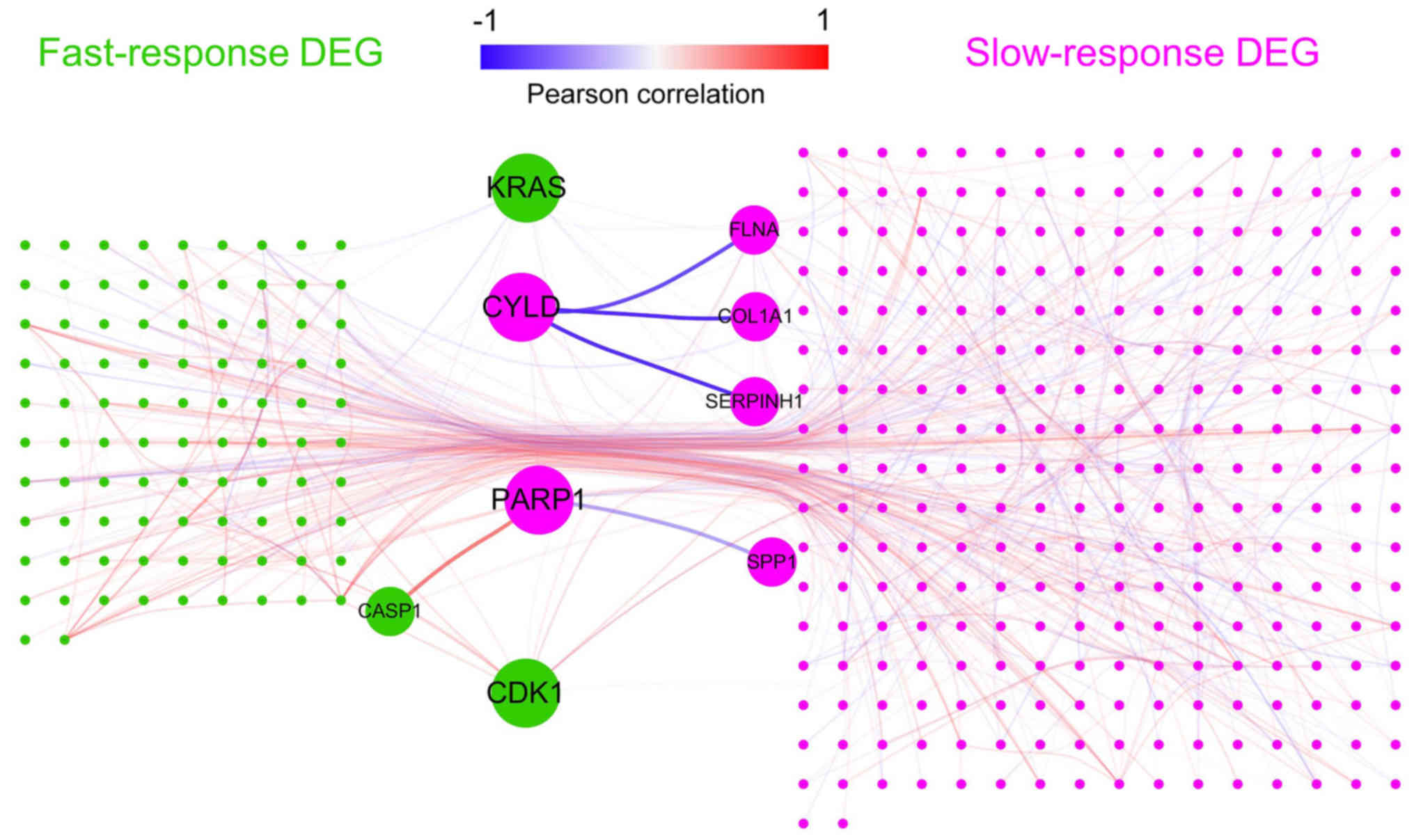
In order to investigate the potential hub genes, a
PPI network of all DEGs was constructed (Fig. 5). It was hypothesized that hub
genes might act as ‘bridges’ between fast- and slow-response DEGs
and connect more slow-response DEGs. If such hub genes exist, they
may be potential drug targets. Thus, the number of slow-response
DEGs connected by each gene was calculated (Table SVIII). The top gene, PARP1,
which connected 14 slow-response DEGs, was an upregulated
slow-response DEG. PARP1 was connected and positively
correlated with caspase-1 (CASP1), an upregulated
fast-response DEG that serves a vital role in response to
inflammation and stress (23).
PARP1 was connected and negatively correlated
with secreted phosphoprotein 1 (SPP1), a gene involved in
the attachment of osteoclasts to the mineralized bone matrix
(24). The CYLD gene, which
connected 13 slow-response DEGs, had three PPI partners that are
involved in collagen biosynthesis [serpin family H member 1
(SERPINH1) (24), filamin A
(FLNA) (25) and
COL1A1]. A negative correlation was observed between
CYLD and these collagen-associated genes, which indicated
that CYLD is upregulated by fast-response genes and may
inhibit downstream extracellular matrix generation of bone.
PARP1 and CYLD were slow-response
DEGs. It was further determined whether any fast-response DEGs
could serve as hub genes. KRAS and CDK1, which
connect 11 slow-response DEGs, were the top genes among
fast-response DEGs. They both connected a number of genes involved
in ‘cell proliferation’ and ‘cell cycle’ (Table SVI). KRAS was negatively
corelated with PPI partners WW and C2 domain containing 1 (PC,
−0.70503), kinetochore scaffold 1 (PC, −0.56551) and cell division
cycle associated 8 (PC, −0.7047), whereas CDK1 exhibited a
notable positive correlation with a number of cell cycle genes,
such as cyclin B1 (PC, 0.922838) and non-SMC condensin I complex
subunit G (PC, 0.975156; Table
SIX).
Discussion
Osteonecrosis of the jaw is one of the most severe
complications in patients using BP (26). The present study reports the
results of a genome-wide landscape of transcriptomic alteration in
PDLSCs during BP usage, which may increase the understanding of the
mechanism of BRONJ and putative drug targets for BRONJ.
With the advantage of time series designation, the
present analysis demonstrated that genes respond to BP exposure at
different rates. The majority of BP-associated genes (676/941)
exhibited significant differential expression levels only at day 5
vs. 0, indicating that they responded to BP in a slow and gradual
manner. The distinct expression level patterns of fast- and
slow-response genes built a ‘two-wave’ alteration, which is in
accordance with the clinical observation that osteonecrosis of the
jaw follows a chronic course following BP exposure (27).
The two-wave expression level alteration affected
different biological functions. As indicated by the GO-BP analysis,
fast-response DEGs were notably enriched in inflammatory and stress
responses, as well as in cell proliferation signaling pathways.
However, slow-response DEGs exhibited enrichment in ossification
and cartilage-associated signaling pathways. As reported by Aghaloo
et al (27) and
Lorenzo-Pouso et al (28),
inflammation and inhibition of bone remodeling have been suggested
as putative mechanisms of BRONJ. While isolated evidence of
resorption-generation imbalance (29), periodontal disease (7) and infection (30) has been discovered in pathological
research of BRONJ, these hypotheses are treated as being exclusive
and require validation (31). The
present results indicated that these signaling pathways may be
different phases of a single uniform pathological procedure and
therefore should not be considered separately.
Another debate regarding BRONJ pathology is the
mutual association between inflammatory damage and infection.
Infection can directly damage the bone structure and exposed bones
are further susceptible to bacterial colonization (32). To date, the causative factors from
this mutual association have not been fully elucidated. Based on
the present sterile in vitro experiments, it was observed
that inflammation and stress responses can arise in the absence of
infection. Despite being a sterile environment, PDLSCs exhibited
signs of fighting against infection (e.g., enrichment in ‘response
to molecules of bacterial origin’). It can be inferred that BP can
induce inflammatory reactions in PDLSCs in the absence of other
causative factors. Therefore, it was hypothesized that inflammation
and stress response are causative factors in the pathology of
BRONJ, and prevention of BP-induced inflammation is a potential
target for the management of BRONJ.
As the stress response occurs prior to inhibition of
ossification, it was hypothesized that the former process gives
rise to the latter via key intermediate molecules, namely hubs. PPI
network analysis identified two potential hub genes: PARP1
and CYLD. PARP1 encodes a chromatin-associated
enzyme, poly(ADP-ribosyl)transferase (33), and acts as a bridge between
fast-response inflammation and slow-response ossification
inhibition by interacting with CASP1 and SPP1,
respectively (23,24). This bridge-like structure suggests
that PARP1-based intervention may have a decoupling effect
on the inflammation-ossification inhibition cascade. Another
potential hub, CYLD, encodes a cytoplasmic protein with
three cytoskeletal-associated protein-glycine-conserved domains
that functions as a deubiquitinating enzyme (34). Previous studies have linked
CYLD with extracellular matrix generation (35) and bone formation/resorption balance
(36). In PDLSCs, CYLD was
observed to interact with slow-response DEGs (SERPINH1, FLNA
and COL1A1) that are involved in collagen biogenesis, a key
process in extracellular generation (37) and bone formation (38). Although no evidence was found to
support the association between CYLD and inflammation in
PDLSCs, the strong connection between CYLD and collagen
function make it a putative hub gene in the network of BP
pathology. As for the fast-response gene, the upregulation of KRAS
and downregulation of CDK1 supported the hypothesis that cell
proliferation was inhibited by BP usage. KRAS and CDK1 may be key
drivers of such inhibition.
The present study has certain limitations. The
duration of BP treatment was relatively short, and all PDLSC
samples were collected from patients aged <30 years. The present
results cannot be extrapolated to older patients or those receiving
long-term BP treatment. Furthermore, the present study only
analyzed BRONJ in terms of transcriptomes, so does not represent
the overall picture of BRONJ-associated pathology. Further
functional validation experiments are required to verify the key
processes and hub genes identified by the present analysis.
BP induced stress response-like transcriptional
alterations in PDLSCs, followed by inhibition of proliferation and
ossification. Such alterations may contribute to the onset of
osteonecrosis of the jaw. PARP1 and CYLD may be two
key genes in this pathological procedure. The present findings
support an anti-inflammatory strategy in management of BP-induced
osteonecrosis and indicate that PARP1 and CYLD may be
potential targets in drug therapy of BRONJ.
Supplementary Material
Supporting Data
Supporting Data
Supporting Data
Supporting Data
Supporting Data
Supporting Data
Supporting Data
Supporting Data
Supporting Data
Acknowledgements
Not applicable.
Funding
This study was supported by the National Natural
Science Foundation of China (grant nos. 81271114 and 31700848) and
Science and Technology Commission of Shanghai Municipality (grant
nos. 19411962000 and 18441903000). The funding organizations had no
role in the design or conduct of this research.
Availability of data and materials
The datasets generated and/or analyzed during the
current study are available in the Github repository,
github.com/WeiCSong/PDLSC.
Authors' contributions
SW designed and supervised the study. ML and YY
collected the samples. HH and ML conducted cellular experiments. YS
collected and analyzed the data and wrote the manuscript. YZ and WZ
interpreted the data and revised the manuscript. All authors read
and approved the manuscript.
Ethics approval and consent to
participate
The experimental protocols were approved by the
ethics committee of Ninth Peoples Hospital affiliated to Shanghai
Jiao Tong University, School of Medicine (approval no.
SH9H-2020-T37-2). Written informed consent was obtained from the
participants prior to participation in the study. Informed consent
was obtained from the parents of the one patient who was a
minor.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Woo SB, Hellstein JW and Kalmar JR:
Systematic review: Bisphosphonates and osteonecrosis of the jaws.
Ann Intern Med. 144:7532006. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Holen I and Coleman RE: Bisphosphonates as
treatment of bone metastases. Curr Pharm Des. 16:1262–1271. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Lo JC, O'Ryan FS, Gordon NP, Yang J, Hui
RL, Martin D, Hutchinson M, Lathon PV, Sanchez G, Silver P, et al:
Prevalence of osteonecrosis of the jaw in patients with oral
bisphosphonate exposure. J Oral Maxillofac Surg. 68:243–253. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Elsayed R, Abraham P, Awad ME, Kurago Z,
Baladhandayutham B, Whitford GM, Pashley DH, McKenna CE and
Elsalanty ME: Removal of matrix-bound zoledronate prevents
post-extraction osteonecrosis of the jaw by rescuing osteoclast
function. Bone. 110:141–149. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Schwartz HC: American association of oral
and maxillofacial surgeons position paper on medication-related
osteonecrosis of the jaw-2014 update and CTX. J Oral Maxillofac
Surg. 73:3772015. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Marx RE: Oral and intravenous
bisphosphonate-induced osteonecrosis of the jaws: History etiology,
prevention, and treatment. Royal College of Surgeons of England.
2009.
|
|
7
|
Aghaloo TL, Kang B, Sung EC, Shoff M,
Ronconi M, Gotcher JE, Bezouglaia O, Dry SM and Tetradis S:
Periodontal disease and bisphosphonates induce osteonecrosis of the
jaws in the rat. J Bone Miner Res. 26:1871–1882. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Oteri G, Bramanti E, Nigrone V and Cicciù
M: Periodontal health in osteoporotic patients affected by BRONJ:
An observational study. J Osteoporos. 2013:2312892013. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Li CL, Lu WW, Seneviratne CJ, Leung WK,
Zwahlen RA and Zheng LW: Role of periodontal disease in
bisphosphonate-related osteonecrosis of the jaws in ovariectomized
rats. Clin Oral Implants Res. 27:1–6. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Thumbigere-Math V, Michalowicz BS, Hodges
JS, Tsai ML, Swenson KK, Rockwell L and Gopalakrishnan R:
Periodontal disease as a risk factor for bisphosphonate-related
osteonecrosis of the jaw. J Periodontol. 85:226–233. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Krimmel M, Ripperger J, Hairass M, Hoefert
S, Kluba S and Reinert S: Does dental and oral health influence the
development and course of bisphosphonate-related osteonecrosis of
the jaws (BRONJ)? Oral Maxillofac Surg. 18:213–218. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Li M, Yu Y, Shi Y, Yuqiong Zhou Y, Zhang
W, Hua H, Ge J, Zhang Z, Ye D, Yang C and Wang S: Decreased
osteogenic ability of periodontal ligament stem cells leading to
impaired periodontal tissue repair in BRONJ patients. Stem Cells
Dev. 29:156–168. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Russell RGG, Watts NB, Ebetino FH and
Rogers MJ: Mechanisms of action of bisphosphonates: Similarities
and differences and their potential influence on clinical efficacy.
Osteoporos Int. 19:733–759. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Otto S, Pautke C, Opelz C, Westphal I,
Drosse I, Schwager J, Bauss F, Ehrenfeld M and Schieker M:
Osteonecrosis of the jaw: Effect of bisphosphonate type, local
concentration, and acidic milieu on the pathomechanism. J Oral
Maxillofac Surg. 68:2837–2845. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Ritchie ME, Phipson B, Wu D, Hu Y, Law CW,
Shi W and Smyth GK: limma powers differential expression analyses
for RNA-sequencing and microarray studies. Nucleic Acids Res.
43:e472015. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
R Core Team: R: A language and environment
for statistical computing. R Foundation for Statistical Computing.
(Vienna, Austria). 2019.
|
|
17
|
Raivo Kolde: Pheatmap: Pretty Heatmaps. R
package version 1.0.8. 2018.
|
|
18
|
Ashburner M, Ball CA, Blake JA, Botstein
D, Butler H, Cherry JM, Davis AP, Dolinski K, Dwight SS, Eppig JT,
et al: Gene ontology: Tool for the unification of biology. Nat
Genet. 25:25–29. 2000. View
Article : Google Scholar : PubMed/NCBI
|
|
19
|
Yu G, Wang LG, Han Y and He QY:
clusterProfiler: An R package for comparing biological themes among
gene clusters. OMICS. 16:284–287. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Chatr-aryamontri A, Oughtred R, Boucher L,
Rust J, Chang C, Kolas NK, O'Donnell L, Oster S, Theesfeld C,
Sellam A, et al: The BioGRID interaction database: 2017 update.
Nucleic Acids Res. 45:D369–D379. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Shannon P, Markiel A, Ozier O, Baliga NS,
Wang JT, Ramage D, Amin N, Schwikowski B and Ideker T: Cytoscape: A
software environment for integrated models of biomolecular
interaction networks. Genome Res. 13:2498–2504. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Katz J, Gong Y, Salmasinia D, Hou W,
Burkley B, Ferreira P, Casanova O, Langaee TY and Moreb JS: Genetic
polymorphisms and other risk factors associated with bisphosphonate
induced osteonecrosis of the jaw. Int J Oral Maxillofac Surg.
40:605–611. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Franchi L, Eigenbrod T, Muñoz-Planillo R
and Nuñez G: The inflammasome: A caspase-1-activation platform that
regulates immune responses and disease pathogenesis. Nat Immunol.
10:241–247. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Chellaiah MA, Kizer N, Biswas R, Alvarez
U, Strauss-Schoenberger J, Rifas L, Rittling SR, Denhardt DT and
Hruska KA: Osteopontin deficiency produces osteoclast dysfunction
due to reduced CD44 surface expression. Mol Biol Cell. 14:173–189.
2003. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Falet H, Pollitt AY, Begonja AJ, Weber SE,
Duerschmied D, Wagner DD, Watson SP and Hartwig JH: A novel
interaction between FlnA and Syk regulates platelet ITAM-mediated
receptor signaling and function. J Exp Med. 207:1967–1979. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Khosla S, Burr D, Cauley J, Dempster DW,
Ebeling PR, Felsenberg D, Gagel RF, Gilsanz V, Guise T, Koka S, et
al: Bisphosphonate-associated osteonecrosis of the jaw: Report of a
task force of the American Society for bone and mineral research. J
Bone Miner Res. 22:1479–1491. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Aghaloo T, Hazboun R and Tetradis S:
Pathophysiology of osteonecrosis of the jaws. Oral Maxillofac Surg
Clin North Am. 27:489–496. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Lorenzo-Pouso AI, Pérez-Sayáns M,
González-Palanca S, Chamorro-Petronacci C, Bagán J and
García-García A: Biomarkers to predict the onset of
biphosphonate-related osteonecrosis of the jaw: A systematic
review. Med Oral Patol Oral Cir Bucal. 24:e26–e36. 2019.PubMed/NCBI
|
|
29
|
Coleman RE, Major P, Lipton A, Brown JE,
Lee KA, Smith M, Saad F, Zheng M, Hei YJ, Seaman J and Cook R:
Predictive value of bone resorption and formation markers in cancer
patients with bone metastases receiving the bisphosphonate
zoledronic acid. J Clin Oncol. 23:4925–4935. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Ruggiero SL, Dodson TB, Fantasia J,
Goodday R, Aghaloo T, Mehrotra B and O'Ryan F; American Association
of Oral and Maxillofacial Surgeons, : American association of oral
and maxillofacial surgeons position paper on medication-related
osteonecrosis of the jaw-2014 update. J Oral Maxillofac Surg.
72:1938–1956. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Allen MR and Burr DB: The pathogenesis of
bisphosphonate-related osteonecrosis of the jaw: So many
hypotheses, so few data. J Oral Maxillofac Surg. 67 (Suppl
5):S61–S70. 2009. View Article : Google Scholar
|
|
32
|
Reid IR and Cornish J: Epidemiology and
pathogenesis of osteonecrosis of the jaw. Nat Rev Rheumatol.
8:90–96. 2012. View Article : Google Scholar
|
|
33
|
Zhu X, Ma X and Hu Y: PARP1: A promising
target for the development of PARP1-based candidates for anticancer
intervention. Curr Med Chem. 23:1756–1774. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Mathis BJ, Lai Y, Qu C, Janicki JS and Cui
T: CYLD-mediated signaling and diseases. Curr Drug Targets.
16:284–294. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Wang H, Lai Y, Mathis BJ, Wang W, Li S, Qu
C, Li B, Shao L, Song H, Janicki JS, et al: Deubiquitinating enzyme
CYLD mediates pressure overload-induced cardiac maladaptive
remodeling and dysfunction via downregulating Nrf2. J Mol Cell
Cardiol. 84:143–153. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Vriend J and Reiter RJ: Melatonin, bone
regulation and the ubiquitin-proteasome connection: A review. Life
Sci. 145:152–160. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Rodriguez-Pascual F and Slatter DA:
Collagen cross-linking: Insights on the evolution of metazoan
extracellular matrix. Sci Rep. 6:373742016. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Saito M and Marumo K: Effects of collagen
crosslinking on bone material properties in health and disease.
Calcif Tissue Int. 97:242–261. 2015. View Article : Google Scholar : PubMed/NCBI
|