Introduction
Hepatic ischemia/reperfusion injury (HIRI) often
occurs as a consequence of liver surgery and numerous types of
injury, including transplantation, trauma, shock and resection
(1). HIRI has been discovered to
be a major cause of liver dysfunction and organ rejection (2). Therefore, the identification of
pharmacological treatments that effectively improve HIRI outcomes
remains a priority over invasive methods.
HIRI usually involves initial ischemia-induced cell
damage, followed by a reperfusion-induced inflammatory response,
which directly injures the liver tissue (3). As the primary producers of ATP and
reactive oxygen species (ROS), hepatocyte mitochondria are critical
for normal liver function (4).
Therefore, clinical interventions targeting the functional
homeostasis of liver mitochondria represent a promising approach
for HIRI treatment.
Previous studies using experimental models have
demonstrated that aldehyde dehydrogenase 2 (ALDH2) served a pivotal
role in the pathophysiology of cardiovascular injury (5,6).
ALDH2 is the primary enzyme that catalyzes ethanol metabolism by
detoxifying acetaldehyde into non-toxic acetate; acetate is then
generated in the mitochondrial matrix (7). In particular, ALDH2 is known have an
important role in the elimination of endogenous oxidized aldehyde
adducts (8). These adducts, which
are formed by lipid peroxidation under oxidative stress, include
4-hydroxy-2-nonenal (4-HNE) and malondialdehyde (MDA) (8,9). The
excessive accumulation of the 4-HNE adduct was discovered to lead
to mitochondrial dysfunction during acute myocardial IRI and
chronic heart failure, which impaired the cardiac contractility,
mitochondrial bioenergetics and redox balance (5,10).
Interestingly, these detrimental outcomes were found to be reversed
through the upregulation of ALDH2 (11). Conversely, 4-HNE and ROS have been
identified to inhibit ALDH2 activity (11).
Guo et al (12) previously demonstrated that ALDH2
was activated by Alda-1, improving chronic alcohol-induced
steatosis and apoptosis. Alda-1, a member of the ALDH2
isozyme-specific activator family, was discovered to increase the
catalytic activity of ALDH2 both in vivo and in vitro
(13). However, whether ALDH2
activation can exert a protective effect during HIRI remains
unclear.
Autophagy is an intracellular degradative process
that targets damaged organelles, dysfunctional proteins and harmful
products through the formation of autophagosomes and autolysosomes
(14). Autophagy has also been
associated with the maintenance of functional liver homeostasis
(14); however, the effects of
autophagy on HIRI remain controversial. Previous studies have
indicated that ALDH2 served a protective role in myocardium IRI and
alcohol-induced chronic hepatic steatosis by inducing autophagy
(6,12). However, to the best of our
knowledge, the potential underlying crosstalk between ALDH2 and
autophagy in HIRI have not yet been characterized.
Therefore, the aim of the present study was to
evaluate the effects of Alda-1 on HIRI and to examine the
underlying mechanisms. It was hypothesized that Alda-1 may
attenuate HIRI by increasing ALDH2 activity, reducing oxidative
stress and apoptosis, inhibiting the inflammatory response and
regulating autophagy.
Materials and methods
Experimental design and animal model
establishment
A total of 48 male inbred Sprague Dawley rats
(weight, 250–300 g) aged 7–8 weeks were obtained from Beijing Vital
River Laboratory Animal Technology Co., Ltd. Rats were raised in
the Animal Experiment Center of The Zhongnan Hospital of Wuhan
University (Wuhan, China). All rats were maintained under standard
animal care conditions at 24±3°C and 60% humidity, with a 12-h
dark/light cycle and free access to food and water. All animal
experiments and protocols were approved by the Committee on the
Experimental Animal Regulations of the Zhongnan Hospital of Wuhan
University (approval no. A237; Wuhan, China) and conformed to the
Guide for the Care and Use of Laboratory Animals (15).
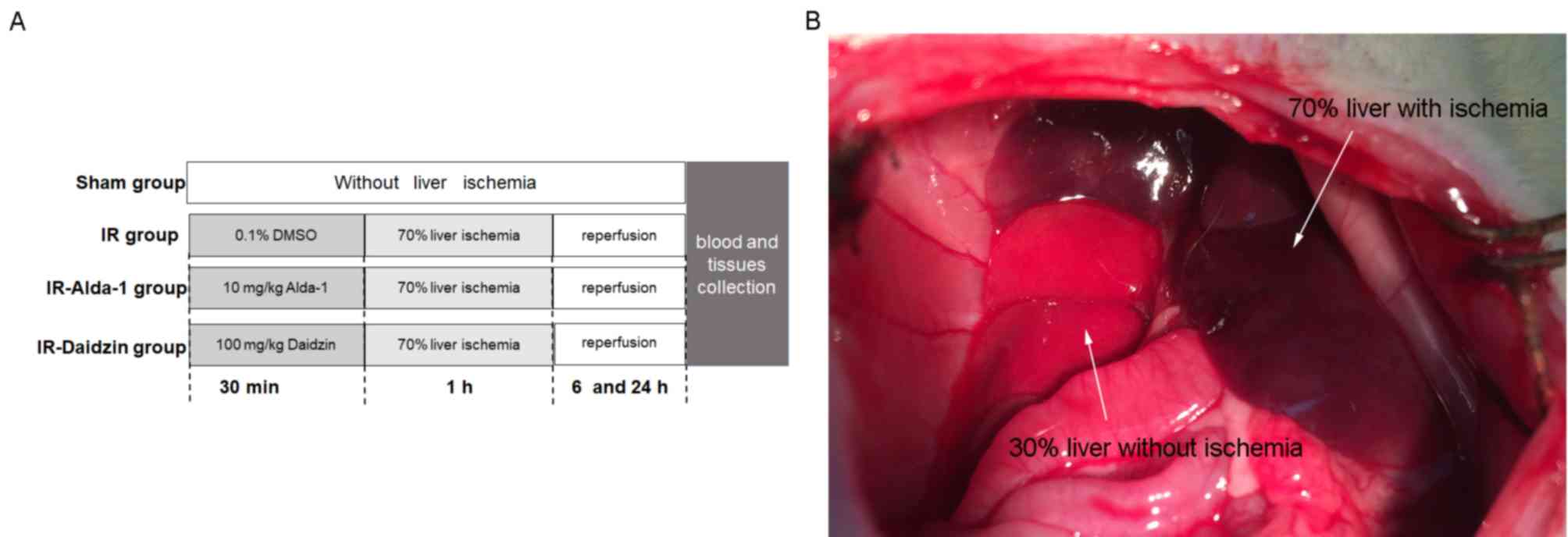
To investigate the effects of Alda-1 on HIRI, rats
were treated intraperitoneally 30 min before warm liver ischemia
with either 10 mg/kg Alda-1 (cat. no. HY-18936; MedChemExpress)
(16) or 100 mg/kg Daidzin (an
ALDH2 inhibitor; cat. no. HY-N0018; MedChemExpress). Both Alda-1
and Daidzin were dissolved in 0.1% DMSO (Sigma-Aldrich; Merck
KGaA). A total of 48 rats were divided into four groups
(n=12/group): i) A sham group, in which rats were given
laparotomies without HIRI; ii) an IR group, where rats were treated
with vehicle (0.1% DMSO) 30 min before laparotomy with HIRI; iii)
an IR-Alda-1 Group, where rats were treated with 10 mg/kg Alda-1 30
min before laparotomy with HIRI; and iv) an IR-Daidzin group, where
rats were treated with 100 mg/kg Daidzin 30 min before laparotomy
with HIRI (Fig. 1A).
Briefly, rats were fasted for 12 h before the
experiment but allowed free access to water. Rats were then
anesthetized with intraperitoneal (IP) injection of 50 mg/kg sodium
pentobarbital. Normal body temperature was maintained using a
heating pad at 37±0.5°C and was monitored with a rectal probe.
Under anesthesia, an abdominal midline laparotomy was performed on
all rats. Partial (70%) liver ischemia was induced in the HIRI
groups by clamping the median and left lobes for 60 min with a
microvascular clip. Hepatic ischemia was confirmed by color changes
in the tissue (Fig. 1B). After the
60 min period of warm ischemia, the clamp was removed, the abdomen
was then closed, and the rats were allowed to recover in a warm
environment. No rats died within 24 h of the laparotomy and no
antibiotics were administrated over the course of the
experiment.
After 6 or 24 h of reperfusion, rats were sacrificed
using 100 mg/kg pentobarbital sodium (IP). After cessation of
breathing and heartbeat, 5 ml blood samples were collected through
inferior vena cava punctures using K2-EDTA test tubes, then
centrifuged at 3,000 × g for 15 min at 4°C to extract serum. Liver
tissue samples were harvested from the median and left lobes of the
ischemic liver. Serum and liver tissue samples were frozen in
liquid nitrogen, then stored at −80°C. A section of each liver
tissue sample was immersed in 4% paraformaldehyde at 25°C for 48 h
for histopathology.
Analysis of liver enzymes and serum
analytes
The levels of alanine aminotransferase (ALT) and
aspartate aminotransferase (AST) released into the serum were
detected using an ALT ELISA kit (cat. no. E-BC-K235-M; Elabscience
Biotechnology, Co., Ltd.) and an AST ELISA kit (cat. no.
E-BC-K236-M; Elabscience Biotechnology, Co., Ltd.) following the
protocols of the clinical laboratory of the Zhongnan Hospital of
Wuhan University. The serum levels of tumor necrosis factor-α
(TNF-α), interleukin (IL)-1β, IL-6 and 4-HNE in the serum were
measured using TNF-α ELISA kit (cat. no. E-EL-R0019c), IL-1β ELISA
kit (cat. no. E-EL-R0012c), IL-6 ELISA kit (cat. no. E-EL-R0015c)
and 4-HNE ELISA kit (cat. no. E-EL-0128c; all from Elabscience
Biotechnology, Co., Ltd.) according to the manufacturers'
protocols.
Histopathology
The samples after paraffin immersion were embedded
in paraffin wax and cut into 5-µm sections for histological
analysis via hematoxylin-eosin staining (HE; hematoxylin staining
for 5–15 min and eosin staining for 1–3 min; all performed at room
temperature). The degree of liver injury was assessed according to
Suzuki's grading scale (17): i)
Grade 0, no congestion, no vacuolization and no necrosis; ii) grade
1, minimal congestion, minimal vacuolization and single-cell
necrosis; iii) grade 2, mild congestion, mild vacuolization and
<30% necrosis; iv) grade 3, moderate congestion, moderate
vacuolization and ≤60% necrosis; and v) grade 4, severe congestion,
severe vacuolization and >60% necrosis. Two blinded pathologists
independently evaluated the sections under a light microscope
(magnification, ×100).
Determination of ALDH2 activity
ALDH2 activity was measured using an ALDH2 activity
assay kit (cat. no. A075-1-1; Nanjing Jiancheng Bioengineering
Institute), according to the manufacturer's protocol. Briefly, the
activity of the ALDH2 enzyme was determined by monitoring the
conversion of NAD+ to NADH at a spectrophotometric
absorbance of 340 nm. ALDH2 activity is expressed as nmol NADH/min
per mg protein.
Oxidative stress analysis
The levels of 4-HNE (cat. no. H268), MDA (cat. no.
A003-1-2) and ROS (cat. no. E004-1-1) in liver tissue samples were
measured using specific rat colorimetric assay kits (Nanjing
Jiancheng Bioengineering Institute), according to the
manufacturers' protocol.
Western blotting
Western blotting was performed as previously
described (18). Briefly, total
protein from liver tissues was extracted using RIPA lysis buffer
(Beyotime Institute of Biotechnology), supplemented with protease
inhibitor and phosphatase inhibitor (Roche Applied Science).
Protein concentration of the supernatant was determined by the
bicinchoninic acid kit (cat. no. G2026; Wuhan Servicebio Technology
Co., Ltd.). Subsequently, 50 µg protein/lane was loaded and
separated by SDS-PAGE on 8–12% gels, then transferred onto PVDF
membranes. The PVDF membranes were blocked with 5% nonfat milk for
1 h at room temperature and incubated overnight at 4°C with one of
the following primary antibodies: Rabbit anti-ALDH2 (1:1,000; cat.
no. 15310-1-AP; ProteinTech Group, Inc.), rabbit anti-4-HNE (1:500;
cat. no. ab46545; Abcam), rabbit anti-high mobility group box 1
(HMGB1; 1:1,000; cat. no. 10829-1-AP; ProteinTech Group, Inc.),
rabbit anti-toll-like receptor 4 (TLR4; 1:1,000; cat. no.
19811-1-AP; ProteinTech Group, Inc.), rabbit anti-beclin1 (1:500;
cat. no. 11306-1-AP; ProteinTech Group, Inc.), rabbit
anti-autophagy-related 7 (ATG7; 1:1,000; cat. no. 67341-1-IG;
ProteinTech Group, Inc.), rabbit anti-p62 (1:1,000; cat. no.
18420-1-AP; ProteinTech Group, Inc.), rabbit anti-Rab7 (1:1,000;
cat. no. 55469-1-AP; ProteinTech Group, Inc.), rabbit
anti-microtubule associated protein 1 light chain 3 α (LC3; 1:500;
cat. no. 14600-1-AP; ProteinTech Group, Inc.), mouse anti-AKT
(1:1,000; cat. no. 10176-2-AP; ProteinTech Group, Inc.), rabbit
anti-phosphorylated (p-)AKT at Thr308 (1:1,000; cat. no. 13038T;
Cell Signaling Technology, Inc.), rabbit anti-mTOR (1:300; cat. no.
20657-1-AP; ProteinTech Group, Inc.), rabbit anti-p-mTOR at Ser2448
(1:1,000; cat. no. 2971S; Cell Signaling Technology, Inc.), rabbit
anti-S6 (1:1,000; cat. no. 14823-1-AP; ProteinTech Group, Inc.),
rabbit anti-p-S6 at Ser240/244 (1:1,000; cat. no. 5364T; Cell
Signaling Technology, Inc.), mouse anti-AMP-activated protein
kinase (AMPK; 1:1,000; cat. no. 2793S; Cell Signaling Technology,
Inc.), rabbit anti-p-AMPK at Thr172 (1:1,000; cat. no. 2535T; Cell
Signaling Technology, Inc.) or rabbit anti-β-actin (1:1,000; cat.
no. 20536-1-AP; ProteinTech Group, Inc.). The following day, the
PVDF membranes were washed three times and incubated with goat
anti-mouse (1:2,000; cat. no. SA00001-1, ProteinTech Group, Inc.)
or goat anti-rabbit (1:2,000; cat. no. SA00001-2, ProteinTech
Group, Inc.) horseradish peroxidase (HRP)-conjugated secondary
antibodies for 2 h at room temperature. Finally, bands were
detected using a chemiluminescent ECL reagent (cat. no. G2020;
Wuhan Servicebio Technology Co., Ltd.). Protein expression bands
were quantified using densitometry analysis with ImageJ v6.0
software (National Institutes of Health) and normalized to
β-actin.
Immunohistochemistry staining and
immunofluorescence
Hepatic HMGB1 and TLR4 expression levels were also
evaluated by immunohistochemical staining. Briefly,
paraffin-embedded 5-µm liver tissue sections were treated with 3%
hydrogen peroxide for 15 min at 37°C to block endogenous peroxidase
activity. Nonspecific binding was blocked by incubating the
sections with 5% BSA (Wuhan Goodbio Technology Co. Ltd.) for 20 min
at room temperature. Liver tissue sections were then incubated with
a polyclonal rabbit anti-HMGB1 (1:100; cat. no. 10829-1-AP;
ProteinTech Group, Inc.) antibody or anti-TLR4 (1:200; cat. no.
19811-1-AP; ProteinTech Group, Inc.) antibody at 4°C overnight,
followed by incubation with polymer-HRP-conjugated anti-rabbit IgG
(1:1,000; cat. no. SA00001-2, ProteinTech Group, Inc.) for 30 min
at room temperature. Lastly, slides were stained for 3 min at room
temperature using a diaminobenzidine (DAB) kit with HRP as the
substrate. Finally, the sections were dehydrated in a graded
ethanol solution and cleared with xylene. The HMGB1 and TLR4
protein expression levels were viewed under a light microscope
(magnification, ×200).
For immunofluorescence, Liver tissue were fixed,
embedded and sectioned using the same method mentioned above.
Sections were blocked in 5% BSA (Beijing Solarbio Science &
Technology Co., Ltd.) at room temperature for 2 h and were
incubated with rabbit anti-IL-6 (cat. no. 21865-1-AP; 1:100;
ProteinTech Group, Inc.) overnight at 4°C. The sections were then
incubated with Alexa Fluor 555-conjugated goat anti-rabbit (cat.
no. P0179; 1:100; Shanghai Biyuntian Biological Co., Ltd.) and DAPI
(cat. no. C1002; Shanghai Biyuntian Biological Co., Ltd.) at room
temperature for 1 h. Results were visualized using a fluorescence
microscope (magnification, ×400) and images were analyzed by ImageJ
software (version 1.51; National Institutes of Health).
Reverse transcription-quantitative PCR
(RT-qPCR)
Total RNA was extracted from 100 mg frozen liver
tissue samples using TRIzol® reagent (Invitrogen; Thermo
Fisher Scientific, Inc.), 20% chloroform and 50% isopropyl alcohol.
Total RNA was subsequently reverse transcribed into cDNA using a
reverse transcription kit (Thermo Fisher Scientific, Inc.),
according to the manufacturer's protocol. RT was performed at 42°C
for 1 h followed by an incubation at 75°C for 5 min. The
thermocycling conditions were as follows: 50°C for 2 min, 95°C for
10 min, followed by 40 cycles of 95°C for 10 sec and 60°C for 30
sec. RT-PCR was subsequently performed using SYBR Green (Shanghai
Yeasen Biotechnology Co. Ltd.) to determine the expression levels
of target genes. The relative expression of genes was calculated
using the 2−∆∆Cq method. Gene expression was normalized
against that of the β-actin gene (19). All primers used for the RT-qPCR are
listed in Table I.
 | Table I.Nucleotide sequences of primers used
for reverse transcription-quantitative PCR. |
Table I.
Nucleotide sequences of primers used
for reverse transcription-quantitative PCR.
| Gene | Primer sequence
(5′→3′) |
|---|
| High
mobility | F:
GGCGGCTGTTTTGTTGACAT |
| group box
1 | R:
ACCCAAAATGGGCAAAAGCA |
|
Toll-like | F:
TGTATCGGTGGTCAGTGTGC |
| receptor
4 | R:
CAGCTCGTTTCTCACCCAGT |
|
Interferon-γ | F:
GAGGAACTGGCAAAAGGACG |
|
| R:
AGGTGCGATTCGATGACACT |
| IL-1β | F:
GAGGCTGACAGACCCCAAAAGA |
|
| R:
TCCACAGCCACAATGAGTGACA |
| IL-6 | F:
AGCGATGATGCACTGTCAGA |
|
| R:
GGAACTCCAGAAGACCAGAGC |
| Tumor | F:
GTGATCGGTCCCAACAAGGA |
|
necrosis-α | R:
TTTGCTACGACGTGGGCTAC |
| β-actin | F:
TGCTATGTTGCCCTAGACTTCG |
|
| R:
GTTGGCATAGGTCTTTACGG |
TUNEL staining
The liver tissue was fixed in 4% paraformaldehyde
solution at room temperature for 24 h, paraffin-embedded,
dehydrated in a graded ethanol series and coronally cut into 5 µm
sections. Sections were deparaffinized using xylene and rehydrated
in a descending ethanol series. Tunel assay (Roche Diagnostics) was
used to detect quantitatively the apoptotic hepatocytes according
to the manufacturer's protocol. Liver tissue sections were
incubated with fluorescein-dUTP at 37°C for 1 h. 6 visual fields
were randomly selected under a fluorescence microscope. The normal
hepatocytes were stained blue and the apoptosis-positive cells
green. Total hepatocytes and TUNEL-positive cells were counted
under a fluorescence microscope (magnification, ×200). The
apoptotic rate was calculated as: (The number of TUNEL-positive
cells/the total number of hepatocytes) ×100%. Analysis was carried
out using Image Pro Plus 6.0 software (Media Cybernetics,
Inc.).
Transmission electron microscopy
(TEM)
The 1–2 mm3 fresh liver tissue fixed in
2.5% glutaraldehyde solution was rinsed in PBS and fixed with 1%
osmium tetroxide for 2 h at room temperature. Following dehydration
with a graded acetone series and embedding in Pon812 epoxy resin
for 12 h at room temperature, the liver tissue was cut into 1 µm
sections. The sections were stained with 3% uranyl acetate at room
temperature for 30 min and washed with double distilled water.
Then, the sections were stained with 3% lead citrate at room
temperature for 10 min and washed with double distilled water. The
mitochondrial injuries was observed by Tecnai G2 20 Twin
TE microscope (FEI; Thermo Fisher Scientific, Inc.) at room
temperature: 5 fields were selected for each specimen for
statistical analysis.
Statistical analysis
Statistical analysis was performed using SPSS 16.0
statistical software (SPSS, Inc.). Data are presented as the mean ±
SD. For experimental data following a normal distribution,
statistical differences were determined using an ANOVA, followed by
a Tukey's post-hoc test. Otherwise, nonparametric data were
analyzed using the Kruskal-Wallis test, followed by a Dunn's
post-hoc test. P<0.05 was considered to indicate a statistically
significant difference.
Results
Alda-1 pretreatment ameliorates HIRI
in rats
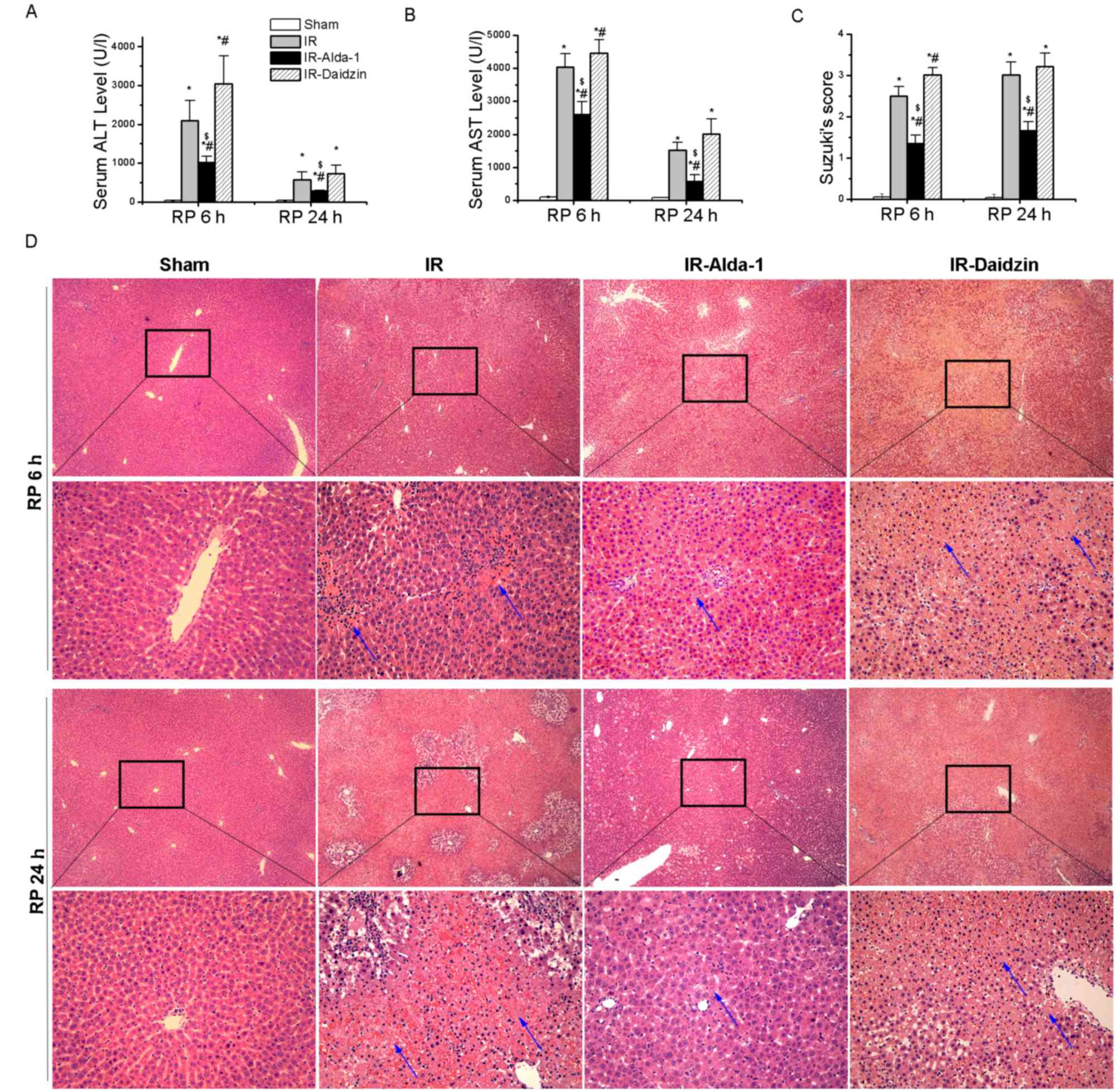
After 70% warm hepatic ischemia for 60 min, followed
by 6 or 24 h of reperfusion, the serum levels of ALT and AST were
significantly increased in the IR group compared with the sham
group (Fig. 2A and B). However,
the pretreatment with Alda-1 prior to HIRI decreased the serum
levels of ALT and AST after 6 and 24 h of reperfusion compared with
the IR and IR-Daidzin groups. These results were supported by the
histological lesions observed in the liver tissues with H&E
staining (Fig. 2C and D).
Extensive necrosis (indicated by the white arrows), congestion and
neutrophil infiltration were observed in the IR and IR-Daidzin
groups after 6 and 24 h of reperfusion. These lesions were
significantly reduced in the IR-Alda-1 group compared with the IR
group, based on Suzuki's HIRI scoring system (17). These results suggested that ALDH2
may serve a role in HIRI and that ALDH2 activation may confer
protection from HIRI.
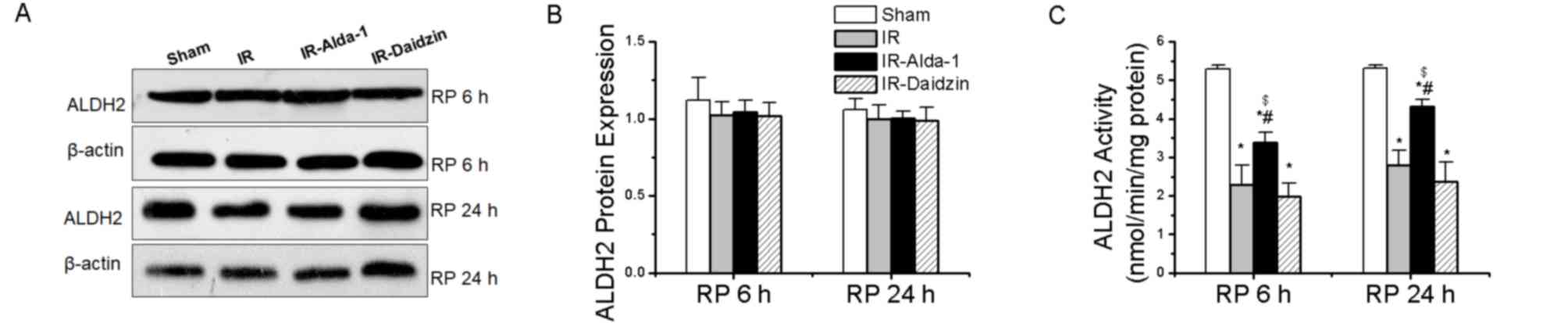
ALDH2 activity and expression in HIRI
model rats
Changes in the ALDH2 expression levels were then
evaluated in HIRI rats. ALDH2 protein expression levels were not
significantly different among the four groups at either 6 or 24 h
(Fig. 3A and B). However, HIRI
significantly decreased ALDH2 activity in the IR group compared
with rats in the sham group at both 6 and 24 h. This effect was
partially rescued by ALDH2 activation in the IR-Alda-1 group
(Fig. 3C).
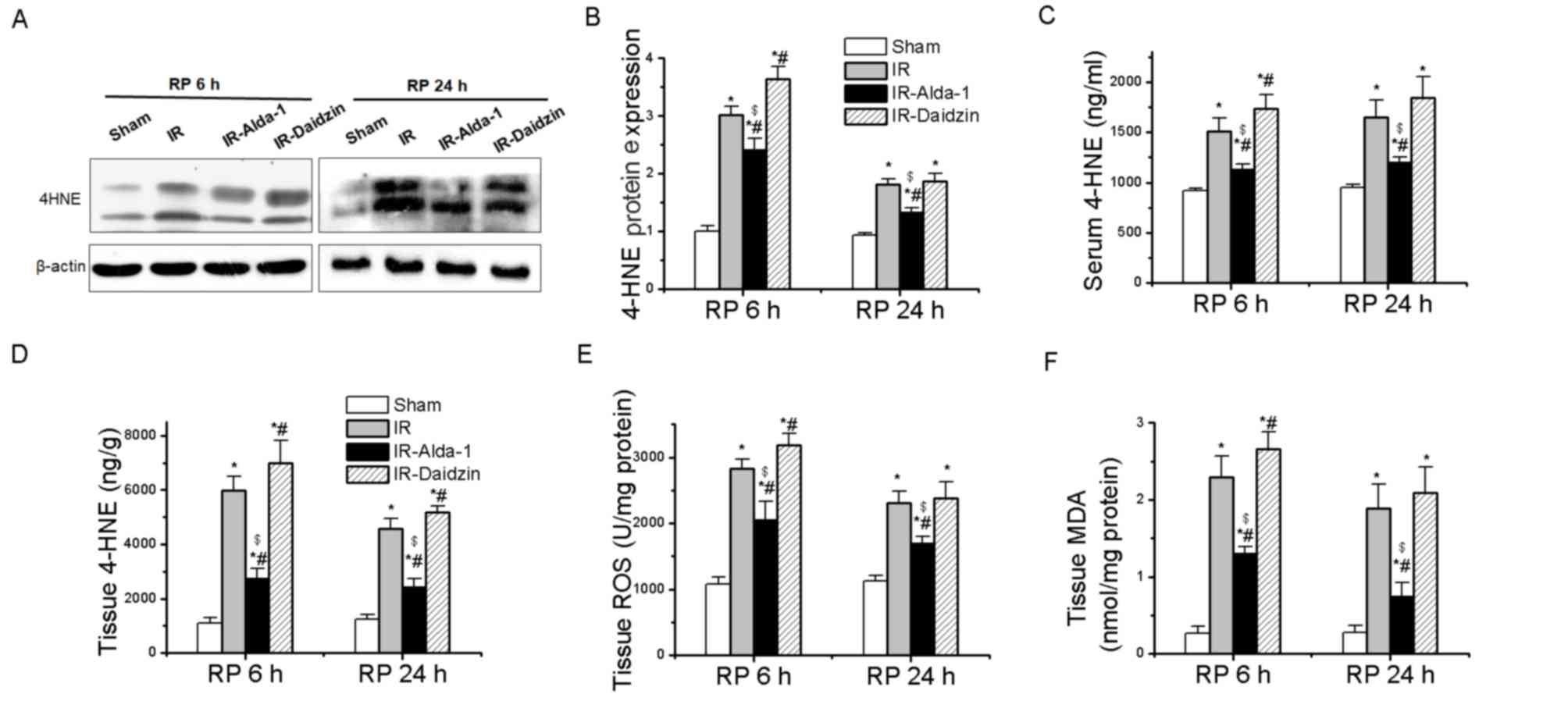
Effects of Alda-1 on 4-HNE
accumulation and oxidative stress in HIRI model rats
The accumulation of ROS, MDA and 4-HNE aldehydes has
been discovered to promote toxic effects associated with HIRI
(4,8,9). A
significant increase in 4-HNE protein expression levels, serum
levels and tissue accumulation was observed following IR and 6 or
24 h reperfusion compared with the sham group (Fig. 4A-D), while these increases were
partially reversed in the IR-Alda-1 group. To investigate the
relationship between the ALDH2-mediated liver protection and
oxidative stress, the levels of ROS and MDA were measured in the
liver tissue; Alda-1 pretreatment reduced the production of ROS and
MDA in hepatic tissues following 6 and 24 h of reperfusion compared
with the IR and IR-Daidzin groups (Fig. 4E and F).
Alda-1 pretreatment improves liver
mitochondrial damage and attenuates hepatocyte apoptosis in HIRI
model rats
Mitochondrial changes in the hepatocytes were
observed after 24 h of reperfusion using TEM. Substantial
mitochondrial damage occurred in the IR group hepatocytes compared
with the sham group (Fig. 5A).
However, decreased mitochondrial edema was observed in the
IR-Alda-1 group (indicated in the figure by arrows). TUNEL staining
also indicated that Alda-1 pretreatment attenuated hepatocyte
apoptosis 24 h after reperfusion compared with the IR and
IR-Daidzin groups (Fig. 5B and
C).
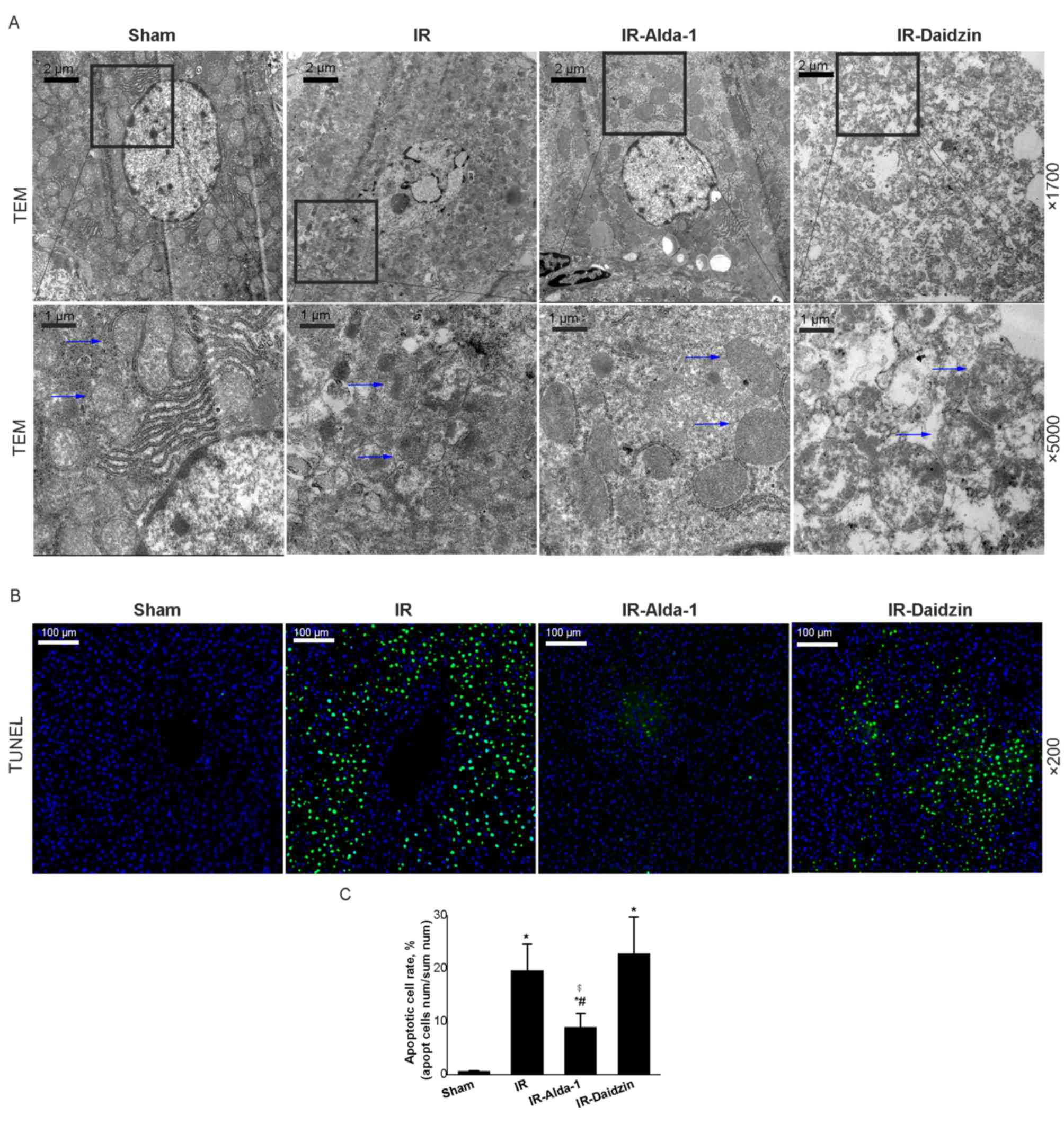 | Figure 5.Alda-1 pretreatment reduces damage to
liver mitochondria and hepatocyte apoptosis in HIRI model rats. (A)
Representative transmission electron micrographs showing
mitochondria in the liver tissue after 24 h of reperfusion. Blue
arrows indicate mitochondria. Magnification, ×1,700 and ×5,000.
Scale bars, 1 or 2 µm. (B) Representative TUNEL-stained images
showing HIRI rat livers after 24 h of reperfusion. Magnification,
×200; scale bar, 100 µm. (C) TUNEL-stained apoptotic cell rate of
each group. Data are presented as the mean ± SD. n=6. *P<0.05
vs. the sham group; #P<0.05 vs. the IR group;
$P<0.05 vs. the IR-Daidzin group. RP, reperfusion;
HIRI, hepatic ischemia/reperfusion injury; IR,
ischemia/reperfusion; TEM, transmission electron microscope. |
Alda-1 pretreatment inhibits
inflammatory responses in HIRI model rats
HMGB1 and TLR4 have been reported to be involved in
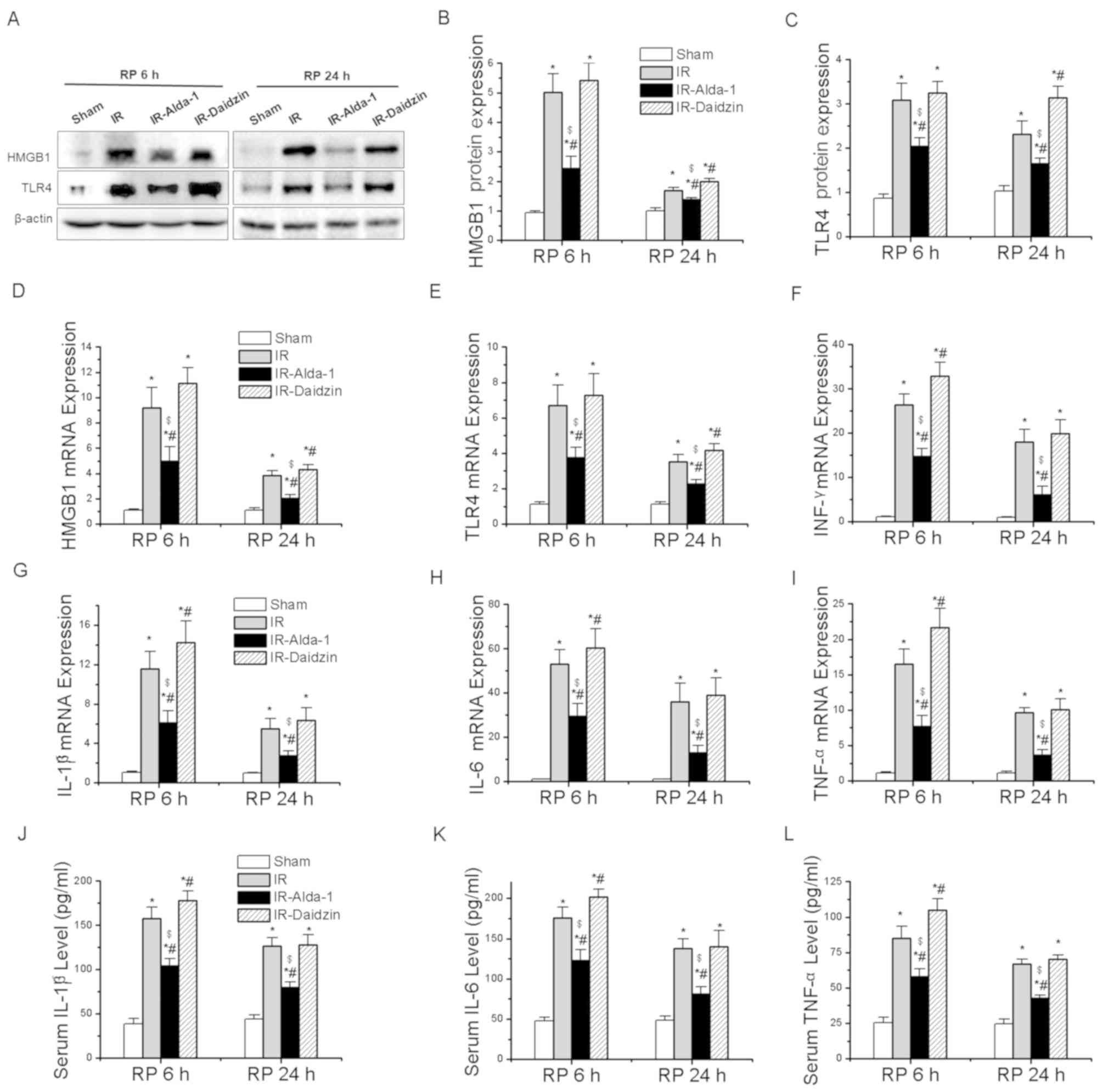
inflammatory responses associated with HIRI (4). After 6 and 24 h of reperfusion, the
protein expression levels of HMGB1 and TLR4 were significantly
upregulated in the IR group compared with the sham group (Fig. 6A-C). Meanwhile, Alda-1 pretreatment
significantly inhibited the protein expression levels of HMGB1 and
TLR4 compared with the IR group.
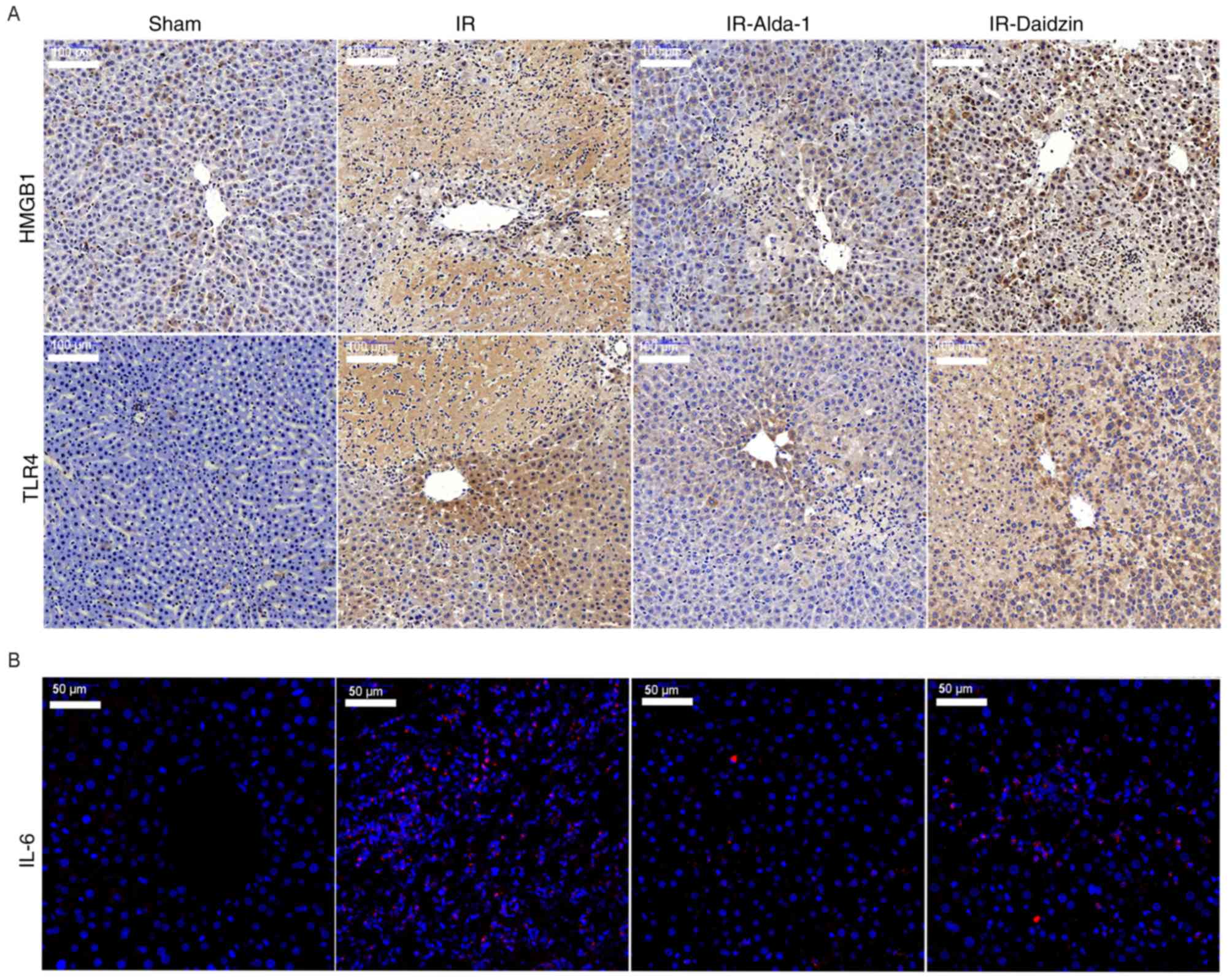
 | Figure 6.Alda-1 pretreatment inhibits the
inflammatory response. (A) Western blotting was used to analyze the
expression levels of HMGB1 and TLR4 in liver tissues after 6 and 24
h of reperfusion. Relative expression was normalized against the
β-actin loading control. Densitometric semi-quantification of (B)
HMGB1 and (C) TLR4 protein expression levels from part. (A) mRNA
expression levels of (D) HMGB1, (E) TLR4, (F) IFN-γ, (G) IL-1β, (H)
IL-6 and (I) TNF-α after 6 and 24 h of reperfusion. Serum levels of
(J) IL-1β, (K) IL-6 and (L) TNF-α after 6 and 24 h of reperfusion.
Data are presented as the mean ± SD. n=6. *P<0.05 vs. the sham
group; #P<0.05 vs. the IR group;
$P<0.05 vs. the IR-Daidzin group. RP, reperfusion;
HMGB1, high mobility group box 1; TLR4, toll-like receptor 4;
IFN-γ, interferon-γ; IL, interleukin; TNF-α, tumor necrosis
factor-α; IR, ischemia/reperfusion. |
RT-qPCR was then used to analyze the mRNA expression
levels of HMGB1, TLR4 and other downstream inflammatory factors.
Alda-1 pretreatment significantly reduced the mRNA expression
levels of HMGB1, TLR4, interferon-γ (IFN-γ), IL-1β, IL-6 and TNF-α
at 6 and 24 h after reperfusion, compared with the mRNA expression
levels of these genes in the IR and IR-Daidzin groups (Fig. 6D-I). Similar trends were also
observed in the serum levels of IL-1β, IL-6 and TNF-α after 6 and
24 h of reperfusion (Fig. 6J-K).
The results of the immunohistochemical staining of HMGB1 and TLR4
expression levels, and of IL-6 immunofluorescence, were consistent
with the western blotting, ELISA and RT-qPCR data (Fig. 7). Thus, these findings indicated
that Alda-1 pretreatment may protect the liver against HIRI by
inhibiting the HMGB1/TLR4 inflammatory pathway.
Alda-1 pretreatment restores
HIRI-induced suppression of autophagy
To evaluate the effects of Alda-1 on HIRI-induced
autophagy, the protein expression levels of Beclin1 (important for
autophagy nucleation) (20), ATG7
(a mediator of autophagosome formation) (12), p62 (a specific protein adaptor
degraded by autophagy) (20), Rab7
(a small GTPase protein that stimulates lysosomal biogenesis and
autophagic vacuole maturation) (12) and LC3 (important for
autophagolysosome formation) (12)
were evaluated. The expression level of Beclin1 protein showed no
significant differences between the groups (Fig. 8B). Compared with the sham group,
the IR group had significantly downregulated expression levels of
ATG7 and Rab7, a significantly reduced LC3II/I ratio and
significantly upregulated protein expression levels of p62 after 6
and 24 h of reperfusion (Fig. 8A,
C-F). These effects were significantly reversed in the
IR-Alda-1 group (Fig. 8A, C-F).
Thus, these results strongly suggested that Alda-1 pretreatment may
significantly alleviate HIRI by inducing autophagy.
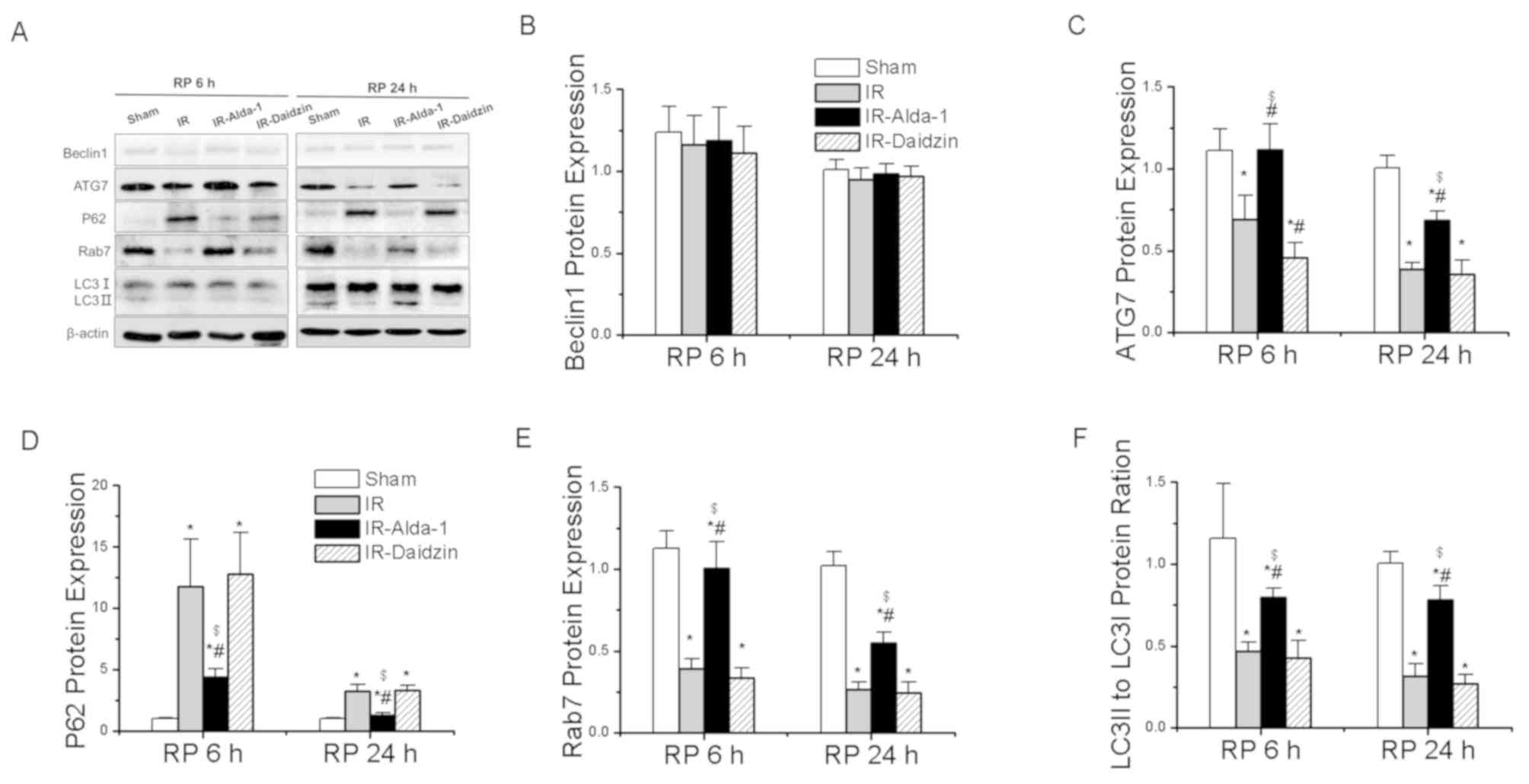 | Figure 8.Alda-1 pretreatment reverses the
hepatic IR injury-induced suppression of autophagy. (A) Western
blotting was used to analyze the expression levels of Beclin1,
ATG7, p62, Rab7 and LC3 in the liver tissues after 6 and 24 h of
reperfusion. Relative expression was normalized against the β-actin
loading control. Densitometric semi-quantification of the
expression levels of (B) Beclin1, (C) ATG7, (D) p62, (E) Rab7 and
(F) LC3 ratio from part. (A) Data are presented as the mean ± SD.
n=6. *P<0.05 vs. the sham group; #P<0.05 vs. the
IR group; $P<0.05 vs. the IR-Daidzin group. ATG7,
autophagy-related 7; LC3, microtubule associated protein 1 light
chain 3 α; IR, ischemia/reperfusion; RP, reperfusion. |
Alda-1 pretreatment-induced autophagy
in HIRI is mediated by AKT/mTOR and AMPK signaling activation
To assess the potential regulatory factors
underlying autophagy induction, the activity levels of the AKT/mTOR
and AMPK signaling pathways were analyzed. Compared with the sham
group, the IR group had significantly downregulated phosphorylation
levels of AKT at Thr308 and of AMPK at Thr172, while upregulated
expression levels of p-mTOR at Ser2448 and p-S6 at Ser240/244 were
observed (Fig. 9A-E). These
effects were significantly reversed in the IR-Alda-1 group. Thus,
these findings suggested that ALDH2-induced autophagy in HIRI may
be dependent on the AKT/mTOR and AMPK signaling pathways.
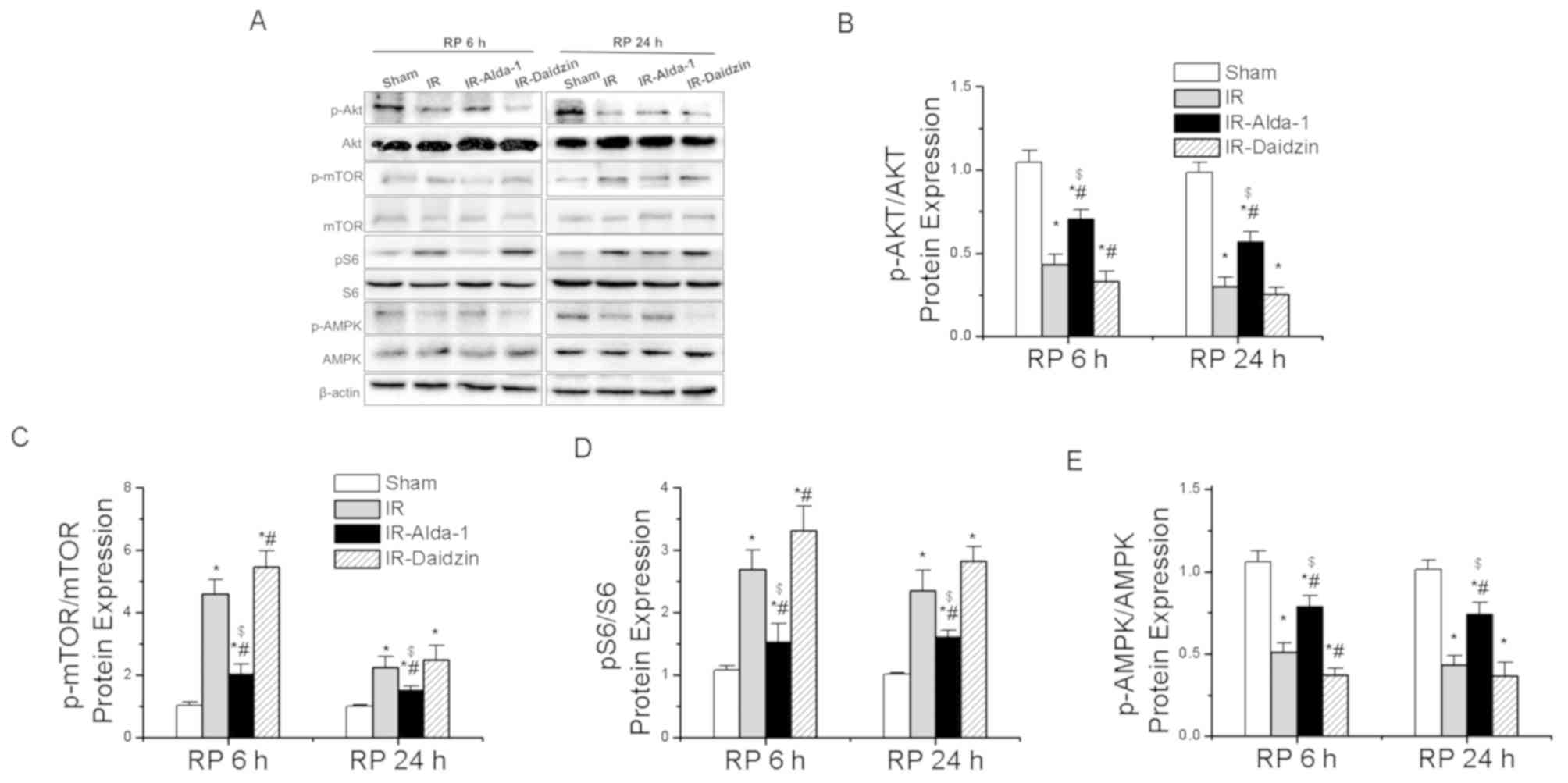 | Figure 9.Alda-1 pretreatment enhances
autophagy in rats with hepatic IR injury through activation of
AKT/mTOR and AMPK signaling. (A) Western blotting was used to
analyze the expression levels of AKT, p-AKT at Thr308, mTOR, p-mTOR
at Ser2448, S6, p-S6 at Ser240/244, AMPK and p-AMPK at Thr172 in
liver tissues after 6 and 24 h of reperfusion. Relative expression
was normalized against the β-actin loading control. Densitometric
semi-quantification of (B) the ratio of p-AKT protein expression to
total AKT protein expression, (C) the ratio of p-mTOR protein
expression to total mTOR protein expression, (D) the ratio of p-S6
protein expression to total S6 protein expression and (E) p-AMPK
protein expression to total AMPK protein expression from part. (A)
Data are presented as the mean ± SD. n=6. *P<0.05 vs. the sham
group; #P<0.05 vs. the IR group;
$P<0.05 vs. the IR-Daidzin group. mTOR, mammalian
target of rapamycin; AMPK, AMP-activated protein kinase α; p,
phosphorylated; IR, ischemia/reperfusion; RP, reperfusion. |
Discussion
HIRI is a complex pathophysiological process that
often leads to poor clinical prognosis. Clinically, there are no
effective means to alleviate HIRI. The aim of the present study was
to evaluate the effects of Alda-1 on HIRI and to examine the
underlying mechanisms. In the present study, the pretreatment with
Alda-1, an ALDH2 activator, reduced the negative effects of HIRI,
including hepatic enzyme injury, 4-HNE levels, oxidative stress,
hepatocyte apoptosis and inflammation. The observed decrease in
HIRI-induced hepatic necrosis, oxidative stress and inflammatory
responses were revealed to be associated with the inhibition of
autophagy. In addition, Alda-1 was discovered to protect the liver
following HIRI by enhancing autophagy and restoring the autophagy
flux. It was also identified that the Alda-1-induced autophagy may
be dependent on the AKT/mTOR and AMPK signaling pathways.
Previous studies have suggested that Alda-1 had a
protective effect on myocardial IRI (5) and liver injuries caused by chronic
alcohol intake (12,21). However, the mechanisms underlying
the effects of Alda-1 pretreatment on HIRI remain poorly
understood. To address this knowledge gap, a rat model of 70% HIRI
was established to investigate whether Alda-1 pretreatment could
reduce the negative effects of HIRI and to characterize the
underlying mechanisms. Warm ischemia caused liver function damage
after 6 and 24 h of reperfusion. The pretreatment with Alda-1 led
to a significant reduction in the serum levels of ALT and AST.
Extensive hepatocyte necrosis, the congestion of the blood cells in
the liver sinusoid and neutrophil infiltration all were identified
in H&E-stained histological sections from the IR group after 6
and 24 h of reperfusion. These effects were markedly reduced in the
group pretreated with Alda-1. Thus, Alda-1 pretreatment was
suggested to exert a strong protective effect against HIRI in the
rat model. However, the mechanism through which Alda-1 protects the
liver against HIRI remains to be clarified.
Previous studies have reported that Alda-1 exerts
beneficial effects on ischemia injury in other organ models,
including the intestine (22),
kidney (23) and brain (24). Changes in ALDH2 expression levels
were evaluated in HIRI rats; however, in the present study, it was
discovered that Alda-1 did not affect the ALDH2 protein expression
levels. Nonetheless, HIRI decreased ALDH2 activity. More
importantly, the ALDH2 activity in HIRI rats was improved by Alda-1
pretreatment.
Reperfusion injury has been reported to results in
the intracellular and extracellular release of ROS, which may be
generated by mitochondrial dysfunction (3,25).
Furthermore, reperfusion injury was discovered to promote the
accumulation of toxic reactive aldehydes, such as MDA and 4-HNE
(26,27). Doorn et al (11) identified that 4-HNE accumulation
was increased by myocardial ischemia injury reperfusion in mice,
and that this increase was reversed by ALDH2. In addition, ROS, MDA
and 4-HNE have all been identified as important markers of
oxidative stress in HIRI (4,8,9,28),
and oxidative stress is associated with all stages of HIRI
(28). Thus, strategies aimed at
reducing oxidative stress may also improve HIRI. In the present
study, consistent with previous studies, HIRI induced mitochondrial
damage, including an increase in 4-HNE, ROS and MDA production.
Restoration of ALDH2 activity eliminated the accumulation of 4-HNE,
ROS and MDA. Traverso et al (29) suggested that the formation of the
4-HNE adduct decreased ALDH2 activity. In addition, during
reperfusion injury, mitochondrial dysfunction and oxidative stress
were identified to contribute to cell death (4). In the present study, TUNEL staining
of the liver sections indicated that fewer apoptosis-positive cells
were present following Alda-1 pretreatment compared with the IR
group. These findings suggested that Alda-1 pretreatment may
improve HIRI by reducing oxidative stress, mitochondrial injury and
hepatocyte apoptosis.
Damage-associated molecular pattern (DAMP) signaling
has been found to serve a critical role in HIRI pathogenesis
(4,30). DAMPs are produced by apoptotic and
necrotic hepatocytes (4). At the
start of reperfusion, DAMPs have been observed to rapidly activate
the innate immune system and initiate an inflammatory response
(4). Moreover, the reintroduction
of blood was found to exacerbate ROS generation, leading to
necrotic liver cell death (31),
further increasing DAMP release (4). The increased release of endogenous
DAMPs was discovered to be detrimental to organ function,
negatively affecting HMGB1 and TLRs activity (32). Furthermore, the release of
hepatocyte HMGB1 has been identified to critically depend on
intracellular ROS accumulation (33) and HMGB1 was discovered to be
scavenged by TLR4 (34). In the
present study, it was hypothesized that Alda-1 pretreatment may
protect liver tissue against HIRI by inhibiting inflammatory
responses. The expression levels of HMGB1, TLR4, IFN-γ, IL-1β, IL-6
and TNF-α were significantly increased in the IR group. However,
ALDH2 activation by Alda-1 pretreatment inhibited the expression
levels of these proteins, indicating that Alda-1 may improve HIRI
by inhibiting the inflammatory response.
Autophagy is a dynamic, intracellular, degradative
process that targets damaged organelles, dysfunctional proteins and
harmful products (20). Autophagy
is induced under hypoxia and ischemia (35), and involves the formation of
autolysosomes and autophagosomes (20). Several previous studies indicated
that autophagy could protect the liver from HIRI, and that the
inhibition of autophagy in HIRI could increase cell death (36–38).
Kim et al (39)
demonstrated that HIRI impaired autophagy in mice, which was
associated with a marked decrease in the expression levels of LC3,
ATG7 and Beclin1. This finding suggested that the enhancement of
autophagy might represent a novel treatment option for HIRI.
Moreover, Wang et al (38)
demonstrated that increasing autophagy ameliorated liver damage and
improved mitochondrial function against HIRI in aged mice. Liu
et al (40) also suggested
that ischemic pre-conditioning ameliorated HIRI in rats by inducing
autophagy. Altogether, these previous studies strongly suggested
that autophagy may be a protective mechanism against HIRI.
Guo et al (12) demonstrated that ALDH2 ameliorated
hepatic damage due to chronic alcohol intake via the induction of
autophagy. However, the potential mechanisms underlying the
association between ALDH2 and autophagy in HIRI remain uncertain.
In the present study, Alda-1 pretreatment increased autophagy in
HIRI. Thus, it could be suggested that Alda-1 pretreatment may
protect the liver via the induction of autophagy.
Further studies are required to identify the
mechanisms regulating autophagy during Alda-1 pretreatment of HIRI.
Zhang et al (41)
demonstrated that activated p-AKT significantly attenuated HIRI. In
addition, activated p-AKT has been demonstrated to serve a pivotal
role in autophagy (42). In
cardiomyocyte IRI, glucose deprivation induced autophagy through
the inhibition of p-mTOR and the activation of p-AMPK (43). Interestingly, p-mTOR was reported
to be negatively regulated by p-AKT (44). Ma et al (6) demonstrated that ALDH2 rescued
myocardial IRI by regulating autophagy via AKT/mTOR and AMPK
signaling. In the present study, Alda-1 pretreatment upregulated
the expression levels of p-AKT and p-AMPK and downregulated the
expression levels of p-mTOR. Thus, ALDH2-induced autophagy might be
dependent on the AKT/mTOR and AMPK signaling pathways, and these
pathways may serve a role in the observed protective effects of
Alda-1 pretreatment on HIRI.
However, the present study had two experimental
limitations. The study design was based on animal experiments in
vivo, and the results were not verified in vitro. In
addition, the role of ALDH2 in conferring protection against HIRI
was not confirmed using ALDH2 overexpression or knockdown
tools.
In conclusion, Alda-1, an ALDH2 activator, was
discovered to alleviate HIRI-induced effects, including hepatic
enzyme injuries, acetaldehyde accumulation, oxidative stress,
hepatocyte apoptosis and inflammation, possibly through the
induction of autophagy. Thus, ALDH2 may be a potential target of
new pharmacological strategies applicable to HIRI clinical
practice.
Acknowledgements
Not applicable.
Funding
The present study was supported by the Medical
Science Advancement Program (Clinical Medicine) of Wuhan University
(grant no. TFLC 2018003) and the Program of Excellent Doctoral
(Postdoctoral) of Zhongnan Hospital of Wuhan University (grant no.
ZNYB2019008).
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
ZL and QY designed and performed the research,
analyzed the data and wrote the article. XF, LL, YX and ZZ
performed the experiments, contributed ideas and helped write the
article. ZL, XZ, PY, CL, JL, XH, WY and WW helped establish the
in vivo model. YW provided guidance and revised the article.
YW, QY and SY designed the experiments, provided overall guidance
and helped with the manuscript. All authors read and approved the
final manuscript.
Ethics approval and consent to
participate
All animal experiments and protocols were approved
by The Committee on the Experimental Animal Regulations of The
Zhongnan Hospital of Wuhan University (approval no. A237; Wuhan,
China) and conformed to The Guide for the Care and Use of
Laboratory Animals (15).
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
Glossary
Abbreviations
Abbreviations:
|
4-HNE
|
4-hydroxy-2-nonenal
|
|
ALDH2
|
aldehyde dehydrogenase 2
|
|
ALT
|
alanine aminotransferase
|
|
AST
|
aspartate aminotransferase
|
|
H&E
|
hematoxylin & eosin
|
|
HIRI
|
hepatic ischemia/reperfusion
injury
|
|
HMGB1
|
high mobility group box 1
|
|
IFN-γ
|
interferon-γ
|
|
IL
|
interleukin
|
|
IP
|
intraperitoneal injection
|
|
MDA
|
malondialdehyde
|
|
mTOR
|
mammalian target of rapamycin
|
|
ROS
|
reactive oxygen species
|
|
TNF-α
|
tumor necrosis factor-α
|
|
TLR4
|
toll-like receptor 4
|
References
|
1
|
Selzner N, Rudiger H, Graf R and Clavien
PA: Protective strategies against ischemic injury of the liver.
Gastroenterology. 125:917–936. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Uchida Y, Ke B, Freitas MC, Yagita H,
Akiba H, Busuttil RW, Najafian N and Kupiec-Weglinski JW: T-cell
immunoglobulin mucin-3 determines severity of liver
ischemia/reperfusion injury in mice in a TLR4-dependent manner.
Gastroenterology. 139:2195–2206. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Peralta C, Jiménez-Castro MB and
Gracia-Sancho J: Hepatic ischemia and reperfusion injury: Effects
on the liver sinusoidal milieu. J Hepatol. 59:1094–1106. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
4
|
van Golen RF, van Gulik TM and Heger M:
The sterile immune response during hepatic ischemia/reperfusion.
Cytokine Growth Factor Rev. 23:69–84. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Gomes KM, Bechara LR, Lima VM, Ribeiro MA,
Campos JC, Dourado PM, Kowaltowski AJ, Mochly-Rosen D and Ferreira
JC: Aldehydic load and aldehyde dehydrogenase 2 profile during the
progression of post-myocardial infarction cardiomyopathy: Benefits
of Alda-1. Int J Cardiol. 179:129–138. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Ma H, Guo R, Yu L, Zhang Y and Ren J:
Aldehyde dehydrogenase 2 (ALDH2) rescues myocardial
ischaemia/reperfusion injury: Role of autophagy paradox and toxic
aldehyde. Eur Heart J. 32:1025–1038. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Chen CH, Ferreira JC, Gross ER and
Mochly-Rosen D: Targeting aldehyde dehydrogenase 2: New therapeutic
opportunities. Physiol Rev. 94:1–34. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Chen CH, Sun L and Mochly-Rosen D:
Mitochondrial aldehyde dehydrogenase and cardiac diseases.
Cardiovasc Res. 88:51–57. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Yoval-Sánchez B and Rodríguez-Zavala JS:
Differences in susceptibility to inactivation of human aldehyde
dehydrogenases by lipid peroxidation byproducts. Chem Res Toxicol.
25:722–729. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Gomes KM, Campos JC, Bechara LR, Queliconi
B, Lima VM, Disatnik MH, Magno P, Chen CH, Brum PC, Kowaltowski AJ,
et al: Aldehyde dehydrogenase 2 activation in heart failure
restores mitochondrial function and improves ventricular function
and remodelling. Cardiovasc Res. 103:498–508. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Doorn JA, Hurley TD and Petersen DR:
Inhibition of human mitochondrial aldehyde dehydrogenase by
4-hydroxynon-2-enal and 4-oxonon-2-enal. Chem Res Toxicol.
19:102–110. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Guo R, Xu X, Babcock SA, Zhang Y and Ren
J: Aldehyde dedydrogenase-2 plays a beneficial role in ameliorating
chronic alcohol-induced hepatic steatosis and inflammation through
regulation of autophagy. J Hepatol. 62:647–656. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Chen CH, Budas GR, Churchill EN, Disatnik
MH, Hurley TD and Mochly-Rosen D: Activation of aldehyde
dehydrogenase-2 reduces ischemic damage to the heart. Science.
321:1493–1495. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Czaja MJ, Ding WX, Donohue TJ Jr, Friedman
SL, Kim JS, Komatsu M, Lemasters JJ, Lemoine A, Lin JD, Ou JH, et
al: Functions of autophagy in normal and diseased liver. Autophagy.
9:1131–1158. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
National Research Council, . Guide for the
care and use of laboratory animals. (8th). The National Academies
Press. (Washington, DC). 2011.
|
|
16
|
Ji W, Wei S, Hao P, Xing J, Yuan Q, Wang
J, Xu F and Chen Y: Aldehyde dehydrogenase 2 has cardioprotective
effects on myocardial ischaemia/reperfusion injury via suppressing
mitophagy. Front Pharmacol. 7:1012016. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Suzuki S, Toledo-Pereyra LH, Rodriguez FJ
and Cejalvo D: Neutrophil infiltration as an important factor in
liver ischemia and reperfusion injury. Modulating effects of FK506
and cyclosporine. Transplantation. 55:1265–1272. 1993. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Liu Z, Zhang X, Xiao Q, Ye S, Lai CH, Luo
J, Huang X, Wang W, Zeng C, Zhong Z, et al: Pretreatment donors
after circulatory death with simvastatin alleviates liver ischemia
reperfusion injury through a KLF2-dependent mechanism in rat. Oxid
Med Cell Longev. 2017:38619142017. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Cursio R, Colosetti P and Gugenheim J:
Autophagy and liver ischemia-reperfusion injury. Biomed Res Int.
2015:4175902015. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Zhong W, Zhang W, Li Q, Xie G, Sun Q, Sun
X, Tan X, Sun X, Jia W and Zhou Z: Pharmacological activation of
aldehyde dehydrogenase 2 by Alda-1 reverses alcohol-induced hepatic
steatosis and cell death in mice. J Hepatol. 62:1375–1381. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Zhu Q, He G, Wang J, Wang Y and Chen W:
Pre-treatment with the ALDH2 agonist Alda-1 reduces intestinal
injury induced by ischaemia and reperfusion in mice. Clin Sci
(Lond). 131:1123–1136. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Zhong Z, Hu Q, Fu Z, Wang R, Xiong Y,
Zhang Y, Liu Z, Wang Y and Ye Q: Increased expression of aldehyde
dehydrogenase 2 reduces renal cell apoptosis during
ischemia/reperfusion injury after hypothermic machine perfusion.
Artif Organs. 40:596–603. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Fu SH, Zhang HF, Yang ZB, Li TB, Liu B,
Lou Z, Ma QL, Luo XJ and Peng J: Alda-1 reduces cerebral
ischemia/reperfusion injury in rat through clearance of reactive
aldehydes. Naunyn Schmiedebergs Arch Pharmacol. 387:87–94. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Caraceni P, Domenicali M, Vendemiale G,
Grattagliano I, Pertosa A, Nardo B, Morselli-Labate AM, Trevisani
F, Palasciano G, Altomare E and Bernardi M: The reduced tolerance
of rat fatty liver to ischemia reperfusion is associated with
mitochondrial oxidative injury. J Surg Res. 124:160–168. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Horton JW and Walker PB: Oxygen radicals,
lipid peroxidation, and permeability changes after intestinal
ischemia and reperfusion. J Appl Physiol (1985). 74:1515–1520.
1993. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Lee H, Ko EH, Lai M, Wei N, Balroop J,
Kashem Z and Zhang M: Delineating the relationships among the
formation of reactive oxygen species, cell membrane instability and
innate autoimmunity in intestinal reperfusion injury. Mol Immunol.
58:151–159. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
van Golen RF, van Gulik TM and Heger M:
Mechanistic overview of reactive species-induced degradation of the
endothelial glycocalyx during hepatic ischemia/reperfusion injury.
Free Radic Biol Med. 52:1382–1402. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Traverso N, Menini S, Odetti P, Pronzato
MA, Cottalasso D and Marinari UM: Diabetes impairs the enzymatic
disposal of 4-hydroxynonenal in rat liver. Free Radic Biol Med.
32:350–359. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Zhang Y, Yuan D, Yao W, Zhu Q, Liu Y,
Huang F, Feng J, Chen X, Huang Y, Chi X and Hei Z: Hyperglycemia
aggravates hepatic ischemia reperfusion injury by inducing chronic
oxidative stress and inflammation. Oxid Med Cell Longev.
2016:39196272016. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Bhogal RH, Curbishley SM, Weston CJ, Adams
DH and Afford SC: Reactive oxygen species mediate human hepatocyte
injury during hypoxia/reoxygenation. Liver Transpl. 16:1303–1313.
2010. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Gracia-Sancho J, Villarreal GJ Jr, Zhang
Y, Yu JX, Liu Y, Tullius SG and García-Cardeña G: Flow cessation
triggers endothelial dysfunction during organ cold storage
conditions: Strategies for pharmacologic intervention.
Transplantation. 90:142–149. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Tsung A, Klune JR, Zhang X, Jeyabalan G,
Cao Z, Peng X, Stolz DB, Geller DA, Rosengart MR and Billiar TR:
HMGB1 release induced by liver ischemia involves Toll-like receptor
4 dependent reactive oxygen species production and calcium-mediated
signaling. J Exp Med. 204:2913–2923. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Tsung A, Sahai R, Tanaka H, Nakao A, Fink
MP, Lotze MT, Yang H, Li J, Tracey KJ, Geller DA and Billiar TR:
The nuclear factor HMGB1 mediates hepatic injury after murine liver
ischemia-reperfusion. J Exp Med. 201:1135–1143. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Dong Y, Undyala VV, Gottlieb RA, Mentzer
RJ Jr and Przyklenk K: Autophagy: Definition, molecular machinery,
and potential role in myocardial ischemia-reperfusion injury. J
Cardiovasc Pharmacol Ther. 15:220–230. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Liu A, Fang H, Dahmen U and Dirsch O:
Chronic lithium treatment protects against liver
ischemia/reperfusion injury in rats. Liver Transpl. 19:762–772.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Wang JH, Ahn IS, Fischer TD, Byeon JI,
Dunn WA Jr, Behrns KE, Leeuwenburgh C and Kim JS: Autophagy
suppresses age-dependent ischemia and reperfusion injury in livers
of mice. Gastroenterology. 141:2188–2199. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Wang JH, Behrns KE, Leeuwenburgh C and Kim
JS: Critical role of autophage in ischemia/reperfusion injury to
aged livers. Autophagy. 8:140–141. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Kim JS, Nitta T, Mohuczy D, O'Malley KA,
Moldawer LL, Dunn WA Jr and Behrns KE: Impaired autophagy: A
mechanism of mitochondrial dysfunction in anoxic rat hepatocytes.
Hepatology. 47:1725–1736. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Liu A, Fang H, Wei W, Dirsch O and Dahmen
U: Ischemic preconditioning protects against liver
ischemia/reperfusion injury via heme oxygenase-1-mediated
autophagy. Crit Care Med. 42:e762–e771. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Zhang R, Zhang L, Manaenko A, Ye Z, Liu W
and Sun X: Helium preconditioning protects mouse liver against
ischemia and reperfusion injury through the PI3K/Akt pathway. J
Hepatol. 61:1048–1055. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Arico S, Petiot A, Bauvy C, Dubbelhuis PF,
Meijer AJ, Codogno P and Ogier-Denis E: The tumor suppressor PTEN
positively regulates macroautophagy by inhibiting the
phosphatidylinositol 3-kinase/protein kinase B pathway. J Biol
Chem. 276:35243–35246. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Matsui Y, Takagi H, Qu X, Abdellatif M,
Sakoda H, Asano T, Levine B and Sadoshima J: Distinct roles of
autophagy in the heart during ischemia and reperfusion: Roles of
AMP-activated protein kinase and Beclin 1 in mediating autophagy.
Circ Res. 100:914–922. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Inoki K, Li Y, Zhu T, Wu J and Guan KL:
TSC2 is phosphorylated and inhibited by Akt and suppresses mTOR
signalling. Nat Cell Biol. 4:648–657. 2002. View Article : Google Scholar : PubMed/NCBI
|