Introduction
Hepatic fibrosis is a risk factor in the progression
of end-stage liver diseases, including cirrhosis and hepatocellular
carcinoma. It is characterized by the progressive accumulation of
extracellular matrix (ECM), such as smooth muscle α-actin (α-SMA)
and type I collagen (1). Hepatic
stellate cells (HSCs) are a major source of ECM and serve important
roles in hepatic fibrosis (2,3).
Generally, the renin angiotensin system (RAS) is
recognized as a manager of blood pressure and hydroelectrolyte
balance. In recent years, accumulating evidence has demonstrated
that RAS can also modulate the proliferation of HSCs, and the
process of hepatic fibrosis via the profibrotic effector,
angiotensin (Ang) II (4–6). The major biological actions of Ang II
are mediated by the type 1 angiotensin receptor (AT1R); Ang II
stimulates HSC migration, pro-collagen α1 expression, and the
secretion of transforming growth factor (TGF) β1 and inflammatory
cytokines (7). In addition,
blocking Ang II activity by lisinopril [an angiotensin-converting
enzyme (ACE) inhibitor] or losartan (an AT1R antagonist) prevents
the pro-fibrogenic effects of RAS (8,9). In
different models of hepatic fibrosis, including bile duct ligation,
CCl4 treatment or continuous Ang II infusion, blocking
the activity of Ang II by ACE inhibitor or AT1R antagonism also
allivates hepatic fibrosis (10–12).
Although angiotensinogen, which is a precursor for all angiotensin
peptides, and its cleavage enzyme, renin, are not found to be
elevated after liver injury, the levels of ACE and AT1R are
markedly upregulated after liver injury, particularly in the
fibrotic areas and activated HSCs (13,14).
These findings suggest that the RAS, especially Ang II, plays a
vital role in liver fibrosis development, mainly through the
activation of HSCs.
Traditional Chinese Medicine has been widely used in
the treatment of a number of liver diseases. Angelica
sinensis has been a commonly used medical herb in Traditional
Chinese Medicine for thousands of years. It has been reported that
Angelica sinensis can reverse high-fat diet-induced liver
steatosis (15) and prevent
CCL4-induced acute hepatic injury in rats (16). Levistilide A (Lev A), the volatile
oil extract of Angelica sinensis, inhibits HSC proliferation
activated by platelet-derived growth factor BB by inducing cell
cycle arrest and apoptosis (17).
In preliminary experiments, Lev A also showed a significant effect
on the proliferation and activation of HSCs induced by Ang II (data
not shown). Therefore, the present study investigated the
anti-hepatic fibrosis effect of Lev A on the the regulation of the
RAS.
Materials and methods
HSC isolation and culture
A total of 10 normal male Sprague-Dawley rats (6–8
weeks old, >400 g) were obtained from The Shanghai Laboratory
Animal Center, Chinese Academy of Sciences Shanghai, China. Rats
were housed in a temperature controlled environment (23±2°C), at a
relative humidity of 60±10%, with a 12-h light/dark cycle and free
access to food and water. Primary HSCs were purified from
Sprague-Dawley rats by perfusion with pronase and collagenase,
followed by Nycodenz density-gradient centrifugation at 1,450 g for
22 min at 25°C (18). The isolated
cells were cultured with Dulbecco's modified Eagle's medium (DMEM;
Gibco; Thermo Fisher Scientific, Inc.) supplemented with 20% fetal
bovine serum (FBS; Gibco; Thermo Fisher Scientific, Inc.). On the
third day, the primary HSCs were used for subsequent experiments.
The protocol was approved by the Committee on the Ethics of Animal
Experiments of Shanghai University of Traditional Chinese Medicine,
China. All animals received humane care during the study.
Cell proliferation analysis
HSCs were plated into 96-well plates and incubated
at 37°C for 24 h. Next, the cells were treated with 0.1 µM Ang II
(Sigma-Aldrich; Merck KGaA) with or without Lev A (2.4, 6 and 15
µM; Shanghai Winherb Medical Technology Co., Ltd.,) or losartan (1
µM; Sigma-Aldrich; Merck KGaA) for another 24 h at 37°C. Cell
proliferation was detected using the Cell-Light
5-ethynyl-2′-deoxyuridine (EdU) Cell Proliferation assay kit (cat.
no. 10310; Guangzhou RiboBio Co., Ltd.) according to the
manufacturer's protocols. The cell nuclei were stained with
4′,6-diamidino-2-phenylindole (Sigma-Aldrich; Merck KGaA) at a
concentration of 1 µg/ml for 10 min at room temperature.
EdU-positive and EdU-negative cells were determined by fluorescent
imaging. Images were taken using Cellomics ArrayScan VTI HCS Reader
and analyzed using Cellomics Cell Health Profiling BioApplication
software (version 3.1; Thermo Fisher Scientific, Inc.).
Immunofluorescence
HSCs were seeded into 96-well plates. After 24 h,
the cells were incubated with Ang II (0.1 µM) with or without Lev A
(2.4, 6 and 15 µM) for another 24 h at 37°C. The cells were then
washed twice with cold phosphate-buffered saline (PBS) and fixed in
4% paraformaldehyde for 10 min at 4°C. F-actin and α-SMA were
determined by immunofluorescence staining as previously described
(19). Images were captured using
Cellomics ArrayScan VTI HCS Reader and analyzed using Cellomics
Cell Health Profiling BioApplication software (Thermo Fisher
Scientific, Inc.).
CCL4-induced hepatic
fibrosis
A total of 24 male Wistar rats (6–8 weeks old;
240–260 g) were obtained from the Shanghai Laboratory Animal
Center, Chinese Academy of Sciences Shanghai, China. The animals
were housed with a 12-h light/dark cycle under temperature control
at 23±2°C, with a relative humidity of 60±10% and free access to
food and water. The rats were randomly assigned into 3 groups:
Control group, model group and Lev A group. The animals in model
and Lev A groups given intraperitoneal injections of
CCL4 (5 ml/kg) for three consecutive days per week over
a period of 4 weeks (20). From
the third week of CCL4 injections, rats in the Lev A
group were administered with Lev A at a dose of 3 mg/kg/day. Rats
in the control and model groups were treated with an equal amount
of vehicle. After 2 weeks of administration, rats were
anaesthetized with 2% pentobarbital sodium and the blood samples
were obtained from the inferior vena cava. A portion of each liver
was fixed in 10% phosphate-buffered formalin for 24 h at room
temperature for histological studies following paraffin embedding.
The remainder was snap-frozen in liquid nitrogen and stored at
−80°C for western blot analysis. The protocol was approved by the
Committee on the Ethics of Animal Experiments of Shanghai
University of Traditional Chinese Medicine, China. All animals
received humane care during the study.
Hematoxylin and eosin (H&E) and
sirius red staining
The hematoxylin-eosin and sirius red staining were
performed as previously described (21). Briefly, paraffin embedded-liver
samples were cut into 4 µm-thick sections and then stained with
H&E and sirius red. A random selection six microscopic fields
were observed at a magnification of ×200 with an inverted
microscope (Mode: IX70, Olympus Corporation).
Serum biochemical measurements
Kits were used to measure levels of serum alanine
aminotransferase (ALT; cat. no. C009-2-1), aspartate
aminotransferase (AST cat. no. C010-2-1), albumin (Alb; cat. no.
A028-2-1) and total bilirubin (TBil; cat. no. C019-1-1), according
to the manufacturer's protocols (Nanjing Jiancheng Bioengineering
Institute), as previously described (21).
Western blot analysis
Total protein was isolated from HSCs and liver
tissue of rats using radioimmunoprecipitation assay buffer (150 mM
NaCl, 1% Nonidet P-40, 0.1% SDS, 50 mM Tris-HCl pH 7.4, 1 mM EDTA,
1 mM PMSF, 1X Roche complete mini protease inhibitor cocktail,
Roche PhosSTOP phosphatase inhibitor cocktail; Beyotime Institute
of Biotechnology). The protein concentration was determined by the
bicinchoninic acid method (Thermo Fisher Scientific, Inc.). Equal
amounts of protein (20 µg) were separated via SDS-PAGE on a 10% gel
under denaturing and non-reducing conditions and then transferred
to a nitrocellulose membrane. The membrane was blocked with odyssey
blocking buffer (LI-COR Biosciences) at room temperature for 1 h
and then incubated with primary antibodies against AT1R (1:200;
cat. no. sc-1173; Santa Cruz Biotechnology, Inc.), phosphorylated
(p)-ERK1/2 (1:200; cat. no. sc-7383; Santa Cruz Biotechnology,
Inc.), total ERK2 (1:200; cat. no. sc-1647; Santa Cruz
Biotechnology, Inc.), GAPDH (1:5,000; cat. no. sc-166574; Santa
Cruz Biotechnology, Inc.), α-SMA (1:1,000; cat. no. ab5649; Abcam),
c-Jun (1:1,000; cat. no. 9165; Cell Signaling Technology, Inc.) and
p-c-Jun (1:1,000; cat. no. 9261; Cell Signaling Technology, Inc.)
at 4°C overnight. Following washing in PBS with 0.1% Tween-20, the
blots were incubated for 1 h at room temperature with a
IRdye® 680CW anti-rabbit (1:20,000; cat. no. 926-32223)
or IRDye 800CW anti-mouse secondary antibody (1:20,000; cat. no.
926-32210), both purchased from LI-COR Biosciences. The signals
were visualized and analyzed using Li-Cor Odyssey Imaging software
(version 2.1; Li-Cor Biosciences).
Statistical analysis
Data are expressed as means ± standard deviation,
and were analyzed using one-way analysis of variance, followed by a
post hoc least significant difference test. Statistical analysis
was conducted using SPSS version 18.0 software (SPSS, Inc.).
P<0.05 was considered to indicate a statistically significant
difference.
Results
Lev A inhibits HSC proliferation
After 24 h incubation, Ang II significantly
increased the cell number of HSCs compared with the control, as
determined by EdU assay (P<0.01; Fig. 1A and B). Co-treatment with Lev A
significantly decreased the number of HSCs proliferating at
concentrations of 6 and 15 µM (P<0.01). However, 2.4 µM of Lev A
exhibited little effect on the proliferation Ang II-induced HSCs.
The blocker of AT1R, losartan, also significantly suppressed the
Ang II-induced proliferation of HSCs (P<0.01; Fig. 1A and B).
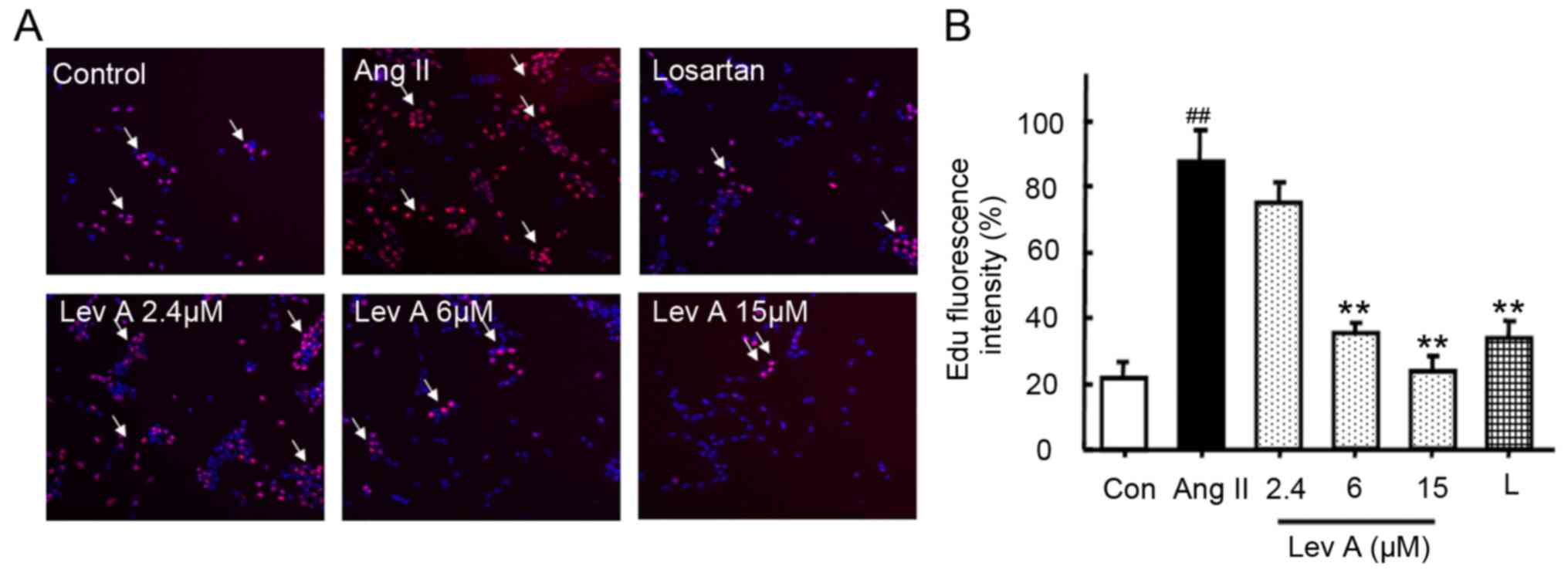 | Figure 1.Lev A inhibits Ang II-induced HSCs
proliferation. Following treatment with Ang II (0.1 µM) and
indicated concentrations of Lev A and losartan for 24 h, cell
proliferation was determined by the EdU method. (A) HSC
proliferation was determined by EdU staining (magnification, ×100;
red, EdU; blue, DAPI). (B) Quantification of the fluorescence
intensity. Each bar represents the means ± standard deviation
(n=6). ##P<0.01 vs. control group; **P<0.01 vs.
Ang II group. Lev A, levistilide A; Ang II, angiotensin II; HSCs,
hepatic stellate cells; EdU, 5-ethynyl-2′-deoxyuridine; DAPI,
4′,6-diamidino-2-phenylindole; L, losartan. |
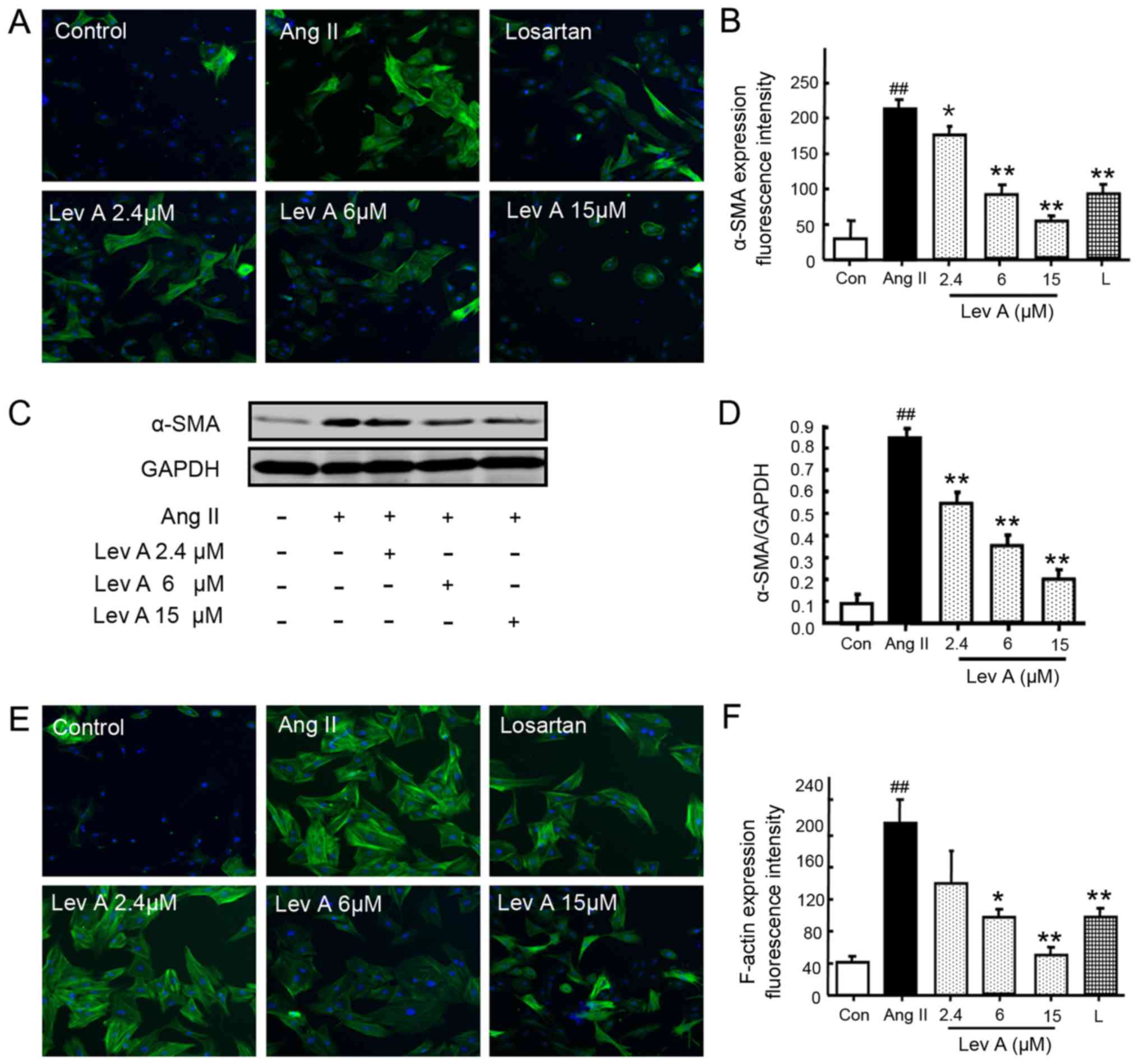
Lev A inhibits the activation of
HSCs
To investigate the effect of Lev A on the activation
of Ang II-induced HSCs, the expression of α-SMA and F-actin was
determined. Ang II treatment significantly enhanced the expression
of α-SMA (P<0.01), whereas co-treatment with losartan or Lev A
significantly suppressed Ang II-induced α-SMA upregulation
(P<0.05; Fig. 2A and B).
Western blot analysis also demonstrated that Lev A inhibited Ang
II-induced α-SMA upregulation (P<0.01; Fig. 2C and D). The present study also
determined the expression of F-actin after Ang II treatment using
immunofluorescence staining. Ang II induced a significant
upregulation of F-actin (P<0.01; Fig. 2E and F) and Lev A partly reversed
the effect of Ang II on F-actin (P<0.05).
Lev A suppresses Ang II-induced AT1R
upregulation and activation of ERK and c-Jun
Our previous study (21) revealed that Ang II is able to
increase the level of its receptor AT1R in HSCs and so the effect
of Lev A on AT1R expression was examined. Ang II treatment
significantly upregulated the expression of AT1R (P<0.01;
Fig. 3A and B). Losartan and Lev A
treatment both repressed Ang II-induced AT1R upregulation
(P<0.01). Activation of ERK and c-Jun is involved in Ang
II-induced HSCs proliferation and activation (12,21).
Therefore, the effect of Lev A on the activation of ERK and c-Jun
was also studied. Ang II treatment induced significant increases of
p-ERK1/2 and p-c-Jun (P<0.01; Fig.
3A, C and D). Pretreatment with Lev A significantly inhibited
the phosphorylation of ERK1/2 and c-Jun induced by Ang II
(P<0.05).
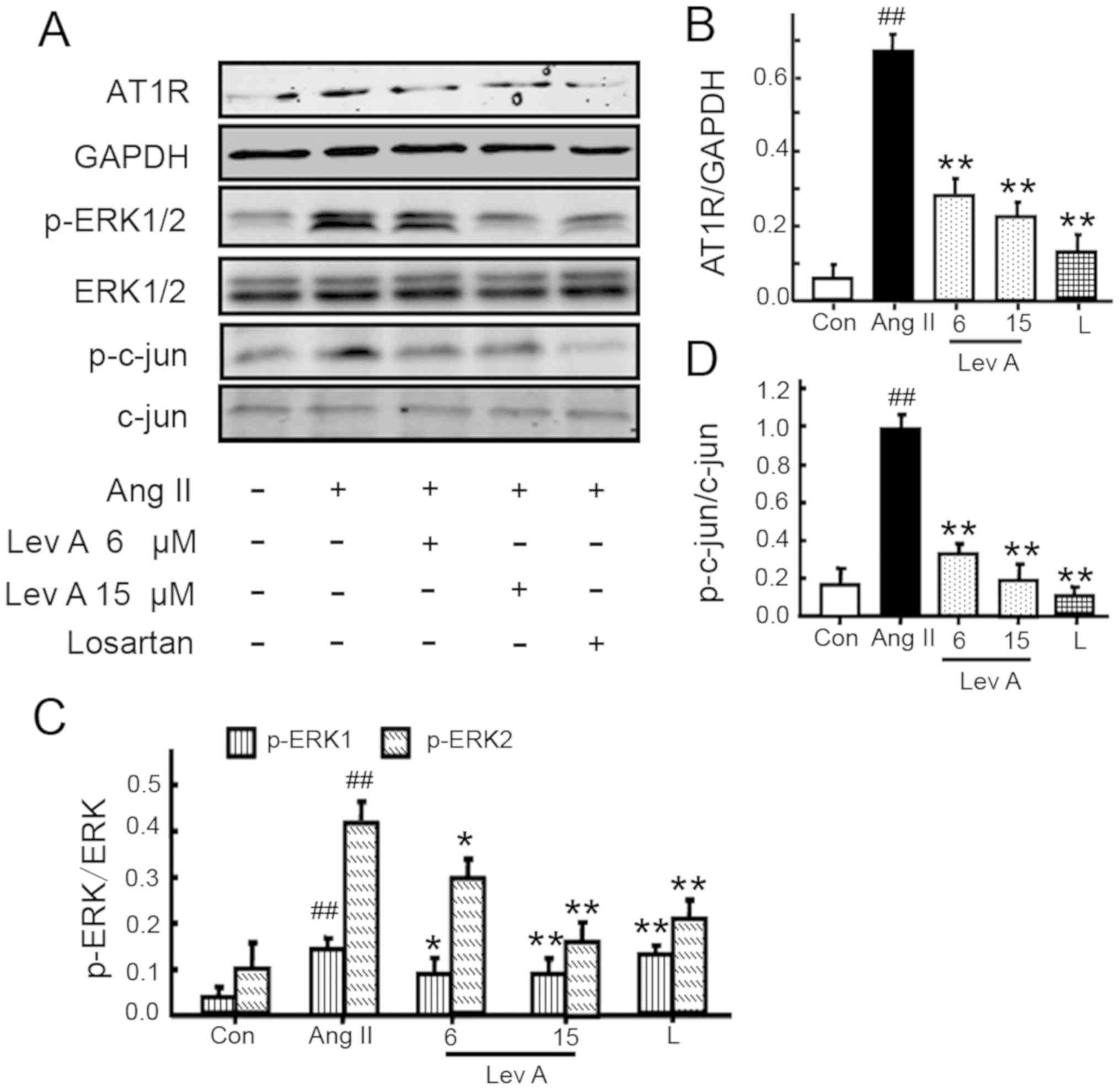 | Figure 3.Effect of Lev A on Ang II-induced RAS
molecule expression in vitro. (A) Effect of Lev A on the
expression of AT1R and phosphorylation of ERK and c-Jun. (B-D)
Quantitative data of AT1R, p-ERK and p-c-Jun, respectively. HSCs
were pre-treated with Lev (6 or 15 µM) or losartan (1 µM) for 2 h,
then treated with Ang II (0.1 µM) for different durations. Total
protein was isolated and subjected to western blot analysis. Each
bar represents the mean ± standard deviation of three independent
experiments. ##P<0.01 vs. control group; *P<0.05,
**P<0.01 vs. Ang II group. Lev A, levistilide A; Ang II,
angiotensin II; RAS, renin angiotensin system; AT1R, type 1
angiotensin receptor; p-, phosphorylated; HSCs, hepatic stellate
cells; Con, control; L, losartan. |
Lev A improves CCL4-induced
rat hepatic injury
To evaluate the in vivo effect of Lev A, a
CCL4-induced rat hepatic fibrosis model was used.
CCL4 significantly decreased the body weight, and
increased the liver/body weight ratio (P<0.01; Fig. 4A and B). Treatment with Lev A
significantly improved the liver/body weight ratio (P<0.01) but
it did not alter the body weight. In the model group,
CCL4 treatment significantly decreased the albumin level
and increased the ALT, AST, γ-GT, IBIL and TBil levels (P<0.01;
Fig. 4C-H). Lev A treatment
significantly improved the liver functions of experimental rats
with hepatic fibrosis. H&E staining demonstrated that
CCL4 induced evident centrilobular hepatic necrosis and
infiltrating lymphocytes, and Lev A treatment partly prevented the
histopathological changes (Fig.
4I).
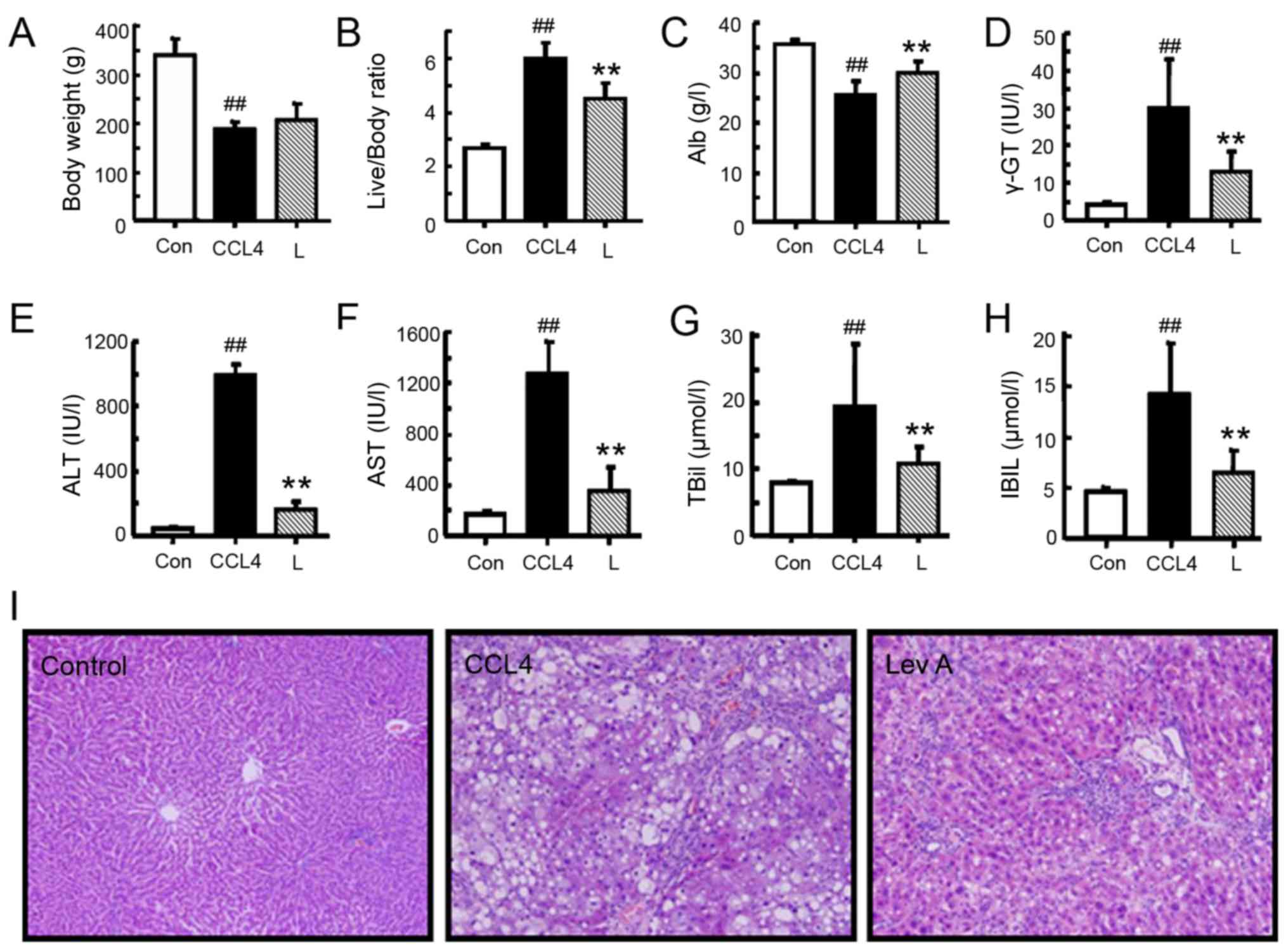 | Figure 4.Effect of Lev A on the body weight
and liver function of CCL4-treated rats. (A) Body weight
and (B) liver/body weight ratio. Levels of (C) Alb, (D) γ-GT, (E)
ALT, (F) AST, (G) TBiL and (H) IBIL. (I) H&E staining of the
rat liver (original magnification, ×200). Each bar represents the
means ± standard deviation (n=8). ##P<0.01 vs.
control group; **P<0.01 vs. model group. Lev A, levistilide A;
Alb, plasma albumin; γ-GT, γ-glutamyl transpeptidase; ALT, alanine
aminotransferase; AST, aspartate aminotransferase; TBiL, total
bilirubin; IBIL, indirect bilirubin; H&E, hematoxylin and
eosin; Con, control; L, losartan. |
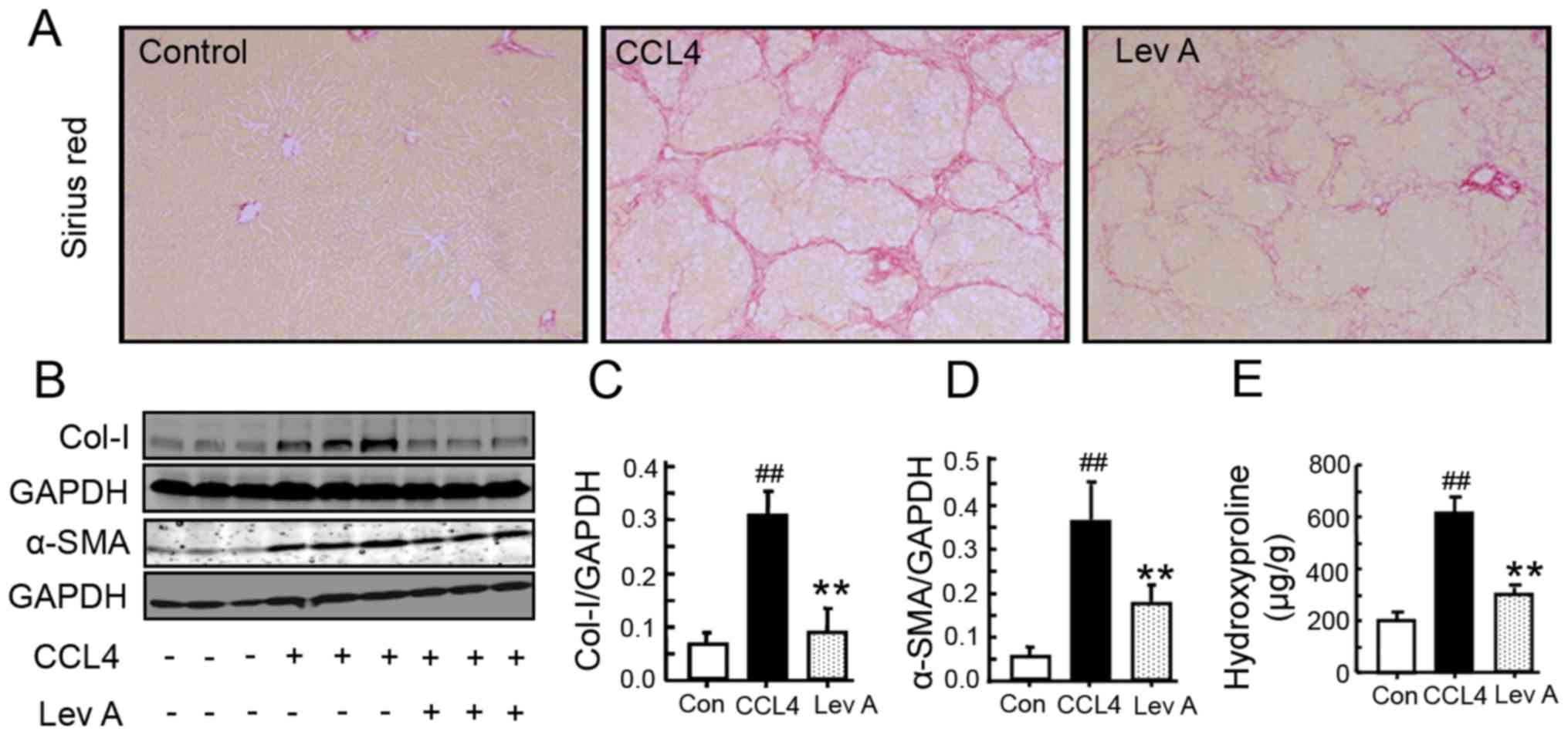
Lev A ameliorates
CCL4-induced liver fibrosis in rats
CCL4 treatment led to a notable
deposition of collagen in rat livers as determined by sirius red
staining (Fig. 5A). Treatment with
Lev A for 2 weeks resulted in an alleviation of collagen deposition
(Fig. 5A). As determined by
western blotting, α-SMA and collagen I expression was significantly
increased in CCL4-treated rats and was suppressed by Lev
A (P<0.01; Fig. 5B-D). In
CCL4-treated rats, the hydroxyproline (Hyp) content was
also significantly upregulated compared with the control group
(P<0.01; Fig. 5E). Treatment
with Lev A significantly decreased the Hyp content (P<0.01).
Lev A regulates the RAS signaling
molecules in vivo
To further evaluate the role of Lev A on RAS in
vivo, the molecules of ACE-Ang II-AT1R axis in rats were next
determined. In the model group, the expression of AT1R was
significantly upregulated compared with the control group
(P<0.01; Fig. 6A and B). p-ERK,
the downstream molecule of Ang II-AT1R, was also significantly
increased in the model group compared with the control (P<0.05;
Fig. 6A and C). Lev A treatment
significantly decreased the CCL4-induced increase of
AT1R, Ang II and p-ERK (P<0.01). Compared with the control
group, the plasma Ang II level was also significantly increased in
the CCL4 group (P<0.01; Fig. 6D).
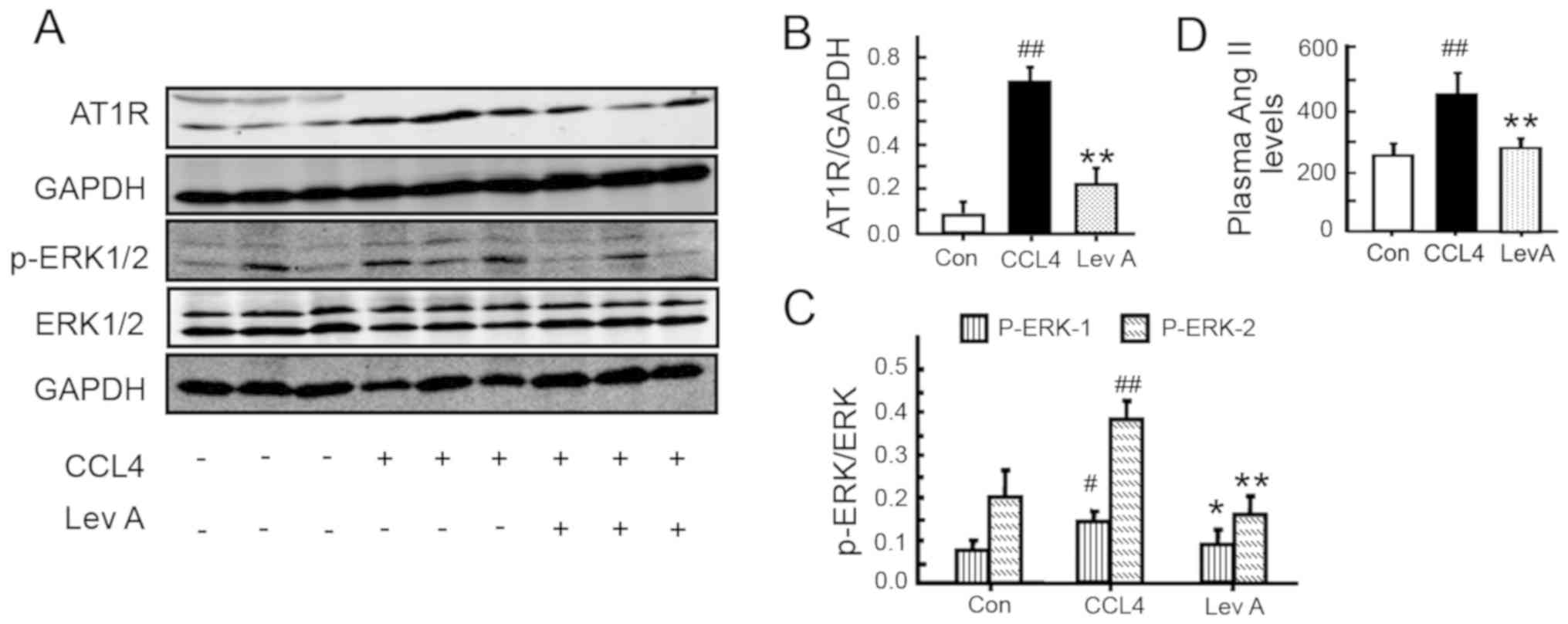 | Figure 6.Effect of Lev A on the molecules of
RAS in vivo. (A) The expression of AT1R, p-ERK1/2 and ERK1/2
in rat liver. (B and C) Relative levels of AT1R and p-ERK1/2,
respectively. (D) Plasma Ang II levels determined by
radioimmunoassay. Each bar represents the means ± standard
deviation (n=8). #P<0.05, ##P<0.01 vs.
control group; *P<0.05, **P<0.01 vs. model group. Lev A,
levistilide A; RAS, renin angiotensin system; AT1R, type 1
angiotensin receptor; p-, phosphorylated; Ang II, angiotensin II;
Con, control. |
Discussion
The data from the present study demonstrated that
Lev A was able to inhibit Ang II-induced proliferation and
activation of HSCs. Lev A also suppressed Ang II-induced
upregulation of AT1R and activation of ERK and c-Jun. In the
CCL4 rat model, treatment with Lev A ameliorated liver
fibrosis and improved liver function. In addition, Lev A also
inhibited CCL4-induced increase of Ang II and AT1R and
phosphorylation of ERK.
Ang II serves a vital role in the activation of HSCs
and secretion of ECM (22).
Therefore, numerous effective therapeutic approaches have focused
on the RAS, particularly Ang II (4,9). In
accordance with previous studies (6,23,24),
Ang II stimulation promoted the proliferation of HSCs and
upregulated the levels of α-SMA and F-actin in HSCs. Lev A
treatment inhibited Ang II-induced HSC proliferation and
activation, indicating that Lev A may be an effective agent in the
treatment of hepatic fibrosis. Subsequently, it was further
investigated whether Lev A could also inhibit hepatic fibrosis
in vivo. In the CCL4-induced rat hepatic fibrosis
model, Lev A significantly inhibited the expression of α-SMA and
collagen, and improved the rat liver function, exhibiting a good
therapeutic effect. These data suggested that Lev A is a good
candidate for the treatment of hepatic fibrosis.
It has been widely recognized that the profibrotic
activity of Ang II is mainly mediated by AT1R (25–27).
Consistent with previous studies (11–13),
the present study also revealed that the AT1R levels were
significantly elevated in Ang II-treated HSCs and
CCL4-treated rat liver. Lev A treatment inhibited Ang
II- and CCL4-induced AT1R upregulation in HSCs and in
the rat liver. Following binding to AT1R, Ang II also activates
intracellular signaling pathways, including MAPKs and transcription
factor AP-1 (12,28). Ang II induces the phosphorylation
of ERK1/2 and activation of AP-1, and subsequently increases the
expression of procollagen α1 and TGF-β1 in HSCs (12,25,29).
The results of the present study revealed that Lev A treatment
significantly suppressed Ang II-induced phosphorylation of ERK1/2
and c-Jun in HSCs. Lev A also inhibited CCL4-induced ERK
and c-Jun phosphorylation in rat livers. These results suggested
that Lev A may exert its anti-hepatic fibrosis effect by inhibiting
Ang II-AT1R signaling and consequently suppressing ERK/c-Jun
activation. In addition to the ACE-Ang II-AT1R pathway, the
ACE2-Ang 1–7-Mas axis also serves an important role in hepatic
fibrosis. ACE2 can degrade Ang II to Ang 1–7, an important,
biologically active component of this system, which exerts opposite
effects to those of Ang II through its receptor, Mas (30,31).
The ACE2-Ang 1–7-Mas axis counteracts the effects of the ACE-Ang
II-AT 1R axis in hepatic fibrosis (30,32).
Ang 1–7 has shown antifibrotic actions in the bile-duct-ligated rat
(33–35). Blockade of the Ang 1–7 receptor has
been demonstrated to aggravate liver fibrosis (36). It has been reported that losartan
may upregulate the cardiac ACE2-Ang(1–7)-Mas axis expression
(37). Therefore, the role of Lev
A on the ACE2-Ang(1–7)-Mas axis should be further investigated.
In summary, the present study demonstrated that Lev
A is able to inhibit Ang II-induced HSC activation and improve
experimental hepatic fibrosis induced by CCL4. Lev A
inhibits the upregulation of AT1R and activation of ERK1/2 and
c-Jun in vitro and in vivo. As a limitation of the
present study, the protein was extracted from liver tissues, not
from isolated HSC cells. Our previous study (21) demonstrated that swertiamarin is
able to attenuate experimental liver fibrosis by regulating the Ang
II/AT1R pathway. The results of the current study also indicated
that Lev A is a potential agent for the treatment of hepatic
fibrosis by suppressing the activation of Ang II/AT1R/ERK/c-Jun
signaling in HSCs. These results imply that the Ang II/AT1R pathway
is an important target for the screening of optimal agents for
reversing liver fibrosis from Chinese herbs.
Acknowledgements
Not applicable.
Funding
The present study was supported by grants from the
National Natural Science Foundation of China (grant nos. 81603458,
81730109 and 81473479), Special Program for Traditional Chinese
Medicine Research of Shanghai Municipal Commission of Health and
Family Planning (grant no. 2016JQ001), Baoshan Branch, Shuguang
Hospital Affiliated to Shanghai University of Traditional Chinese
Medicine (grant no. GZRPYZZ-201603), Construction plan of medical
specialty in Baoshan District of Shanghai (grant no.
BSZK-2018-A03), National Science and Technology Major Project
(grant no. 2014ZX10005001) and ‘Three-Year Action Plan’ for
Development of TCM in Shanghai (grant no. 16CR1026B).
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
ShuaL and CL were involved in conception and design
of the study. ShuL, WZ and ZZ performed the experiments. ShuL, BC
and ShuaL analyzed the data. ShuL and BC wrote the manuscript. All
authors read and approved the final manuscript.
Ethics approval and consent to
participate
The protocols were approved by the Committee on the
Ethics of Animal Experiments of Shanghai University of Traditional
Chinese Medicine, China. All animals received humane care during
the study.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Khomich O, Ivanov AV and Bartosch B:
Metabolic hallmarks of hepatic stellate cells in liver fibrosis.
Cells. 9(pii): E242019. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Huang Y, Deng X and Liang J: Modulation of
hepatic stellate cells and reversibility of hepatic fibrosis. Exp
Cell Res. 352:420–426. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Cai X, Wang J, Zhou Q, Yang B, He Q and
Weng Q: Intercellular crosstalk of hepatic stellate cells in liver
fibrosis: New insights into therapy. Pharmacol Res. 155:1047202020.
View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Sancho-Bru P and Ginès P: Targeting the
renin-angiotensin system in liver fibrosis. Hepatol Int.
10:730–732. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Li S, Zhao W, Tao Y and Liu C: Fugan Wan
alleviates hepatic fibrosis by inhibiting ACE/Ang II/AT-1R
signaling pathway and enhancing ACE2/Ang 1-7/Mas signaling pathway
in hepatic fibrosis rat models. Am J Transl Res. 12:592–601.
2020.PubMed/NCBI
|
|
6
|
Bataller R, Ginès P, Nicolás JM, Görbig
MN, Garcia-Ramallo E, Gasull X, Bosch J, Arroyo V and Rodés J:
Angiotensin II induces contraction and proliferation of human
hepatic stellate cells. Gastroenterology. 118:1149–1156. 2000.
View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Bataller R, Schwabe RF, Choi YH, Yang L,
Paik YH, Lindquist J, Qian T, Schoonhoven R, Hagedorn CH, Lemasters
JJ and Brenner DA: NADPH oxidase signal transduces angiotensin II
in hepatic stellate cells and is critical in hepatic fibrosis. J
Clin Invest. 112:1383–1394. 2003. View
Article : Google Scholar : PubMed/NCBI
|
|
8
|
Saber S, Goda R, El-Tanbouly GS and Ezzat
D: Lisinopril inhibits nuclear transcription factor kappa B and
augments sensitivity to silymarin in experimental liver fibrosis.
Int Immunopharmacol. 64:340–349. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Czechowska G, Celinski K, Korolczuk A,
Wojcicka G, Dudka J, Bojarska A, Madro A and Brzozowski T: The
effect of the angiotensin II receptor, type 1 receptor antagonists,
losartan and telmisartan, on thioacetamide-induced liver fibrosis
in rats. J Physiol Pharmacol. 67:575–586. 2016.PubMed/NCBI
|
|
10
|
Kim MY, Baik SK, Park DH, Jang YO, Suk KT,
Yea CJ, Lee IY, Kim JW, Kim HS, Kwon SO, et al: Angiotensin
receptor blockers are superior to angiotensin-converting enzyme
inhibitors in the suppression of hepatic fibrosis in a bile
duct-ligated rat model. J Gastroenterol. 43:889–896. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Reza HM, Tabassum N, Sagor MA, Chowdhury
MR, Rahman M, Jain P and Alam MA: Angiotensin-converting enzyme
inhibitor prevents oxidative stress, inflammation, and fibrosis in
carbon tetrachloride-treated rat liver. Toxicol Mech Methods.
26:46–53. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Bataller R, Gäbele E, Schoonhoven R,
Morris T, Lehnert M, Yang L, Brenner DA and Rippe RA: Prolonged
infusion of angiotensin II into normal rats induces stellate cell
activation and proinflammatory events in liver. Am J Physiol
Gastrointest Liver Physiol. 285:G642–G651. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Huang Y, Li Y, Lou A, Wang GZ, Hu Y, Zhang
Y, Huang W, Wang J, Li Y, Zhu X, et al: Alamandine attenuates
hepatic fibrosis by regulating autophagy induced by NOX4-dependent
ROS. Clin Sci (Lond). 134:853–869. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Wei H, Lu H, Li D, Zhan Y, Wang Z and
Huang X: The expression of AT1 receptor on hepatic stellate cells
in rat fibrosis induced by CCl4. Chin Med J (Engl). 114:583–587.
2001.PubMed/NCBI
|
|
15
|
Wang K, Cao P, Wang H, Tang Z, Wang N,
Wang J and Zhang Y: Chronic administration of Angelica
sinensis polysaccharide effectively improves fatty liver and
glucose homeostasis in high-fat diet-fed mice. Sci Rep.
6:262292016. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Pu X, Fan W, Yu S, Li Y, Ma X, Liu L, Ren
J and Zhang W: Polysaccharides from Angelica and Astragalus exert
hepatoprotective effects against carbon-tetrachloride-induced
intoxication in mice. Can J Physiol Pharmacol. 93:39–43. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Lee TF, Lin YL and Huang YT: Studies on
antiproliferative effects of phthalides from Ligusticum chuanxiong
in hepatic stellate cells. Planta Med. 73:527–534. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Friedman SL, Rockey DC, McGuire RF, Maher
JJ, Boyles JK and Yamasaki G: Isolated hepatic lipocytes and
Kupffer cells from normal human liver: Morphological and functional
characteristics in primary culture. Hepatology. 15:234–243. 1992.
View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Sohail MA, Hashmi AZ, Hakim W, Watanabe A,
Zipprich A, Groszmann RJ, Dranoff JA, Torok NJ and Mehal WZ:
Adenosine induces loss of actin stress fibers and inhibits
contraction in hepatic stellate cells via Rho inhibition.
Hepatology. 49:185–194. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Ala-Kokko L, Pihlajaniemi T, Myers JC,
Kivirikko KI and Savolainen ER: Gene expression of type I, III and
IV collagens in hepatic fibrosis induced by dimethylnitrosamine in
the rat. Biochem J. 244:75–79. 1987. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Li S, Wang Q, Tao Y and Liu C:
Swertiamarin attenuates experimental rat hepatic fibrosis by
suppressing angiotensin II-angiotensin type 1
receptor-extracellular signal-regulated kinase signaling. J
Pharmacol Exp Ther. 359:247–255. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Wei HS, Lu HM, Li DG, Zhan YT, Wang ZR,
Huang X, Cheng JL and Xu QF: The regulatory role of AT 1 receptor
on activated HSCs in hepatic fibrogenesis:Effects of RAS inhibitors
on hepatic fibrosis induced by CCl(4). World J Gastroenterol.
6:824–828. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Wu Y, Li Z, Wang S, Xiu A and Zhang C:
Carvedilol inhibits angiotensin II-induced proliferation and
contraction in hepatic stellate cells through the RhoA/Rho-kinase
pathway. Biomed Res Int. 2019:79320462019. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Zhang X, Zhang F, Kong D, Wu X, Lian N,
Chen L, Lu Y and Zheng S: Tetramethylpyrazine inhibits angiotensin
II-induced activation of hepatic stellate cells associated with
interference of platelet-derived growth factor β receptor pathways.
FEBS J. 281:2754–2768. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Li X, Meng Y, Wu P, Zhang Z and Yang X:
Angiotensin II and aldosterone stimulating NF-kappaB and AP-1
activation in hepatic fibrosis of rat. Regul Pept. 138:15–25. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Yang L, Bataller R, Dulyx J, Coffman TM,
Ginès P, Rippe RA and Brenner DA: Attenuated hepatic inflammation
and fibrosis in angiotensin type 1a receptor deficient mice. J
Hepatol. 43:317–323. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Wang XN, Hu YY, Liu CH, Liu P and Zhu DY:
Effects of salvianolic acid B on expressions of TGF-beta1 and its
receptors in liver of rats with dimethylnitrosamine-induced hepatic
fibrosis. Zhong Xi Yi Jie He Xue Bao. 3:286–289. 2005.(In Chinese).
View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Ruiz-Ortega M, Lorenzo O, Rupérez M,
Suzuki Y and Egido J: Angiotensin II activates nuclear
transcription factor-kappaB in aorta of normal rats and in vascular
smooth muscle cells of AT1 knockout mice. Nephrol Dial Transplant.
16 (Suppl 1):S27–S33. 2001. View Article : Google Scholar
|
|
29
|
Vogt PK: Jun, the oncoprotein. Oncogene.
20:2365–2377. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Donoghue M, Hsieh F, Baronas E, Godbout K,
Gosselin M, Stagliano N, Donovan M, Woolf B, Robison K, Jeyaseelan
R, et al: A novel angiotensin-converting enzyme-related
carboxypeptidase (ACE2) converts angiotensin I to angiotensin 1–9.
Circ Res. 87:E1–E9. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Tipnis SR, Hooper NM, Hyde R, Karran E,
Christie G and Turner AJ: A human homolog of angiotensin-converting
enzyme. Cloning and functional expression as a captopril-
insensitive carboxypeptidase. J Biol Chem. 275:33238–33243. 2000.
View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Santos RA, Ferreira AJ, Verano-Braga T and
Bader M: Angiotensin-converting enzyme 2, angiotensin-(1–7) and
Mas: New players of the renin-angiotensin system. J Endocrinol.
216:R1–R17. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Lubel JS, Herath CB, Tchongue J, Grace J,
Jia Z, Spencer K, Casley D, Crowley P, Sievert W, Burrell LM and
Angus PW: Angiotensin-(1–7), an alternative metabolite of the
renin-angiotensin system, is up-regulated in human liver disease
and has antifibrotic activity in the bile-duct-ligated rat. Clin
Sci (Lond). 117:375–386. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Dasuri K, Zhang L and Keller JN: Oxidative
stress, neurodegeneration, and the balance of protein degradation
and protein synthesis. Free Radic Biol Med. 62:170–185. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Herath CB, Warner FJ, Lubel JS, Dean RG,
Jia Z, Lew RA, Smith AI, Burrell LM and Angus PW: Upregulation of
hepatic angiotensin-converting enzyme 2 (ACE2) and
angiotensin-(1–7) levels in experimental biliary fibrosis. J
Hepatol. 47:387–395. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Pereira RM, Dos Santos RA, Teixeira MM,
Leite VH, Costa LP, da Costa Dias FL, Barcelos LS, Collares GB and
Simões e Silva AC: The renin-angiotensin system in a rat model of
hepatic fibrosis: Evidence for a protective role of
Angiotensin-(1-7). J Hepatol. 46:674–681. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Wang X, Ye Y, Gong H, Wu J, Yuan J, Wang
S, Yin P, Ding Z, Kang L, Jiang Q, et al: The effects of different
angiotensin II type 1 receptor blockers on the regulation of the
ACE-AngII-AT1 and ACE2-Ang(1–7)-Mas axes in pressure
overload-induced cardiac remodeling in male mice. J Mol Cell
Cardiol. 97:180–190. 2016. View Article : Google Scholar : PubMed/NCBI
|