Introduction
Long noncoding RNAs (lncRNAs), which are a major
class of transcripts >200 nucleotides in length, drive several
important physiological processes, including proliferation,
metastasis and apoptosis of tumor cells (1,2).
Numerous studies have revealed that lncRNAs have critical functions
in the regulation of malignant tumor behavior, including in liver
(3), pancreatic (4) and breast cancer (5). Growth arrest-specific 5
(GAS5), located at chromosome 1q25.1, is an antitumor lncRNA
that is frequently downregulated in various cancer types and
associated with clinicopathological characteristics, including
tumour size, staging and invasion (6). In bladder cancer cells, GAS5
was revealed to function as a transcriptional factor promoting the
transcriptional activity of enhancer of zeste homolog 2 by
physically interacting with E2F transcription factor 4, thus
inducing apoptosis (7). In gastric
cancer cells, GAS5 blocked cell cycle phases by interacting
with Y-box binding protein 1 and thus upregulating p21 expression,
which has a demonstrable role in inhibiting tumor growth (8).
Autophagy is a regular physiological process of
metabolic degradation, and the process occurs in double-membrane
autophagosomes (9). Autophagy
serves a critical role in numerous biochemical processes, including
pro-survival signaling and pro-apoptotic signaling (10). In several cancer cell types, the
modulation of autophagy serves a tumor-promoting or
tumor-inhibiting role depending on the different modulators and the
specific autophagy pathway (11).
During cellular stress, including hypoxia and nutrient deprivation,
these factors function as tumor-promoting modulators of autophagy
by helping cells overcome these stresses (12,13).
For example, in RAS-mutated cancer cells, the basal level of
autophagy was revealed to be markedly high to compensate for the
loss of RAS-modulated signaling pathways, including pathways
related to proliferation, survival and metabolism (14–16).
In addition, the basal level of autophagy has also been revealed to
serve as a regulatory mechanism for tumor suppression by inducing
cellular damage and imbalanced cellular homeostasis (17). Studies have revealed that in
several cancer types, including cervical squamous-cell carcinoma
(18) and hepatocellular carcinoma
(19), Beclin1, which is an
important protein for autophagosome formation, is upregulated and
thus acts as a tumor suppressor.
Unc-51 like autophagy activating kinase (ULK)1 plays
a key role in the regulation of autophagy initiation in an
autophagy-related (Atg)5- and Atg7-independent macroautophagic
manner (20). The human genome
contains two ULK1 homologs, ULK1 and ULK2, which are
known to initiate autophagy (21,22).
Accumulating evidence has demonstrated that ULK1 tightly regulates
tumor progression by modulating autophagy in cancer cells. For
example, Blessing et al (23) reported that increased autophagy
upregulated ULK1 expression, further promoting autophagy in
prostate cancer. In glioma, ULK1 and ULK2 were
revealed to be methylated and silenced; subsequently, ULK1 and/or
ULK2 failed to induce autophagy, which resulted in promotion of
proliferation and tumor progression, indicating that downregulation
of ULK1/2 is essential for glioma development (24). These results indicated that ULK1/2
may tightly regulate tumor progression by initiating autophagy.
In the present study, the expression levels of GAS5,
ULK1 and ULK2 were measured in breast cancer (BC) clinical samples
and adjacent samples, also aiming to analyze any correlation
between these proteins. Additionally, the present study
investigated the possible molecular mechanism of GAS5 involved in
the regulation of autophagy in breast cancer cells. The results
should increase understanding of the relationship between GAS5 and
autophagy in BC cells, and provide new insights into the impact of
GAS5 on the chemoresistance to cisplatin, which may facilitate the
development of more effective clinical treatments for BC.
Materials and methods
Clinical samples
Paired human breast cancer clinical samples and
adjacent normal samples used in the present study were obtained
from 39 female patients, aged from 30 to 82 (median age, 52.9) at
the Affiliated Hospital of Southwest Medical University (Luzhou,
China) between December 2018 and March 2019. The present study was
approved by The Medical Ethics Committee of the Institutional
Review Board of the Affiliated Hospital of Southwest Medical
University (Luzhou, China). Written informed consent was provided
by all patients. All types of breast cancer were invasive ductal
carcinoma as confirmed by pathologists. Inclusion criteria were
histological diagnosis of invasive ductal carcinoma of the breast
before operation. None of the patients received chemotherapy or
radiotherapy prior to surgical excision Detailed
clinicopathological information were obtained, including sex, age,
tumor depth, tumor size, differentiation, lymph node invasion and
distant metastasis (data not shown).
Cell cultures
Three human cell lines, including breast cancer cell
lines MCF-7 and MDA-MB-231, and breast non-tumor cell line MCF-10a,
were used in the present study. All cell lines were frozen in
liquid nitrogen and cultured in MEM supplemented with 10% fetal
bovine serum (FBS, Gibco; Thermo Fisher Scientific, Inc.), 100 µ/ml
penicillin and 100 µ/ml streptomycin (Amresco, LLC) in a 5%
CO2 incubator at 37°C. Cells were passaged once every
three days. To inhibit autophagy, 2 mM 3-methyladenine (3-MA,
Sigma-Aldrich; Merck KGaA) was added into culturing medium for 24
h.
Reverse transcription-quantitative
(RT-q)PCR
Total RNA was extracted from BC clinical samples and
adjacent normal samples using TRIzol® (Thermo Fisher
Scientific, Inc). A total of 5 mg of each sample was suspended in 1
ml RNA EasyOut kit (Chengdu Daosheng Biological Technology Co.,
Ltd.) and sonicated at room temperature for 5 sec using a
SoniConvert™ system (Chengdu Daosheng Biological Technology Co.,
Ltd.), both according to the manufacturer's instructions. For
RT-qPCR analysis, 1 µg total RNA was reverse-transcribed into cDNA
using an Superscript III RT-qPCR kit (Thermo Fisher Scientific,
Inc.). Then the cDNA was used as template under the following
conditions: 40 cycles at 95°C for 30 sec and 60°C for 1 min. An ABI
7500 Fast qPCR system (Applied Biosystems; Thermo Fisher
Scientific, Inc.) was used for qPCR. For amplification, the
following procedure was performed: Predenaturation at 98°C for 2
min for each cycle, denaturation at 98°C for 10 sec, annealing and
extension at 60°C for 60 sec, 40 cycles; final extension at 60°C
for 5 min. The primers used for qPCR were as follows: GAS5 forward,
5′-GAGCAAGCCTAACTCAAGCC-3′ and reverse, 5′-ACACAGTGTAGTCAAGCCGA-3′;
ULK1 forward, 5′-GGCAAGTTCGAGTTCTCCCG-3′ and reverse,
5′-CGACCTCCAAATCGTGCTTCT-3′; ULK2 forward,
5′-TGGAGACCTCGCAGATTATTTGC-3′ and reverse,
5′-CTGTGCAGGATTCGCATGG-3′; β-actin forward,
5′-CATGTACGTTGCTATCCAGGC-3′ and reverse,
5′-CTCCTTAATGTCACGCACGAT-3′. The qPCR results were analyzed and
expressed relative to the Cq (threshold cycle) values (25) and then converted to fold change
values. A fold-change of 2.0 was considered significant.
Western blotting
Cells (1×106) were suspended in 1 ml of
RIPA buffer (Sigma-Aldrich; Merck KGaA) and lysed using
SoniConvert® Sonicator (Chengdu Daosheng Biological
Technology Co., Ltd.). Protein concentration was determined via BCA
assay (Sigma-Aldrich; Merck KGaA) according to the manufacturer's
instructions. In each lane, 20 µg total protein was applied to a
5–15% SDS-PAGE gel. Fractionated proteins were transferred onto a
nitrocellulose membrane and blocked using 5% BSA containing PBS at
room temperature for 1 h. Membranes were then incubated with
primary antibodies overnight at 4°C. The primary antibodies used
were obtained from Abcam and are listed as follows: Rabbit
monoclonal anti-ULK1 antibody (1:2,000; cat. no. ab133766), rabbit
polyclonal anti-ULK2 antibody (1:3,000; cat. no. ab97695), rabbit
monoclonal anti-Atg3 antibody (1:500; cat. no. ab108251), rabbit
polyclonal anti-Atg5 antibody (1:2,000; cat. no. ab228668), rabbit
monoclonal anti-Atg7 antibody (1:2,000; cat. no. ab52472), rabbit
monoclonal anti-Beclin 1 antibody (1:2,000; cat. no. ab207612),
rabbit polyclonal anti-LC3B antibody (1:500; cat. no. ab48394) and
mouse monoclonal anti-β-actin antibody (1:5,000; cat. no. ab8226).
Following three washes with TBST, secondary antibody was incubated
with the membrane at room temperature for 1 h. Goat anti-rabbit IgG
H&L antibody (HRP-labelled; 1:10,000; cat. no. ab7090) and goat
anti-mouse IgG H&L antibody (HRP-labelled; 1:10,000; cat. no.
ab97040) were used as the secondary antibodies. Blot bands were
quantified via densitometry using ImageJ software (v2.0; National
Institutes of Health). β-actin was used as an internal
reference.
Tandem mRFP/mCherry-GFP fluorescence
microscopy
MCF-7 cells were seeded at 2×105
cells/well in a 12-well plate and attached overnight at 37°C in a
5% CO2 incubator. Cells were transfected using pre-made
lentiviral particles expressing a fusion target of GFP-RFP-LC3
(cat. no. GM-1314L204H-S, Genomeditech Co., Ltd.) according to the
manufacturer's instructions. After 2 h incubation at 37°C in the
dark, the medium was refreshed and cells were incubated in MEM
supplemented with 10% FBS at 37°C for 48 h. Cells were visualized
under an X71 (U-RFL-T) fluorescence microscope (magnification,
×100; Olympus Corporation) and analyzed with ImageJ software (v2.0;
National Institutes of Health).
Double staining of lysotracker green
and mitotracker sox red
MCF-7 cells were seeded at 2×105
cells/well in a 12-well plate and incubated with Lysotracker green
and mitotracker sox red according to the manufacturer's
instructions. After 1 h incubation at 37°C, 1 µg/ml
4′,6-diamidino-2-phenylindole (DAPI) was used to stain nuclei for 5
min at room temperature in the dark. Cellular fluorescence was
observed using an X71 (U-RFL-T) fluorescence microscope
(magnification, ×100; Olympus Corporation).
Cell Counting Kit (CCK)-8 assay for
proliferation
A total of 5×103 cells/well were seeded
in 96-well plates and allowed to attach overnight. After addition
of 1, 2, 3, 4 and 5 µmol/L of cisplatin (Thermo Fisher Scientific,
Inc.), for 1–5 days, the cell proliferation assay was performed by
adding 10 µl CCK-8 solution (Guangzhou RiboBio Co., Ltd.) according
to the manufacturer's protocol. After 1 h incubation at 37°C, the
absorbance of each well was measured at a wavelength of 450–620 nm
using a Multiskan spectrum microplate reader (Thermo Fisher
Scientific, Inc.).
Cell cycle analysis
A total of 1×106 MCF-7 cells were
collected and washed with ice-cold PBS twice, and fixed using 1 ml
70% ethanol and stored at 4°C overnight. After washing with
ice-cold PBS three times, the staining solution containing 100
ng/ml RNase A and 5 µg/ml propidium iodide (PI) (Sigma-Aldrich;
Merck KGaA) was added to cells and incubated in the dark for 30 min
at 4°C. Cells were the screened by a three laser Navios flow
cytometer (Beckman Coulter, Inc.) and analyzed by ModFit LT
software (v2.0, Verity Software House).
Annexin V-FITC/PI double staining and
flow cytometric analysis
MCF-7 Cells were treated with 2 µM cisplatin for 24
h, and untreated cells were considered as control group. Cells were
resuspended in trypsin and washed twice with ice-cold PBS. Then,
Annexin V-FITC/PI double staining was performed according to the
instructions of the Annexin V-FITC/PI apoptosis detection kit
(Thermo Fisher Scientific, Inc.). For each sample,
~2×104 cells were collected and scanned by a three laser
Navios flow cytometer (Beckman Coulter, Inc.). Data files was
analyzed using FlowJo v9.7.6 (FlowJo LLC).
Transwell cell invasion assay
Transwell membranes were precoated with 60 µl
Matrigel (8%) in MEM and incubated at room temperature for 2 h. For
each well, 1×104 cells were added into upper chamber.
Subsequently, 600 µl of MEM supplemented with 10% FBS was added to
the lower chamber. After 24 h, medium was removed and 4%
paraformaldehyde was used for fixation at room temperature for 10
min. After washing with PBS twice, cells were stained with 0.1%
crystal violet (in PBS) at room temperature for 30 min. After
staining, the crystal violet solution was removed and the cells
were washed with PBS five times. Then the cells in five random
views were counted under an X71 fluorescence microscope
(magnification, ×100; Olympus Corporation).
Transfection
A total of 0.8 µg GAS5-expressing plasmid or empty
vector (Addgene, Inc.) was mixed with 4 µl Lipofectamine™ 2000
transfection reagent (Thermo Fisher Scientific, Inc.) in 0.5 ml
OptiMEM medium (Thermo Fisher Scientific, Inc.), and this was added
to MCF-7 cells for 4 h followed by replacing the medium. After 24
h, the culture media was refreshed containing 500 µg/ml geneticin
sulfate 418 (G418, Sigma-Aldrich; Merck KGaA) for antibiotic
selection. Two weeks later, the transfected cells that had survived
were collected and maintained in 4,000 µg/ml G418. Individual
clones (GAS5 clone-1 and clone-2) were detected, separately.
Tumor formation in soft agar
Low-melting agarose (2 ml 0.6%, Sigma-Aldrich; Merck
KGaA) diluted in MEM supplemented with 10% FBS was added to 6-well
plates. For each well, a total of 5×103 cells were added
in 1 ml of 0.3% (weight/volume) low-melting agarose diluted in MEM
supplemented with 10% FBS. After 2 weeks of incubation at 37°C,
cells were stained with 10 ng/ml nitro blue tetrazolium at 37°C and
colony number and diameter were quantified under an X71
fluorescence microscope (magnification, ×100; Olympus
Corporation).
Statistical analysis
Statistical analyses were carried out using SPSS
v19.0 for Windows (IMB Corp). Student's paired t-tests and ANOVA
were used to compare two or more groups for statistical analysis
followed by Tukey's post hoc test, respectively. Pearson's
correlation analysis was used for the correlation between GAS5,
ULK1 and ULK2 levels. Data are presented as the mean ± standard
deviation. P<0.05 was considered to indicate a statistically
significant difference. All experiments were repeated at least
three times.
Results
GAS5 is downregulated in BC clinical
samples and BC cell lines
The present study detected the expression levels of
GAS5 in paired BC clinical samples and adjacent normal
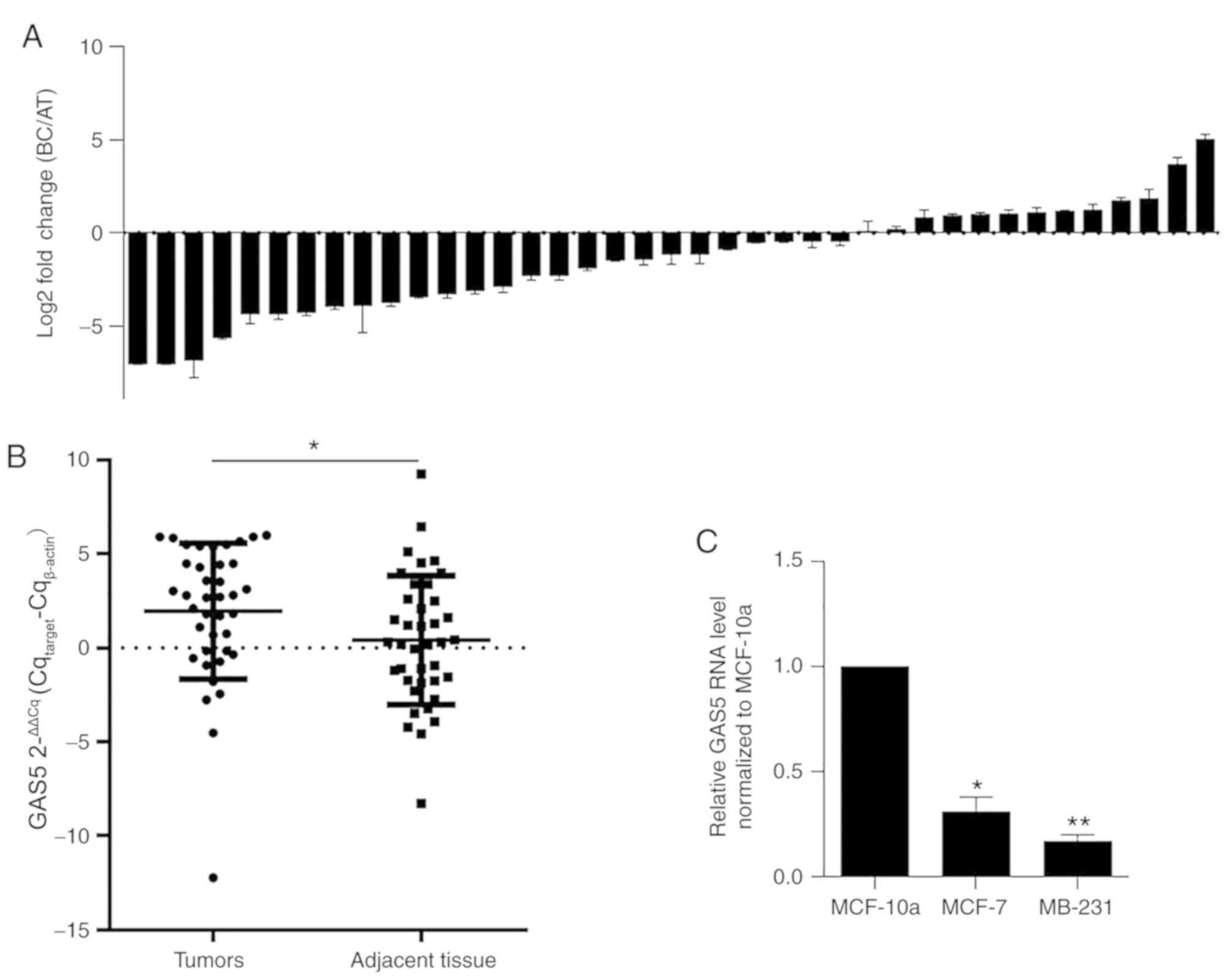
tissues by performing RT-qPCR. As revealed in Fig. 1A, among the 39 cases of BC, ~66.7%
exhibited lower levels of GAS5 compared with matched
controls, and GAS5 was significantly downregulated in BC
tissue samples compared with those in adjacent normal tissues
(1.95±0.36 vs. 0.40±0.22, P=0.002, Fig. 1B). To confirm whether similar
expression patterns of GAS5 were presented in BC cell lines,
GAS5 expression levels were analyzed in MCF-7 and MDA-MB-231 cells
compared with the non-tumor cell line MCF-10a. As anticipated, the
expression levels of GAS5 in both MCF-7 and MDA-MB-231 was
significantly lower compared with those in MCF-10a cells (*P=0.037;
**P 0.006, Fig. 1C), indicating
that GAS5 may exert antitumor effects in BC.
Expression levels of GAS5, ULK1 and
ULK2 are significantly correlated in BC clinical samples
The present study also assessed the expression
levels of ULK1 and ULK2 in BC clinical samples and adjacent
samples. Results revealed that GAS5 expression in BC clinical
samples was positively and significantly correlated with ULK1 and
ULK2 levels (Fig. 2A). To confirm
whether the expression levels of GAS5 positively regulated ULK1,
ULK2 and other autophagy-associated proteins, including
AuTophaGy-related (Atg)3, Atg5, Atg7, Beclin-1 and
microtubule-associated proteins 1A/1B light chain (LC)3B, cells
stably overexpressing GAS5 were selected and analyzed for the
expression levels of the aforementioned proteins. As revealed in
Fig. 2B, ULK1 and ULK2 protein
levels were both significantly upregulated in GAS5 clone-1 and
clone-2 (*P<0.05), which was consistent with the tendency
observed in clinical samples. In addition, GAS5 overexpression
induced an increase in Beclin-1 and LC3B levels with no effect on
Atg3, Atg5 and Atg7, indicating that GAS5 potentially initiates
autophagy by upregulating ULK1 and ULK2, but not Atgs (data not
shown).
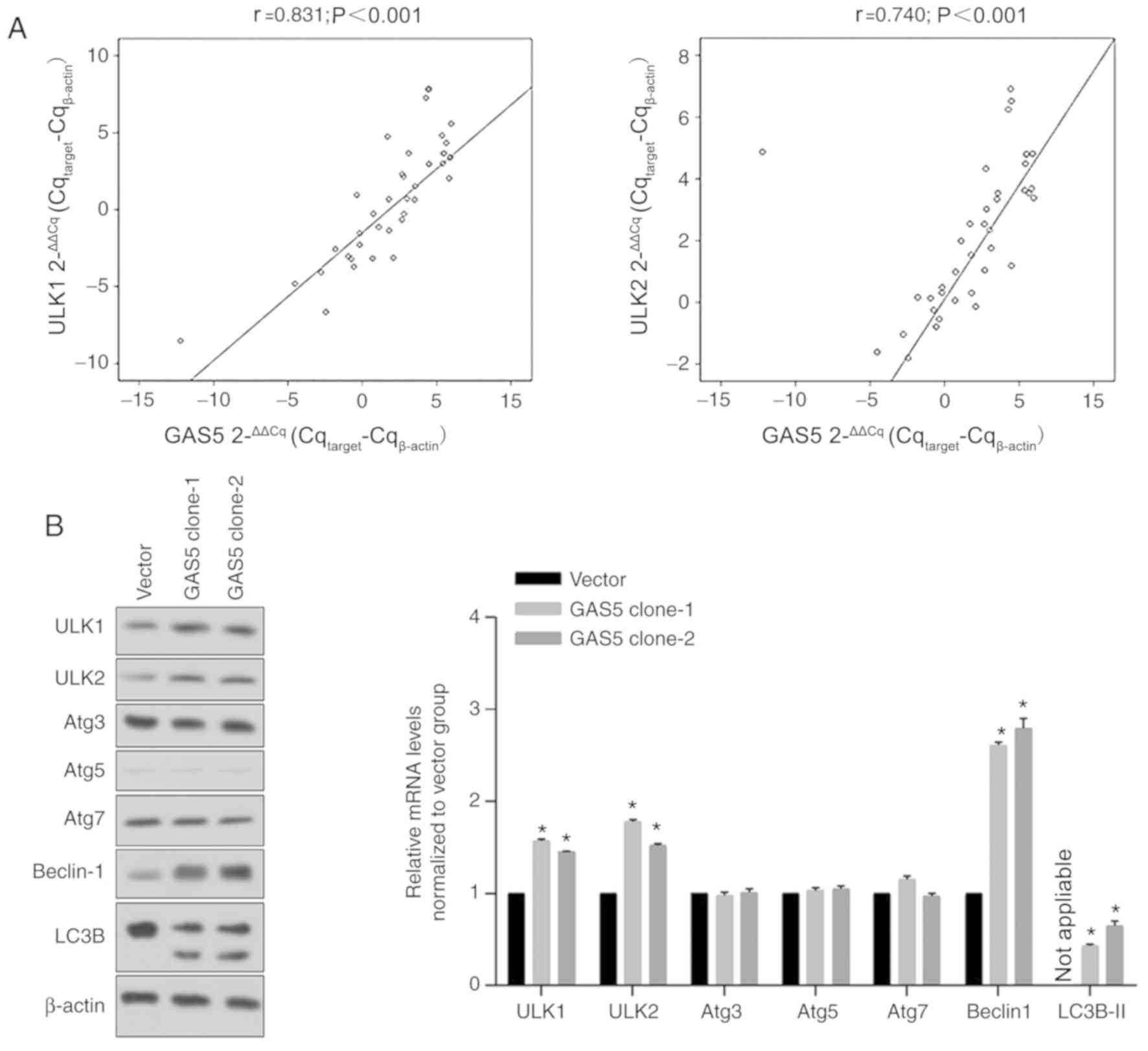 | Figure 2.Correlation between GAS5, ULK1 and
ULK2 levels. (A) Interactions between GAS5, ULK1 and ULK2 levels
were assessed using Pearson's correlation analysis. (B) After
stable expression of GAS5 in MCF-7 cells, the autophagic proteins,
including ULK1, ULK2, Atg3, Atg5, Atg7, Beclin-1 and LC3B, were
analyzed using western blotting (left panel). The bands were
analyzed and normalized to the Vector group. *P<0.05, vs. the
Vector group. GAS5, growth arrest-specific 5; ULK, unc-51-like
autophagy activating kinase; Atg, autophagy-related; LC3B,
microtubule-associated proteins 1A/1B light chain 3B. |
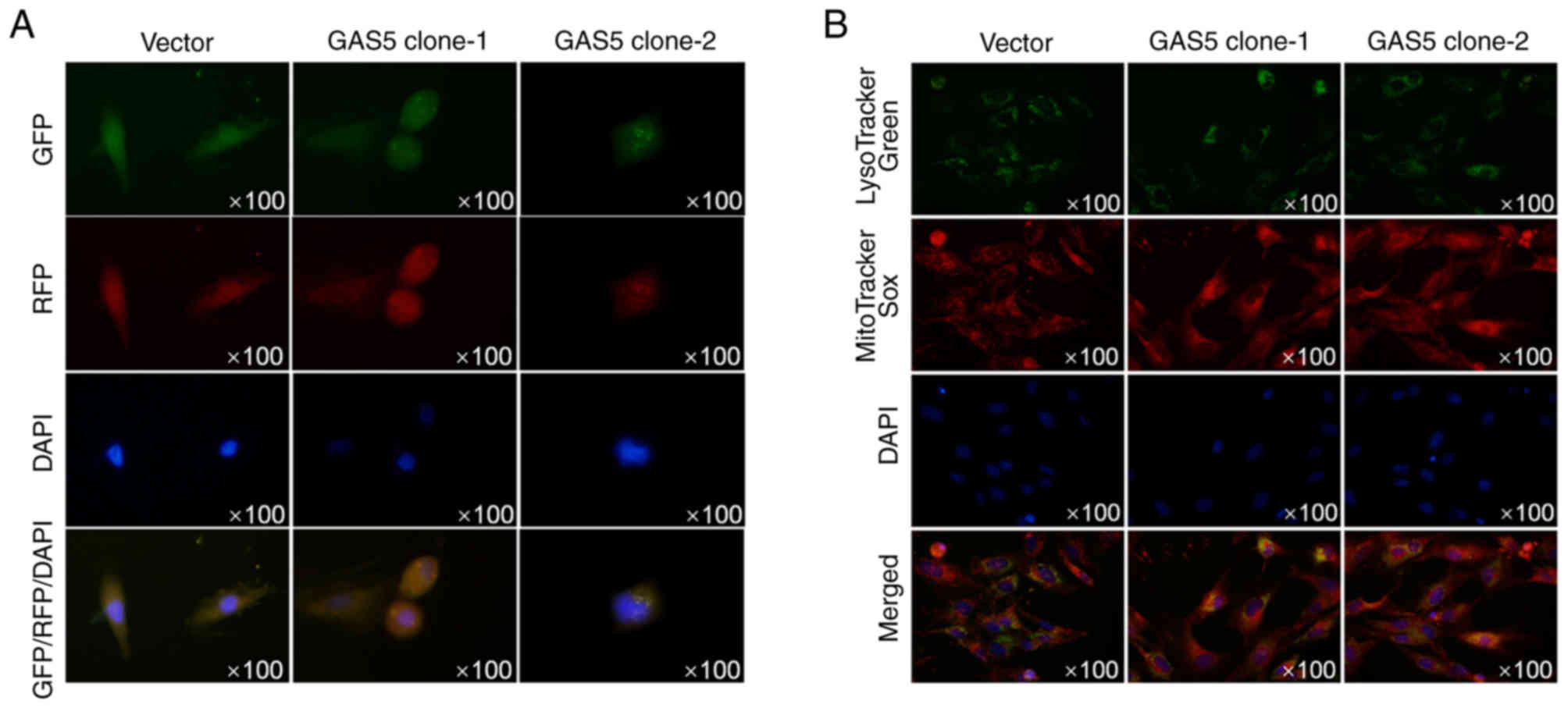
To further confirm whether autophagosome formation
was promoted by overexpressed GAS5, images of MCF-7 cells stably
overexpressing GAS5 and transiently expressed LC3-RFP-GFP were
captured using confocal microscopy to view the autophagosomes. As
revealed in Fig. 3A, without
autophagy promoter treatment, overexpression of GAS5 promoted the
formation of autophagosomes. Double staining of lysotracker green
and mitotracker sox red also demonstrated that overexpression of
GAS5 promoted colocalization of mitochondria and lysosomes,
indicating the promoting effect of GAS5 on autophagy/mitophagy
(Fig. 3B).
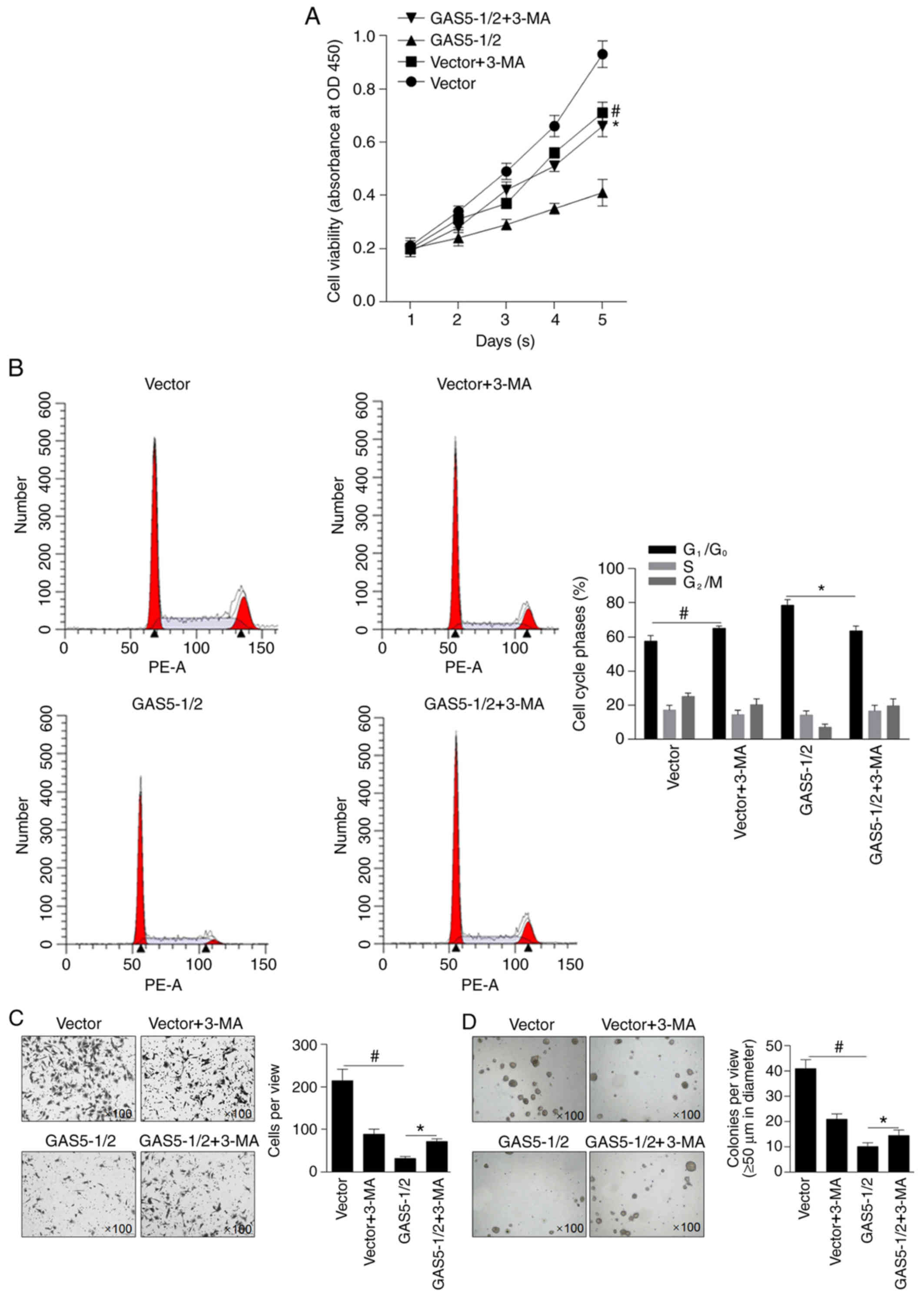
GAS5 exerts antitumor effects
partially by inducing autophagy in MCF-7 cells
To evaluate the effects of overexpressed GAS5 on the
malignant behaviors of MCF-7 cells, cell viability was analyzed. As
revealed in Fig. 4A, mixed clone-1
and clone-2 (GAS5-1/2) inhibited cell viability compared with the
vector-transfected clone (P<0.05). Pre-treatment of 10 µM 3-MA
was performed to demonstrate whether autophagy induced by GAS5
affected cell viability. 3-MA treatment notably decreased cell
viability in the vector group, indicating the inhibitory effect of
3-MA on proliferation alone, and this inhibitory effect on cell
viability was reversed by GAS5 overexpression (Fig. 4A). By performing cell cycle
analysis, it was observed that GAS5-induced block of the cells at
G1/G0 was reversed by 3-MA treatment. The
effects of GAS5 on invasion and colony formation in MCF-7 cells was
then investigated, and results demonstrated that invasion (Fig. 4C) and colony formation (Fig. 4D) of MCF-7 cells were significantly
inhibited by GAS5 overexpression compared with vector group and
partially reversed by inhibiting autophagy compared with GAS5-1/2
group, indicating that GAS5 may exert antitumor effects by
partially inducing autophagy in MCF-7 cells.
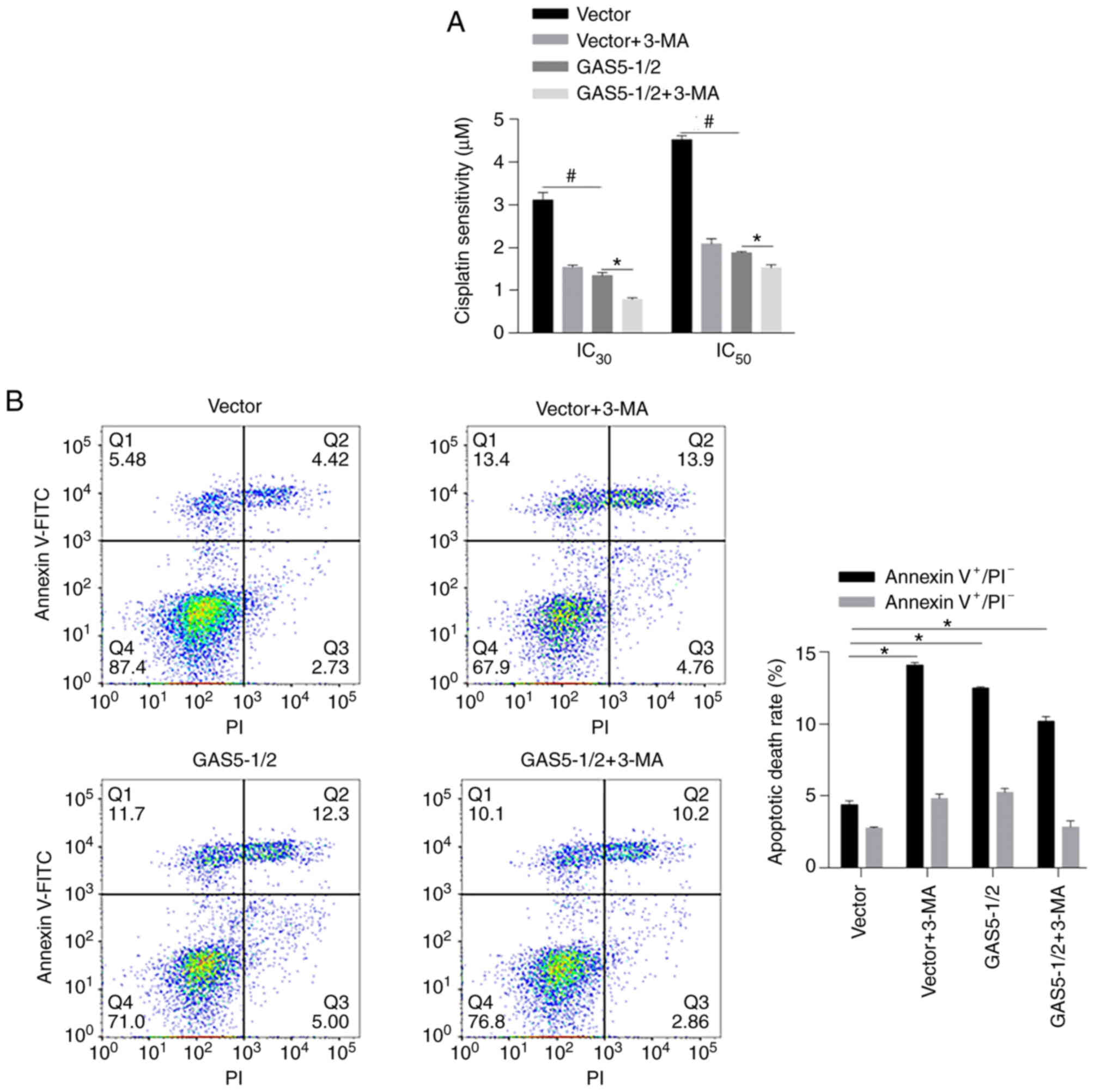
GAS5 sensitizes MCF-7 cells to
chemotreatment in an autophagy-independent manner
Since autophagy is a critical physiological process
for chemosensitivity in cancer cells, it was then evaluated whether
GAS5 affects chemosensitivity by regulating autophagy in MCF-7
cells. By performing chemosensitivity on cisplatin, as anticipated,
both 3-MA treatment and overexpression of GAS5 desensitized MCF-7
cells to cisplatin (Fig. 5A,
*P<0.05, vs. GAS5-1/2 group; #P<0.05, vs. Vector
group). However, 3-MA treatment decreased chemosensitivity in MCF-7
cells overexpressing GAS5 instead of reversing the promoting effect
of GAS5 on chemosensitivity. To analyze the apoptotic rate induced
by cisplatin, Annexin V-FITC/PI double staining was performed after
24-h treatment with 2 µM cisplatin. Autophagy inhibition
significantly increased apoptotic cell death compared with mock
group, and GAS5-promoted apoptosis was not notably affected by
autophagy inhibition, indicating that the effect of GAS5 on
chemosensitivity may be autophagy-independent (Fig. 5B).
Discussion
The present study is the first to demonstrate the
positive correlation between GAS5 and ULK1/2 levels in BC
pathogenesis, to the best of the authors' knowledge. GAS5 is not
only expressed in multiple cancers including breast cancer, but
also widely expressed in a variety of normal tissues, such as lung,
esophagus, colon, ovary and kidney (26). GAS5 is involved in the regulation
of physiological processes; for example, GAS5 transcriptionally
regulates mTOR and thus activates downstream signaling, which
promotes cell proliferation by promoting entry into the
G2/M phase of the cell cycle (27). GAS5 is also well known as a tumor
suppressor in various cancer types, including gastric (8), lung (2), pancreatic (14) and BC (7). In these cancer types, GAS5 was
revealed to be transcriptionally downregulated, and GAS5
upregulation inhibited malignant behaviors, including
proliferation, migration, invasion and tumor formation. Given these
reports, several studies investigating GAS5 have aimed to increase
cellular GAS5 levels via specific methods to control the
proliferation of cancer cells (26,7,14).
In the present study, it was reported that the expression levels of
GAS5 were significantly downregulated in BC clinical samples
compared to those in adjacent normal samples, and the expression of
GAS5 was positively correlated with the expression levels of ULK1
and ULK2, which are key initiators of autophagy (20–22).
Further experiments also revealed that GAS5 exerted suppressive
effects on the malignant behaviors of BC cells, including
proliferation, invasion and tumor formation, partially by inducing
autophagy. However, one limitation of the present study is that no
triple-negative breast cancer tissues were collected; thus, the
functions of GAS5 in these tissues is still unknown and further
studies should focus on the effect of GAS5 in triple-negative
breast cancer.
lncRNAs are also involved in autophagy regulation,
including autophagy initiation (28), phagophore nucleation (29), autophagosome elongation/closure
(30) and autolysosome fusion
(31). Several types of lncRNAs
are involved in these processes, including GAS5 (32). Song et al (32), reported that in osteoarthritis,
GAS5 was overexpressed and thus stimulated apoptosis and suppressed
autophagic responses, which was determined by the detection of
downregulated Beclin 1, Atg7, Atg12 and LC3B. In the present study,
GAS5 was positively correlated with the expression levels of ULK1/2
in BC clinical samples, but no correlation was observed between
GAS5 and with Atgs (data not shown). It was also demonstrated that
overexpression of GAS5 in MCF-7 cells caused upregulation of ULK1
and ULK2, but it was not determined whether ULK1/2 levels were
regulated directly or indirectly. Moreover, GAS5 overexpression did
not affect Atg3, Atg5 or Atg7 levels. This may be due to the
different expression patterns of microRNA-21, which is suppressed
by GAS5 and regulates Atg expression (32). As a result of overexpressed GAS5,
autophagy was promoted via increased levels of LC3B and Beclin-1
and the formation of autophagosomes. It is still unclear how GAS5
affects the expression of ULK1 and ULK2; however, the present study
demonstrated the promoting effect of GAS5 on autophagy, and GAS5
exerted antitumor effects by promoting autophagy, at least in
part.
lncRNA-induced autophagy is involved in modulating
chemosensitivity. Hu et al (33), reported that metastasis-associated
lung adenocarcinoma transcript 1, which is widely involved in the
regulation of biological and cellular processes, including
autophagy, induced chemoresistance by modulating autophagy. In
bladder cancer cells, the lncRNA gallbladder cancer drug
resistance-associated lncRNA1, which is upregulated in bladder
cancer cells, promoted autophagic flux and thus induced
chemoresistance (34). In the
present study, it was reported that both overexpression of GAS5 and
autophagy inhibition by 3-MA pretreatment desensitized MCF-7 cells
to cisplatin, and that 3-MA pretreatment failed to disturb
GAS5-induced chemoresistance, indicating that GAS5 may induce
chemoresistance in an autophagy-independent manner. The results
demonstrated that GAS5 may not be a promising indicator for
predicting chemosensitivity in BC.
Collectively, the present study may provide a
possible mechanism by which GAS5 regulates breast cancer by
promoting autophagy and affecting proliferation, invasion and tumor
formation.
Acknowledgements
Not applicable.
Funding
The present study was funded by Initiation Fund for
Doctors of the Affiliated Hospital of Southwest Medical University
(grant no. 18110) and The Undergraduate Training Program for
Innovation and Entrepreneurship of the Southwest Medical University
(grant no. 2019272)
Availability of data and materials
The datasets used and/or analyzed during the present
study are available from the corresponding author on reasonable
request.
Authors' contributions
GL and LQ performed cellular and molecular
experiments. XT and YC contributed to molecular experiments and
were responsible for data collection. ZZ performed cellular
experiments, analyzed and interpreted the data. CZ conceived and
designed the study. ZZ and CZ performed molecular experiments and
wrote the manuscript. All authors read and approved the final
manuscript.
Ethics approval and consent to
participate
The present study was approved by The Medical Ethics
Committee of The Institutional Review Board of the Affiliated
Hospital of Southwest Medical University (Luzhou, China; approval
no. KY2019040). Written informed consent was provided by all
patients.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
D'Angelo E and Agostini M: Long non-coding
RNA and extracellular matrix: The hidden players in cancer-stroma
cross-talk. Noncoding RNA Res. 3:174–177. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Lu T, Wang Y, Chen D, Liu J and Jiao W:
Potential clinical application of lncRNAs in non-small cell lung
cancer. OncoTargets Ther. 11:8045–8052. 2018. View Article : Google Scholar
|
|
3
|
Yang Y, Chen L, Gu J, Zhang H, Yuan J,
Lian Q, Lv G, Wang S, Wu Y, Yang YT, et al: Recurrently deregulated
lncRNAs in hepatocellular carcinoma. Nat Commun. 8:144212017.
View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Tang YT, Xu XH, Yang XD, Hao J, Cao H, Zhu
W, Zhang SY and Cao JP: Role of non-coding RNAs in pancreatic
cancer: The bane of the microworld. World J Gastroenterol.
20:9405–9417. 2014.PubMed/NCBI
|
|
5
|
Zhao Z, Chen C, Liu Y and Wu C:
17β-Estradiol treatment inhibits breast cell proliferation,
migration and invasion by decreasing MALAT-1 RNA level. Biochem
Biophys Res Commun. 445:388–393. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Pickard MR and Williams GT: Molecular and
Cellular Mechanisms of action of tumour suppressor GAS5 LncRNA.
genes (Basel). 6:484–499. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Wang M, Guo C, Wang L, Luo G, Huang C, Li
Y, Liu D, Zeng F, Jiang G and Xiao X: Long noncoding RNA GAS5
promotes bladder cancer cells apoptosis through inhibiting EZH2
transcription. Cell Death Dis. 9:2382018. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Liu Y, Zhao J, Zhang W, Gan J, Hu C, Huang
G and Zhang Y: lncRNA GAS5 enhances G1 cell cycle arrest via
binding to YBX1 to regulate p21 expression in stomach cancer. Sci
Rep. 5:101592015. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Zhang J, Wang P, Wan L, Xu S and Pang D:
The emergence of noncoding RNAs as Heracles in autophagy.
Autophagy. 13:1004–1024. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Klionsky DJ, Kotb A, Akihisa A, et al:
Guidelines for the use and interpretation of assays for monitoring
autophagy (3rd edition). Autophagy. 12:1–222. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Zhang L, Guo YF, Liu YZ, Liu YJ, Xiong DH,
Liu XG, Wang L, Yang TL, Lei SF, Guo Y, et al: Pathway-based
genome-wide association analysis identified the importance of
regulation-of-autophagy pathway for ultradistal radius BMD. J Bone
Miner Res. 25:1572–1580. 2010. View
Article : Google Scholar : PubMed/NCBI
|
|
12
|
Luo T, Fu J, Xu A, Su B, Ren Y, Li N, Zhu
J, Zhao X, Dai R, Cao J, et al: PSMD10/gankyrin induces autophagy
to promote tumor progression through cytoplasmic interaction with
ATG7 and nuclear transactivation of ATG7 expression. Autophagy.
12:1355–1371. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Liu M, Jiang L, Fu X, Wang W, Ma J, Tian
T, Nan K and Liang X: Cytoplasmic liver kinase B1 promotes the
growth of human lung adenocarcinoma by enhancing autophagy. Cancer
Sci. 109:3055–3067. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Su C-C: Tanshinone IIA can inhibit
MiaPaCa-2 human pancreatic cancer cells by dual blockade of the
Ras/Raf/MEK/ERK and PI3K/AKT/mTOR pathways. Oncol Rep.
40:3102–3111. 2018.PubMed/NCBI
|
|
15
|
Zhu D, Zhou J, Zhao J, Jiang G, Zhang X,
Zhang Y and Dong M: ZC3H13 suppresses colorectal cancer
proliferation and invasion via inactivating Ras-ERK signaling. J
Cell Physiol. 234:8899–8907. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Natan S, Tsarfaty G, Horev J, Haklai R,
Kloog Y and Tsarfaty I: Interplay between HGF/SF-Met-Ras signaling,
tumor metabolism and blood flow as a potential target for breast
cancer therapy. Oncoscience. 1:30–38. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Tang D, Kang R, Livesey KM, Cheh C-W,
Farkas A, Loughran P, Hoppe G, Bianchi ME, Tracey KJ, Zeh HJ III,
et al: Endogenous HMGB1 regulates autophagy. J Cell Biol.
190:881–892. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Cai M, Hu Z, Liu J, Gao J, Liu C, Liu D,
Tan M, Zhang D and Lin B: Beclin 1 expression in ovarian tissues
and its effects on ovarian cancer prognosis. Int J Mol Sci.
15:5292–5303. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Qiu DM, Wang GL, Chen L, Xu YY, He S, Cao
XL, Qin J, Zhou JM, Zhang YX and Qun E: The expression of beclin-1,
an autophagic gene, in hepatocellular carcinoma associated with
clinical pathological and prognostic significance. BMC Cancer.
14:3272014. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Nishida Y, Arakawa S, Fujitani K,
Yamaguchi H, Mizuta T, Kanaseki T, Komatsu M, Otsu K, Tsujimoto Y
and Shimizu S: Corrigendum: Discovery of Atg5/Atg7-independent
alternative macroautophagy. Nature. 533:1302016. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Ganley IG, Lam H, Wang J, Ding X, Chen S
and Jiang X: ULK1.ATG13.FIP200 complex mediates mTOR signaling and
is essential for autophagy. J Biol Chem. 284:12297–12305. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Chan EY, Kir S and Tooze SA: siRNA
screening of the kinome identifies ULK1 as a multidomain modulator
of autophagy. J Biol Chem. 282:25464–25474. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Blessing AM, Rajapakshe K, Reddy Bollu L,
Shi Y, White MA, Pham AH, Lin C, Jonsson P, Cortes CJ, Cheung E, et
al: Transcriptional regulation of core autophagy and lysosomal
genes by the androgen receptor promotes prostate cancer
progression. Autophagy. 13:506–521. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Shukla S, Patric IRP, Patil V, Shwetha SD,
Hegde AS, Chandramouli BA, Arivazhagan A, Santosh V and
Somasundaram K: Methylation silencing of ULK2, an autophagy gene,
is essential for astrocyte transformation and tumor growth. J Biol
Chem. 289:22306–22318. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) Method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Yu Y and Hann SS: Novel tumor suppressor
lncRNA growth arrest-specific 5 (GAS5) in human cancer. OncoTargets
Ther. 12:8421–8436. 2019. View Article : Google Scholar
|
|
27
|
Mirna MM, Hasan AM, Farzin F and Williams
Gwyn T: Inhibition of human T-cell proliferation by mammalian
target of rapamycin (mTOR) antagonists requires noncoding RNA
growth-arrest-specific transcript 5 (GAS5). Pharmacol. 78:19–28.
2010.
|
|
28
|
Dunlop EA and Tee AR: mTOR and autophagy:
A dynamic relationship governed by nutrients and energy. Semin Cell
Dev Biol. 36:121–129. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
He C and Beth L: The Beclin 1 interactome.
Curr Opin Cell Biol. 22:140–149. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Mizushima N, Yoshimori T and Ohsumi Y: The
role of Atg proteins in autophagosome formation. Annu Rev Cell Dev
Biol. 27:107–132. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
McEwan DG, Popovic D, Gubas A, Terawaki S,
Suzuki H, Stadel D, Coxon FP, Miranda de Stegmann D, Bhogaraju S,
Maddi K, et al: PLEKHM1 regulates autophagosome-lysosome fusion
through HOPS complex and LC3/GABARAP proteins. Mol Cell. 57:39–54.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Song J, Ahn C, Chun C-H and Jin E-J: A
long non-coding RNA, GAS5, plays a critical role in the regulation
of miR-21 during osteoarthritis. J Orthop Res. 32:1628–1635. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Hu YR, Yu YC, You SW, Li KQ, Tong XC, Chen
SR, Chen ED, Lin XZ and Chen YY: Long noncoding RNA MALAT1
regulates autophagy associated chemoresistance via miR-23b-3p
sequestration in gastric cancer. Mol Cancer. 16:1742017. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Cai Q, Wang S, Jin L, Weng M, Zhou D, Wang
J, Tang Z and Quan Z: Long non-coding RNA GBCDRlnc1 induces
chemoresistance of gallbladder cancer cells by activating
autophagy. Mol Cancer. 18:822019. View Article : Google Scholar : PubMed/NCBI
|