Introduction
Thyroid carcinoma (THCA) is a common type of
malignant tumor of the endocrine system, representing 3.4% of all
cancer types diagnosed annual (1,2). The
overall incidence of THCA is increasing in recent decades (3), with ~567,000 incident cases of THCA
reported in 2018 worldwide (4).
Moreover, the morbidity of THCA in women is three times higher
compared with men (1,5), and despite the advancement for the
treatment of THCA, the prognosis of patients remains poor (6). Therefore, it is important to provide
novel insights into the gene regulatory circuits underlying THCA
and identify new effective therapy targets.
MicroRNAs (miRNAs/miRs), a type of endogenous RNAs
with a length of 18–22 nucleotides, have been reported to be vital
regulators in various tumors (7).
For example, miRNAs are involved in numerous processes of
tumorigenesis such as cell proliferation, cell apoptosis, cell
differentiation and development (8,9). In
addition, the expression of certain genes can be regulated by
miRNAs via the binding between miRNAs and the 3′-untranslated
region (3′-UTR) of the genes (10,11).
Previous studies have reported that miRNAs can not only serve as
oncogenes, but also can exert a tumor suppressive role by
regulating the expression of the target genes (12–14).
As a mature miRNA, miR-15b-5p is spliced from the 5′-end of
pre-miR-15b, which is a member of miR-16 family (15,16).
Moreover, miR-15b-5p is upregulated in gastric cancer (15), liver cancer (17) and hepatocellular carcinoma
(18–22), while it is lowly expressed in
neuroblastoma (18) and prostate
cancer (23). However, the
biological functions and underlying mechanisms of miR-15b-5p in
THCA are yet to be elucidated.
GDP dissociation inhibitor 2 (GDI2) belongs to a
small family of chaperone proteins and is mainly expresses in
hematopoietic, endothelial and epithelial cells (24,25).
Although the dysregulation of GDI2 has been revealed in numerous
cancer types, including pancreatic carcinoma, ovarian cancer,
gastric cancer and esophageal squamous cell carcinoma (26–30),
the functions of GDI2 in cancer have been rarely reported. While
Onda et al (31)
demonstrated that GDI2 was upregulated in anaplastic thyroid
cancer, the detailed effects of GDI2 and its regulatory mechanism
in THCA remain unknown.
In the present study, the effects of miR-15b-5p on
the prognosis of THCA patients, viability and invasion of THCA
cells were identified. Moreover, the potential target gene of
miR-15b-5p in THCA was verified by bioinformatics prediction,
luciferase reporter assay and a series of rescue experiments. The
results of the present study provide a novel insight in the
pathological mechanism of THCA.
Materials and methods
Data collection and analysis
The data of tissue samples including 509 patients
with THCA (age: 15–89 years; 136 males and 373 females) and 58
healthy individuals were downloaded from the Cancer Genome Atlas
project using starbase2.0 (http://starbase.sysu.edu.cn/index.php). The
expressions of miRNAs or genes in different groups were analyzed
using starbase2.0. Then, the patients of THCA were divided into
high miR-15b-5p expression group (value ≥194.72) and low miR-15b-5p
expression group (value <194.72) according to the median
expression of miR-15b-5p. The overall survival time of patients
with THCA in two groups were analyzed using Kaplan-Meier analysis
followed by log rank test.
Prediction of target gene for
miR-15b-5p
The miRNAs target gene prediction websites
TargetScan Release 7.2 (http://www.targetscan.org/vert_72/), miRanda 2010
Release (http://www.microrna.org/microrna/getGeneForm.do) and
miRWalk version 3.0 (http://mirwalk.umm.uni-heidelberg.de) were used to
identify the target gene of miR-15b-5p. The starbase2.0 database
online tool was used to analyze the association between miR-15b-5p
and the predicted targets. Gene Expression Profiling Interactive
Analysis 2 (GEPIA2; http://gepia.cancer-pku.cn/) was used to assess the
expression of the target gene in THCA. Subsequently, log-rank test
was performed to analyze the differences of the overall survival
time between patients with high and low expression of the target
gene.
Cell lines and culture
Human THCA cell lines FTC133 (derived from
follicular THCA; cat. no. ECACC 94060901), SW1736 (derived from
anaplastic THCA; cat. no. CLS 300453) and K1 (derived from a
primary papillary THCA; cat. no. ECACC 92030501; authenticated by
Shanghai Genechem Co., Ltd. using short tandem repeat profile
analysis), as well as the thyroid epithelial cell line Nthy-ori3-1
(Nthy3; cat. no. SA 90011609) were purchased from the European
Collection of Cell Cultures, CLS Cell Lines Service GmbH or
Sigma-Aldrich (Merck KGaA). FTC133 and K1 were maintained in
DMEM/modified HAM-F12 medium (Invitrogen; Thermo Fisher Scientific,
Inc.), while SW1736 and Nthy3 were cultured in RPMI-1640 medium
(Thermo Fisher Scientific, Inc.) at 37°C with an atmosphere of 5%
CO2. Both of mediums were supplemented with 10% FBS
(Gibco; Thermo Fisher Scientific, Inc.).
Cell transfection
miR-15b-5p agomir (5′-UAGCAGCACAUCAUGGUUUACA-3′, 20
nM), miR-15b-5p antagomir (5′-TGTAAAUUATGATGTGUTGUTA-3′, 50 nM) and
corresponding negative controls (NC, 50 nM), pcDNA3.1 and
pcDNA3.1-GDI2 (1 µg/well), and small interfering RNA (si)-NC and
si-GDI2 (5′-CCAAGTTCCTTATGGCTAA-3′, 50 nM) were designed and
synthesized by Guangzhou RiboBio Co., Ltd. Then, the transfection
assays were performed using Lipofectamine® 3000
(Invitrogen; Thermo Fisher Scientific, Inc.) following the
manufacturer's instructions. The cells were harvested for
subsequent experiments, 24 h post transfection.
Assessment of cell viability using
Cell Counting Kit (CCK-8) assay
SW1736 and K1 cells with different treatments were
plated into 24-well plates (3,000 cells/well) in DMEM containing
10% FBS at 37°C. Then, 10 µl CCK-8 regent (Beijing Solarbio Science
& Technology Co., Ltd.) was added into each well and mixed for
1.5 h at 37°C, and the cell viability was measured according to the
manufacture's protocol every 24 h, from 0 to 96 h. A microplate
reader was utilized to measure the optical density value at 450
nm.
Transwell assays
Transwell assays were performed to examine cell
invasion. In brief, 100 µl Matrigel, which was incubated in
serum-free medium overnight at 37°C, was added to the upper
Transwell chamber (EMD Millipore) and subsequently inserted into
24-well plates. The cells were cultured at 37°C for 4–6 h. Then,
500 µl serum-free medium was added into lower chamber and incubated
for 1.5 h at 37°C. Subsequently, 100 µl cell suspension
(1×105 transfected cells) was prepared using medium
without serum and plated into the upper chamber, with 500 µl
complete culture solution plated into the lower chamber. After 24
h, the chambers were removed and washed with PBS, followed by
fixation with 4% paraformaldehyde for 30 min at room temperature.
Staining was performed using 0.1% crystal violet for 20 min at room
temperature and the number of invading cells was counted under a
light microscope at ×200 magnification.
Reverse transcription-quantitative PCR
(RT-qPCR)
Total RNA of the cells was extracted using
TRIzol® (Invitrogen; Thermo Fisher Scientific, Inc.). RT
was conducted using different kits: For mRNAs, PrimeScript RT
Reagent kit (Takara Bio, Inc.); and for miRNAs, MiScript RT kit
(Qiagen, Inc.). The RT reaction conditions were as follows:
Incubation at 42°C for 50 min, followed by incubation at 85°C for 5
min to terminate the reaction. Subsequently, RT-qPCR was performed
with SYBR Premix Ex Taq II (Takara Bio, Inc.) for mRNA and miScript
SYBR-Green PCR kit (Qiagen, Inc.) for miRNA in a 7900HT RT PCR
system (Applied Biosystems; Thermo Fisher Scientific, Inc.). GAPDH
and U6 were used the internal references of mRNA and miRNA,
respectively. The following thermocycling conditions were used for
qPCR: Initial denaturation at 95°C for 30 sec; 40 cycles of 95°C
for 5 sec, 60°C for 30 sec. The relative mRNA expression was
calculated with 2−ΔΔCq analysis method (32). Primers used were as follows: GDI2
forward (F), 5′-TTTCGTAAGCTTATGGACGAGGAATACGATGT-3′ and reverse
(R), 5′-TTTCGTCTCGAGGCTGTTAGTCTTCCCCATAG-3′; GAPDH F,
5′-CAACTTTGGCATCGTGGAAGGG-3′ and R, 5′-CAACGGATACATTGGGGGTAGG-3′.
miR-15b-5p F, 5′-TAGCAGCACATCATGGTTTACA-3′ and R,
5′-TGCGTGTCGTGGAGTC-3′; and U6 F, 5′-CTCGCTTCGGCAGCACA-3′ and R,
5′-AACGCTTCACGAATTTGCGT-3′.
Western blot analysis
Total protein was isolated using RIPA buffer
(Beijing Solarbio Science & Technology Co., Ltd., with protease
inhibitor) and the protein concentration was measured with a
bicinchoninic acid protein assay kit (Sigma-Aldrich; Merck KGaA).
Then, 20 µg protein/lane was added into each well of the vertical
electrophoresis tank and separated using 10% SDS-PAGE.
Subsequently, the protein was transferred onto the PVDF membrane.
After being blocked with 5% skim-milk for 1 h at room temperature,
the membranes were incubated with the primary antibodies as
follows: Matrix metalloproteinase (MMP)2 (1:300; cat. no. sc-13594;
Santa Cruz Biotechnology, Inc.), MMP9 (1:300; cat. no. sc-21733;
Santa Cruz Biotechnology, Inc.), GDI2 (1:1,000; cat. no. ab234986;
Abcam) and GAPDH (1:4,000; cat. no. sc-47724; Santa Cruz
Biotechnology) at 4°C for overnight. The membranes were washed
three times with 0.1% TBS-Tween-20 (each time 5 min) and mixed with
goat anti-rabbit/mouse horseradish peroxide-conjugated secondary
antibodies (1:5,000; cat. nos. ab205718 and ab205719; Abcam) at
room temperature for 1 h. Then, the membranes were rinsed with TBS
and developed using ECL (Thermo Fisher Scientific, Inc.). Protein
expression was semi-quantified using Quantity One software (version
4.6.6, Bio-rad Laboratories, Inc.) and normalized to GAPDH.
Dual-luciferase assay
The 3′-UTR of GDI2, harboring either the wild-type
(WT) or mutant (MT) miR-15b-5p binding site was cloned into the
downstream of pmirGLO Dual-Luciferase vector (Promega Corporation)
to generate the double luciferase reporter plasmid. Subsequently,
the co-transfections of 20 nM miR-15b-5p agomir or NC and 100 ng
double luciferase reporter plasmid into the cells were performed
utilizing Lipofectamine® 2000 (Invitrogen; Thermo Fisher
Scientific, Inc.). Luciferase activities were measured using the
Dual-Luciferase® Reporter Assay System (Promega
Corporation) after being transfected for 48 h. The results were
normalized to the Renilla luciferase activity and expressed as
relative luciferase activity (firefly luminescence/Renilla
luminescence).
Statistical analysis
SPSS 22.0 (IBM Corp.) was used to analyze the data,
while GraphPad Prism 6.0 (GraphPad Software, Inc.) was used to
create the figures. Each experiment was repeated ≥3 times. The
differences between two groups were estimated using an unpaired
Student's t-test, and ≥3 groups were analyzed using one-way ANOVA
analysis followed by Tukey's multiple comparisons post hoc test.
P<0.05 was considered to indicate a statistically significant
difference.
Results
Expression and prognostic value of
miR-15b-5p in THCA
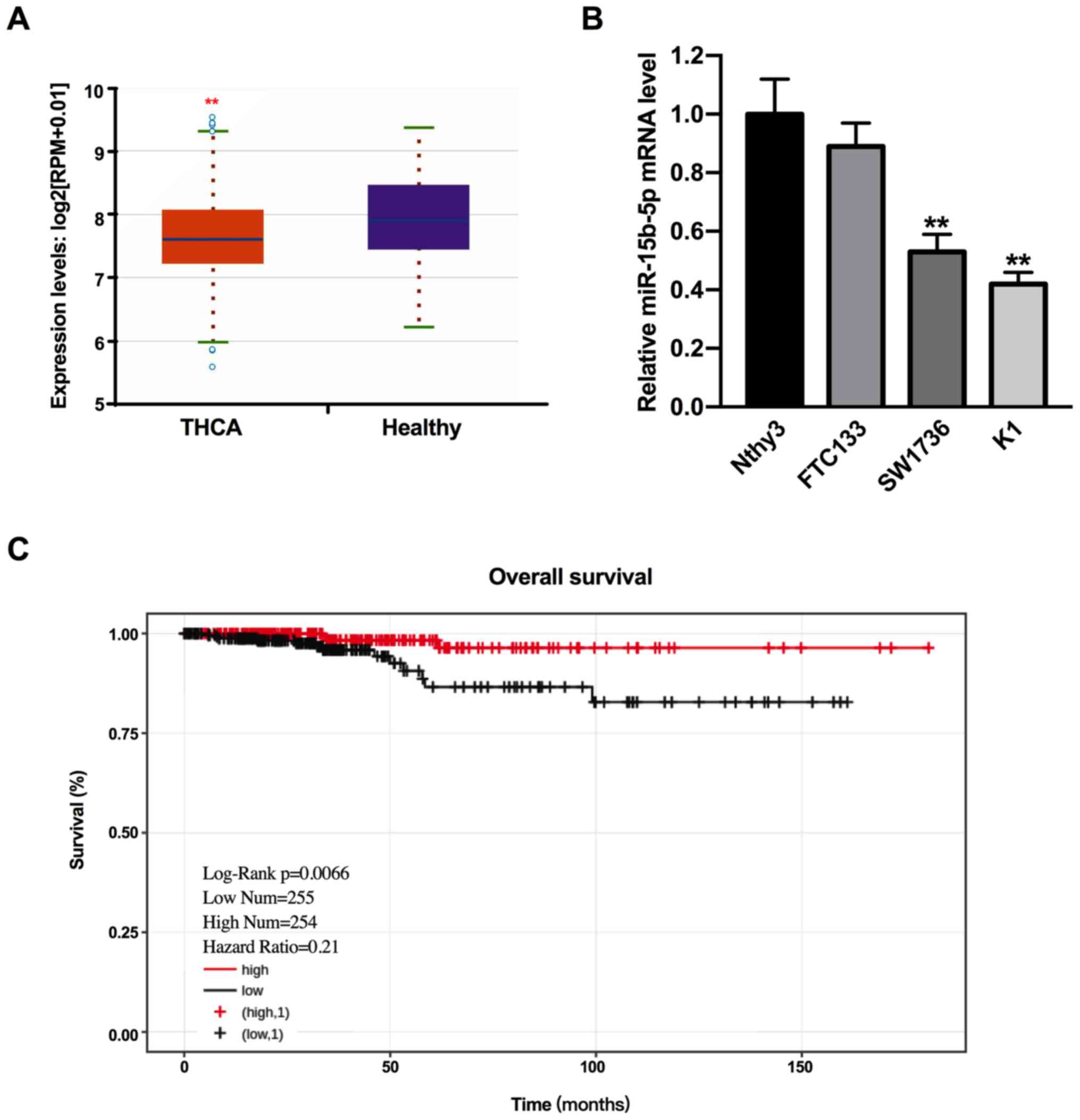
By searching the starbase2.0 database, it was found
that miR-15b-5p expression was reduced in THCA compared with
healthy individuals (Fig. 1A;
P<0.01). To assess this result, miR-15b-5p expression was
determined in human the THCA cell lines FTC133, SW1736 and K1, and
a normal cell line Nthy3. The results demonstrated that miR-15b-5p
expression was significantly decreased expressed in SW1736 and K1
cells compared with Nthy3 cells (Fig.
1B; P<0.01). While the expression of miR-15b-5p was lower in
FTC133 cells compared with Nthy3 cells, the difference was not
significant (P>0.05). Therefore, SW1736 and K1 cells were used
in the subsequent experiments.
To analyze the impacts of miR-15b-5p on the overall
survival of patients with THCA, Kaplan-Meier analysis was
performed. In total, two groups (high miR-15b-5p expression, n=254;
low miR-15b-5p expression, n=255) were determined based on the
median expression of miR-15b-5p in THCA tissues. It was
demonstrated that patients with high miR-15b-5p expression had a
longer overall survival time compared with patients with low
miR-15b-5p expression (Fig. 1C;
P<0.01). Thus, the results suggested that miR-15b-5p was
downregulated in THCA, and lower miR-15b-5p expression was related
to the tumor progression and poor prognosis of THCA.
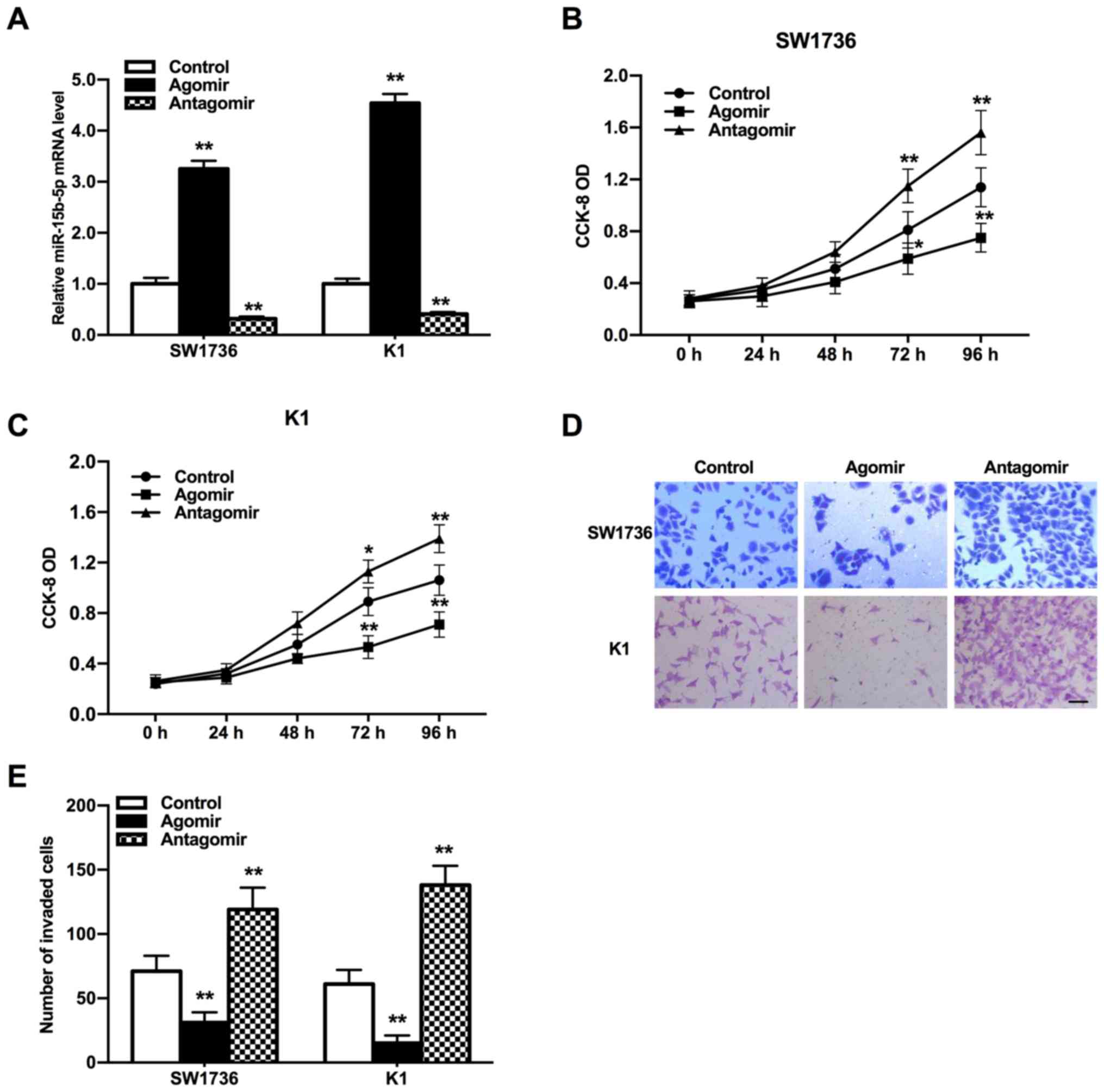
Suppression of THCA cell proliferation
and invasion induced by miR-15b-5p
To estimate the efficacy of miR-15b-5p in THCA
progression, the expression of miR-15b-5p in SW1736 and K1 cells
was altered using miR-15b-5p agomir or antagomir. Then, the
proliferative and invasive abilities of THCA cells were evaluated
using CCK-8 and Transwell assays, respectively. miR-15b-5p
expression was significantly increased in miR-15b-5p agomir group,
while it was significantly reduced in miR-15b-5p antagomir group
compared with the control group (Fig.
2A; P<0.01). CCK-8 assay results indicated that the THCA
cell proliferation was significantly accelerated by the miR-15b-5p
antagomir, but was suppressed by the miR-15b-5p agomir compared
with the control (Fig. 2B and C;
P<0.05 and P<0.01). According to Transwell assay results, the
miR-15b-5p agomir reduced the invasion of THCA cells, while the
invasive ability of those treated by miR-15b-5p antagomir was
increased (Fig. 2D and E;
P<0.01). Therefore, it was indicated that miR-15b-5p may be an
inhibitor in the tumor development of THCA.
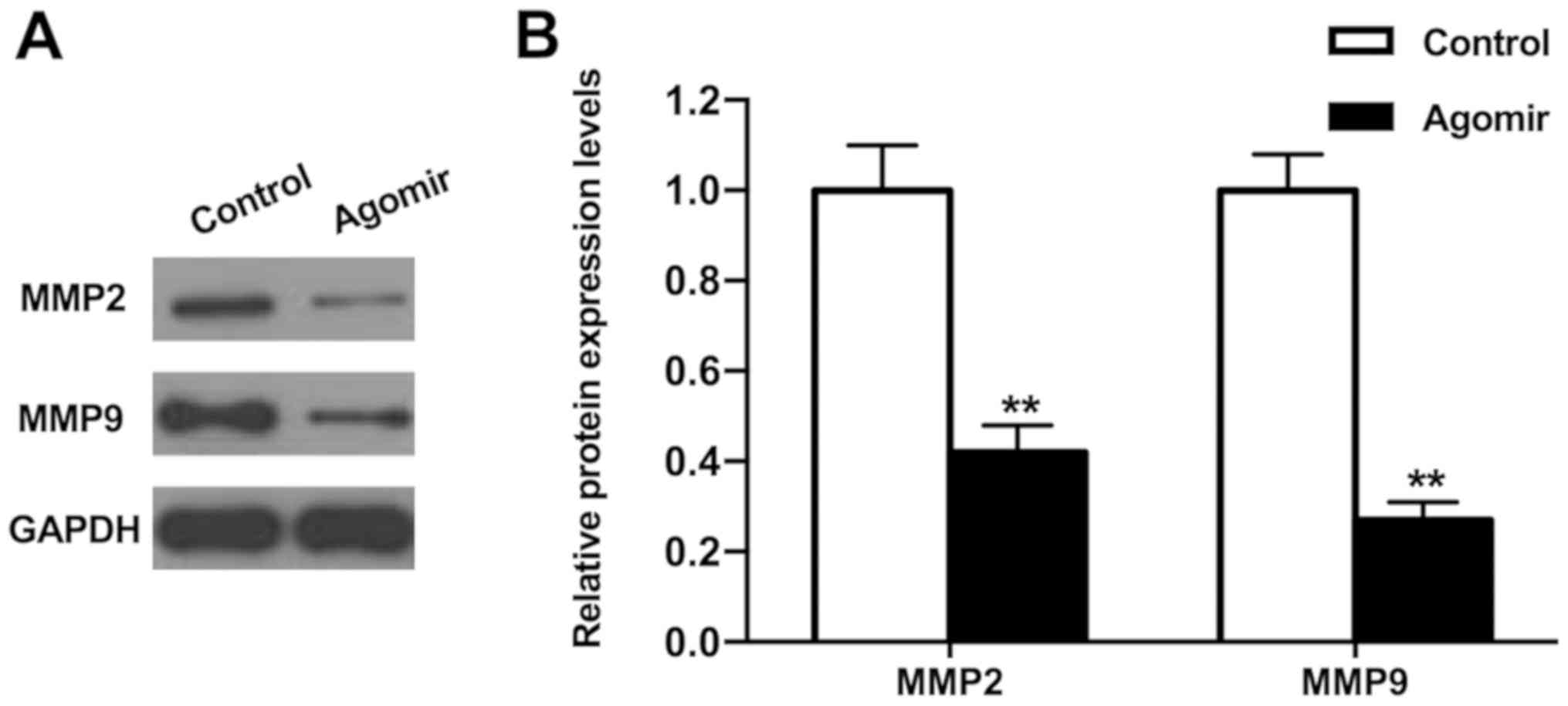
miR-15b-5p regulates the expression
levels of invasion-associated proteins MMP2 and MMP9
To investigate how miR-15b-5p exerted its effects on
cell invasion, the expression levels of invasion-associated
proteins MMP2 and MMP9 were measured using western blot analysis.
Overexpression of miR-15b-5p significantly reduced the protein
expression levels of MMP2 and MMP9 compared with the control
(Fig. 3; P<0.01). Thus,
miR-15b-5p may suppress the invasion of THCA cells by inhibiting
the expression levels of MMP2 and MMP9.
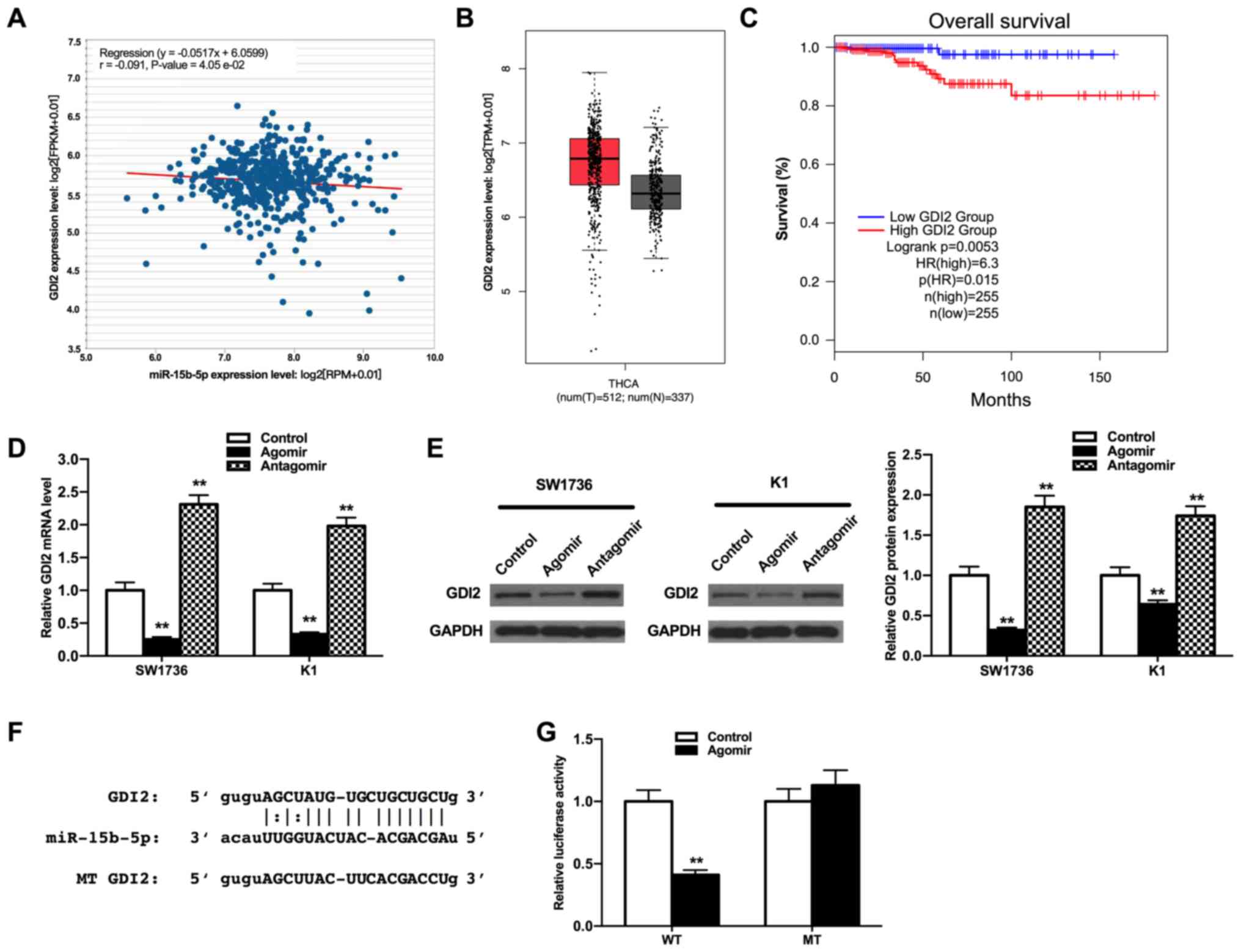
GDI2 is targeted by miR-15b-5p and
related to the prognosis of patients with THCA
Next, the target genes of miR-15b-5p were examined
using TargetScan, miRanda and miRWalk online tools, and GDI2 was
obtained via the intersection of these databases. Then, the
relevance of miR-15b-5p and GDI2 expression levels were analyzed
using starbase website. The results demonstrated that miR-15b-5p
expression was negatively associated with GDI2 expression (Fig. 4A; P<0.05). Moreover, there was a
slight increase in GDI2 expression in patients with THCA compared
with healthy controls, as determined via the online analysis tool
of the GEPIA website (Fig. 4B;
P>0.05), but the high expression of GDI2 was found to predict a
poor prognosis of patients with THCA (Fig. 4C; P<0.01), suggesting the
involvement of GDI2 in THCA progression.
Subsequently, the effect of miR-15b-5p on the
expression of GDI2 in THCA cells were measured using RT-qPCR and
western blot analyses. The miR-15b-5p agomir repressed the relative
mRNA and protein expression of GDI2, while the miR-15b-5p antagomir
increased this expression (Fig. 4D and
E; all P<0.01), which indicated that the expression of GDI2
was negatively modulated by miR-15b-5p.
To assess whether miR-15b-5p regulated GDI2 by
binding to the 3′-UTR of GDI2, a dual-luciferase reporter assay was
performed. The binding sites between miR-15b-5p and GDI2 were
predicted via the bioinformatics analysis as aforementioned
(Fig. 4F). The luciferase activity
of pmirGLO-GDI2-WT in the miR-15b-5p agomir group was significantly
lower compared with the control group (Fig. 4G; P<0.01); however, the
luciferase activity of pmirGLO-GDI2-MT had no significant
differences between the two groups, suggesting that GDI2 may be the
downstream target genes of miR-15b-5p in the tumor progression of
THCA. Collectively, these results indicated that GDI2 was
upregulated in THCA and may be involved in the progression of THCA
modulated by miR-15b-5p.
GDI2 abolishes the effects of
miR-15b-5p on the phenotype of THCA cells
To examine whether GDI2 was involved in the effects
of miR-15b-5p during the tumor progression, a transfection assay
was conducted on the SW1736 and K1 cells with si-con and si-GDI2,
as well as pcDNA3.1 and pcDNA3.1-GDI2. RT-qPCR and western blot
analyses demonstrated that the mRNA and protein expression levels
of GDI2 were both downregulated after the cells were transfected
with si-GDI2, while these were upregulated in cells transfected
with pcDNA3.1-GDI2 compared with the control (Fig. 5A-C; all P<0.01).
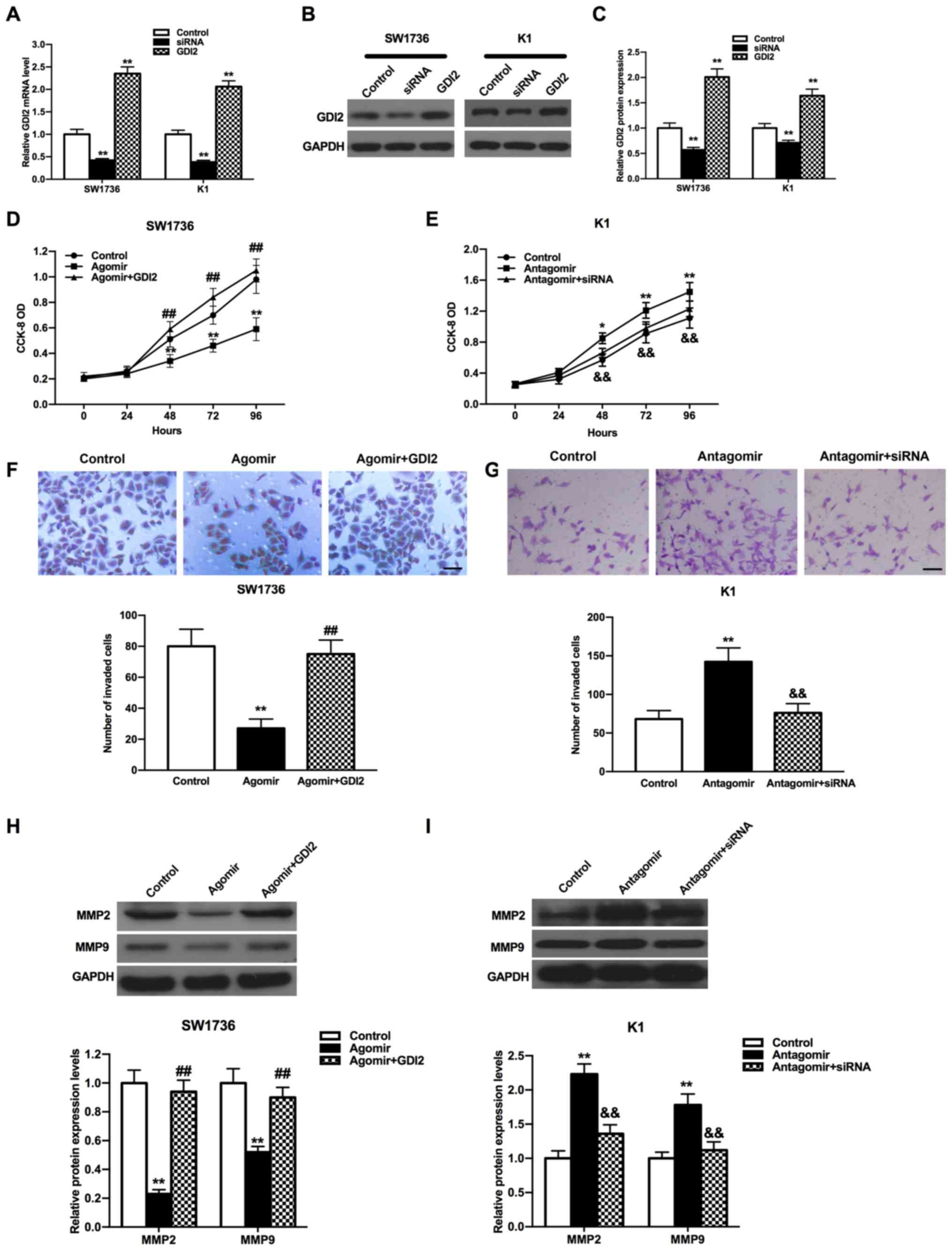 | Figure 5.Impacts of miR-15b-5p and GDI2 on the
proliferation and invasion of THCA cells. GDI2 expression was
reduced or induced by si-GDI2 or pcDNA3.1-GDI2 in SW1736 and K1
cells both at (A) mRNA and (B) protein levels, (C) which were
semi-quantified from western blot analysis. Control group cells
were treated with a mixture of the scrambled siRNAs and pcDNA3.1.
Proliferation of (D) SW1736 and (E) K1 cells were attenuated or
accelerated by miR-15b-5p agomir or miR-15b-5p antagomir, which was
restored by pcDNA3.1-GDI2 or si-GDI2. Inhibitory or stimulative
effect of miR-15b-5p agomir or miR-15b-5p antagomir on (F) SW1736
and (G) K1 cell invasion was weakened by pcDNA3.1-GDI2 or si-GDI2.
Scale bar, 200 µm. pcDNA3.1-GDI2 or si-GDI2 increased or decreased
the expression levels of MMP2 and MMP9, which were caused by
miR-15b-5p agomir or miR-15b-5p antagomir, in (H) SW1736 and (I) K1
cells. Control group cells were treated with the mixture of
miR-15b-5p agomir control and pcDNA3.1 in D, F and H, or the
mixture of miR-15b-5p antagomir control and the scrambled siRNAs in
E, G and I. All experiments were repeated three times. *P<0.05,
**P<0.01 vs. control group; ##P<0.01 vs. agomir
group; &&P<0.01 vs. the antagomir group. miR,
microRNA; GDI2, GDP dissociation inhibitor 2; siRNA, small
interfering RNA; MMP, matrix metalloproteinase; CCK-8, Cell
Counting Kit-8; OD, optical density. |
CCK-8 and Transwell assays were performed to
evaluate the functions of the miR-15b-5p/GDI2 axis in the
proliferation and invasion of THCA cells. It was found that the
proliferation of THCA cells was increased in the miR-15b-5p agomir
+ pcDNA3.1-GDI2 group compared with the miR-15b-5p agomir group,
but the proliferative ability in miR-15b-5p antagomir + si-GDI2
group was significantly reduced compared with the miR-15b-5p
antagomir group (Fig. 5D and E;
all P<0.01). Furthermore, the invasive abilities of THCA cells
in the miR-15b-5p agomir + pcDNA3.1-GDI2 group were significantly
enhanced compared with the miR-15b-5p agomir group, while the
knockdown of GDI2 and miR-15b-5p together suppressed the invasion
of THCA cells compared with the miR-15b-5p antagomir group
(Fig. 5F and G; all P<0.01). It
was also found that the proliferative and invasive abilities of
THCA cells in miR-15b-5p agomir + pcDNA3.1-GDI2 group and
miR-15b-5p antagomir + si-GDI2 group were both almost return to the
level of control group.
To determine the relative expression levels of the
invasion-associated proteins MMP2 and MMP9, western blot analysis
was conducted. MMP2 and MMP9 expression levels were both increased
in the miR-15b-5p agomir + pcDNA3.1-GDI2 group compared with the
miR-15b-5p agomir group in SW1736 cells (Fig. 5H; P<0.01). In addition, the
expression levels of MMP2 and MMP9 were both decreased in
miR-15b-5p antagomir + si-GDI2 group compared with the miR-15b-5p
antagomir group in K1 cells (Fig.
5I; P<0.01). It was also demonstrated that the expression
levels of MMP2 and MMP9 could be both recovered to similar levels
as the control in miR-15b-5p agomir + pcDNA3.1-GDI2 group or
miR-15b-5p antagomir + si-GDI2 group (P>0.05). Collectively, the
results suggested that miR-15b-5p could inhibit cell proliferation
and invasion, and regulate the expression levels of MMP2 and MMP9
via targeting GDI2.
Discussion
Recently, the incidence of THCA has been continually
increasing and is higher compared with other cancer types (33). Despite THCA having a relative low
mortality rate, patients with advanced THCA still have an
undesirable prognosis (34–36).
Thus, identifying novel molecular markers of THCA is of importance
for its effective treatment. The present study identified the low
expression of miR-15b-5p in THCA, which was associated with a
shorter overall survival time of patients with THCA, as well as
promoted the proliferation and invasion of THCA cells via
regulating the expression levels of the invasion-associated
proteins MMP2 and MMP9.
Previous studies have reported that miRNAs serve
vital roles in the oncogenesis of THCA due to their effects on the
regulation of biological features in THCA (37). For example, miR-136-5p, miR-26a,
miR-214 and miR-1266 are abnormally expressed in THCA and modulate
the tumor progression of THCA (37–40).
miR-15b-5p is the dominant isoform of miR-15b, which is located on
the long arm of chromosome 3 (41), and has been revealed to be
dysregulated and involved in the tumor development processes of
various cancer types. For instance, miR-15b-5p is upregulated in
gastric cancer and facilitates tumor metastasis by targeting
progestin and adipoQ receptor family member 3 (15). Moreover, the overexpression of
miR-15b-5p has been shown to promote the proliferation and invasion
of liver cancer cells via targeting axis formation inhibitor 2
(17). The upregulation of
miR-15b-5p has also been reported to be involved in
hepatocarcinogenesis via various mechanisms, including inducing
endoplasmic reticulum stress and apoptosis by regulating Rab1A
(21), and acts as a diagnostic
marker in hepatocellular carcinoma (19–22).
Furthermore, knockdown of miR-15b-5p could inhibit the tumor growth
of prostate cancer by targeting reversion inducing cysteine rich
protein with kazal motifs (23).
The inhibitory effects of miR-15b-5p have been
revealed in neuroblastoma, and it can restrain tumor progression by
directly targeting MYCN proto-oncogene, bHLH transcription factor
(18). Luo et al (42) demonstrated that miR-15b-5p reduced
proliferation, but induced apoptosis and cytotoxic activities of
PC12 cells. Moreover, Ahmad et al (43) reported that miR-15b-5p could be a
potential biomarker for the treatment of squamous cell carcinoma.
miR-15p-5b expression also exhibits a downward trend in
osteosarcoma (44), colon cancer
(45) and squamous cell carcinoma
(46). Consistent with the
decrease in miR-15b-5p expression in these aforementioned cancer
types, the present results suggested that miR-15b-5p was
downregulated in THCA. Furthermore, miR-15b-5p was found to inhibit
cell proliferation and invasion, which was consistent with the
suppressive effect observed in neuroblastoma (18). MMP2 and MMP9 are important
inflammatory biomarkers and the increased expression of these
factors has been shown to facilitate cell invasion in THCA
(47,48). In line with this finding, the
current results indicated that miR-15b-5p suppressed THCA cell
invasion via regulating the expression levels of MMP2 and MMP9. To
the best of our knowledge, the present study was the first to
investigate the functions of miR-15b-5p in THCA and may provide a
novel therapy target site for THCA.
miRNAs can mediate the expression of the target
genes via binding to their 3′-UTR (49). Previous studies have revealed that
miR-15b-5p can exert effects in the tumor progression via targeting
different genes, such as Rab1A, MYCN and Axin2 (17,18,21).
Therefore, the present study examined the potential targets of
miR-15b-5p and GDI2 was recognized as the target gene of
miR-15b-5p. GDI2 has been reported to serve a key role in several
cancer types. For instance, the upregulation of GDI2 has been
identified in pancreatic cancer (26) and esophageal squamous cell
carcinoma (30), indicating that
GDI2 could be potential molecular targets or biomarkers for these
cancer types. In addition, the downregulation of GDI2 has been
observed in ovarian cancer and it may be a potential biomarker of
the paclitaxel-resistant ovarian cancer for tailored cancer therapy
(29). The present results
identified a high expression of GDI2 in THCA and its negative
association with the overall survival of patients with THCA. It was
also found that the inhibitory effects of miR-15b-5p on the
proliferative and invasive abilities of THCA cells were both
weakened by the overexpression of GDI2. Furthermore, GDI2 increased
the low expression levels of MMP2 and MMP9, which were inhibited by
the overexpression of miR-15b-5p. Therefore, it was speculated that
the regulatory roles of miR-15b-5p were mediated by GDI2 in THCA.
However, the mechanism via which the miR-15b-5p/GDI2 axis
communicates with MMP2 and MMP9 requires further investigation.
While the expression levels of miRNAs-15b-5 and GDI2 in the THCA
were changed significantly compared with healthy tissues, the
extensive overlapping between the individual values suggested the
use of miRNAs-15b-5 and GDI2 values as tumor biomarkers should be
moderated.
In conclusion, the current results demonstrated that
miR-15b-5p could prevent the progression of THCA by binding and
negatively regulating GDI2 expression. Therefore, these preliminary
findings indicated the functions and underlying mechanisms of
miR-15b-5p in THCA mainly via in vitro cells experiments,
and provided a novel potential therapeutic target for THCA
treatment in the future. However, additional studies are required
to verify the validity of aforementioned conclusions in
vivo.
Acknowledgements
Not applicable.
Funding
The present study was funded by the Research Award
Fund for Outstanding Young and Middle-Aged Scientists in Shandong
Province (grant no. BS2010YY059).
Availability of data and materials
The datasets used and/or analyzed during the present
study are available from the corresponding author on reasonable
request.
Authors' contributions
JZ designed the study and wrote the manuscript. JQ,
HF, FY, WZ and LX performed the experiments and statistical
analysis. All authors read and approved the final manuscript.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Ward EM, Sherman RL, Henley SJ, Jemal A,
Siegal DA, Feuer EJ, Firth AU, Kohler BA, Scott S, et al: Annual
report to the nation on the status of cancer, featuring cancer in
men and women age 20–49 years. J Natl Cancer Inst. 111:2723–1297.
2019. View Article : Google Scholar
|
|
2
|
Torre LA, Bray F, Siegel RL, Ferlay J,
Lortet-Tieulent J and Jemal A: Global cancer statistics, 2012. CA
Cancer J Clin. 65:87–108. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Huk DJ, Ashtekar A, Magner A, La Perle K
and Kirschner LS: Deletion of Rap1b, but not Rap1a or Epac1,
reduces protein kinase A-mediated thyroid cancer. Thyroid.
28:1153–1161. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Bray F, Ferlay J, Soerjomataram I, Siegel
RL, Torre LA and Jemal A: Global cancer statistics 2018: GLOBOCAN
estimates of incidence and mortality worldwide for 36 cancers in
185 countries. CA Cancer J Clin. 68:394–424. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Luzon-Toro B, Fernandez RM,
Villalba-Benito L, Torroglosa A, Antinolo G and Borrego S:
Influencers on thyroid cancer onset: Molecular genetic basis. Genes
(Basel). 10:9132019. View Article : Google Scholar
|
|
6
|
Tella SH, Kommalapati A, Esquivel MA and
Correa R: Potential role of metabolic intervention in the
management of advanced differentiated thyroid cancer. Front Oncol.
7:1602017. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Ma Y and Sun Y: miR-29a-3p inhibits
growth, proliferation, and invasion of papillary thyroid carcinoma
by suppressing NF-κB signaling via direct targeting of OTUB2.
Cancer Manag Res. 11:13–23. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
McKenna LB, Schug J, Vourekas A, McKenna
JB, Bramswig NC, Friedman JR and Kaestner KH: MicroRNAs control
intestinal epithelial differentiation, architecture, and barrier
function. Gastroenterology. 139:1654–1664. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Wojtas B, Ferraz C, Stokowy T, Hauptmann
S, Lange D, Dralle H, Musholt T, Jarzab B, Paschke R and Eszlinger
M: Differential miRNA expression defines migration and reduced
apoptosis in follicular thyroid carcinomas. Mol Cell Endocrinol.
388:1–9. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Guo H, Ingolia NT, Weissman JS and Bartel
DP: Mammalian microRNAs predominantly act to decrease target mRNA
levels. Nature. 466:835–840. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Ling H, Fabbri M and Calin GA: MicroRNAs
and other non-coding RNAs as targets for anticancer drug
development. Nat Rev Drug Discov. 12:847–865. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Takahashi RU, Prieto-Vila M, Hironaka A
and Ochiya T: The role of extracellular vesicle microRNAs in cancer
biology. Clin Chem Lab Med. 55:648–656. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Gao XB, Chen CL, Tian ZL, Yuan FK and Jia
GL: MicroRNA-791 is an independent prognostic factor of papillary
thyroid carcinoma and inhibits the proliferation of PTC cells. Eur
Rev Med Pharmacol Sci. 22:5562–5568. 2018.PubMed/NCBI
|
|
14
|
Mishra S, Yadav T and Rani V: Exploring
miRNA based approaches in cancer diagnostics and therapeutics. Crit
Rev Oncol Hematol. 98:12–23. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Zhao C, Li Y, Chen G, Wang F, Shen Z and
Zhou R: Overexpression of miR-15b-5p promotes gastric cancer
metastasis by regulating PAQR3. Oncol Rep. 38:352–358. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Rissland OS, Hong SJ and Bartel DP:
MicroRNA destabilization enables dynamic regulation of the miR-16
family in response to cell-cycle changes. Mol Cell. 43:993–1004.
2011. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Dong Y, Zhang N, Zhao S, Chen X, Li F and
Tao X: miR-221-3p and miR-15b-5p promote cell proliferation and
invasion by targeting Axin2 in liver cancer. Oncol Lett.
18:6491–6500. 2019.PubMed/NCBI
|
|
18
|
Chava S, Reynolds PC, Pathania AS,
Gorantla S, Poluektova LY, Coulter DW, Gupta SC, Pandey MK and
Challagundla KB: miR-15a-5p, miR-15b-5p, and miR-16-5p inhibit
tumor progression by directly targeting MYCN in neuroblastoma. Mol
Oncol. 14:180–196. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Pan WY, Zeng JH, Wen DY, Wang JY, Wang PP,
Chen G and Feng ZB: Oncogenic value of microRNA-15b-5p in
hepatocellular carcinoma and a bioinformatics investigation. Oncol
Lett. 17:1695–1713. 2019.PubMed/NCBI
|
|
20
|
Li H, Zhang J, Lee MJ, Yu GR, Han X and
Kim DG: OIP5, a target of miR-15b-5p, regulates hepatocellular
carcinoma growth and metastasis through the AKT/mTORC1 and
b-catenin signaling pathways. Oncotarget. 8:18129–18144. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Yang Y, Hou N, Wang X, Wang L, Chang S, He
K, Zhao Z, Zhao X, Song T and Huang C: miR-15b-5p induces
endoplasmic reticulum stress and apoptosis in human hepatocellular
carcinoma, both in vitro and in vivo, by suppressing Rab1A.
Oncotarget. 6:16227–16238. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Chen Y, Chen J, Liu Y, Li S and Huang P:
Plasma miR-15b-5p, miR-338-5p, and miR-764 as biomarkers for
hepatocellular carcinoma. Med Sci Monit. 21:1864–1871. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Chen R, Sheng L, Zhang HJ, Ji M and Qian
WQ: miR-15b-5p facilitates the tumorigenicity by targeting RECK and
predicts tumour recurrence in prostate cancer. J Cell Mol Med.
22:1855–1863. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Theodorescu D, Sapinoso LM, Conaway MR,
Oxford G, Hampton GM and Frierson HF Jr: Reduced expression of
metastasis suppressor RhoGDI2 is associated with decreased survival
for patients with bladder cancer. Clin Cancer Res. 10:3800–3806.
2004. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Alcántara-Hernández R, Casas-González P
and García-Sáinz JA: Roles of c-Src in alpha1B-adrenoceptor
phosphorylation and desensitization. Auton Autacoid Pharmacol.
28:29–39. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Sun ZL, Zhu Y, Wang FQ, Chen R, Peng T,
Fan ZN, Xu ZK and Miao Y: Serum proteomic-based analysis of
pancreatic carcinoma for the identification of potential cancer
biomarkers. Biochim Biophys Acta. 1774:764–771. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Zhang XY, Hong SS, Zhang M, Cai QQ, Zhang
MX and Xu CJ: Proteomic alterations of fibroblasts induced by
ovarian cancer cells reveal potential cancer targets. Neoplasma.
65:104–112. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Bai Z, Ye Y, Liang B, Xu F, Zhang H, Zhang
Y, Peng J, Shen D, Cui Z, Zhang Z and Wang S: Proteomics-based
identification of a group of apoptosis-related proteins and
biomarkers in gastric cancer. Int J Oncol. 38:375–383.
2011.PubMed/NCBI
|
|
29
|
Lee DH, Chung K, Song JA, Kim TH, Kang H,
Huh JH, Jung SG, Ko JJ and An HJ: Proteomic identification of
paclitaxel-resistance associated hnRNP A2 and GDI 2 proteins in
human ovarian cancer cells. J Proteome Res. 9:5668–5676. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Kashyap MK, Harsha HC, Renuse S, Pawar H,
Sahasrabuddhe NA, Kim MS, Marimuthu A, Keerthikumar S, Muthusamy B,
Kandasamy K, et al: SILAC-based quantitative proteomic approach to
identify potential biomarkers from the esophageal squamous cell
carcinoma secretome. Cancer Biol Ther. 10:796–810. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Onda M, Emi M, Yoshida A, Miyamoto S,
Akaishi J, Asaka S, Mizutani K, Shimizu K, Nagahama M, Ito K, et
al: Comprehensive gene expression profiling of anaplastic thyroid
cancers with cDNA microarray of 25 344 genes. Endocr Relat Cancer.
11:843–854. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Lubitz CC, Zhan T, Gunda V, Amin S,
Gigliotti BJ, Fingeret AL, Holm TM, Wachtel H, Sadow PM, Wirth LJ,
et al: Circulating BRAF(V600E) Levels Correlate with Treatment in
Patients with Thyroid Carcinoma. Thyroid. 28:328–339. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Wei WJ, Zhang GQ and Luo QY: Postsurgical
management of differentiated thyroid cancer in China. Trends
Endocrinol Metab. 29:71–73. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Fu G, Polyakova O, MacMillan C, Ralhan R
and Walfish PG: Programmed death-ligand 1 expression distinguishes
invasive encapsulated follicular variant of papillary thyroid
carcinoma from noninvasive follicular thyroid neoplasm with
papillary-like nuclear features. EBioMedicine. 18:50–55. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Lin P, He Y, Wen DY, Li XJ, Zeng JJ, Mo
WJ, Li Q, Peng JB, Wu YQ, Pan DH, et al: Comprehensive analysis of
the clinical significance and prospective molecular mechanisms of
differentially expressed autophagy-related genes in thyroid cancer.
Int J Oncol. 53:603–619. 2018.PubMed/NCBI
|
|
37
|
Gao RZ, Que Q, Lin P, Pang YY, Wu HY, Li
XJ, Chen G, He Y and Yang H: Clinical roles of miR-136-5p and its
target metadherin in thyroid carcinoma. Am J Transl Res.
11:6754–6774. 2019.PubMed/NCBI
|
|
38
|
Wu YC, Li SY and Jia YF: MicroRNA-26a
suppresses the malignant biological behaviors of papillary thyroid
carcinoma by targeting ROCK1 and regulating PI3K/AKT signaling
pathway. Eur Rev Med Pharmacol Sci. 23:8940–8949. 2019.PubMed/NCBI
|
|
39
|
Liu F, Lou K, Zhao X, Zhang J, Chen W,
Qian Y, Zhao Y, Zhu Y and Zhang Y: miR-214 regulates papillary
thyroid carcinoma cell proliferation and metastasis by targeting
PSMD10. Int J Mol Med. 42:3027–3036. 2018.PubMed/NCBI
|
|
40
|
Fu YT, Zheng HB, Zhang DQ, Zhou L and Sun
H: MicroRNA-1266 suppresses papillary thyroid carcinoma cell
metastasis and growth via targeting FGFR2. Eur Rev Med Pharmacol
Sci. 22:3430–3438. 2018.PubMed/NCBI
|
|
41
|
Salimi S, Noorbakhsh F, Faghihzadeh S,
Ghaffarpour S and Ghazanfari T: Expression of miR-15b-5p,
miR-21-5p, and SMAD7 in lung tissue of sulfur mustard-exposed
individuals with long-term pulmonary complications. Iran J Allergy
Asthma Immunol. 18:332–339. 2019.PubMed/NCBI
|
|
42
|
Luo H, Li Y, Liu B, Yang Y and Xu ZD:
MicroRNA-15b-5p targets ERK1 to regulate proliferation and
apoptosis in rat PC12 cells. Biomed Pharmacother. 92:1023–1029.
2017. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Ahmad P, Sana J, Slavik M, Gurin D, Radova
L, Gablo NA, Kazda T, Smilek P, Horakova Z, Gal B, et al:
MicroRNA-15b-5p predicts locoregional relapse in head and neck
carcinoma patients treated with intensity-modulated radiotherapy.
Cancer Genomics Proteomics. 16:139–146. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Weng Y, Shen Y, He Y, Pan X, Xu J, Jiang
Y, Zhang Q, Wang S, Kong F, Zhao S, et al: The miR-15b-5p/PDK4 axis
regulates osteosarcoma proliferation through modulation of the
Warburg effect. Biochem Biophys Res Commun. 503:2749–2757. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Zhao C, Zhao Q, Zhang C, Wang G, Yao Y,
Huang X, Zhan F, Zhu Y, Shi J, Chen J, et al: miR-15b-5p
resensitizes colon cancer cells to 5-fluorouracil by promoting
apoptosis via the NF-κB/XIAP axis. Sci Rep. 7:41942017. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Jin X, Chen Y, Chen H, Fei S, Chen D, Cai
X, Liu L, Lin B, Su H, Zhao L, et al: Evaluation of tumor-derived
exosomal miRNA as potential diagnostic biomarkers for early-stage
non-small cell lung cancer using next-generation sequencing. Clin
Cancer Res. 23:5311–5319. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Gu M: IL13Ralpha2 siRNA inhibited cell
proliferation, induced cell apoptosis, and suppressed cell invasion
in papillary thyroid carcinoma cells. Onco Targets Ther.
11:1345–1352. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Huang LL, Wang Z, Cao CJ, Ke ZF, Wang F,
Wang R, Luo CQ, Lu X and Wang LT: AEG-1 associates with metastasis
in papillary thyroid cancer through upregulation of MMP2/9. Int J
Oncol. 51:812–822. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Liu ZM, Wu ZY, Li WH, Wang LQ, Wan JN and
Zhong Y: MiR-96-5p promotes the proliferation, invasion and
metastasis of papillary thyroid carcinoma through down-regulating
CCDC67. Eur Rev Med Pharmacol Sci. 23:3421–3430. 2019.PubMed/NCBI
|