Introduction
Breast cancer accounts for 25% of cancer cases in
women and is one of the four most common cancers in women worldwide
(1). The majority of individuals
affected include patients from developing countries, which
represent 53% of the total breast cancer cases (1). Therefore, breast cancer has emerged
as the second leading cause of cancer-related deaths among women in
developing countries, following lung cancer (2). It has been reported that >80% of
the genome is actively transcribed to non-coding RNAs (ncRNAs).
This includes RNA molecules that serve no protein-coding function.
Based on their size, ncRNAs are classified into long ncRNAs
(lncRNAs) and small ncRNAs (3).
Aberrant expression of lncRNAs can be observed in almost all tumors
and can affect tumor development via an oncogenic or tumor
suppressive mechanism of action. Moreover, the expression levels of
lncRNAs closely correlate with malignancy status and disease
prognosis (4).
High expression in hepatocellular carcinoma (HEIH)
is a lncRNA that was initially identified in hepatitis B
virus-induced hepatocellular carcinoma (5). High HEIH expression is associated
with an increased risk of recurrence and significantly reduced
overall postoperative survival in patients with hepatocellular
carcinoma. Moreover, HEIH acts as a tumor promoter by accelerating
the cell cycle progression of hepatocellular carcinoma cells
(5,6). Recent studies have suggested that
HEIH is highly expressed in other cancer types, including
colorectal cancer (7), melanoma
(8) and non-small cell lung cancer
(9). However, the expression,
pathophysiological roles and clinical significance of HEIH in
breast cancer remain unknown.
In the present study, the expression pattern of HEIH
was explored in breast cancer and the data revealed the
upregulation of HEIH expression in breast cancer tissues compared
with that noted in the paired adjacent normal tissues. The
association between HEIH expression and clinical variables in
patients with breast cancer was further investigated. To determine
the functional role of HEIH in the development of breast cancer,
the effects of HEIH knockdown on the proliferation, apoptosis and
metastasis of breast cancer cells were investigated. The present
study provides a more in-depth understanding of HEIH function,
which may aid the diagnosis and targeted treatment of breast
cancer.
Materials and methods
Patient samples
Cancer tissues and paired adjacent normal tissues
(~5 cm from tumor) were obtained from 160 female patients (age,
40–68 years) with breast cancer who underwent surgical operation at
the First Hospital of Lanzhou University (Gansu, China) between
January 2013 and December 2016. The clinical information of the
patients, including age, family history, tumor grade,
tumor-node-metastasis (TNM) stage, lymph node status, estrogen
receptor (ER) status, progesterone receptor (PR) status and human
epidermal growth factor receptor-2 (HER2) status, were collected
(Table I). Following surgery, the
tissue samples obtained were stored immediately in liquid nitrogen,
and then transferred into a freezer at an ultra-low temperature
(−80°C) until further processing for reverse
transcription-quantitative PCR (RT-qPCR).
 | Table I.Association between lncRNA HEIH and
clinical characteristics. |
Table I.
Association between lncRNA HEIH and
clinical characteristics.
|
|
| HEIH expression |
|
|---|
|
|
|
|
|
|---|
| Factors | Number of
patients | Low (n=77) | High (n=83) | P-valuea |
|---|
| Age, years |
|
|
| 0.188 |
|
<60 | 80 | 42 | 38 |
|
| ≥60 | 80 | 35 | 45 |
|
| Family history |
|
|
| 0.472 |
|
Absent | 147 | 72 | 75 |
|
|
Present | 13 | 5 | 8 |
|
| Tumor grade |
|
|
| 0.776 |
| I–II | 125 | 62 | 63 |
|
| III | 35 | 15 | 20 |
|
| TNM stage |
|
|
| 0.006 |
| I | 39 | 28 | 11 |
|
| II | 55 | 32 | 23 |
|
|
III | 45 | 13 | 32 |
|
| IV | 21 | 4 | 17 |
|
| Lymph node
metastasis |
|
|
| 0.017 |
|
Negative | 78 | 27 | 51 |
|
|
Positive | 82 | 50 | 32 |
|
| ER status |
|
|
| 0.696 |
|
Negative | 87 | 43 | 44 |
|
|
Positive | 73 | 34 | 39 |
|
| PR status |
|
|
| 0.574 |
|
Negative | 20 | 8 | 12 |
|
|
Positive | 140 | 69 | 71 |
|
| HER2 status |
|
|
| 0.726 |
|
Negative | 47 | 22 | 25 |
|
|
Positive | 113 | 55 | 58 |
|
RT-qPCR
Total RNA was extracted from the tissues and cells
using TRIzol® reagent (Invitrogen; Thermo Fisher
Scientific, Inc.). Total RNA (0.2-0.5 µg) was reverse transcribed
into cDNA using the PrimeScript RT Reagent kit (cat. no. RR037B;
Takara Biotechnology Co., Ltd.) at 37°C for 15 min. qPCR was
subsequently performed using SYBR Premix Ex Taq™ (cat. no. RR420B;
Takara Biotechnology Co., Ltd.) on an ABI 7300 Real-Time PCR system
(Applied Biosystems; Thermo Fisher Scientific, Inc.). The following
thermocycling conditions were used for the qPCR: Initial incubation
at 95°C for 15 sec; followed by 40 cycles of denaturation at 95°C
for 5 sec and annellation at 60°C for 31 sec. The primer sequences
used were as follows: HEIH forward 5′-CCTCTTGTGCCCCTTTCT-3′ and
reverse, 5′-AGGTCTCATGGCTTCTCG-3′; and β-actin forward,
5′-GGGAAATCGTGCGTGACATTAAG-3′ and reverse,
5′-TGTGTTGGCGTACAGGTCTTTG-3′. Expression levels were quantified
using the 2−ΔΔCq method (10) and normalized to the β-actin loading
control.
Cell culture
MCF-10A (normal human mammary epithelial cell line)
and four breast cancer cell lines, including MCF-7, SK-BR-3,
MDA-MB-231 and MDA-MB-468, were purchased from The Cell Bank of
Type Culture Collection of the Chinese Academy of Sciences. These
cell lines were cultured in DMEM (Gibco; Thermo Fisher Scientific,
Inc.) containing 10% FBS (Gibco; Thermo Fisher Scientific, Inc.) at
37°C in the presence of 5% CO2.
HEIH small interfering (si)RNA
transfection
Briefly, 50 nM HEIH siRNA or negative control (NC)
siRNA (Guangzhou RiboBio Co., Ltd.) sequences were synthesized and
transfected into 1×105 MDA-MB-231 cells/well seeded into
12-well plates. Transfection was achieved using Lipofectamine™ 2000
(Guangzhou RiboBio Co., Ltd.). The transfection was performed for
24 h at 37°C, and then used for subsequent experiments. The
transfection efficiency was determined by measuring the HEIH
expression via RT-qPCR. The siRNA segment sequences were obtained
by a previous study (9).
Cell viability assay
Cell viability was confirmed by the CellTiter
96® AQueous One Solution Cell Proliferation assay kit
(MTS; Promega Corporation). The transfected cells were seeded in
96-well plates at a concentration of 5×104 cells in each
well. Following incubation for 24 h, 20 µl reagent was added into
each well and the cells were subsequently incubated at 37°C for 2
h. The cell viability was determined by the OD value at 490 nm. The
detailed protocol was performed following the manufacturer's
instructions as described previously (11).
5-Ethynyl-20-deoxyuridine (EdU)
incorporation assay
In addition to the cell viability assay, the EdU
incorporation assay kit (Guangzhou RiboBio Co., Ltd.) was used to
assess the proliferative activity of the cells. Briefly, the
transfected cells were seeded in 96-well plates at a density of
1×104 cells/well, following incubation for 24 h at 37°C.
A total of 100 µl medium containing EdU solution (50 µM) was added
into each well. The cells were incubated for 3 h and
paraformaldehyde (4%) with Triton X-100 (0.5%) was added to the
cells. Moreover, the cell nuclei were stained with DAPI for 15 min
at room temperature. The ratio of EdU positive cells (green cells)
to total DAPI positive cells (blue cells) corresponded to
proliferative activity.
Western blot analysis
Total protein was extracted using NP-40 lysis buffer
(Beyotime Institute of Biotechnology). Total protein was quantified
using a BCA protein quantification kit (cat. no. P0012S; Beyotime
Institute of Biotechnology) and 20 µg protein/lane was resolved via
10% SDS-PAGE and electroblotted onto nitrocellulose membranes.
Membranes were blocked with 5% non-fat milk solution at room
temperature for 1.5 h, followed by overnight incubation with
primary antibodies at 4°C. The blots were subsequently incubated
with horseradish peroxidase-conjugated goat anti-mouse or goat
anti-rabbit secondary antibodies (1:5,000; Abcam; cat. nos.
ab205718 and ab205719) at room temperature for 1 h. The following
primary antibodies (all purchased from Abcam) were used: Bcl-2
(1:2,000; cat. no. ab692), Bax (1:2,000; cat. no. ab32503),
E-cadherin (1:1,000; cat. no. ab40772), N-cadherin (1:1,000; cat.
no. ab18203), Vimentin (1:1,000; cat. no. ab92547), matrix
metallopeptidase 9 (MMP-9; 1:1,000; cat. no. ab137867) and β-actin
(1:2,000; cat. no. ab8227).
Caspase-3 activity assay
A Caspase-3 Activity kit (Beyotime Institute of
Biotechnology; cat. no. C1168M) was used to determine caspase-3
activity. Total cellular protein was obtained using a lysis buffer.
A total of 40 µg protein was diluted to a 50-µl final solution that
was subsequently mixed with 75 µl caspase-3 substrate and incubated
for 3 h. The hydrolysis of Ac-DEVD-pNA resulted in caspase-3
released free pNA (yellow formazan product) that was detected at
405 nm. Caspase-3 activity was expressed as the fold of enzyme
activity compared with that of the synchronized cells.
Flow cytometry assay
The apoptotic assays were performed using an Annexin
V-FITC/PI Apoptosis Detection kit (Miltenyi Biotec GmbH), according
to the manufacturer's protocols. The apoptotic rate was calculated
using both early and late apoptotic cells. Briefly,
1×106 cells were washed using 500 µl binding buffer,
centrifuged at 560 × g, and stained with 10 µl Annexin V-FITC
solution at room temperature for 30 min. A total of 5 µl PI
solution was added to each sample and incubated at room temperature
for 15 min. The apoptotic rate was evaluated using a BD FACSCalibur
flow cytometer (BD Biosciences) and ModFit LT software version 2.0
(Verity Software House, Inc.). At least 10,000 events were analyzed
for each sample. The analysis was repeated in three cell
samples.
Transwell assay
MDA-MB-231 cells (5×104) were seeded in
triplicate onto a 24-well plate with Transwell Boyden chambers (BD
Biosciences) coated with Matrigel (37°C for 30 min) or without
Matrigel for the invasion and migration assay, respectively. The
upper chamber contained serum-free DMEM, and the lower chamber
contained DMEM supplemented with 10% FBS as a chemoattractant.
Cells were incubated for 36 h at 37°C, fixed with 4%
paraformaldehyde at room temperature for 15 min and then stained
with 0.5% crystal violet for 5 min at room temperature. Stained
cells were visualized in five randomly selected fields using a
light IX71 inverted research microscope (magnification, ×200;
Olympus Corporation).
Scratch assay
MDA-MB-231 cells (5×104) were grown to
90–100% confluence in 6-well culture plates. A scratch on the cell
layer was created using a 200 µl pipette tip. Cells were cultured
in serum-free DMEM, and photographed immediately (0 h) after
scratching and 24 h after scratching. The scratch closure was
monitored using a light IX71 inverted research microscope
(magnification, ×200; Olympus Corporation). An ocular ruler was
used to verify the scratch sizes.
Statistical analysis
A paired Wilcoxon signed-rank test was employed to
evaluate significant differences of HEIH expression in breast
cancer tissues compared with adjacent normal tissues. Associations
between HEIH expression and clinical features were performed by a
Pearson's χ2 test. The optimal cut-off value of HEIH
expression in tumor/normal tissues was determined by a receiver
operating characteristic (ROC) curve analysis. The survival curve
was evaluated by the Kaplan-Meier method and the log-rank test was
used to compare differences between groups. The prognostic value of
HEIH expression was assessed based on the high-HEIH vs. low-HEIH
expression categorical definition. Hazard ratios (HRs) and 95%
confidence intervals (95% CIs) were calculated with the Cox
proportional hazard regression model and the data were adjusted for
the clinicopathological prognostic factors. All the statistical
tests were two-sided and considered significant when the P-value
was <0.05. The data in Table I
are presented as number (percentage). The cell experiment data were
analyzed by one-way ANOVA with a Dunnett's post hoc test for
multiple comparisons (≥3 groups). An unpaired Student's t-test was
used to analyze the statistical differences between two groups,
except for Fig. 1A, which was
analyzed using a paired Student's t-test. Data are presented as the
mean ± SEM. P<0.05 was considered to indicate a statistically
significant difference.
Results
Upregulation of HEIH expression is
positively associated with malignancy status and poor disease
prognosis
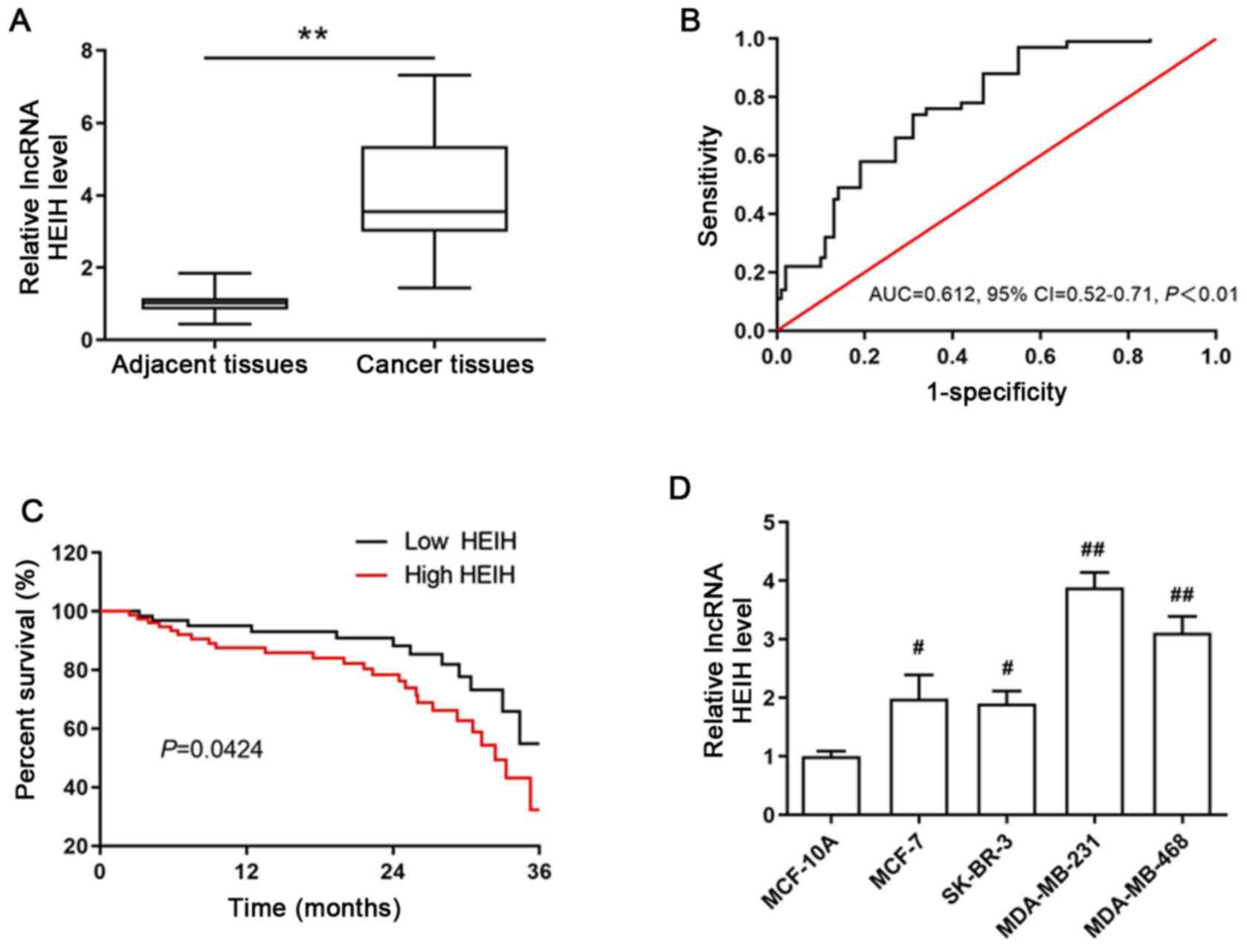
To determine the aberrant expression of HEIH in
breast cancer, the expression levels of HEIH in cancer tissues and
paired adjacent normal tissues from 160 patients with breast cancer
were measured by RT-qPCR. HEIH was upregulated in breast cancer
tissues (Fig. 1A). The area under
the ROC was 0.612 (95% CI, 0.52-0.71; P<0.01; Fig. 1B). In order to explore the
association of HEIH expression with the clinical characteristics of
patients with breast cancer, they were divided into high or low
expression groups according to the optimal cut-off value (0.43) of
fold-expression change of HEIH. The results of the Pearson's
χ2 analysis between HEIH expression and patient clinical
characteristics indicated that high HEIH expression levels were
associated with TNM stage and lymph node metastasis, whereas this
type of association was not noted for patient age, family history,
tumor grade, ER, PR and HER2 status (Table I).
In addition, the 3-year overall survival of patients
was estimated using Kaplan-Meier analysis by the log-rank test. The
data indicated that HEIH expression was negatively associated with
3-year overall survival and that high HEIH expression levels were
positively associated with a poor prognosis of patients with breast
cancer (log rank=4.118, P=0.0424; Fig.
1C). This finding was also confirmed by univariate analysis for
the significant factors described in Table II. HEIH expression was an
independent prognostic factor of the 3-year overall survival in
patients with breast cancer (HR, 4.97, 95% CI, 3.19-9.66;
P<0.001). Taken together, these results suggested that HEIH
expression is positively associated with malignancy status and poor
prognosis.
| Table II.Univariate and multivariate analysis
of clinical characteristics of overall survival using Cox
regression model. |
Table II.
Univariate and multivariate analysis
of clinical characteristics of overall survival using Cox
regression model.
|
| Univariate
analysis | Multivariate
analysis |
|---|
|
|
|
|
|---|
|
Characteristics | HR (95% CI) | P-value | HR (95% CI) | P-value |
|---|
| Age | 1.51
(0.88-2.60) | 0.212 | 1.34
(0.70-2.25) | 0.375 |
| Family history | 1.34
(0.51-3.44) | 0.513 |
|
|
| Tumor grade | 1.89
(1.03-3.44) | 0.032 | 1.70
(0.93-3.11) | 0.082 |
| TNM stage | 2.42
(1.41-4.20) | 0.004 | 2.31
(1.34-4.02) | 0.006 |
| Lymph node
metastasis | 1.98
(1.11-3.62) | 0.017 | 1.23
(0.37-3.86) | 0.672 |
| ER status | 0.74
(0.36-1.19) | 0.323 |
|
|
| PR status | 0.77
(0.38-1.21) | 0.513 |
|
|
| HER2 status | 0.81
(0.42-1.39) | 0.472 |
|
|
| HEIH
expression | 5.19
(3.21-9.84) | <0.001 | 4.97
(3.19-9.66) | <0.001 |
Proliferation of MDA-MB-231 cells can
be inhibited by HEIH knockdown
The aforementioned results allowed the determination
of HEIH expression in breast cancer cells. The expression levels of
HEIH were upregulated in four breast cancer cell lines (MCF-7,
SK-BR-3, MDA-MB-231 and MDA-MB-468), which was most notable in
MDA-MB-231 cells (Fig. 1D).
Therefore, a functional study was performed in MDA-MB-231 cells.
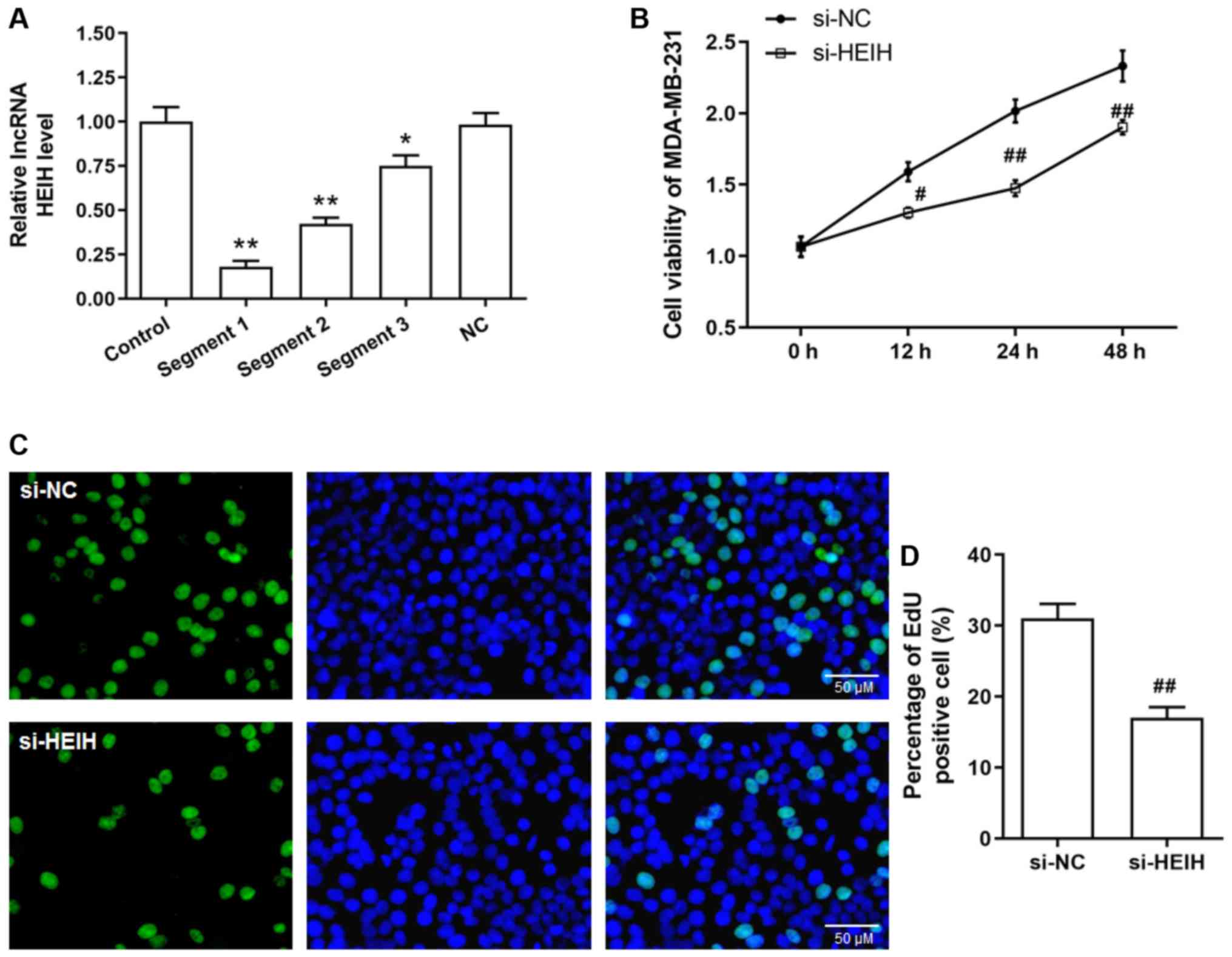
Initially, siRNA sequences were transfected into MDA-MB-231 cells
in order to achieve HEIH knockdown. Among the three siRNA sequences
tested, the first sequence indicated the maximum interference
efficiency and was used for the following studies (Fig. 2A). Following transfection of the
siRNAs into cells, cell viability and EdU-positive cell percentage
were decreased in MDA-MB-231 cells, suggesting that HEIH knockdown
inhibited the proliferation of breast cancer cells.
Apoptosis of MDA-MB-231 cells is
promoted by HEIH knockdown
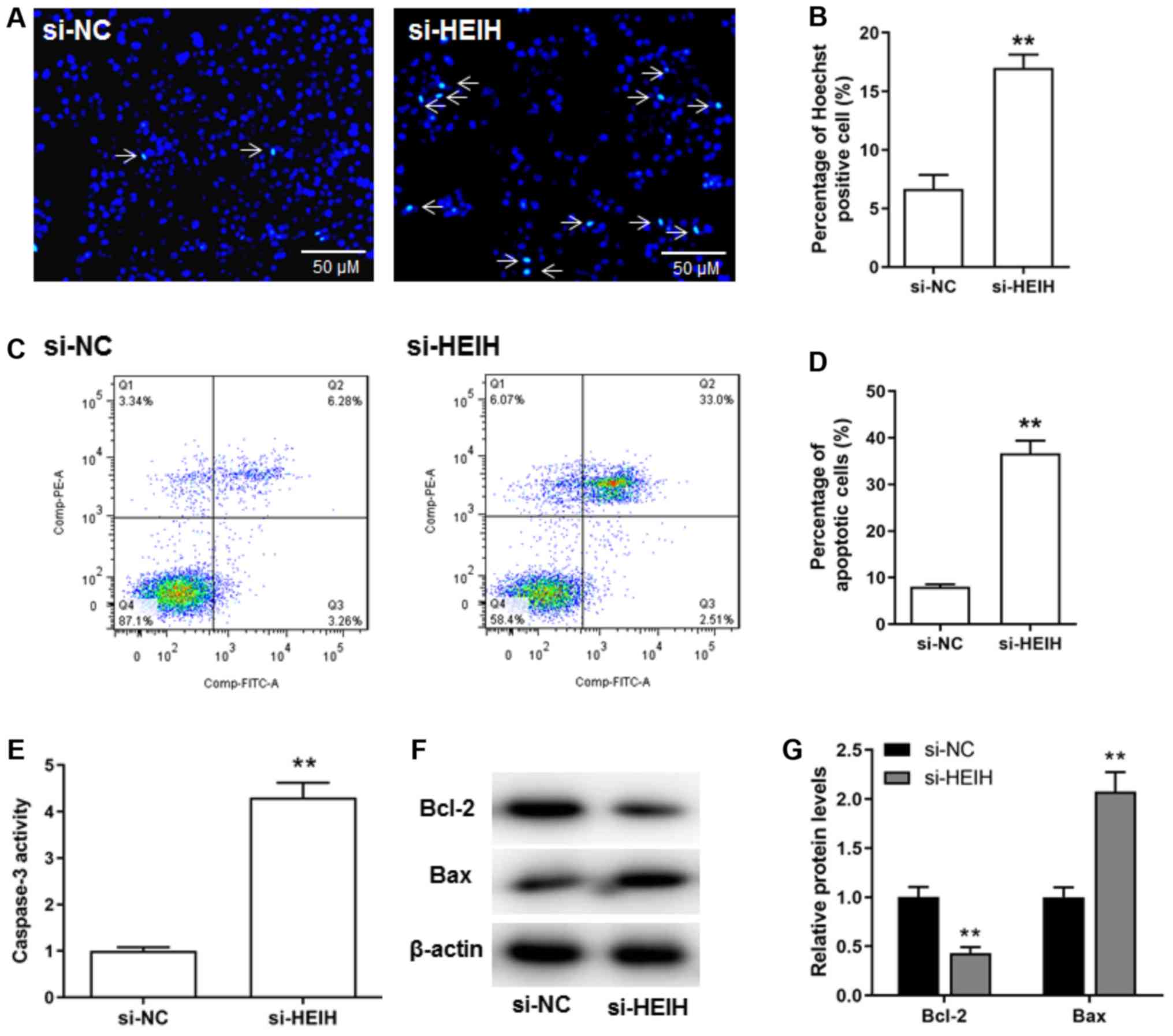
The effects of HEIH knockdown on breast cancer cell
apoptosis were investigated. The percentage of Hoechst positive
cells was increased following HEIH knockdown (Fig. 3A and B). The results of flow
cytometry confirmed that HEIH knockdown increased the number of
apoptotic cells (Fig. 3C and D).
Moreover, upregulation of Bax protein expression and caspase-3
activity, as well as downregulation of Bcl-2 protein expression
could be observed following HEIH knockdown in MDA-MB-231 cells
(Fig. 3E-G). Taken collectively,
the data suggested that HEIH knockdown promoted the apoptosis of
breast cancer cells.
MDA-MB-231 cell metastasis can be
inhibited by HEIH knockdown
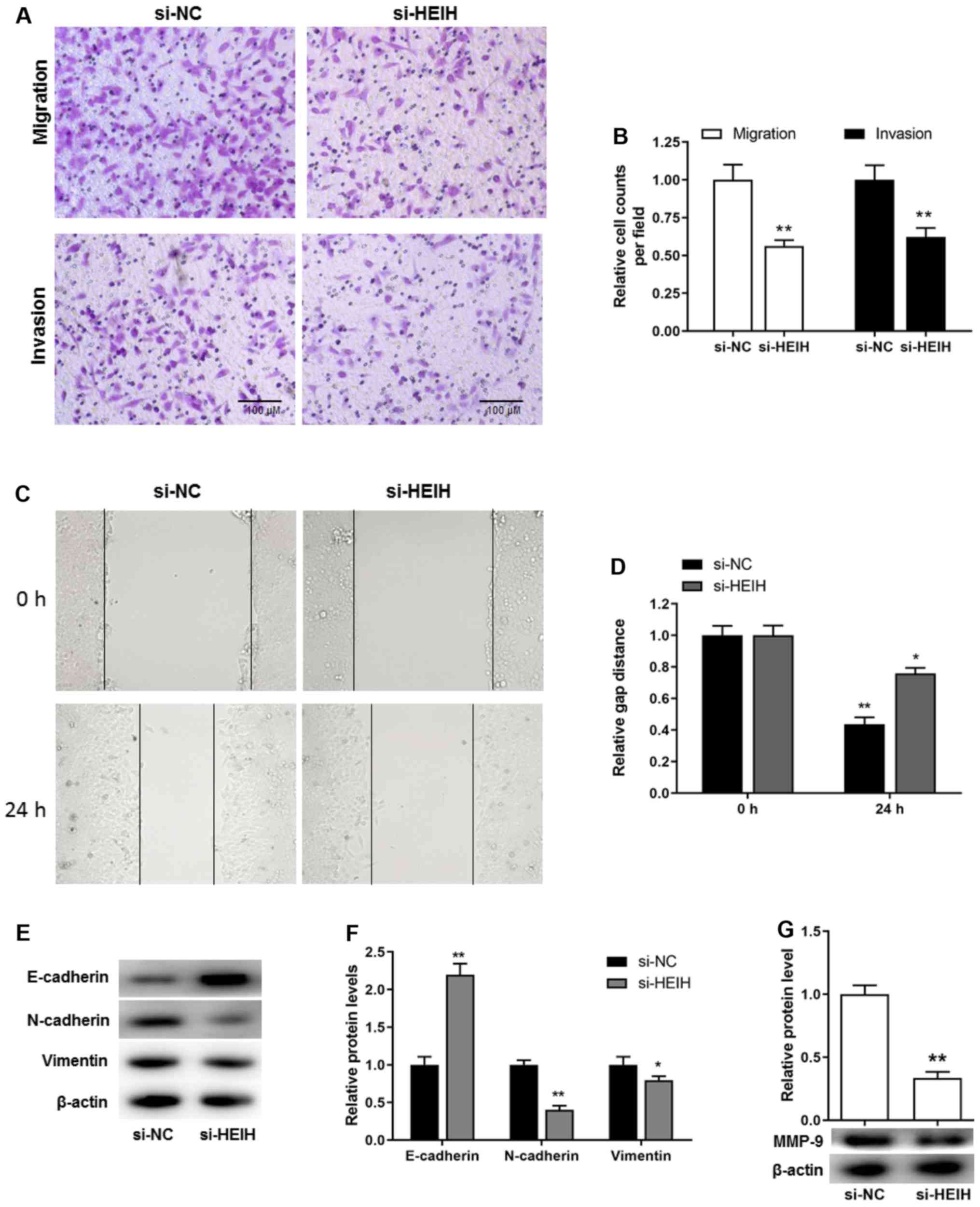
In addition to the proliferative potential and
apoptosis resistance, cell metastasis is considered a key
pathological feature during cancer development. Transwell assays,
including migration and invasion assays, were used to investigate
cell motility. The cell number, which corresponded with the cells
passing through the membrane, was decreased following HEIH
knockdown in MDA-MB-231 cells. This was determined by migration and
invasion assays (Fig. 4A and B).
In order to exclude that these effects were attributed to decreased
proliferation and/or increased apoptosis, a scratch assay was
performed to further investigate cell motility. The results
demonstrated that the distance covering the wound in si-HEIH cells
was higher than that noted in si-NC cells, suggesting that HEIH
knockdown decreased cell motility (Fig. 4C and D). Furthermore, considering
the key role of the epithelial-mesenchymal transition (EMT) in the
development of metastasis, the expression levels of EMT-related
proteins were determined following HEIH knockdown. HEIH knockdown
upregulated E-cadherin protein expression, whereas it downregulated
the expression of N-cadherin, Vimentin and MMP-9 proteins (Fig. 4E-G). These results indicated that
HEIH knockdown inhibited metastasis of breast cancer cells.
Discussion
As an intermediate between DNA and protein, RNA
transmits genetic information that is translated into various
biological processes. However, these intermediate RNAs (mRNAs)
compose <3% of human DNA, as demonstrated from the development
of human genome sequencing and the discovery of ncRNAs. Emerging
evidence suggests that ncRNAs are key RNA molecules that play
prominent roles in cell biology (12). According to their size, ncRNAs can
be classified into small ncRNAs (<200 nt in length) and lncRNAs
(≥200 nt in length) (4). The
microRNAs are considered one of the well-known small ncRNAs that
have been reported to regulate carcinogenesis (13–15).
The investigation of the expression levels of ncRNAs can be used
for diagnostic and prognostic purposes in cancer. In recent years,
the investigation of the role of lncRNAs in cancer has become a
research hot spot due to their functional relevance in
physiological and pathological processes involved in this disease
(4). An increasing number of
lncRNAs are aberrantly expressed in breast cancer (16–18).
Previously, Yang et al (19) sequenced >1,300 lncRNAs that were
aberrantly expressed in breast cancer. Notably, the lncRNA with the
highest upregulation was identified as AFAP1-AS1 (19). Our previous study further
investigated the functional role of AFAP1-AS1 in breast cancer and
found its diagnostic and prognostic value for this disease
(20). Similarly, in the present
study it was reported that the expression levels of HEIH were
upregulated in breast cancer tissues. Moreover, high HEIH
expression levels were shown to be positively associated with the
malignancy status, including TNM stage and lymph node metastasis,
which was also indicative of poor disease prognosis. However,
assessing patient survival from the first three years may not be
enough to support the aforementioned conclusion. The follow-up of
patients with breast cancer from the present study is still
on-going, and future studies would assess overall survival over a
longer time period.
It has been previously reported that lncRNA HEIH is
upregulated in hepatocellular carcinoma, indicating poor disease
outcome of patients with hepatocellular carcinoma (5,6).
Moreover, previous studies have proposed that it can promote cell
cycle progression of hepatocellular carcinoma cells (5,6).
Upregulation of HEIH was further observed in colorectal cancer
(7), melanoma (8) and non-small cell lung cancer
(9). The expression levels of HEIH
are significantly increased in colorectal cancer tissues, and HEIH
expression is positively associated with malignancy status and poor
prognosis in patients with colorectal cancer. Furthermore,
upregulation of HEIH has been reported to upregulate colorectal
cancer cell proliferation and decrease apoptosis in vitro,
whereas it further accelerates colorectal cancer tumor growth in
vivo (7). Moreover, HEIH has
been demonstrated to be highly expressed in melanoma tissues and
cell lines and is associated with advanced clinical stage and poor
disease outcome in patients with melanoma. Knockdown of HEIH
inhibits melanoma cell proliferation, migration and invasion
(8). In non-small cell lung
cancer, it was revealed that HEIH accelerates cell proliferation
and metastasis (9). These studies
collectively suggest that HEIH serves as an oncogene in multiple
cancer types. The present study demonstrated that HEIH expression
was upregulated in breast cancer cell lines, and HEIH knockdown
inhibited proliferation and induced apoptosis of breast cancer
cells, suggesting that it acted as an oncogene in breast
cancer.
Among the four breast cancer cell lines examined,
upregulation of HEIH in MDA-MB-231 and MDA-MB-468 cells was higher
than that noted in other types of breast cancer cell lines (MCF-7
and SK-BR-3). The highest expression was found in MDA-MB-231 cells.
MDA-MB-231 and MDA-MB-468 cells belong to triple-negative breast
cancer (TNBC) cell lines. It is well known that TNBC is an
aggressive form of breast cancer that contains cells that
overexpress HER2. TNBC cells do not express ER or PR and are
consequently resistant to cancer therapy (21). The development of targeted therapy
(hormonal agents and trastuzumab) has made significant improvements
in the outcome of other subtypes of breast cancer, including
ER-positive/HER2 overexpressing tumors. However, current treatments
for TNBC do not contain specific tumor-targeting therapeutic agents
(22). Moreover, TNBC easily
recurs and its metastasis leads to a poor prognosis during
treatment (23). Therefore, the
present study further explored the role of HEIH in the metastatic
activity of MDA-MB-231 cells. Although, it was demonstrated that
high HEIH expression was not associated with ER, PR and HER2
status, HEIH knockdown inhibited the invasion and migration of
MDA-MB-231 cells, suggesting the contribution of HEIH in TNBC
metastasis.
In conclusion, in the present study, the
upregulation of HEIH expression was confirmed in breast cancer
tissues. This lncRNA was positively associated with the malignancy
status and poor prognosis of patients with breast cancer.
Furthermore, HEIH knockdown induced apoptosis and suppressed
proliferation and metastatic activity in vitro. These
findings suggested the oncogenic role of HEIH in breast cancer,
which may be used as a novel diagnostic and prognostic indicator
for this disease.
Acknowledgements
Not applicable.
Funding
The present work was supported by the Scientific
Research Project of Gansu Provincial Health Commission (grant no.
GSWSKY-2019-02) and Gansu Provincial Natural Scientific Foundation
(grant no. 18JR3RA352).
Availability of data and materials
All data generated or analyzed during this study are
included in this published article.
Authors' contributions
CC and LW designed the study; CG, QR, FGD, QP and
YCN performed the experiments; DCM, QP and YCN performed the data
analysis; CC and LW wrote the manuscript; and CC, CG, QR and LW
revised the manuscript. All authors read and approved the final
manuscript.
Ethics approval and consent to
participate
All patients provided signed informed consent prior
to surgery. Clinicopathological follow-up information of patients
with breast cancer was available. All clinical investigations in
the present study were approved by the Human Ethics Committee of
the First Hospital of Lanzhou University (Lanzhou, China). The
study protocols were conducted according to the principles
described in the Declaration of Helsinki.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Gardezi SJS, Elazab A, Lei B and Wang T:
Breast cancer detection and diagnosis using mammographic data:
Systematic review. J Med Internet Res. 21:28692019. View Article : Google Scholar
|
|
2
|
Merino Bonilla JA, Torres Tabanera M and
Ros Mendoza LH: Breast cancer in the 21st century: From early
detection to new therapies. Radiologia. 59:368–379. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Kumar M, DeVaux RS and Herschkowitz JI:
Molecular and cellular changes in breast cancer and new roles of
lncRNAs in breast cancer initiation and progression. Prog Mol Biol
Transl Sci. 144:563–586. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Bhan A, Soleimani M and Mandal SS: Long
noncoding RNA and cancer: A new paradigm. Cancer Res. 77:3965–3981.
2017. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Yang F, Zhang L, Huo XS, Yuan JH, Xu D,
Yuan SX, Zhu N, Zhou WP, Yang GS, Wang YZ, et al: Long noncoding
RNA high expression in hepatocellular carcinoma facilitates tumor
growth through enhancer of zeste homolog 2 in humans. Hepatology.
54:1679–1689. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Ma Y, Cao D, Li G, Hu J, Liu X and Liu J:
Silence of lncRNA HEIH suppressed liver cancer cell growth and
metastasis through miR-199a-3p/mTOR axis. J Cell Biochem.
120:17757–17766. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Cui C, Zhai D, Cai L, Duan Q, Xie L and Yu
J: Long noncoding RNA HEIH promotes colorectal cancer tumorigenesis
via counteracting miR-939mediated transcriptional repression of
Bcl-xL. Cancer Res Treat. 50:992–1008. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Zhao H, Xing G, Wang Y, Luo Z, Liu G and
Meng H: Long noncoding RNA HEIH promotes melanoma cell
proliferation, migration and invasion via inhibition of
miR-200b/a/429. Biosci Rep. 37:BSR201706822017. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Jia K, Chen F and Xu L: Long noncoding RNA
HEIH promotes the proliferation and metastasis of non-small cell
lung cancer. J Cell Biochem. 120:3529–3538. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Li WQ, Li XH, Wu YH, Du J, Wang AP, Li D
and Li YJ: Role of eukaryotic translation initiation factors 3a in
hypoxia-induced right ventricular remodeling of rats. Life Sci.
144:61–68. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Ling H, Vincent K, Pichler M, Fodde R,
Berindan-Neagoe I, Slack FJ and Calin GA: Junk DNA and the long
non-coding RNA twist in cancer genetics. Oncogene. 34:5003–5011.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Ling H, Krassnig L, Bullock MD and Pichler
M: MicroRNAs in testicular cancer diagnosis and prognosis. Urol
Clin North Am. 43:127–134. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Thomas J, Ohtsuka M, Pichler M and Ling H:
MicroRNAs: Clinical relevance in colorectal cancer. Int J Mol Sci.
16:28063–28076. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Troppan K, Wenzl K, Deutsch A, Ling H,
Neumeister P and Pichler M: MicroRNAs in diffuse large B-cell
lymphoma: Implications for pathogenesis, diagnosis, prognosis and
therapy. Anticancer Res. 34:557–564. 2014.PubMed/NCBI
|
|
16
|
Youness RA and Gad MZ: Long non-coding
RNAs: Functional regulatory players in breast cancer. Noncoding RNA
Res. 4:36–44. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Xu N, Wang F, Lv M and Cheng L: Microarray
expression profile analysis of long non-coding RNAs in human breast
cancer: A study of Chinese women. Biomed Pharmacother. 69:221–227.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Shen X, Xie B, Ma Z, Yu W, Wang W, Xu D,
Yan X, Chen B, Yu L, Li J, et al: Identification of novel long
non-coding RNAs in triple-negative breast cancer. Oncotarget.
6:21730–21739. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Yang F, Lyu S, Dong S, Liu Y, Zhang X and
Wang O: Expression profile analysis of long noncoding RNA in
HER-2-enriched subtype breast cancer by next-generation sequencing
and bioinformatics. OncoTargets Ther. 9:761–772. 2016. View Article : Google Scholar
|
|
20
|
Ma D, Chen C, Wu J, Wang H and Wu D:
Up-regulated lncRNA AFAP1-AS1 indicates a poor prognosis and
promotes carcinogenesis of breast cancer. Breast Cancer. 26:74–83.
2019. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Park SY, Choi JH and Nam JS: Targeting
cancer stem cells in triple-negative breast cancer. Cancers
(Basel). 11:9652019. View Article : Google Scholar
|
|
22
|
McCann KE, Hurvitz SA and McAndrew N:
Advances in targeted therapies for triple-negative breast cancer.
Drugs. 79:1217–1230. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Saleh R, Taha RZ, Sasidharan Nair V,
Alajez NM and Elkord E: PD-L1 blockade by atezolizumab
downregulates signaling pathways associated with tumor growth,
metastasis, and hypoxia in human triple negative breast cancer.
Cancers (Basel). 11:10502019. View Article : Google Scholar
|