Introduction
Lung cancer is one of the leading causes of
cancer-related mortality in China (1). Among the two major lung cancer
subtypes, non-small-cell lung cancer (NSCLC) accounts for ≥80% of
all lung cancer cases (2). Over
the past several years, surgical resection has remained the main
choice of treatment for patients with NSCLC, but the survival rate
of these patients remains ≤18% due to rapid tumor metastasis
(3,4). Therefore, there is a pressing need to
identify new highly sensitive biomarkers and targets for
therapeutic intervention to aid in the diagnosis of NSCLC and
improve patient outcomes.
MicroRNAs (miRNAs/miRs) are single-stranded
non-coding RNAs that negatively regulate gene expression by binding
to the 3′-untranslated region (UTR) of their target gene mRNAs at
the post-transcriptional level (5). Previous studies have revealed that
miRNAs are crucial for various biological and pathological
processes, and aberrant expression or function of miRNAs are
commonly observed in various types of cancer, including NSCLC
(6–8). It has been shown that the
overexpression of miR-30e could inhibit cell viability and invasion
by targeting SRY-box transcription factor 9 in NSCLC cells
(9). Liu et al (10) also revealed that miR-661 promoted
NSCLC viability, migration and metastasis by targeting
retinoblastoma 1. However, the role of miRNAs in NSCLC and the
molecular mechanisms by which they affect this type of cancer
remain largely unknown.
miR-92a, a putative oncogene, is frequently
upregulated in numerous types of human cancer (11). For example, Yu et al
(12) demonstrated that
upregulation of miR-92a promoted the viability, migration and
invasion of osteosarcoma cells. Shigoka et al (13) reported that miR-92a was upregulated
in hepatocellular carcinoma (HCC) and overexpression of miR-92a
enhanced HCC cell viability. A recent study also confirmed the
oncogenic role of miR-92a in colorectal cancer (CRC), and revealed
its pro-proliferative and anti-apoptotic functions in CRC cells
(14). Jiang et al
(15) demonstrated that
overexpression of miR-92a promoted osteosarcoma tumor growth in
vivo. In a previous study, miR-92a was more highly expressed in
NSCLC tumor tissues than in adjacent tissue samples and plasma from
healthy donors (16). However, the
mechanisms by which miR-92a affects NSCLC tumorigenesis remains
unclear. Therefore, the aim of the present study was to examine the
potential molecular mechanisms by which miR-92a affects the
pathogenesis of NSCLC cells by examining miR-92a expression in
NSCLC tissues and cell lines. The regulatory role of miR-92a in
NSCLC cell viability and apoptosis, and the relevant mechanisms by
which it affects NSCLC cells, were also investigated.
Materials and methods
Patients and sample collection
The present study was approved by the Research
Ethics Committee of Shanghai Ninth People's Hospital, Shanghai
JiaoTong University School of Medicine (Shanghai, China). All
patients provided written, informed consent. Paired NSCLC tissues
and adjacent normal tissues were obtained from 15 patients (9 men,
6 women; age, 60.3±8.5 years) during surgery between January and
June 2018. None of the patients with NSCLC received treatment
before surgery. In addition, SCLC tissues and adjacent normal
tissue were collected from 15 patients (8 men, 7 women; age,
62.1±9.2 years) before chemotherapy between August and November
2019. All samples were collected at the Department of Thoracic
Surgery, Shanghai Ninth People's Hospital, Shanghai JiaoTong
University School of Medicine, and both tumor and adjacent normal
tissues were confirmed by pathological examinations. Patients were
excluded if they had recurrent tumors or had primary tumors but
received chemoradiotherapy before surgical operation. Tissues were
immediately snap frozen in liquid nitrogen and stored at −80°C
prior to RNA extraction.
Cell lines and culture conditions
The four NSCLC cell lines (A549, H358, NCI-H520 and
H1299), a normal human bronchial epithelial cell line (16HBE) and
293T cells were purchased from The Cell Bank of Type Culture
Collection of the Chinese Academy of Sciences. All cells were
cultured in DMEM (Gibco; Thermo Fisher Scientific, Inc.)
supplemented with 10% fetal bovine serum (Invitrogen; Thermo Fisher
Scientific, Inc.), 100 U/ml penicillin and 100 mg/ml streptomycin,
in humidified air at 37°C with 5% CO2.
RNA extraction and reverse
transcription-quantitative PCR (RT-qPCR)
Total RNA was extracted from NSCLC tissues and cell
lines with TRIzol® reagent (Invitrogen; Thermo Fisher
Scientific, Inc.), according to the manufacturer's instructions.
For RT-qPCR, RNA was reverse transcribed to cDNA from 100 ng total
RNA using a RT kit (Tiangen Biotech Co., Ltd.), and qPCR was
performed with SYBRGreen (Tiangen Biotech Co., Ltd.). All protocols
were carried out according to the manufacturer's instructions. The
primer sequences are listed as follows: miR-92a, forward
5′-CCGCGTGCTGGGATTC-3′, reverse, 5′-TCCAGAAGGCTGCAAATGG-3′; U6,
forward 5′-CTCGCTTCGGCAGCACA-3′, reverse
5′-GTCATACTCCTGCTTGCTGAT-3′; F-box/WD repeat-containing protein 7
(FBXW7), forward 5′-GTCCCGAGAAGCGGTTTGATA-3′, reverse
5′-TGCTCAGGCACGTCAGAAAAG-3′; and GAPDH, forward
5′-GAAGATGGTGATGGGATTTC-3′, and reverse 5′-AACGCTTCACGAATTTGCGT-3′.
RT-qPCR was performed on a Step-One Plus Real-Time PCR system
(Applied Biosystems; Thermo Fisher Scientific, Inc.), and each
RT-qPCR reaction was performed in triplicate, including no-template
controls. The reaction was performed under the following
conditions: 95°C for 5 min, followed by 40 cycles at 95°C for 15
sec and 60°C for 50 sec, and a final extension at 72°C for 10 min.
The relative quantification of miR-92a and FBXW7 were normalized to
the expression of U6 and GAPDH, respectively using the
2−ΔΔCq method (17).
Cell transfection
The miR-92a mimics (5′-UAUUGCACUUGUCCCGGCCUGU-3′),
mimics negative control (NC; 5′-CGGTGUGUUCAGACUACCUGUUC-3′),
miR-92a inhibitor (5′-ACAGGCCGGGACAAGUGCAAUA-3′) and inhibitor NC
(5′-TAACACGTCTATACGCCCA-3′) were obtained from Guangzhou RiboBio
Co., Ltd. FBXW7 small interfering (si)RNA (si-FBXW7; sense,
5′-TAAAGAGTTGGCACTCTAT-3′ and antisense, 5′-ATAGAGTGCCAACTCTTTA-3′)
and corresponding NC siRNA (si-scramble; sense,
5′-TTCTCCGAACGTGTCACGT-3′ and antisense, 5′-ACGTGACACGTTCGGAGAA-3′)
were also purchased from Guangzhou RiboBio Co., Ltd.
Upon A549, H1299, H358 and NCI-H520 cells in
six-well plates reaching 80% confluence, 1×106
cells/well were transfected with the transfectants using
Lipofectamine® 2000 reagent (Invitrogen; Thermo Fisher
Scientific, Inc.), according to the manufacturer's protocol. A
final concentration of 50 nM miR-92a mimics, 100 nM mimics NC, 200
nM miR-92a inhibitor, 100 nM inhibitor NC, 100 nM si-FBXW7 or 100
nM si-Scramble were used for each transfection. A blank control
(untransfected cells) was set up for each transfection. Following
transfection at 37°C for 24 h, the transfection efficiency was
analyzed using RT-qPCR or western blotting.
Cell viability
The effect of miR-92a on NSCLC cell viability was
measured using an MTT assay. After transfection with the indicated
miRNA mimics, inhibitor or siRNA, 20 µl MTT solution
(Sigma-Aldrich; Merck KGaA) was added to each well
(2×105/well), and A549, H1299, H358 and NCI-H520 cells
were cultured for an additional 2 h at 24, 36 and 48 h
post-transfection. Then, 150 µl dimethyl sulfoxide was used to
dissolve the purple formazan. Subsequently, the absorbance of the
samples at 450 nm was detected using a microplate reader (Bio-Rad
Laboratories, Inc.).
Flow cytometry
At 24-h post-transfection, apoptotic cells were
detected using Annexin V/APC and propidium iodide (PI) apoptosis
detection kit I (BD Pharmingen; BD Biosciences), according to the
manufacturer's protocol. Briefly, NSCLC cells were collected after
centrifugation at 400 × g for 10 min at room temperature, and then
washed with cold PBS. Then, 1×105 cells/ml were stained
with binding buffer, containing 5 µl Annexin V-FITC and 10 µl PI at
4°C in the dark for 15 min. Following the incubation, cell
apoptosis was analyzed using a FACScan flow cytometer (BD
Biosciences) and CellQuest software version 3.3 (BD
Biosciences).
Luciferase reporter assay
The targets of miR-92a were predicted using
TargetScan 7.0 (http://www.targetscan.org) and miRanda (http://www.microrna.org). The 3′-untranslated region
(UTR) fragment of FBXW7 containing the putative wild-type (wt)
sequence was amplified by PCR. The amplified product was inserted
into the pGL3 luciferase reporter vector (Promega Corporation),
namely pGL-FBXW7-wt. The QuikChange Lightning Site-Directed
Mutagenesis kit (Stratagene; Agilent Technologies, Inc.) was used
to construct the miR-92a binding site mutation in the 3′-UTR of
FBXW7, according to the manufacturer's protocol; this construct was
named pGL-FBXW7-mutant (mut). A total of 2×105 293T
cells/well were seeded into 24-well plates and transfected with
miR-92a mimics or mimics NC, and 100 ng pGL-FBXW7-wt or
pGL-FBXW7-mut, together with 100 ng pRL-TK Renilla plasmids
(Promega Corporation) using Lipofectamine® 2000
(Invitrogen; Thermo Fisher Scientific, Inc.) at 37°C. A total of 48
h after transfection, the luciferase activity was determined using
a Dual-Luciferase Reporter Assay system (Promega Corporation),
according to the manufacturer's protocol. Relative luciferase
activity was normalized to Renilla luciferase activity.
Western blot analysis
Total protein was extracted from cells using RIPA
buffer (Sigma-Aldrich; Merck KGaA) with 1% phenylmethylsulfonyl
fluoride. The protein concentration was determined using a BCA
protein assay kit (Pierce; Thermo Fisher Scientific, Inc.). Total
protein samples (30 µg) were separated by SDS-PAGE on 8% gels and
transferred to a polyvinylidene fluoride membrane (EMD Millipore).
The membranes were blocked with 5% non-fat milk at 4°C overnight
and incubated with primary antibodies overnight at 4°C. Primary
antibodies against Bax (1:1,000; cat. no. sc-70408), Bcl-2
(1:1,000; cat. no. sc-7382), proliferating cell nuclear antigen
(PCNA; 1:1,000; cat. no. sc-9857) and β-actin (1:2,000; cat. no.
sc-8432) were purchased from Santa Cruz Biotechnology, Inc., while
cleaved caspase-3 (1:1,000; cat. no. 9661) and Ki-67 (1:1,000; cat.
no. 12075) were purchased from Cell Signaling Technology, Inc.
After washing with PBS, the membrane was incubated with
HRP-conjugated antibodies (1:2,000; cat. nos. ab205718 and ab97040;
Abcam) for 1 h at room temperature and the bands were detected with
an ECL Advance reagent (GE Healthcare). The intensity of the bands
of interest was analyzed with ImageJ software version 1.46
(National Institutes of Health).
Statistical analyses
SPSS 13.0 software (SPSS, Inc.) was used to analyze
the data. Data were expressed as the mean ± SD of three independent
experiments. Differences between two groups were analyzed by a
paired t-test when comparing paired NSCLC tissues and adjacent
normal tissues, and an unpaired t-test for comparing two groups in
other cases. Differences between multiple groups were analyzed by
one-way analysis of variance, followed by Tukey's post-hoc test.
Pearson's correlation analysis was carried out to determine the
association between miR-92a and FBXW7. P<0.05 was considered to
indicate a statistically significant difference.
Results
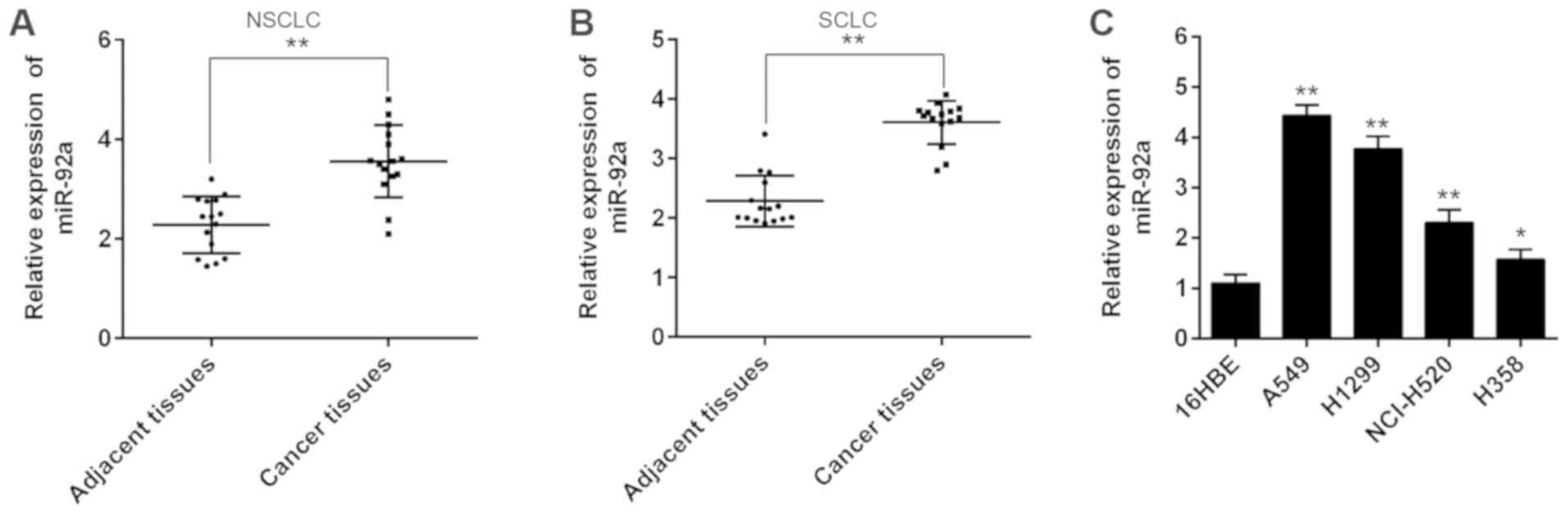
miR-92a is upregulated in NSCLC
tissues and cell lines
To investigate the potential involvement of miR-92a
in NSCLC development, RT-qPCR analysis was conducted to determine
miR-92a expression levels in 15 paired NSCLC and adjacent tissues.
As shown in Fig. 1A, the
expression levels of miR-92a were significantly upregulated in
NSCLC tissues compared with in adjacent tissues. In addition,
higher expression levels of miR-92a were also observed in SCLC
tissues compared with adjacent tissues (Fig. 1B), which is consistent with the
results of a previous study (18).
This analysis was also extended to four NSCLC cell lines; A549,
H1299, NCI-H520 and H358, with 16HBE as a control. Consistent with
the results of miR-92a expression levels in clinical tissues,
miR-92a expression was markedly increased in these cell lines
compared with in 16HBE cells (Fig.
1C). Together, these data suggested that miR-92a may be
involved in NSCLC progression.
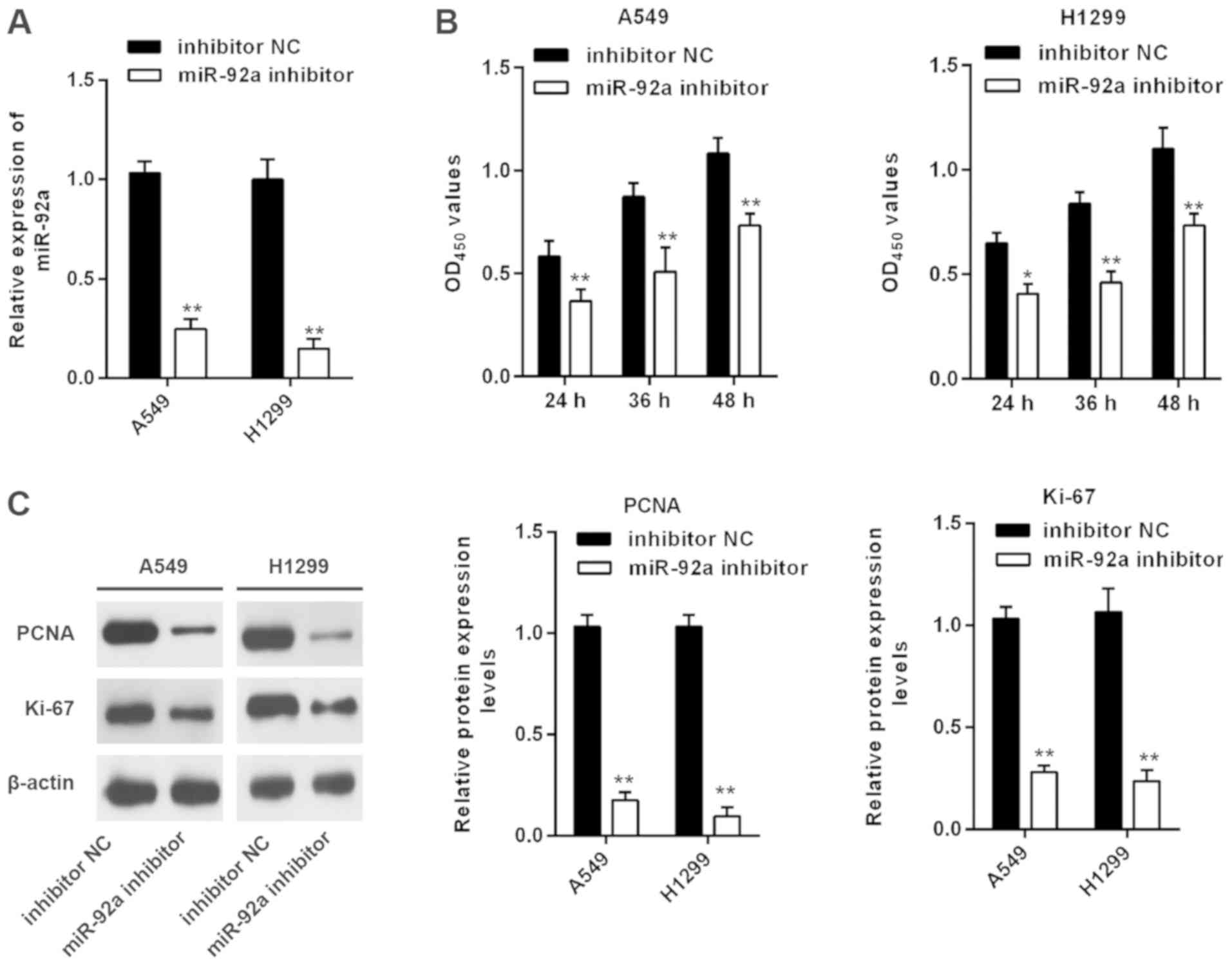
Inhibition of miR-92a suppresses the
viability of NSCLC cells
According to the aforementioned detection of miR-92a
expression in the four NSCLC cell lines, the highly metastatic A549
and H1299 cell lines, in which miR-92a was highly expressed, were
chosen to conduct loss-of-function experiments. miRNA transfection
efficiency was evaluated using RT-qPCR. As shown in Fig. 2A, miR-92a was significantly
decreased in A549 and H1299 cells post-transfection with the
miR-92a inhibitor. The biological effects of miR-92a inhibition on
regulating cell viability was then assessed using an MTT assay.
Inhibition of miR-92a significantly suppressed A549 and H1299 cell
viability compared with inhibitor NC-transfected cells (Fig. 2B). PCNA and Ki-67 expression may
reflect the degree of viability because their rate of synthesis is
directly associated with the rates of cellular proliferation and
DNA synthesis (19). Therefore, to
further confirm the miR-92a inhibitor-induced reduction in cell
viability, the protein expression levels of PCNA and Ki-67 were
examined. Western blot analysis indicated that miR-92a inhibition
resulted in a significant decrease in PCNA and Ki-67 protein
expression (Fig. 2C). Therefore,
these results demonstrated that inhibition of miR-92a could
suppress the viability of NSCLC cells.
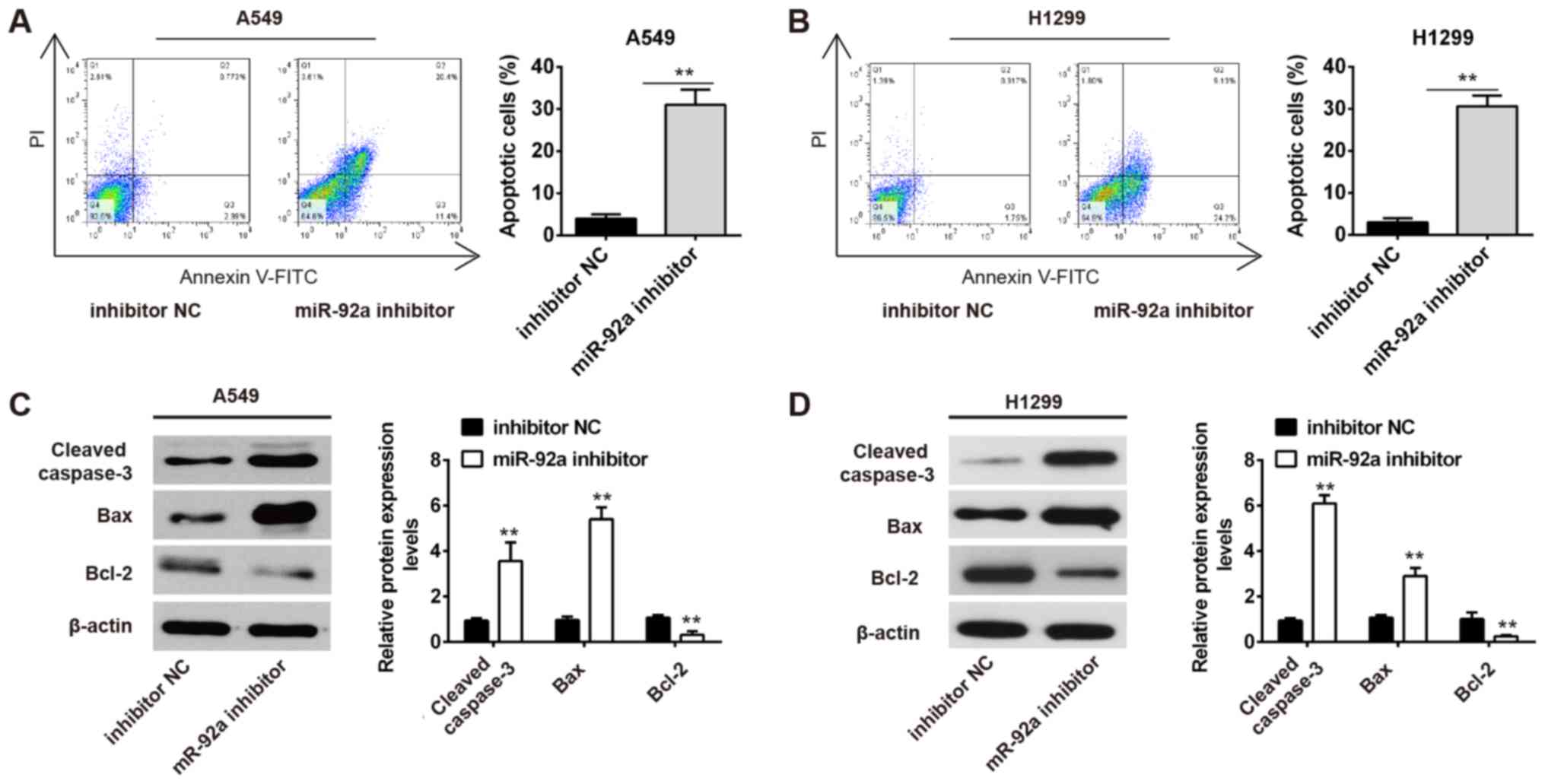
Inhibition of miR-92a promotes
apoptosis of NSCLC cells
To determine whether apoptosis inhibited growth, the
effects of miR-92a inhibitor on cell apoptosis were assessed. It
was demonstrated that miR-92a inhibition markedly promoted
apoptosis of A549 and H1299 cells compared in the inhibitor NC
group (Fig. 3A and B). In
addition, miR-92a inhibition significantly increased the expression
levels of pro-apoptotic proteins, cleaved caspase-3 and Bax, and
reduced the expression levels of the anti-apoptotic protein Bcl-2
(Fig. 3C and D). These data
suggested that miR-92a inhibition promoted apoptosis of NSCLC
cells.
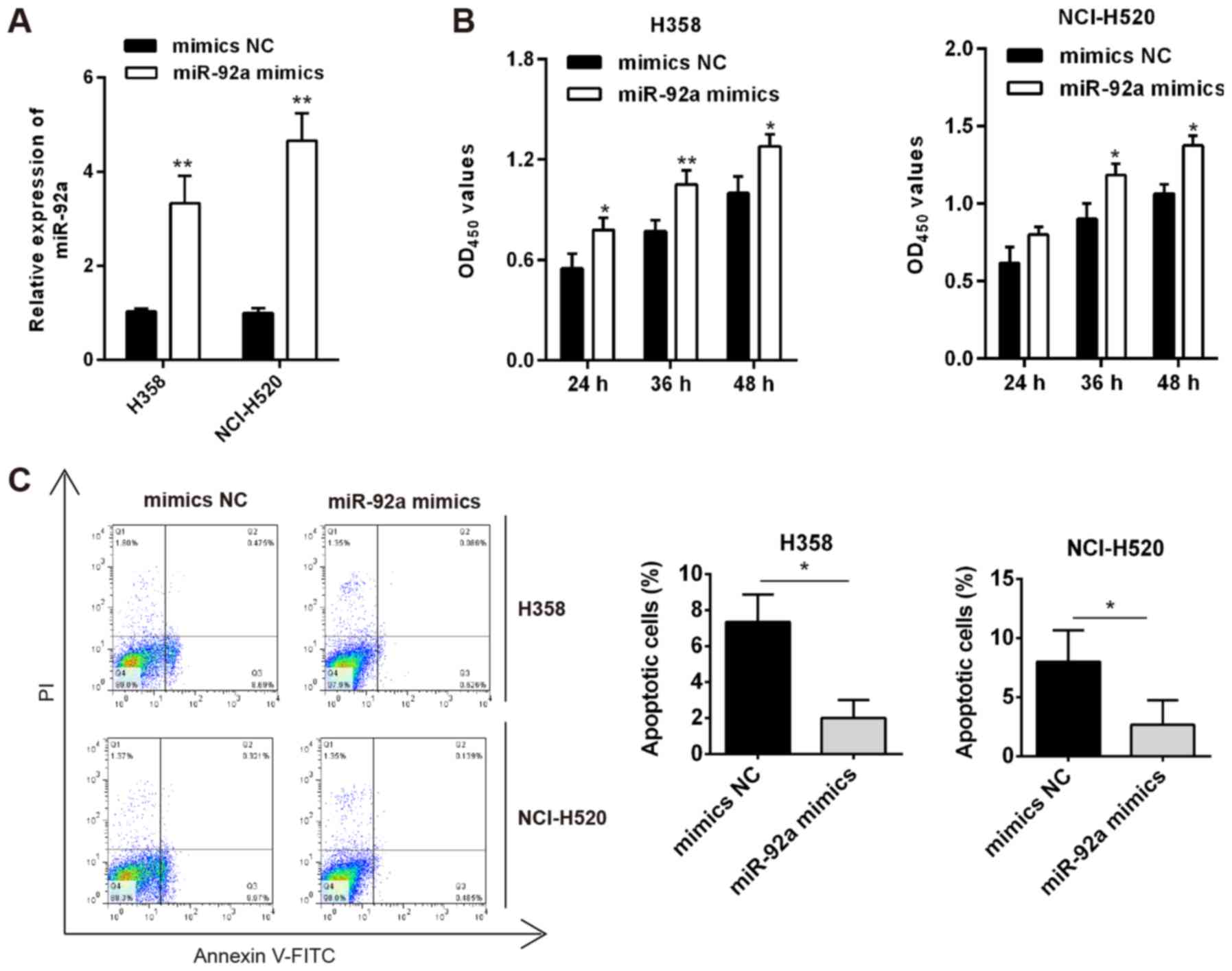
Overexpression of miR-92a inhibits the
viability and induces cell apoptosis of NSCLC cells
Next, weakly metastatic cells H358 and NCI-H520,
which expressed lower levels of miR-92a compared with the other
NSCLC cell lines, were used for gain-of-function experiments. As
shown in Fig. 4A, miR-92a was
significantly increased in H358 and NCI-H520 cells
post-transfection with miR-92a mimics. The MTT assay revealed that
miR-92a overexpression significantly promoted cell viability,
compared with that in mimics NC-transfected cells (Fig. 4B). Apoptosis of H358 and NCI-H520
cells was also markedly suppressed with miR-92a overexpression
(Fig. 4C). Collectively, miR-92a
overexpression may promote cell viability and suppress cell
apoptosis in vitro.
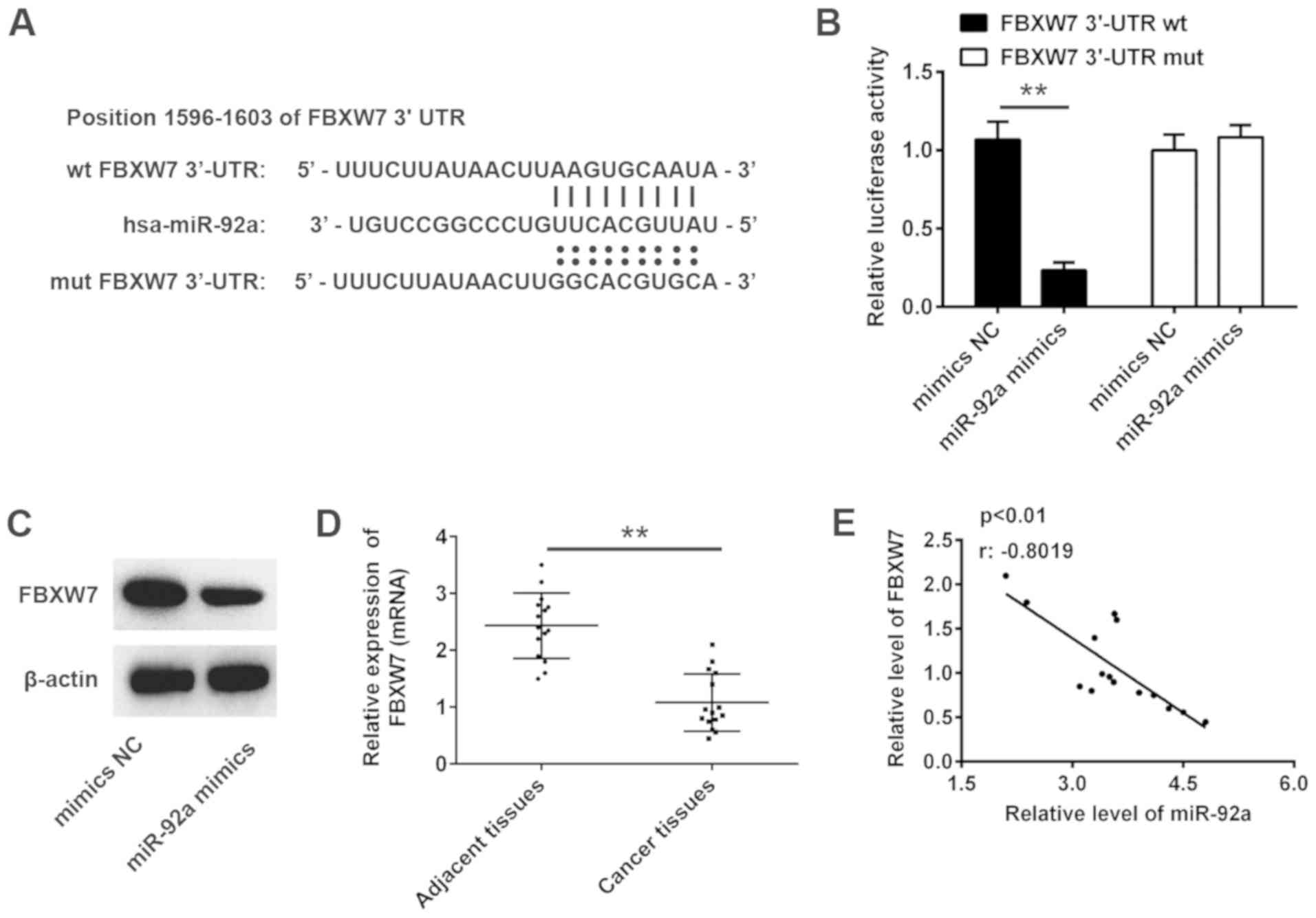
miR-92a directly targets FBXW7
To explore the molecular mechanism by which miR-92a
mediates its inhibitory effect on NSCLC, TargetScan 7.0 and miRanda
analyses were conducted to predict the target genes of miR-92a.
FBXW7, a well-known tumor suppressor, was identified as a potential
target of miR-92a, with the target site located in the 3′-UTR of
FBXW7 mRNA (Fig. 5A). To validate
whether FBXW7 is a direct target gene of miR-92a, the 3-UTR region
of FBXW7 was fused to a luciferase system. As shown in Fig. 5B, miR-92a mimics significantly
suppressed the luciferase activities of the 3′-UTR segment of
FBXW7, but not those of the construct containing a mut binding site
(mut FBXW7 3′-UTR), compared with the NC group.
Western blotting was conducted to further determine
whether miR-92a regulated the expression of FBXW7. The results
revealed that FBXW7 expression levels were markedly downregulated
by the miR-92a mimics (Fig. 5C).
The mRNA expression levels of FBXW7 were detected in 15 pairs of
NSCLC and adjacent tissues using RT-qPCR. The results revealed that
FBXW7 mRNA expression was significantly downregulated in NSCLC
tissues compared with in adjacent tissues (Fig. 5D). Further analyses indicated that
FBXW7 expression was negatively correlated with miR-92a expression
in NSCLC tissues (Fig. 5E). These
data suggested that miR-92a directly targets FBXW7 and suppresses
its expression in NSCLC cells.
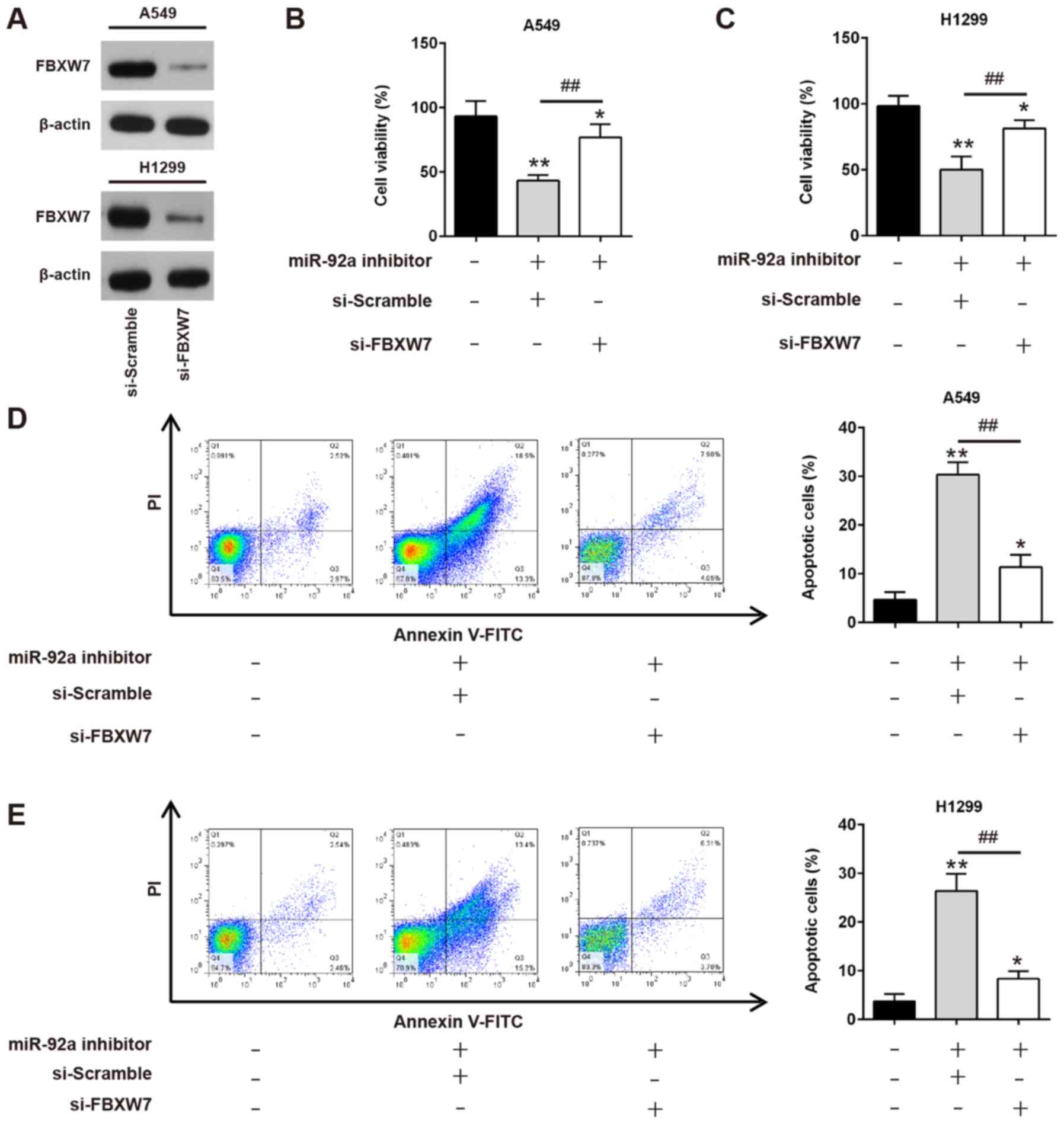
Downregulation of miR-92a inhibits
cell viability and promotes apoptosis by targeting FBXW7
As aforementioned, FBXW7 was a direct target of
miR-92a in NSCLC cells; therefore, it was further investigated
whether downregulation of miR-92a exerted its antiproliferative and
pro-apoptotic effects by upregulating FBXW7. A549 and H1299 cells
were co-transfected with siRNA-FBXW7 and miR-92a inhibitor, and
cell viability and apoptosis were evaluated. Firstly, siRNA-FBXW7
transfection efficiency was confirmed by western blotting. The
results revealed that siRNA-FBXW7 transfection markedly inhibited
FBXW7 levels compared with siRNA-scramble transfection in A549
cells and H1299 cells (Fig. 6A).
Compared with untransfected cells, miR-92a inhibitor significantly
reduced cell viability, whereas this inhibitory effect was
attenuated by siRNA-mediated FBXW7 knockdown (Fig. 6B and C). Meanwhile, siRNA-FBXW7
could significantly weaken apoptosis enhancement caused by miR-92a
inhibitor in A549 and H1299 cells (Fig. 6D and E). Collectively, these data
suggested that miR-92a downregulation may suppress NSCLC cell
growth and enhance apoptosis by targeting FBXW7.
Discussion
The present study demonstrated that miR-92a was
upregulated in NSCLC tissues and cell lines. Moreover, miR-92a
downregulation suppressed cell viability and promoted cell
apoptosis in vitro by targeting the tumor suppressor FBXW7.
These findings suggested that miR-92a may potentially provide a
novel strategy for the treatment of NSCLC.
Emerging evidence has indicated that aberrant
expression of miRNAs can affect tumorigenesis processes by
regulating key oncogenes or tumor suppressors (20); therefore, miRNAs may have great
potential as prognostic indicators and therapeutic targets
(21). Previous studies have
demonstrated that aberrant miRNA expression may contribute to NSCLC
progression (22,23). For example, Yang et al
(24) showed that miR-218 was
significantly downregulated in lung cancer tissues and was
associated with prognosis of patients with lung cancer. Zhuang
et al (25) reported that
serum levels of miR-484 could help screen patients for NSCLC with a
high sensitivity and specificity using receiver operating curve
analyses. In the present study, miR-92a was significantly
upregulated in NSCLC tissues and cell lines, which suggested that
miR-92a could potentially act as a new diagnostic marker and
therapeutic target in NSCLC treatment.
Previous studies have also examined the oncogenic
role of miR-92a in other types of human cancer (14,26,27).
For example, Ke et al (28)
showed that miR-92a promoted CRC cell metastasis through the
PTEN-mediated PI3K/AKT pathway. Chen et al (29) demonstrated that miR-92a promoted
the migration and invasion of human esophageal squamous cell
carcinoma by, at least partially, suppressing cadherin-1
expression. Notably, miR-92a has been observed to be highly
expressed in NSCLC tissues and cell lines (30,31),
which indicates that miR-92a may be involved in NSCLC progression.
Lu et al (32) revealed
that miR-92a regulated cell migration and invasion of NSCLC cells
by targeting PTEN; therefore, it was hypothesized that miR-92a may
affect NSCLC progression, which had not been previously
investigated. The results of the present study demonstrated that
miR-92a inhibition could suppress cancer cell viability (with
suppressed PCNA and Ki-67 protein expression) and promote apoptosis
(with increased caspase-3 and Bax, and decreased Bcl-2 protein
expression). Together, these results suggested that miR-92a may
function as an oncogene in NSCLC progression.
To elucidate the mechanisms by which NSCLC
progression is induced by miR-92a, bioinformatics analyses were
conducted to predict the putative targets of miR-92a, and FBXW7 was
identified as one of these potential targets. FBXW7 has been
reported to be a tumor suppressor in human tumorigenesis, with
marked effects on cell cycle progression, cell growth and invasion
(33,34). For example, Li et al
(35) revealed that upregulation
of FBXW7 attenuated osteosarcoma cell proliferation and its low
expression was associated with a worse patient outcome. Xiao et
al (36) also demonstrated
that FBXW7 knockdown enhanced NSCLC tumorigenesis and resistance to
gefitinib in a xenograft mouse model. Notably, a previous study
(15) showed that overexpression
of miR-92a promoted osteosarcoma growth by targeting FBXW7. Zhou
et al (37) also reported
that miR-92a was upregulated in cervical cancer, and promoted cell
proliferation and invasion by targeting FBXW7. Yang et al
(38) reported that miR-92a
promoted the tumor growth of HCC by targeting FBXW7. However, to
the best of our knowledge, whether FBXW7 is a functional target of
miR-92a in NSCLC cells has not been elucidated. In the present
study, FBXW7 was validated as a target gene of miR-92a in NSCLC
cells. In addition, the expression levels of FBXW7 in NSCLC tissues
were low, and negatively correlated with miR-92a expression.
Moreover, knockdown of FBXW7 in NSCLC cells attenuated the miR-92a
inhibitor-induced reduction in cell viability. This indicated that
an miR-92a inhibitor might inhibit NSCLC cell viability by
targeting FBXW7.
In conclusion, in the present study, miR-92a was
upregulated in NSCLC tissues and cell lines. Notably, miR-92a
inhibition suppressed cell viability and promoted cell apoptosis by
increasing FBXW7 expression in NSCLC cells. As a result, reducing
miR-92a expression could have important implications for clinical
management of NSCLC.
There are some limitations to the present study. For
example, the clinical sample size was relatively small, and a
larger sample size should be used in future studies to validate the
accuracy of miR-92a as a biomarker for NSCLC. Furthermore,
additional experiments in vivo should be performed to
confirm whether miR-92a may be utilized as a therapeutic target for
NSCLC. It has been shown that several targets can also be
controlled by miR-92a in NSCLC, such as PTEN and
double-strand-break repair protein rad21 homolog 1 (31,39),
which remains to be determined in future studies. Thus, further
research is required to gain deeper insight into these
mechanisms.
Acknowledgements
Not applicable.
Funding
No funding was received.
Availability of data and materials
All data generated or analyzed during this study are
included in this published article.
Authors' contributions
DN, JT, YC, ZZ and BZ performed the experiments,
contributed to data analysis and wrote the paper. ZY conceptualized
the study design, contributed to data analysis and experimental
materials. All authors read and approved the final manuscript.
Ethics approval and consent to
participate
All individuals provided written, informed consent
for the use of human specimens for clinical research. The present
study was approved by Shanghai Ninth People's Hospital, Shanghai
JiaoTong University School of Medicine.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Siegel R, Naishadham D and Jemal A: Cancer
statistics, 2013. CA Cancer J Clin. 63:2817–30. 2013. View Article : Google Scholar
|
|
2
|
Torre LA, Bray F, Siegel RL, Ferlay J,
Lortet-Tieulent J and Jemal A: Global cancer statistics, 2012. CA
Cancer J Clin. 65:87–108. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Minna JD, Roth JA and Gazdar AF: Focus on
lung cancer. Cancer Cell. 1:49–52. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
International Early Lung Cancer Action
Program Investigators, ; Henschke CI, Yankelevitz DF, Libby DM,
Pasmantier MW, Smith JP and Miettinen OS: Survival of patients with
stage I lung cancer detected on CT screening. N Engl J Med.
355:1763–1771. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Ambros V: The functions of animal
microRNAs. Nature. 431:350–355. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Chen CZ: MicroRNAs as oncogenes and tumor
suppressors. N Engl J Med. 353:1768–1771. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Ma Y, Pan X, Xu P, Mi Y, Wang W, Wu X, He
Q, Liu X, Tang W and An HX: Plasma microRNA alterations between
EGFR-activating mutational NSCLC patients with and without primary
resistance to TKI. Oncotarget. 8:88529–88536. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Krentz Gober M, Collard JP, Thompson K and
Black EP: A microRNA signature of response to erlotinib is
descriptive of TGFβ behaviour in NSCLC. Sci Rep. 7:42022017.
View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Cui Y, Zhao L, Zhao S, Guo T, Li F, Li Z,
Fang L, Wu T and Gu C: MicroRNA-30e inhibits proliferation and
invasion of non-small cell lung cancer via targeting SOX9. Hum
Cell. 32:326–333. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Liu F, Cai Y, Rong X, Chen J, Zheng D,
Chen L, Zhang J, Luo R, Zhao P and Ruan J: miR-661 promotes tumor
invasion and metastasis by directly inhibiting RB1 in non small
cell lung cancer. Mol Cancer. 16:1222017. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Liu PJ, Ye YX, Wang YX, Du JX, Pan YH and
Fang XB: miRNA-92a promotes cell proliferation and invasion through
binding to KLF4 in glioma. Eur Rev Med Pharmacol Sci. 23:6612–6620.
2019.PubMed/NCBI
|
|
12
|
Yu H, Song H, Liu L, Hu S, Liao Y, Li G,
Xiao X, Chen X and He S: miR-92a modulates proliferation,
apoptosis, migration, and invasion of osteosarcoma cell lines by
targeting Dickkopf-related protein 3. Biosci Rep.
39:BSR201904102019. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Shigoka M, Tsuchida A, Matsudo T, Nagakawa
Y, Saito H, Suzuki Y, Aoki T, Murakami Y, Toyoda H, Kumada T, et
al: Deregulation of miR-92a expression is implicated in
hepatocellular carcinoma development. Pathol Int. 60:351–357. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Chen E, Li Q, Wang H, Yang F, Min L and
Yang J: miR-92a promotes tumorigenesis of colorectal cancer, a
transcriptomic and functional based study. Biomed Pharmacother.
106:1370–1377. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Jiang X, Li X, Wu F, Gao H, Wang G, Zheng
H, Wang H, Li J and Chen C: Overexpression of miR-92a promotes the
tumor growth of osteosarcoma by suppressing F-box and WD
repeat-containing protein 7. Gene. 606:10–16. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Zhu Q, Zang Q and Jiang ZM: Enhanced
expression of non coding miR 92a expression is implicated in the
development of lung cancer. Eur Rev Med Pharmacol Sci.
22:1028–1034. 2018.PubMed/NCBI
|
|
17
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Yu Y, Zuo J, Tan Q, Zar Thin K, Li P, Zhu
M, Yu M, Fu Z, Liang C and Tu J: Plasma miR-92a-2 as a biomarker
for small cell lung cancer. Cancer Biomark. 18:319–327. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Coşarcă AS, Mocan SL, Păcurar M, Fülöp E
and Ormenişan A: The evaluation of Ki67, p53, MCM3 and PCNA
immunoexpressions at the level of the dental follicle of impacted
teeth, dentigerous cysts and keratocystic odontogenic tumors. Rom J
Morphol Embryol. 57:407–412. 2016.PubMed/NCBI
|
|
20
|
Kent OA and Mendell JT: A small piece in
the cancer puzzle: microRNAs as tumor suppressors and oncogenes.
Oncogene. 25:6188–6196. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Kong YW, Ferland-McCollough D, Jackson TJ
and Bushell M: microRNAs in cancer management. Lancet Oncol.
13:e249–e258. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Nasser MW, Datta J, Nuovo G, Kutay H,
Motiwala T, Majumder S, Wang B, Suster S, Jacob ST and Ghoshal K:
Down-regulation of micro-RNA-1 (miR-1) in lung cancer. Suppression
of tumorigenic property of lung cancer cells and their
sensitization to doxorubicin-induced apoptosis by miR-1. J Biol
Chem. 283:33394–33405. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Takamizawa J, Konishi H, Yanagisawa K,
Tomida S, Osada H, Endoh H, Harano T, Yatabe Y, Nagino M, Nimura Y,
et al: Reduced expression of the let-7 microRNAs in human lung
cancers in association with shortened postoperative survival.
Cancer Res. 64:3753–3756. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Yang Y, Ding L, Hu Q, Xia J, Sun J, Wang
X, Xiong H, Gurbani D, Li L, Liu Y and Liu A: MicroRNA-218
functions as a tumor suppressor in lung cancer by targeting
IL-6/STAT3 and negatively correlates with poor prognosis. Mol
Cancer. 16:1412017. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Zhuang Z, Sun C and Gong H: High serum
miR-484 expression is associated with the diagnosis and prognosis
of patients with non-small cell lung cancer. Exp Ther Med.
18:4095–4102. 2019.PubMed/NCBI
|
|
26
|
Li M, Guan X, Sun Y, Mi J, Shu X, Liu F
and Li C: miR-92a family and their target genes in tumorigenesis
and metastasis. Exp Cell Res. 323:1–6. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Guo J, Wen N, Yang S, Guan X and Cang S:
miR-92a regulates oral squamous cell carcinoma (OSCC) cell growth
by targeting FOXP1 expression. Biomed Pharmacother. 104:77–86.
2018. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Ke TW, Wei PL, Yeh KT, Chen WT and Cheng
YW: miR-92a promotes cell metastasis of colorectal cancer through
PTEN-mediated PI3K/AKT pathway. Ann Surg Oncol. 22:2649–2655. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Chen ZL, Zhao XH, Wang JW, Li BZ, Wang Z,
Sun J, Tan FW, Ding DP, Xu XH, Zhou F, et al: microRNA-92a promotes
lymph node metastasis of human esophageal squamous cell carcinoma
via E-cadherin. J Biol Chem. 286:10725–10734. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Bae S, Lee EM, Cha HJ, Kim K, Yoon Y, Lee
H, Kim J, Kim YJ, Lee HG, Jeung HK, et al: Resveratrol alters
microRNA expression profiles in A549 human non-small cell lung
cancer cells. Mol Cells. 32:243–249. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Zhao J, Fu W, Liao H, Dai L, Jiang Z, Pan
Y, Huang H, Mo Y, Li S, Yang G and Yin J: The regulatory and
predictive functions of miR-17 and miR-92 families on cisplatin
resistance of non-small cell lung cancer. BMC Cancer. 15:7312015.
View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Lu C, Shan Z, Hong J and Yang L:
MicroRNA-92a promotes epithelial-mesenchymal transition through
activation of PTEN/PI3K/AKT signaling pathway in non-small cell
lung cancer metastasis. Int J Oncol. 51:235–244. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Welcker M and Clurman BE: FBW7 ubiquitin
ligase: A tumour suppressor at the crossroads of cell division,
growth and differentiation. Nat Rev Cancer. 8:83–93. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Minella AC and Clurman BE: Mechanisms of
tumor suppression by the SCF(Fbw7). Cell Cycle. 4:1356–1359. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Li Z, Xiao J, Hu K, Wang G, Li M, Zhang J
and Cheng G: FBXW7 acts as an independent prognostic marker and
inhibits tumor growth in human osteosarcoma. Int J Mol Sci.
16:2294–2306. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Xiao Y, Yin C, Wang Y, Lv H, Wang W, Huang
Y, Perez-Losada J, Snijders AM, Mao JH and Zhang P: FBXW7 deletion
contributes to lung tumor development and confers resistance to
gefitinib therapy. Mol Oncol. 12:883–895. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Zhou C, Shen L, Mao L, Wang B, Li Y and Yu
H: miR-92a is upregulated in cervical cancer and promotes cell
proliferation and invasion by targeting FBXW7. Biochem Biophys Res
Commun. 458:63–69. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Yang W, Dou C, Wang Y, Jia Y, Li C, Zheng
X and Tu K: MicroRNA-92a contributes to tumor growth of human
hepatocellular carcinoma by targeting FBXW7. Oncol Rep.
34:2576–2584. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Ren P, Gong F, Zhang Y, Jiang J and Zhang
H: MicroRNA-92a promotes growth, metastasis, and chemoresistance in
non-small cell lung cancer cells by targeting PTEN. Tumour Biol.
37:3215–3225. 2016. View Article : Google Scholar : PubMed/NCBI
|