Introduction
Atopic dermatitis (AD) is a chronic inflammatory
skin disease characterized by erythema, edema, thickening of the
skin and pruritic, eczematous skin lesions (1,2). The
prevalence of AD is increasing worldwide (3). It generally occurs in infancy and
childhood, but can also appear in adults. When AD begins in
childhood, it is usually followed by other allergic diseases, such
as allergic rhinitis and asthma (4). A number of remedies for AD involve
the topical or systemic administration of steroids and
antihistamines (5).
Corticosteroids, the most commonly used steroids, are the main
anti-inflammatory therapy and they are effective at inhibiting both
acute and chronic skin inflammation (6). Steroids are most commonly used for
anti-inflammatory therapy; however, their long-term use is limited
due to side-effects such as osteoporosis, brittle skin, muscle
weaknesses and diabetes (7).
Therefore, alternative treatment strategies are required for
AD.
AD is caused by inflammatory reactions to
environmental factors and allergen stimulation (8). The inflammatory reaction begins when
allergens bind to mast cells, which play an important role in early
inflammatory responses (9). When
mast cells are activated, they express inflammatory cytokine, such
as granulocyte-macrophage colony-stimulating factor (GM-CSF)
(10). Subsequently, activated
dendritic cells also express cytokines, such as thymus and
activation regulated chemokine (TARC/CCL17) and C-X-C motif
chemokine ligand 8 (CXCL-8/IL-8) (11). These actions cause the thickening
of the skin, erythema and itching.
1-Chloro-2,4-dinitrobenzene (DNCB) is a chemical
substance that causes chronic contact dermatitis and is widely used
in human studies of AD (12). When
DNCB is topically applied to mice, it causes dermatopathy similar
to symptoms of AD, such as an increased IgE expression,
inflammatory cytokines, edema and itching (13). Human adult low calcium, high
temperature (HaCaT) cells are human keratinocytes and have also
been widely used in dermatological studies (14). TNF-α/IFN-γ activate the
phagocytosis of granulocytes and macrophages, causing them to
secrete various cytokines. All these cytokines strongly influence
the recruitment of inflammatory cells, neutrophils and monocyte
chemotactic activity (15).
Traditionally, Solanum nigrum Linne (SNL) has
been used for decoding, urination, swelling, and heat treatment in
a number of traditional medicine systems worldwide, including
traditional Chinese medicine (16). Recently, SNL has been reported to
exert several positive biotic effects, such as anti-inflammatory,
tumor growth inhibitory and hepatoprotective effects (17–19).
However, the inhibitory effects of SNL on AD have not yet been
proven, at least to the best of our knowledge.
The present study aimed to investigate the
anti-inflammatory response by inhibiting mast cell and keratinocyte
activation during SNL treatment. For this purpose, BALB/c mice with
DNCB-induced AD and human keratinocyte lineage cell line HaCaT
cells were used as models of AD. The thickness of the epidermis and
dermis, the infiltration of inflammatory mediators, and the serum
expression levels of IgE were evaluated in BALB/c mice with
DNCB-induced AD. Moreover, the expression of the inflammatory
cytokines TARC, GM-CSF and CXCL-8, were investigated in
TNF-α/IFN-γ-stimulated HaCaT cells.
Materials and methods
Reagents
HaCaT cells were obtained from the cell line service
(CLS, Eppelheim, Germany). Penicillin-streptomycin (P/S), fetal
bovine serum (FBS) and goat serum (16210-064) were obtained from
Gibco; Thermo fisher Scientific, Inc. Dulbecco's modified Eagle's
medium (DMEM) was purchased form Welgene. CD4 (ab183685) and CD8
(ab209775) antibodies were obtained from Abcam. Antibodies to
phosphorylated-extracellular signal-regulated kinase (p-ERK,
#4370), extracellular signal-regulated kinase (ERK, #4695),
phosphorylated-p38 (p-p38, #4511), p38 (#9212), phosphorylated-Jun
N-terminal kinase (p-JNK, #4668), Jun N-terminal kinase (JNK,
#9258) and phosphorylated-NF-κB (p-NF-κB, #3033) were
purchased from Cell Signaling Technology, Inc. Lamin B was
purchased from Santa Cruz Biotechnology, Inc. Protease inhibitor
cocktail (#P8340) and phosphatase inhibitor cocktail (#P0044,
#P5726) were purchased from Sigma-Aldrich; Merck KGaA. ECL solution
(RPN2106) was obtained from GE Healthcare Life Sciences. PCR
primers glyceraldehyde 3-phosphate dehydrogenase (GAPDH),
TARC/CCL17, GM-CSF, CXCL-8 were obtained from Genotech. Taq
polymerase was obtained from KaPa BioSystems. TNF-α
(#285-IF), IFN-γ (#210-TA) were purchased from R&D
Systems.
Preparation of SNL
SNL was purchased from Omniherb. SNL was prepared as
a dried powder with lyophilization following sonication in 80%
ethanol for 2 h. The extract was concentrated in a rotary
evaporator and lyophilized (yield ratio 13.88%). The extracts were
stored at −20°C until use.
High-performance liquid chromatography
(HPLC) analysis of SNL
The separation and determination of SNL were
performed on an HPLC instrument (waters 2695 Alliance system with a
2996 UV detector) connected to a Photodiode Array detector (PDA).
The SNL extraction was separated by an Xbridge C18 column (4.6×250
mm, 5 µm). The temperature was maintained at 30°C, the
injection volume was 10 µl, and the flow rate was 1 ml/min.
The mobile phase consisted of acetonitrile (sol A) and 1% acetic
acid/H2O (sol B). The mobile phase solvent was used at a
9:1 ratio of sol A and sol B. The PDA detector wavelength was set
at 192 nm and the running time was 20 min.
Animals
All animal experiments were approved by the Kyung
Hee University Animal Care and Use Committee (KHMC-IACUC-18-016)
and were performed for 50 days in accordance with the guidelines of
Kyung Hee University Institutional Animal Care and Use Committee.
(KHMC-IACUC-18-016). BALB/c mice (6 weeks old) male were obtained
from KOATECH. All mice were bred in a controlled room (22±2°C
temperature, 50±10% humidity, 12-h light/dark cycle). They were
also stabilized for 7 days in a controlled room. All animals were
checked the daily for the health and sighs of morbidity. The
criteria for determining the animal's euthanasia period (endpoint)
are rapid weight loss, lethargy, debilitating diarrhea, sizable
abdominal enlargement and ascites. No animals died during the
experiment period.
AD model and drug treatment in
mice
The mice were anesthetized by the use of 5%
isoflurane of inhalant anesthetics in 100% oxygen and anesthesia
was maintained at 2~2.5% isoflurane inhalation. After anesthesia,
the dorsal skin of the mice was shaved with a clipper. The mice
were divided into 4 groups (n=8/group) as follows: The untreated
group (Normal), the DNCB-sensitized group (Control), the SNL 1
mg/ml-treated group (SNL_L), and the SNL 10 mg/ml-treated group
(SNL_H). The mice in the Normal group were treated with 9:1
phosphate-buffered saline (PBS)/olive oil. To induce AD,
1-chloro-2,4-dinitrobenzene (DNCB) was diluted at 0.5 and 1%. The
DNCB solvent was used at a 3:1 ratio of olive oil and acetone. The
mice in the Control, SNL_L and SNL_H groups were subjected to
initial sensitization with 1% DNCB. After 5 days, in the mice in
the Control, SNL_L and SNL_H groups, were 0.5% DNCB was dorsally
applied 3 times a week for 4 weeks. Following 2 h of the second
sensitization, SNL preparations of 1 and 10 mg/ml were applied on
the dorsal skin of the mice in the SNL_L and SNL_H groups once
every day for 35 days. The SNL solvent was used a 9:1 ratio of
PBS/olive oil. On day 36 after SNL treatments, all animals were
sacrificed. When the mouse was sacrificed, the isoflurane
concentration was adjusted to 5% to expose it, and the isoflurane
was exposed until one minute after breathing stopped. After
confirming that the heart and breathing has stopped, either 0.8 or
1 ml blood was collected and body weight was 30–35 g at the time of
sacrifice. According to the existing research, herbal medicines
were improved atopy dermatitis when the concentration of 1 to 10
mg/ml was applied to 200 µl. Therefore, we decided on the
concentration of SNL referring to the results of previous studies
(20–22).
Histological analysis
The skin tissues were fixed in 10% neutral buffered
formalin (NBF) for one day. After fixation, the skin tissues were
washed under running water for 24 h, then embedded with paraffin.
The paraffin block was sectioned at 5 µm thickness using a
microtome (ZEISS). The skin tissues were stained with hematoxylin
and eosin (H&E), Masson trichrome and toluidine blue to measure
the infiltration of eosinophils and mast cells and thickness of
epidermis and dermis.
Immunohistochemistry (IHC)
The skin tissue was deparaffinized and hydrated in
xylene and ethanol series. Skin tissues were heated with a 0.01 M
sodium citrate buffer (0.1 M citric acid, 0.1 M sodium citrate)
using an Electric Pressure Cooker (CPC-600; Cuisinart). After being
washed three times with tris-buffered saline (TBS), the skin tissue
was reacted with 0.3% H2O2 in methanol at
room temperature for 30 min to inhibit the activity of peroxidase.
The tissues were blocked with 10% goat serum in PBS for 10 min.
They were then incubated in CD4 and CD8 antibodies at 4°C for 24 h.
After 24 h, CD4- and CD8-positive cells were colored red using the
Polink-2 Plus AP rabbit kit (D70-18, GBI Labs) according to the
manufacturer's protocol, and were then counterstained with H&E.
CD8- and CD4-positive cells were counted in 1- fields using a light
microscope at ×400 magnification (BX51, Olympus Corp.).
Enzyme-linked immunosorbent assay
Blood samples were collected from the mice by
cardiac puncture following anesthesia, and the serum was separated
by centrifugation at 2,000 rpm for 10 min. IgE levels were
determined using a mouse enzyme-linked immunosorbent assay (ELISA)
kit, according to the manufacturer's protocol.
Cell culture and assessment of cell
viability
HaCaT cells were cultured in DMEM with 10% FBS and
1% P/S. They were then incubated at 37°C, 5% CO2 and 95%
humidity. HaCaT cells were seeded in a 96-well plate with
1.5×104 cells/well. After 24 h, the cells were treated
with SNL at 6.25, 12.5, 25, 50 and 100 µg/ml for 24 h. A
3-(4,5-dimethylthiazol-2-yl)-5-(3-carboxymethoxyphenyl)-2-(4-sulfophenyl)-2H-tetrazolium,
inner salt (MTS) solution was then added at 20 µl per well and the
HaCaT cells were incubated for 2 h at 37°C. Cell viability was
measured at a wavelength of 490 nm using an ELISA reader (Versamax;
Molecular Devices, LLC).
Reverse transcription-quantitative
polymerase chain reaction (RT-qPCR)
HaCaT cells were seeded in a 6-well plate at
1×106 cells/well. After 24 h, the HaCaT cells were
pretreated with various concentrations of SNL (6.25, 12.5, 25 and
50 µg/ml) for 1 h and stimulated with 10 ng/ml TNF-α/IFN-γ for 24
h. Total RNA was extracted using TRIzol reagent (TAKARA BIO)
according to the manufacturer's protocol. cDNA was prepared using
SuperScript II reverse transcriptase (Invitrogen; Thermo Fisher
Scientific, Inc.). The primer sequences for TARC, GM-CSF, CXCL-8,
and GAPDH are presented in Table
I. The cDNA samples were separated on a 1.2% agarose gel and
determined using NαBI (Neoscience). mRNA expression was measured
using ImageJ software (Ver. 1.52a, National Institutes of
Health).
 | Table I.Primer sequence for RT-PCR
analysis. |
Table I.
Primer sequence for RT-PCR
analysis.
| Primer name | Orientation | Sequence |
|---|
| TARC | Forward |
5′-ACTGCTCCAGGGATGCCATCGTTTTT-3′ |
|
| Reverse |
5′-ACAAGGGGATGGGATCTCCCTCACTG-3′ |
| GM-CSF | Forward |
5′-CAGCCTCACCAAGCTCAAGG-3′ |
|
| Reverse |
5′-TCATGAGAGAGCAGCTCCCC-3′ |
| CXCL-8 | Forward |
5′-ACATGACTTCCAAGCTGGCCG-′3 |
|
| Reverse |
5′-TTTATGAATTCTCAGCCCTC-′3 |
| GAPDH | Forward |
5′-ACTTTGTCAAGCTCATTTCC-′3 |
|
| Reverse |
5′-TGCAGCGAACTTTATTGATG-′3 |
Western blot analysis
The phosphorylation of MAPK and the activation of
NF-κB were examined by western blot analysis. The HaCaT cells,
incubated with diverse concentrations of SNL, were washed in cold
Dulbecco's phosphate-buffered saline (DPBS). The HaCaT cells were
lysed in a RIPA buffer (0.1% SDS, 150 mM NaCl, 50 mM Tris-Cl, 1%
NP-40, 0.5% Na-deoxycholate, a protease inhibitor cocktail, and a
phosphatase inhibitor cocktail) and incubated in ice for 30 min.
Following centrifugation at 13,200 rpm for 20 min at 4°C, the
protein concentration was calculated by bicinchoninic acid (BCA)
assay. Protein samples (40 µg) were separated by 10% SDS-PAGE. The
protein was transferred to a nitrocellulose membrane and then
blocked with 5% skim milk for 1 h. After blocking, the membrane was
incubated with primary antibodies, such as p-ERK, t-ERK, p-JNK,
t-JNK, p-p38, t-p38, p-NF-κB and Lamin B in a 1% bovine serum
albumin (BSA) solution at 4°C overnight. After the membrane was
incubated with secondary antibodies, the protein was detected using
an ECL solution.
Statistical analysis
Each experiment was repeated at least 3 times. The
data are presented as the mean ± standard error of the mean
(SEM). All data were analyzed using the Graph Pad PRISM software
(GraphPad Software, Inc.). One-way ANOVA was used to evaluate the
treatment effect, followed by Tukey's post-hoc test. P<0.05 was
considered to indicate a statistically significant difference.
Results
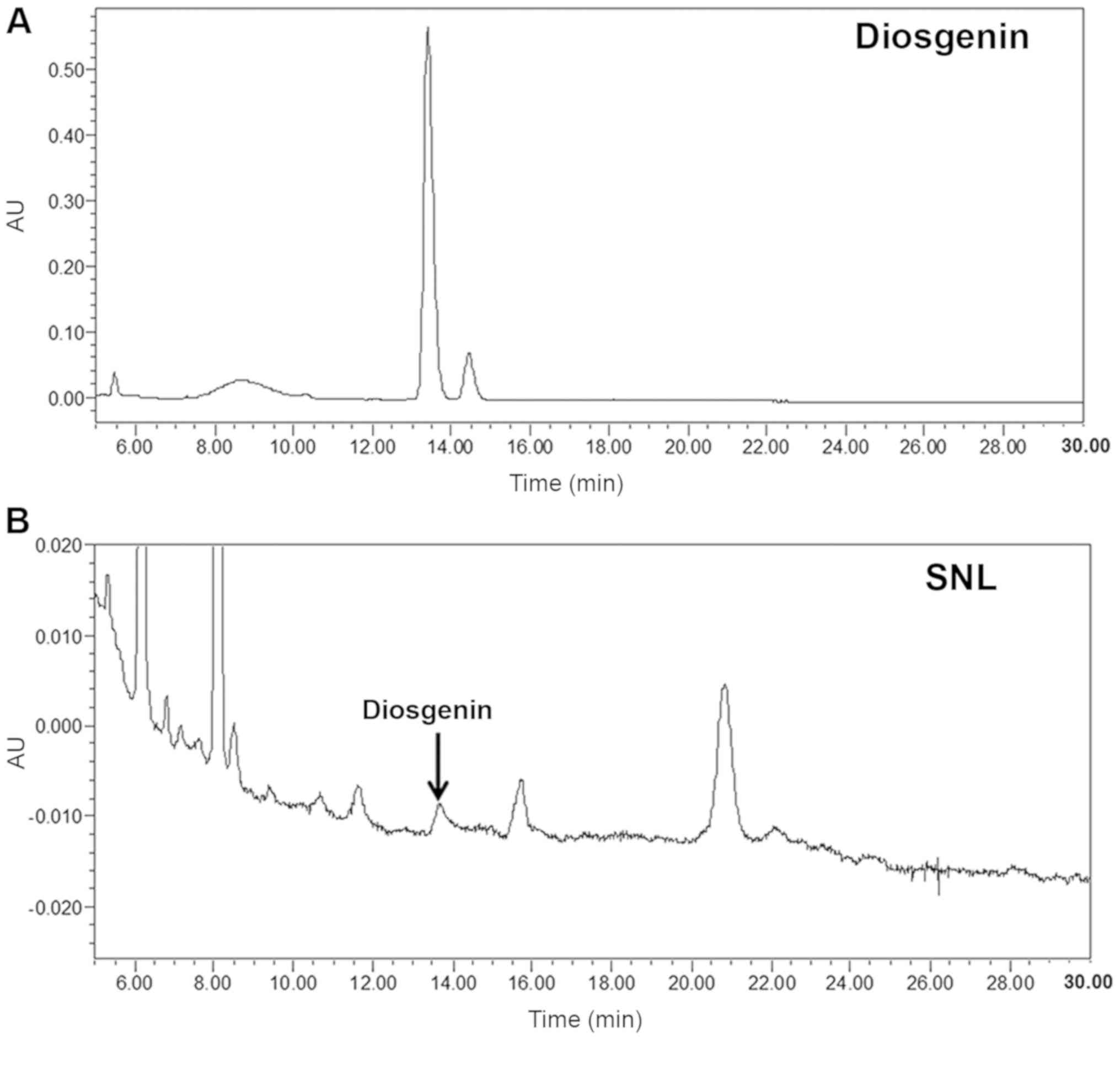
HPLC analysis of SNL
According to previous studies, the diosgenin is one
of the ingredients of SNL, which has excellent anti-inflammatory
and anticancer effects and is detected in SNL to 0.16–3% (23,24).
In the present study, the quality and purity of SNL were measured
using a HPLC analyzer. The typical chromatography profile of the
diosgenin standard and SNL extract are depicted in Fig. 1. The retention time of the
diosgenin standard and SNL extract was 13.439 min. Therefore, SNL
was identified as diosgenin based on the HPLC-PDA data.
Effects of SNL on DNCB-induced AD-like
symptoms in BALB/c mice
All animal experiments were based on the schedule
illustrated in Fig. 2. The dorsal
skin on which the SNL was applied for 5 weeks in the model of
DNCB-induced atopic dermatitis is illustrated in Fig. 3A. Body weight, liver weight and
aspartate aminotransferase (AST) and alanine aminotransferase (ALT)
levels were measured to determine the toxicity of SNL in the mouse
model of AD (Fig. 3B-E). The body
weight of the mice did not exhibit any marked differences between
the Control, SNL_L and SNL_H groups were compared. Following
sacrifice, the livers were rapidly removed and weighed. The liver
weight was measured to evaluate the toxicity of SNL. The liver
weight of the mice in the Control group was significantly increased
compared with the mice in the Normal group. The mice in the SNL-L
group exhibited a decreased liver weight compared with the mice in
the Control and the SNL-H groups. Total AST and ALT levels are
indicators of hepatic function. No significant changes in the
levels of AST and ALT were observed in the SNL_L and SNL_H compared
to the Control group. This result indicated that SNL was an
effective sample for atopic dermatitis at a non-toxic
concentration. The levels of serum IgE were measured by ELISA
(Fig. 3F). The serum IgE levels
were increased in the Control group compared with the Normal group.
However, the serum IgE levels were significantly decreased in the
SNL_L compared with the Control group.
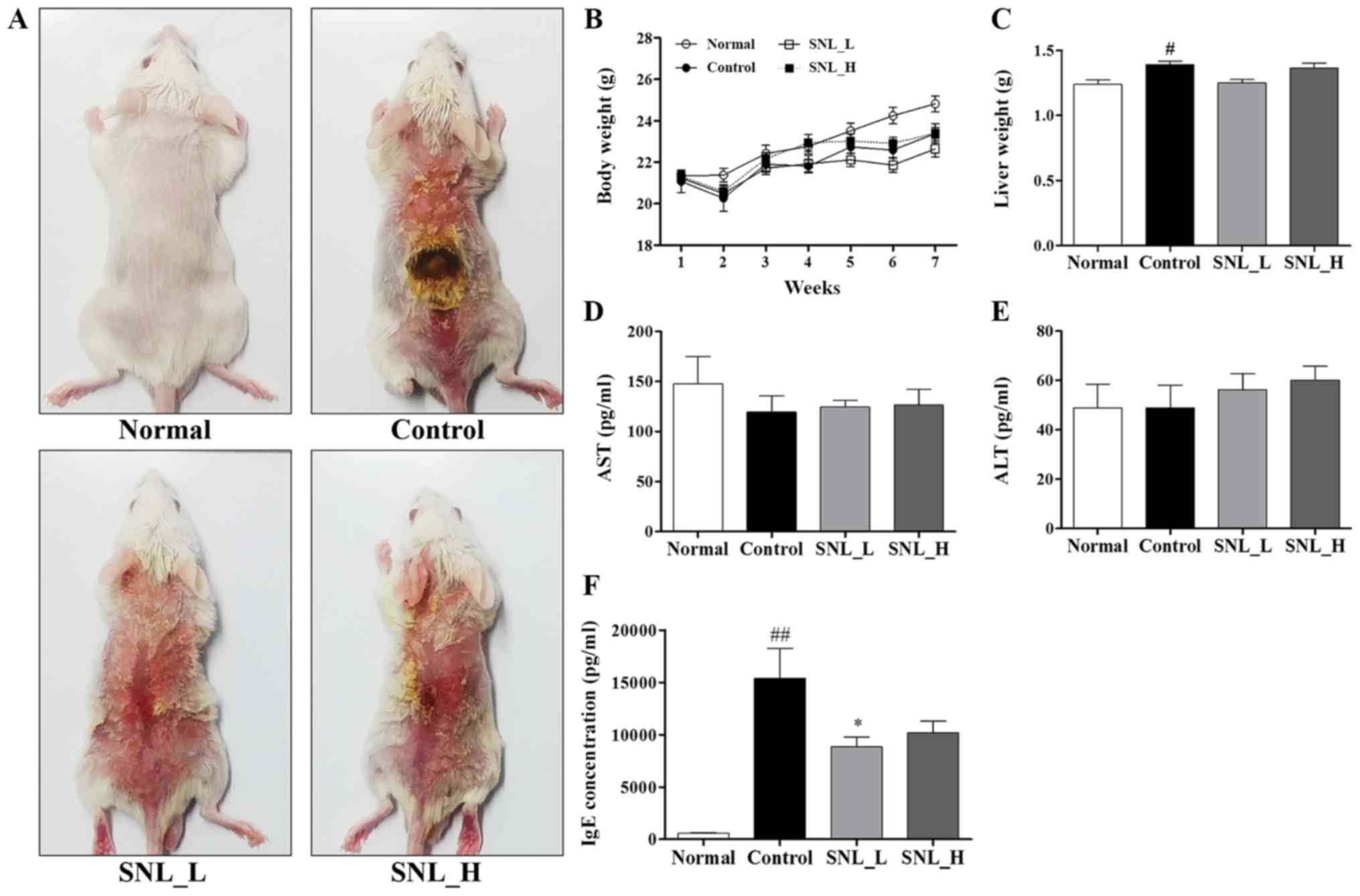 | Figure 3.Treatment with SNL inhibits
DNCB-induced AD-like symptoms in BALB/c mice. (A) SNL was applied
for 5 weeks to the dorsal skins of mice with DNCB-induced AD. (B)
Body weight was measured once a week. (C) After the mice were
sacrificed, the liver weight was measured. (D) AST and (E) ALT
levels were measured to determine the toxicity of the drug in the
blood following sacrifice. (F) Total serum IgE levels. Data
represent the means ± SEM. #P<0.05 and
##P<0.01, compared with the Normal group. *P<0.05,
compared with the Control group. Normal, untreated group; Control,
DNCB-sensitized group; SNL_L, Solanum nigrum Linne 1
mg/ml-treated group; SNL_H, Solanum nigrum Linne 10
mg/ml-treated group; AST, aspartate aminotransferase; ALT, alanine
aminotransferase; IgE, immunoglobulin E; AD, atopic dermatitis;
SEM, standard error of the mean. |
Effects of SNL on epidermal and dermal
thickness, and mast cells in mice with DNCB-induced AD
The mouse skin was stained with Masson's trichrome
stain to evaluate the effects of SNL on epidermal and dermal
thickness (Fig. 4A). The
thicknesses of the epidermis and dermis were measured using ImageJ
software. The thickness of the epidermis and dermis increased in
the Control group compared to the Normal group. The thickness of
the epidermis was significantly decreased in the SNL_H compared
with the Control group. The thickness of the dermis was
significantly decreased in both the SNL_L and SNL_H groups compared
to the Control group (Fig. 4F and
G). To measure the degree of mast cell infiltration, skin
tissues were stained with toluidine blue (Fig. 4B). The infiltration of mast cells
increased in the Control group compared with the Normal group. The
numbers of mast cells were decreased in the SNL_L compared with the
Control group (Fig. 4H). To
measure the degree of eosinophils infiltration, skin tissues were
stained with H&E (Fig. 4C).
The infiltration of eosinophils increased in the Control group
compared with the Normal group. However, eosinophils did not affect
the SNL_L and SNL_H compared to the Control group (Fig. 4I). IHC staining was performed on
the skin with atopic dermatitis to observe the extent of CD4 and
CD8 infiltration. CD4 and CD8 infiltration increased in the Control
group compared with the Normal group (Fig. 4D and E). However, the number of
CD8-positive cells decreased in the SNL_L compared with the Control
group (Fig. 4J). No significant
difference was observed in the number of CD4-positive cells between
the Control, SNL_L and SNL_H groups (Fig. 4K).
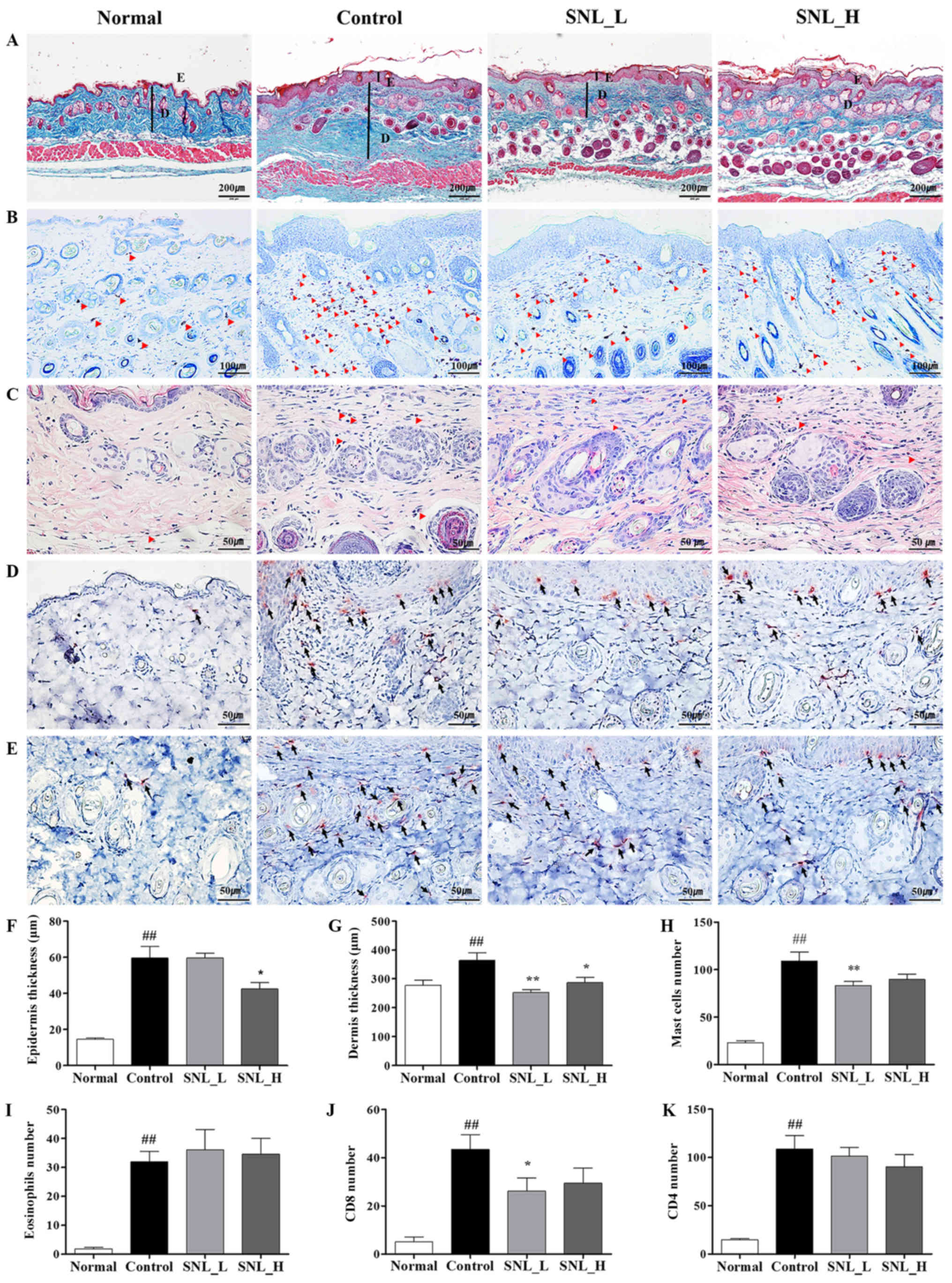 | Figure 4.SNL suppresses the infiltration of
immune cells in the skins of mice with atopic dermatitis. (A, F and
G) The thicknesses of the epidermis and dermis were examined by
Masson's trichrome staining of the skin sections (magnification,
×100; scale bar, 200 µm). (B) The infiltration of mast cells in the
dermis was examined by toluidine blue staining of the skin section
(magnification, ×200; scale bar, 100 µm). (H) The mast cells were
counted in 3 fields. (C) The infiltration of eosinophils in the
dermis was examined by H&E staining of skin sections
(magnification, ×400; scale bar, 50 µm). (I) The eosinophils were
counted in 10 fields. (D) The infiltration of CD8 in the epidermis
was examined by IHC staining of the skin sections (magnification,
×400; scale bar, 50 µm). (E) The infiltration of CD4 in the dermis
was examined by IHC staining of the skin sections (magnification,
×400; scale bar, 50 µm). (J and K) CD8- and CD4-positive cells were
counted in 10 fields. Data represent the means ± SEM.
##P<0.01, compared with the Normal group. *P<0.05
and **P<0.01, compared with the Control group. Normal, untreated
group; Control, DNCB-sensitized group; SNL, Solanum nigrum
Linne; SNL_L, Solanum nigrum Linne 1 mg/ml treated group;
SNL_H, Solanum nigrum Linne 10 mg/ml-treated group; E,
epidermal thickness; D, dermal thickness; CD4, cluster of
differentiation 4; CD8, cluster of differentiation 8; SEM, standard
error of the mean. |
Cytotoxicity of SNL in HaCaT
cells
MTS assay was used to measure the cytotoxicity of
SNL in HaCaT cells. HaCaT cells were treated with various
concentrations of SNL (0–100 µg/ml). SNL was found to be cytotoxic
at 100 µg/ml (Fig. 5A). Therefore,
SNL was used at a concentration <50 µg/ml in following
experiments, which was not cytotoxic to HaCaT cells.
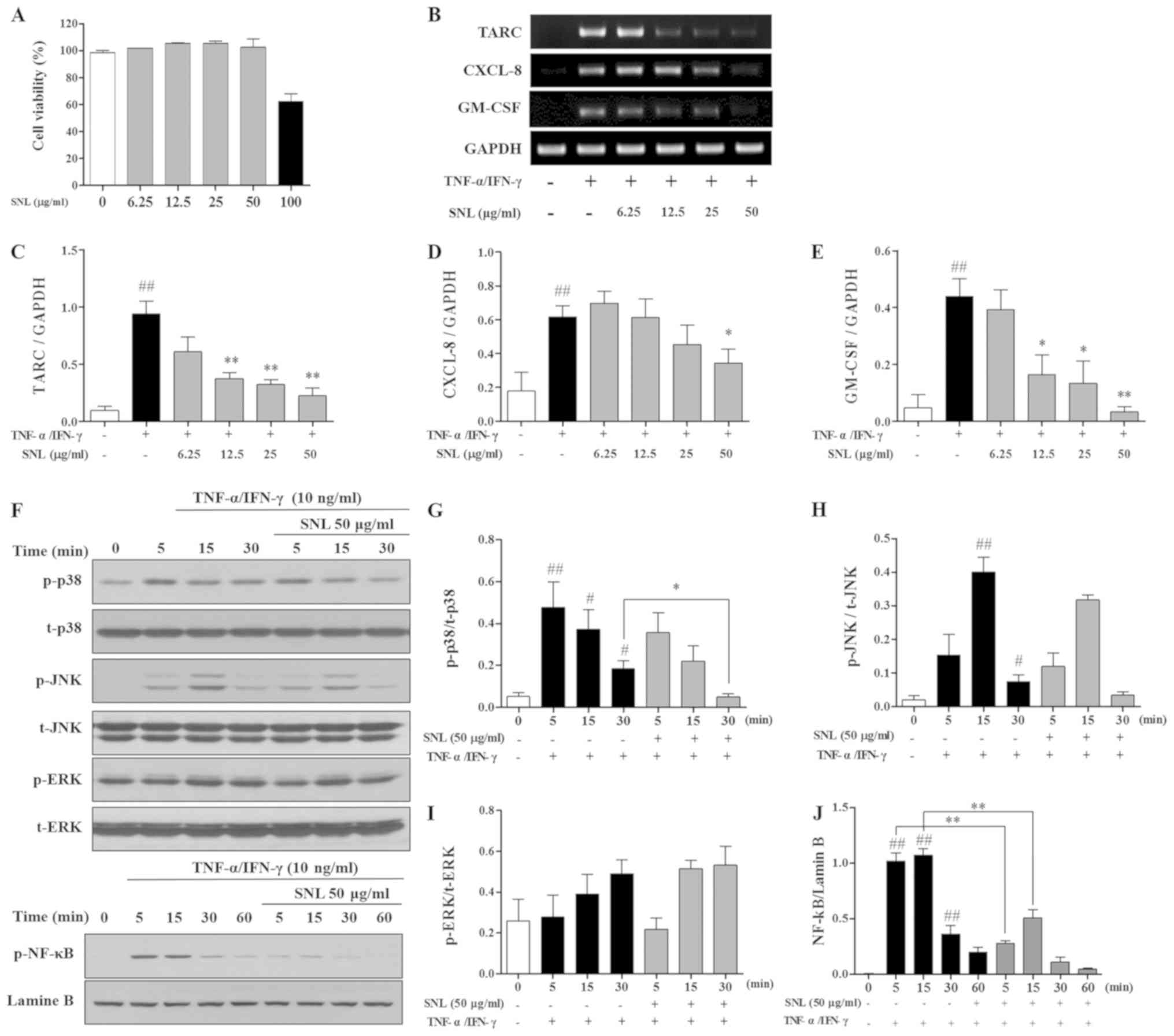 | Figure 5.Effects of SNL on
TNF-α/IFN-γ-stimulated HaCaT cells. (A) Viability of HaCaT cells
following SNL treatment was determined by an MTS assay after 24 h.
(B-E) Gene expression of TARC, CXCL-8 and GM-CSF in HaCaT cells
detected by RT-qPCR. HaCaT cells were treated with SNL for 1 h
prior to stimulation with TNF-α (10 ng/ml) and IFN-γ (10 ng/ml).
(F-J) Phosphorylation and degradation of MAPK and NF-κB were
examined by western blot analysis. mRNA and protein expression were
measured using ImageJ software. Data represents the means ± SEM.
#P<0.05 and ##P<0.01, compared with the
Normal group (not stimulated with TNF-α/IFN-γ). *P<0.05 and
**P<0.01, compared with the TNF-α/IFN-γ-stimulated HaCaT cells.
SNL, Solanum nigrum Linne; TNF-α, tumor necrosis factor-α;
IFN-γ, Interferon-γ; HaCaT, human adult low calcium high
temperature; TARC, thymus and activation regulated chemokine;
CXCL-8, C-X-C motif chemokine ligand 8; GM-CSF,
granulocyte-macrophage colony-stimulating factor; GAPDH,
glyceraldehyde 3-phosphate dehydrogenase; RT-qPCR, reverse
transcription-quantitative polymerase chain reaction; MAPK,
mitogen-activated protein kinase; p-, phosphorylated; t- total;
ERK1/2, extracellular signal-regulated kinase 1/2; JNK, c-Jun
N-terminal kinase; NF-κB, nuclear factor-κB; SEM, standard error of
the mean. |
Effects of SNL on mRNA expression in
TNF-α/IFN-γ stimulated HaCaT cells
mRNA expression in TNF-α/IFN-γ-stimulated HaCaT
cells was measured by RT-qPCR. SNL at concentrations of 12.5, 25,
and 50 µg/ml significantly decreased the level of TARC in the HaCaT
cells. The expression of the inflammatory cytokine, CXCL-8, was
inhibited with SNL at 50 µg/ml. In addition, SNL at 12.5, 25 and 50
µg/ml decreased the GM-CSF level in the TNF-α/IFN-γ-stimulated
HaCaT cells (Fig. 5B-E).
Effects of SNL on phosphorylation of
MAPKs and NF-κB in TNF-α/IFN-γ stimulated HaCaT cells
The present study examined the mechanisms through
which SNL affects the expression of the MAPK and NF-κB pathway. As
shown in Fig. 5F-J, the expression
of p-p38 was significantly inhibited at 30 min of TNF-α/IFN-γ
stimulation; however, SNL treatment increased p-p38 expression
after 5 min. However, the levels of JNK and ERK were not affected
by SNL. The expression of p-NF-κB was significantly inhibited at 5,
15 min compared with the TNF-α/IFN-γ-stimulated HaCaT cells.
Discussion
The present study aimed to investigate the effects
of SNL on a model of DNCB-induced AD and TNF-α/IFN-γ-stimulated
HaCaT cells. Symptoms of AD include the thickening of the epidermis
and dermis, the infiltration of mast cells and eosinophils, the
overexpression of Th2 cytokines, and increased IgE production
(12,13,25,26).
In the present study, BALB/c mice with DNCB-induced AD and human
keratinocyte HaCaT cells were used, which are widely used as models
of AD, to determine whether SNL decreases the inflammatory
response.
Skin thickening is a well-known symptom of AD, both
clinically and historically (27).
Continuous inflammation and allergic reactions can cause the skin
to become thick and hard (28). In
the present study, it was found that SNL inhibited the thickening
of the epidermis and dermis in the model of DNCB-induced AD. As
shown Fig. 4, the skin of the mice
in the Control group thickened compared to that of the Normal
group, and the skin of the mice in the SNL_L and SNL_H groups was
thinner than that of the Control group. These results indicate that
SNL reduces the hyperkeratosis of AD.
The expression of IgE is known to cause acute and
chronic skin inflammation (8). In
particular, the increase in IgE levels has been reported as a
characteristic of AD. IgE binds to high-affinity receptors on the
surface of mast cells to activate them (29). In the present study, the serum IgE
levels were significantly increased in mice with DNCB-induced AD.
SNL decreased the level of IgE in the serum of mice with
DNCB-induced AD. Mast cells are an early indicator of the
inflammatory response (30). Mast
cells are activated by the stimulation of IgE molecules, and
activated mast cells increase the production of inflammatory
cytokines, such as IL-5, IL-6, IL-13 and GM-CSF (31). The findings of the present study
demonstrated that the infiltration of mast cells was decreased in
the SNL_L group. This suggests that SNL inhibits mast cell
infiltration and may improve tissue-related changes, such as
characteristic edema and skin thickening in AD. CD8 is a
transmembrane glycoprotein that serves as a co-receptor for the T
cell receptor (32). Cytotoxic T
cells that express CD8 on the cell surface are termed
CD8+ T cells. CD8+ T cells play an immune
defense against intracellular pathogens, including viruses and
bacteria (33). Previous studies
have demonstrated that CD8+ T cells are essential for
eczema formation in a mouse model, they appear in human skin (even
prior to Th2 cells) following allergen exposure, and are a source
of inflammatory cytokines (34).
Hijnen et al reported that inflammatory cytokines are
overexpressed by CD8+ T cells in AD-affected skin
(35). The results of the present
study confirmed the increase production of CD8 in the model of
DNCB-induced AD. The infiltration of CD8+ T cells was
significantly increased in DNCB-exposed mice, and SNL treatment of
the DNCB-exposed mice significantly reduced the infiltration
CD8+ T cells compared to the Control group. The
expression of IgE is known to cause acute and chronic skin
inflammation.
The inflammatory cytokine, GM-CSF, is secreted by
mast cells and induces the infiltration of eosinophils (36). This causes the skin to thicken as a
result of swelling and edema. TARC secreted from dendritic cells
plays an important role in recruiting and activating Th2 cells,
which, in turn, play an important in the inflammatory response
(18). CXCL-8 is an
eosinophil-activating cytokine. CXCL-8 is expressed by various
tissue and phagocytic cells when exposed to inflammatory stimulants
(37). CXCL-8 recruits and
activates eosinophils from inflammatory lesions (38). These eosinophils amplify the
inflammatory response (39). In
the present study, SNL significantly decreased the levels of
inflammatory cytokines, such as TARC, CXCL-8 and GM-CSF in the
TNF-α/IFN-γ-stimulated HaCaT cells. This result suggests that SNL
decreased the inflammatory reaction in BALB/c mice by inhibiting
TARC, CXCL-8, and GM-CSF in keratinocytes. MAPKs are signal
transduction pathways, and they are important mediators of
transcriptional responses to extracellular signals, including
hormones, cytokines, and environmental stress. In particular, p38
MAPK is activated by cellular stress and modulates the expression
of inflammatory cytokines, such as CXCL-8, TARC and GM-CSF
(40,41). NF-κB is an important transcription
factor activated by various stimuli, such as TNF-α and IFN-γ
(42). In the present study, p38
MAPK and NF-κB signaling in the TNF-α/IFN-γ-stimulated HaCaT cells
were decreased by SNL treatment.
In conclusion, the findings of the present study
suggest that p38 MAPK induces the activity of NF-κB and activated
NF-κB produces inflammatory cytokines. Immune cell infiltration, as
well as cytokine and keratinocyte production were confirmed in the
skins of mice with AD. However, SNL relieved the symptoms of
AD.
Acknowledgements
Not applicable.
Funding
The current study was supported by the National
Research Foundation of Korea (NRF) grant funded by the Korea
government (MSIT) (grant no. 2020R1A2C2005836).
Availability of data and materials
All data generated or analyzed during this study are
included in this published article.
Authors' contributions
YS and HSJ conceptualized the study. SH was been
involved in drafting the manuscript. SH, BL, BK and HC performed
all experiments and verified the analytical data. BK, BL and HC
involved in critically revising the manuscript. EYK, JHK and MK
contributed to the statistical analysis and helped interpret the
results. EYK supervised the experiments in discussion with SH and
BL. All authors read and approved the final manuscript.
Ethics approval and consent to
participate
All animal experiments were approved by the Kyung
Hee University Animal Care and Use Committee (approval no.
KHMC-IACUC-18-016).
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Jung M, Lee TH, Oh HJ, Kim H, Son Y, Lee
EH and Kim J: Inhibitory effect of 5,6-dihydroergosteol-glucoside
on atopic dermatitis-like skin lesions via suppression of NF-kappaB
and STAT activation. J Dermatol Sci. 79:2878–261. 2015. View Article : Google Scholar
|
|
2
|
Lim SJ, Kim M, Randy A, Nam EJ and Nho CW:
Effects of Hovenia dulcis Thunb. Extract and methyl vanillate on
atopic dermatitis-like skin lesions and TNF-α/IFN-γ-induced
chemokines production in HaCaT cells. J Pharm Pharmacol.
68:1465–1479. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Spergel JM and Paller AS: Atopic
dermatitis and the atopic march. J Allergy Clin Immunol. 112 (Suppl
6):S118–S127. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Nutten S: Atopic dermatitis: Global
epidemiology and risk factors. Ann Nutr Metab. 66 (Suppl 1):S8–S16.
2015. View Article : Google Scholar
|
|
5
|
Charman CR, Morris AD and Williams HC:
Topical corticosteroid phobia in patients with atopic eczema. Br J
Dermatol. 142:931–936. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Leung DYM, Boguniewicz M, Howell MD,
Nomura I and Hamid OA: New insights into atopic dermatitis. J Clin
Invest. 113:651–657. 2004. View
Article : Google Scholar : PubMed/NCBI
|
|
7
|
Kleiman A and Tuckermann JP:
Glucocorticoid receptor action in beneficial and side effects of
steroid therapy: Lessons from conditional knockout mice. Mol Cell
Endocrinol. 275:98–108. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Galli SJ, Tsai M and Piliponsky AM: The
development of allergic inflammation. Nature. 454:445–454. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Kraneveld AD, Sagar S, Garssen J and
Folkerts G: The two faces of mast cells in food allergy and
allergic asthma: The possible concept of Yin Yang. Biochim Biophys
Acta. 1822:93–99. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Dahl C, Hoffmann HJ, Saito H and Schiotz
PO: Human mast cells express receptors for IL-3, IL-5 and GM-CSF; a
partial map of receptors on human mast cells cultured in vitro.
Allergy. 59:1087–1096. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Liu YJ: Thymic stromal lymphopoietin:
Master switch for allergic inflammation. J Exp Med. 203:269–273.
2006. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Fujii Y, Takeuchi H, Sakuma S, Sengoku T
and Takakura S: Characterization of a
2,4-dinitrochlorobenzene-induced chronic dermatitis model in rats.
Skin Pharmacol Physiol. 22:240–247. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Inagaki N, Shiraishi N, Igeta K, Itoh T,
Chikumoto T, Nagao M, Kim JF and Nagai H: Inhibition of scratching
behavior associated with allergic dermatitis in mice by tacrolimus,
but not by dexamethasone. Eur J Pharmacol. 546:189–196. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Yang CT, Yang ZL, Zhang MF, Dong Q, Wang
XY, Lan AP, Zeng FQ, Chen PX, Wang CH and Feng JQ: Hydrogen sulfide
protects against chemical hypoxia-induced cytotoxicity and
inflammation in HaCaT cells through inhibition of ROS/NF-κB/COX-2
pathway. PLoS One. 6:e219712011. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Chodorowska G: Plasma concentrations of
IFN-gamma and TNF-alpha in psoriatic patients before and after
local treatment with dithranol ointment. J Eur Acad Dermatol
Venereol. 10:147–151. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Loganayaki N, Siddhuraju P and Manian S:
Antioxidant activity of two traditional Indian vegetables:
Solanum nigrum L. and Solanum torvum L. Food Sci
Biotechnol. 19:121–127. 2010. View Article : Google Scholar
|
|
17
|
Proksch E, Folster-Holst R and Jensen JM:
Skin barrier function, epidermal proliferation and differentiation
in eczema. J Dermatol Sci. 43:159–169. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Lambrecht BN and Hammad H: The role of
dendritic and epithelial cells as master regulators of allergic
airway inflammation. Lancet. 376:835–843. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Galli SJ: The Mast Cell-IgE Paradox: From
homeostasis to anaphylaxis. Am J Pathol. 186:212–224. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Kim SY, Yohannes SB, Damte D, Lee SJ,
Hossain MA, Kim JY, Rhee MH, Suh JW and Park SC: Effect of
fermented rhus verniciflua extract on DNCB induced-atopy like
dermatitis in BALB/c mice. Pak Vet J. 34:333–336. 2014.
|
|
21
|
Choi YY, Kim MH, Ahn KS, Um JY, Lee SG and
Yang WM: Immunomodulatory effects of Pseudostellaria heterophylla
(Miquel) Pax on regulation of Th1/Th2 levels in mice with atopic
dermatitis. Mol Med Rep. 15:649–656. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Lee HG, Cho NC, Jeong AJ, Li YC, Rhie SJ,
Choi JS, Lee KH, Kim Y, Kim YN, Kim MH, et al: Immunomodulatory
activities of the benzoxathiole derivative BOT-4-one ameliorate
pathogenic skin inflammation in mice. J Invest Dermatol.
136:107–116. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Suthar AC and Mulani RM: A high
performance thin layer chromatography method for quantitative
estimation of Diosgenin in Solanum nigrum Linn. Pharmacog
Magazine. 4:112–115. 2008.
|
|
24
|
Desai S, Tatke P and Gabhe SY:
Quantification of diosgenin in extracts and formulations containing
Solanum Nigrum. Int J Pharm Sci Res. 6:676–681. 2015.
|
|
25
|
Matsuda H, Watanabe N, Geba GP, Sperl J,
Tsudzuki M, Hiroi J, Matsumoto M, Ushio H, Saito S, Askenase PW and
Ra C: Development of atopic dermatitis-like skin lesion with IgE
hyperproduction in NC/Nga mice. Int Immunol. 9:461–466. 1997.
View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Kabashima K: New concept of the
pathogenesis of atopic dermatitis: Interplay among the barrier,
allergy, and pruritus as a trinity. J Dermatol Sci. 70:3–11. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Wang G, Savinko T, Wolff H, Dieu-Nosjean
MC, Kemeny L, Homey B, Lauerma AI and Alenius H: Repeated
epicutaneous exposures to ovalbumin progressively induce atopic
dermatitis-like skin lesions in mice. Clin Exp Allergy. 37:151–161.
2007. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Skoner DR: Allergic rhinitis: Definition,
epidemiology, detection, and pathophysiology, diagnosis. J Allergy
Clin Immun. 108 (Suppl 1):S2–S8. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Amarasekera M: Immunoglobulin E in health
and disease. Asia Pac Allergy. 1:12–15. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
De Filippo K, Dudeck A, Hasenberg M, Nye
E, van Rooijen N, Hartmann K, Gunzer M, Roers A and Hogg N: Mast
cell and macrophage chemokines CXCL1/CXCL2 control the early stage
of neutrophil recruitment during tissue inflammation. Blood.
121:4930–4937. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Stone KD, Prussin C and Metcalfe DD: IgE,
mast cells, basophils, and eosinophils. J Allergy Clin Immun. 125
(2 Suppl 2):S73–S80. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Janeway CA Jr: The T cell receptor as a
multicomponent signalling machine: CD4/CD8 coreceptors and CD45 in
T cell activation. Annu Rev Immunol. 10:645–674. 1992. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Nagata T and Koide Y: Induction of
specific CD8(+) T cells against intracellular bacteria by CD8(+)
T-cell-oriented immunization approaches. J Biomed Biotechnol.
2010:7645422010. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Roesner LM, Heratizadeh A, Wieschowski S,
Mittermann I, Valenta R, Eiz-Vesper B, Hennig C, Hansen G, Falk CS
and Werfel T: α-NAC-specific autoreactive CD8+ T cells in atopic
dermatitis are of an effector memory type and secrete IL-4 and
IFN-γ. J Immunol. 196:3245–3252. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Hijnen D, Knol EF, Gent YY, Giovannone B,
Beijn SJ, Kupper TS, Bruijnzeel-Koomen CA and Clark RA: CD8(+) T
cells in the lesional skin of atopic dermatitis and psoriasis
patients are an important source of IFN-γ, IL-13, IL-17, and IL-22.
J Invest Dermatol. 133:973–979. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Gordon JR, Burd PR and Galli SJ: Mast
cells as a source of multifunctional cytokines. Immunol Today.
11:458–464. 1990. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Baggiolini M and Clark-Lewis I:
Interleukin-8, a chemotactic and inflammatory cytokine. FEBS Lett.
307:97–101. 1992. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Hedges JC, Singer CA and Gerthoffer WT:
Mitogen-activated protein kinases regulate cytokine gene expression
in human airway myocytes. Am J Respir Cell Mol Biol. 23:86–94.
2000. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Jacobsen EA, Lee NA and Lee JJ:
Re-defining the unique roles for eosinophils in allergic
respiratory inflammation. Clin Exp Allergy. 44:1119–1136. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Barnes PJ and Stockley RA: COPD: Current
therapeutic interventions and future approaches. Eur Respir J.
25:1084–1106. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Kwon DJ, Bae YS, Ju SM, Goh AR, Youn GS,
Choi SY and Park J: Casuarinin suppresses TARC/CCL17 and MDC/CCL22
production via blockade of NF-κB and STAT1 activation in HaCaT
cells. Biochem Biophys Res Commun. 417:1254–1259. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Ju SM, Song HY, Lee SJ, Seo WY, Sin DH,
Goh AR, Kang YH, Kang IJ, Won MH, Yi JS, et al: Suppression of
thymus- and activation-regulated chemokine (TARC/CCL17) production
by 1,2,3,4,6-penta-O-galloyl-beta-D-glucose via blockade of
NF-kappaB and STAT1 activation in the HaCaT cells. Biochem Biophys
Res Commun. 387:115–120. 2009. View Article : Google Scholar : PubMed/NCBI
|