Introduction
Colorectal cancer (CRC) is one of the most frequent
malignant gastrointestinal tumors that threatens human health
(1). CRC has a high incidence
rate; the second highest (9.5% of 8.6 million new cases) in females
and the third highest (10.9% of 9.5 million new cases) in males
worldwide (2). Therefore,
investigating the pathogenesis of CRC can contribute to the
treatment and diagnosis of the disease.
With the development of sequencing technology, the
functions of non-coding RNAs (ncRNAs) in the development,
metastasis and drug resistance in multiple cancer types have been
increasingly revealed (3–9). Long ncRNAs (lncRNAs) are a type of
ncRNAs with a length of >200 nucleotides (nts) and serve
important regulatory roles in tumor cell progression, including
cell proliferation, migration, invasion and apoptosis in CRC
(10–12). For example, Zhang et al
(13) reported that lncRNA CPS-IT1
is downregulated in CRC tissues and cells, which could inhibit CRC
cell proliferation, invasion and metastasis. MicroRNAs
(miRNAs/miRs), a type of ncRNAs with a length of ~22 nts, are also
closely related to the proliferation, invasion and apoptosis of CRC
cells (14–16). Moreover, Luo et al (17) demonstrated that miR-17 upregulation
contributes to cell proliferation, tumor growth and cycle
progression in CRC.
lncRNAs and miRNAs have become widely investigated
in the research of cancer pathogenesis. A number of studies have
revealed that lncRNAs and miRNAs serve important regulatory roles
in tumor occurrence and migration by forming regulatory networks
(18–21). For example, lncRNA PVT1 promotes
cell proliferation and migration by regulating miR-448 in
pancreatic cancer (22).
TMPO-antisense (AS)1 is also associated with CRC and may serve a
role in CRC (23). Cui et
al (24) reported that
TMPO-AS1 acts as a competing endogenous RNA to regulate
osteosarcoma tumorigenesis via the miR-199a-5p/Wnt family member 7B
axis. The present study found that TMPO-AS1 expression was
increased in CRC cells. However, the underlying regulatory
mechanism of TMPO-AS1 has not been fully investigated in CRC.
In the current study, the effects of TMPO-AS1 and
miR-143-3p on cell proliferation, migration and invasion were
investigated, and the regulatory relationship between TMPO-AS1 and
miR-143-3p was further evaluated. The present study demonstrated
that the regulatory networks of TMPO-AS1 and miR-143-3p affected
the cellular proliferation of CRC in vitro and in
vivo.
Materials and methods
Cell culture and transfection
Human CRC cell lines (SW480, HCT15, SW1116 and
HCT116) and normal connective mucosal epithelial cells (NCM460)
were purchased from the Cell Bank of the Chinese Academy of
Science. All cells were cultured in DMEM (Thermo Fisher Scientific,
Inc.) containing 10% FBS (Invitrogen; Thermo Fisher Scientific,
Inc.) and 1% penicillin and streptomycin under a 5% CO2
atmosphere at 37°C.
Small interfering (si)-TMPO-AS1, pcDNA-TMPO-AS1,
miR-143-3p, anti-miR-143-3p and their negative controls (si-NC,
pcDNA, miR-NC and anti-miR-NC) were purchased from Shanghai
GenePharma Co., Ltd. All plasmids (100 nM) and (si)-TMPO-AS1 (50
nM), miR-143-3p (50 nM), anti-miR-143-3p (50 nM) and their controls
were transfected into SW480 and HCT15 cells using
Lipofectamine® 2000 reagent (Invitrogen; Thermo Fisher
Scientific, Inc.) according to the manufacturer's instructions. The
siRNA for TMPO-AS1 (si-TMPO-AS1, 5′-GAAGACUAGUGACCUAUAAUU-3′),
miR-143-3p mimic (5′-UGAGAUGAAGCACUGUAGCUC-3′), miR-143-3p mimic
inhibitor (anti-miR-143-3p, 5′-GAGCUACAGUGCUUCAUCUCA-3′). Cells
(1×105/well) were collected after 48 h for use in
subsequent experiments.
Reverse transcription-quantitative
(RT-q) PCR
Total RNA from 5×106 cells was separated
using TRIzol® reagent (Invitrogen; Thermo Fisher
Scientific, Inc.). TaqMan miRNA Reverse Transcription kit (Applied
Biosystems; Thermo Fisher Scientific, Inc.) was used to reverse
transcribe the total RNA into cDNA of miR-143-3p, and the M-MLV
Reverse Transcriptase, M-MLV 5X Reaction Buffer, dNTPs (Invitrogen;
Thermo Fisher Scientific, Inc.) were used to synthesize
first-strand cDNA of TMPO-AS1. The reaction volume was 12 µl. U6
snRNA and GAPDH served as the internal reference genes for
normalization of miR-143-3p and TMPO-AS1, respectively. SYBR Green
Real-time PCR kit (Invitrogen; Thermo Fisher Scientific, Inc.) was
used to perform RT-qPCR on an iQTM5 Multicolor Real-Time PCR
Detection System (Bio-Rad Laboratories, Inc.). The following
thermocycling conditions were used for PCR: 95°C for 10 min,
followed by denaturation at 95°C for 10 sec, annealing at 60°C for
60 sec, and extension at 72°C for 30 sec, with a total of 45
cycles. The expression levels of miR-143-3p and TMPO-AS1 were
calculated using the 2−ΔΔCq method (25). The following primers were used:
miR-143-3p: Forward, 5′-GUCACGAAGUAGAGU-3′ and reverse,
5′-CTCAACTGGTGTCGTGGA-3′; GAPDH: Forward,
5′-TGACTTCAACAGCGACACCCA-3′ and reverse,
5′-CACCCTGTTGCTGTAGCCAAA-3′; TMPO-AS1: Forward
5′-CAGTTTAAAAGGCGCTGGGG-3′ and reverse 5′-CCTTATCGGCGTCTAAGGGG′;
and U6: Forward, 5′-GCTTCGGCAGCACATATACTAA-3′ and reverse,
5′-AACGCTTCACGAATTTGCGT-3′. A total of three independent repeats
was performed.
MTT assay
MTT assay was used to detect cell proliferation.
Transfected cells in each well (2×103) were seeded into
96-well plates. After culture for 24, 48 and 72 h at 37°C, 15 µl
MTT solution was added into each well. Then, cells were incubated
for another 4 h at room temperature. Next, 150 µl DMSO solution was
added into each well to dissolve formazan for 10 min at 37°C. The
absorbance was detected using SpectraMax 360 pc microplate reader
(Molecular Devices, LLC) at a wavelength of 490 nm.
Western blot analysis
Cells were lysed in RIPA buffer (Cell Signaling
Technology, Inc.) and the proteins extracted. Then, the protein
concentration was detected using a bicinchoninic acid Protein assay
kit (Beyotime Institute of Biotechnology). Next, 10% SDS-PAGE was
used to separate the proteins (20 mg/lane), which were then
transferred onto a PVDF membrane (EMD Millipore). Next, 5% non-fat
milk was employed to block the membrane for 2 h at room
temperature. The membrane was incubated with the primary antibodies
against cyclin D1 (cat. no. 55506), p21 (cat. no. 2947), p27 (cat.
no. 3686), matrix metalloproteinase (MMP)-2 (cat. no. 40994), MMP-9
(cat. no. 13667), MMP-14 (cat. no. 13130) and GAPDH (cat. no. 5174)
(all 1:2,000; all Cell Signaling Technology, Inc.) at 4°C
overnight, and then incubated with the secondary antibody
HRP-conjugated goat anti-rabbit IgG; 1:2,000; Cell Signaling
Technology, Inc.) for 1 h at room temperature. The protein blots
were resolved using enhanced chemiluminescence reagents (Pierce;
Thermo Fisher Scientific, Inc.) according to the manufacturer's
instructions. In addition, Image Studio Lite software (version 3.1;
LI-COR Biosciences) was used to quantify the density of protein
bands, as previously described (26).
Cell migration and invasion
Transwell migration and invasion assay were used to
measure cell migration and invasion. 24-Transwell inserts (8 µm
pores; Corning Inc.) were used to assess cell migration, and
Transwell inserts coated with Matrigel (BD Biosciences) at 37°C for
6 h were applied for cell invasion measurement. The cells were
seeded onto the upper chamber of a Transwell supplemented with the
serum-free medium at 5.0×104 for migration and
1.0×105 for invasion and then incubated in a 5% of
CO2 atmosphere at 37°C for 24 h. The lower chamber was
filled with complete medium containing 10% FBS. Next, cells in the
upper chamber supplemented with medium containing 10% FBS were
removed. The numbers of migratory and invasive cells in the lower
chambers were fixed with 4% paraformaldehyde for 30 min at 37°C and
stained with 0.1% crystal violet for 10 min at room temperature.
The cells were counted in 5 random fields under a Leica DM3000
routine light microscope (magnification, ×100, Leica Microsystems
GmbH).
Luciferase reporter assay
The targeted sequence between TMPO-AS1 and
miR-143-3p was predicted using StarBase database (27). Fragments of TMPO-AS1 containing the
wild-type (WT) or mutant (MUT) were synthesized and cloned into
pMIR-REPORT™ (Thermo Fisher Scientific, Inc.) to construct the
vector of WT-TMPO-AS1 or MUT-TMPO-AS1, respectively, according to
the manufacturer's protocols. Next, the vector of WT-TMPO-AS1 or
MUT-TMPO-AS1 (100 ng) was co-transfected with miR-NC or miR-143-3p
into SW480 and HCT15 cells at a final concentration of 50 nM using
Lipofectamine® 2000 reagent (Invitrogen; Thermo Fisher
Scientific, Inc.), according to the manufacturer's instructions.
Following 36 h transfection, cells were harvested. The luciferase
activities were detected using a Dual-Luciferase Reporter Assay
system (Promega Corporation). The normalization of firefly
luciferase activity utilized the Renilla luciferase activity as the
control.
Tumor xenograft model
All animal experiments were approved by the Animal
Ethics Committee of Lanzhou University Second Hospital, China. A
total of 12 female BALB/c nude mice (age, 4–5 weeks; weight 15–18
g) were obtained from the Animal Center of Central South
University. Mice were maintained at a temperature of 18–23°C, 12-h
light/dark cycle, 45–65% humidity, and free access to water and
food. A total of 12 female BALB/c nude mice were randomly divided
into two groups (6 mice/group). The flanks of the mice were
injected with cells (1×106 cell/ml) transfected with
short hairpin (sh)-NC or sh-TMPO-AS1 (Shanghai GenePharma Co.,
Ltd.), respectively. Following injection, the tumor size (length
and width) was measured every week. After 5 weeks, the mice were
sacrificed and the tumor weight was measured. The tumor volume=0.5
× length × width2. The volume for the maximum tumor was
729 mm3 with the length for 18.0 mm and the width for
9.0 mm.
Statistical analysis
Data are presented as the mean ± standard deviation,
and analyzed using GraphPad Prism 7.0 (GraphPad Software, Inc.).
All experiments were performed in triplicate. Statistical
comparison of data was analyzed with an unpaired Student's t-test
and one-way ANOVA followed by Tukey's test. P<0.05 was
considered to indicate a statistically significant difference.
Results
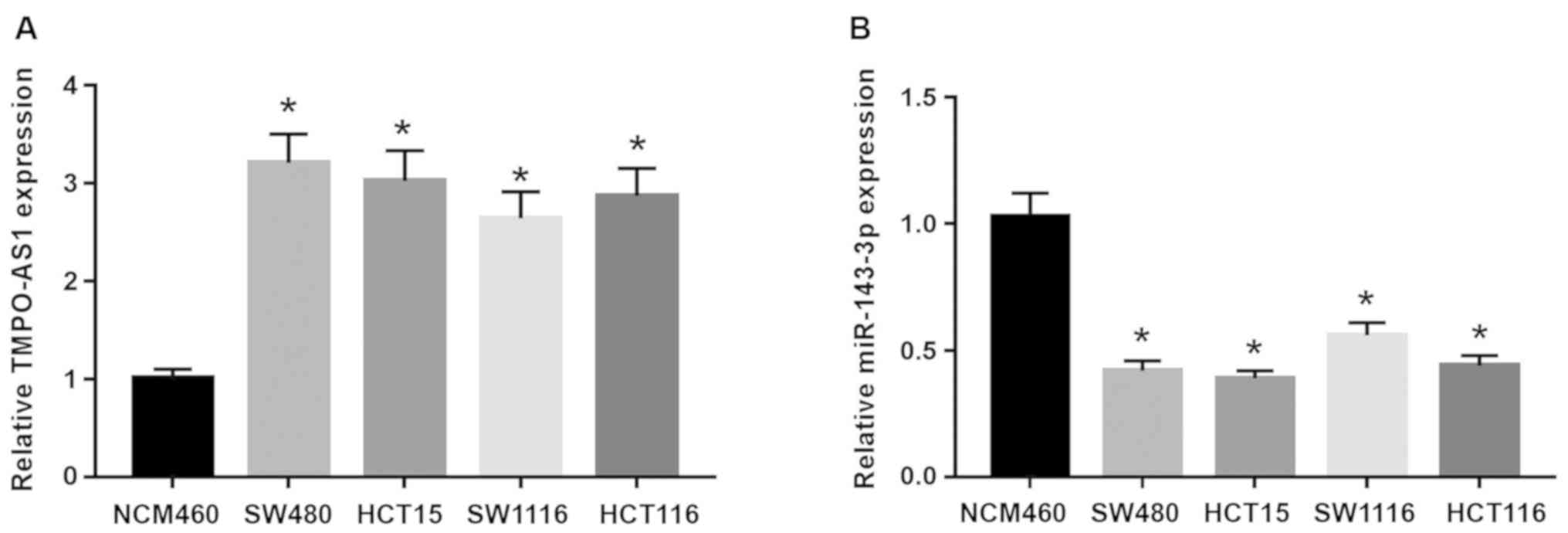
TMPO-AS1 expression is increased and
miR-143-3p expression is decreased in CRC cells
First, the expression levels of TMPO-AS1 and
miR-143-3p in CRC cell lines (SW480, HCT15, SW1116 and HCT116) and
normal connective mucosal epithelial cells (NCM460) were determined
using RT-qPCR. The data demonstrated that TMPO-AS1 was
significantly increased, while the expression of miR-143-3p was
significantly decreased, in SW480, HCT15, SW1116 and HCT116 cells
compared with NCM460 cells (Fig. 1A
and B). Thus, it was indicated that TMPO-AS1 and miR-143-3p may
serve crucial roles in CRC cells. To further investigate the
functions of TMPO-AS1 and miR-143-3p in CRC cells, SW480 and HCT15
CRC cell lines were selected for subsequent functional studies.
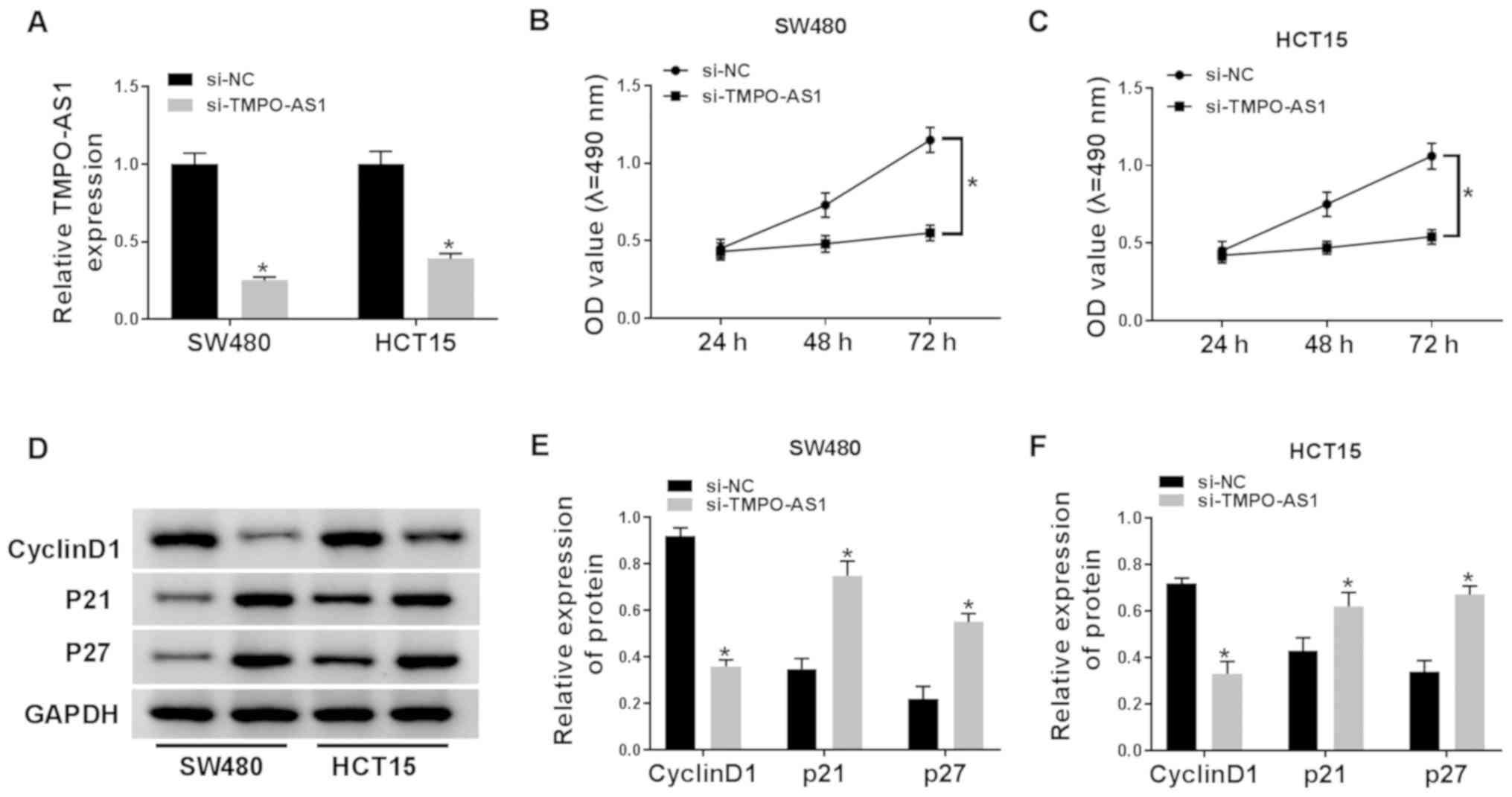
Knockdown of TMPO-AS1 suppresses cell
proliferation in CRC cells
si-NC or si-TMPO-AS1 was transfected into SW480 and
HCT15 cells. RT-qPCR results indicated that the expression of
TOMP-AS1 was significantly reduced in cells transfected with
si-TMPO-AS1 (Fig. 2A). Then, cell
proliferation was determined using a MTT assay, which was found to
be significantly decreased by TMPO-AS1 knockdown in SW480 and HCT15
cells (Fig. 2B and C). In
addition, western blot analysis demonstrated that the protein
expression of cyclin D1 was inhibited, while the protein expression
levels of p21 and p27 were increased, in SW480 and HCT15 cells
transfected with si-TMPO-AS1 compared with the si-NC (Fig. 2D-F). Therefore, these data
suggested that downregulation TMPO-AS1 inhibited cell proliferation
of CRC cells.
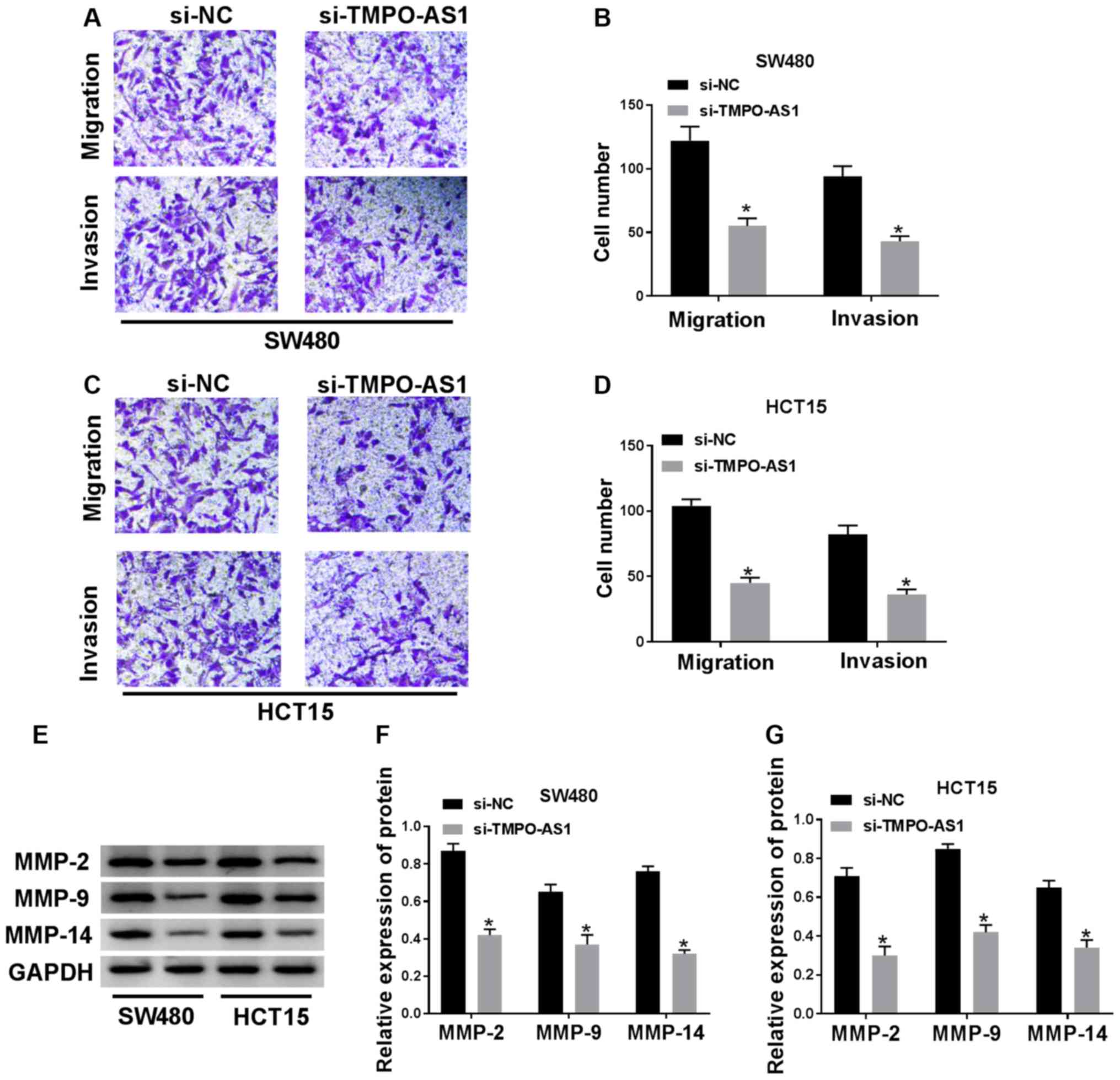
Knockdown of TMPO-AS1 reduces cell
migration and invasion in CRC cells
Transwell migration and invasion assays demonstrated
that cell migratory and invasive capacities of SW480 and HCT15
cells were suppressed by TMPO-AS1 knockdown (Fig. 3A-D). The MMP protein family is
closely associated with cell metastasis (28). Thus, MMP-2, MMP-9 and MMP-14
protein expression levels were determined in SW480 and HCT15 cells
transfected with si-NC or si-TMPO-AS1. It was demonstrated that
MMP-2, MMP-9 and MMP-14 protein expression levels were
significantly reduced by si-TMPO-AS1 transfection compared with the
si-NC group (Fig. 3E-G). Thus,
knockdown of TMPO-AS1 could inhibit cell migration and invasion in
CRC cells.
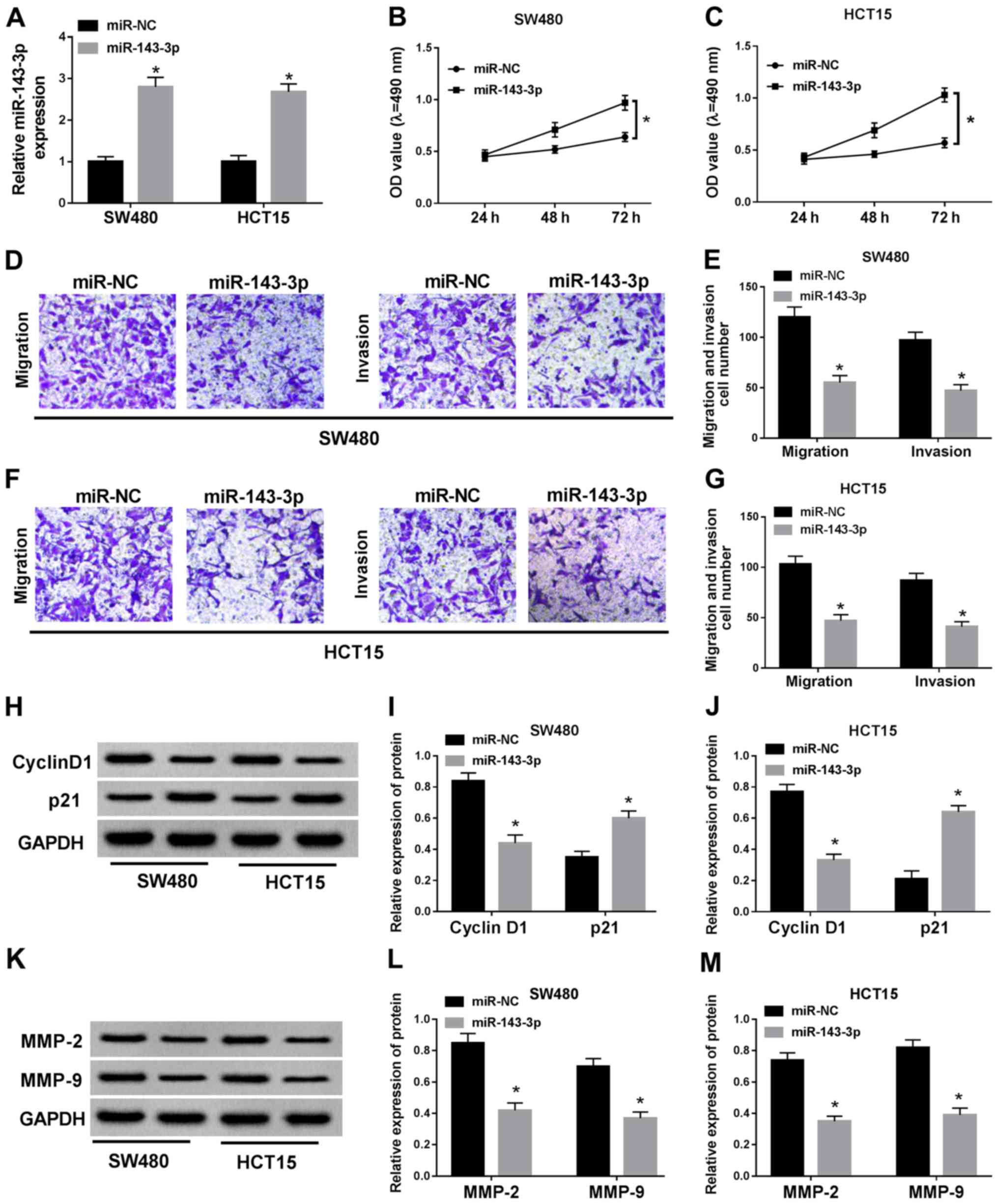
Overexpression of miR-143-3p inhibits
cell proliferation, migration and invasion of CRC cells
To confirm the function of miR-143-3p, miR-143-3p
was overexpressed in SW480 and HCT15 cells (Fig. 4A). It was found that miR-143-3p
overexpression significantly decreased proliferation of SW480 and
HCT15 cells (Fig. 4B and C). In
addition, cell migration and invasion were significantly inhibited
by overexpression of miR-143-3p in SW480 and HCT15 cells (Fig. 4E-G). The miR-143-3p mimic also
reduced the protein expression levels of cyclin D1, MMP-2 and
MMP-9, but promoted the protein expression of p21 (Fig. 4H-N). These results indicated that
overexpression of miR-143-3p could inhibit cell proliferation,
migration and invasion in CRC cells.
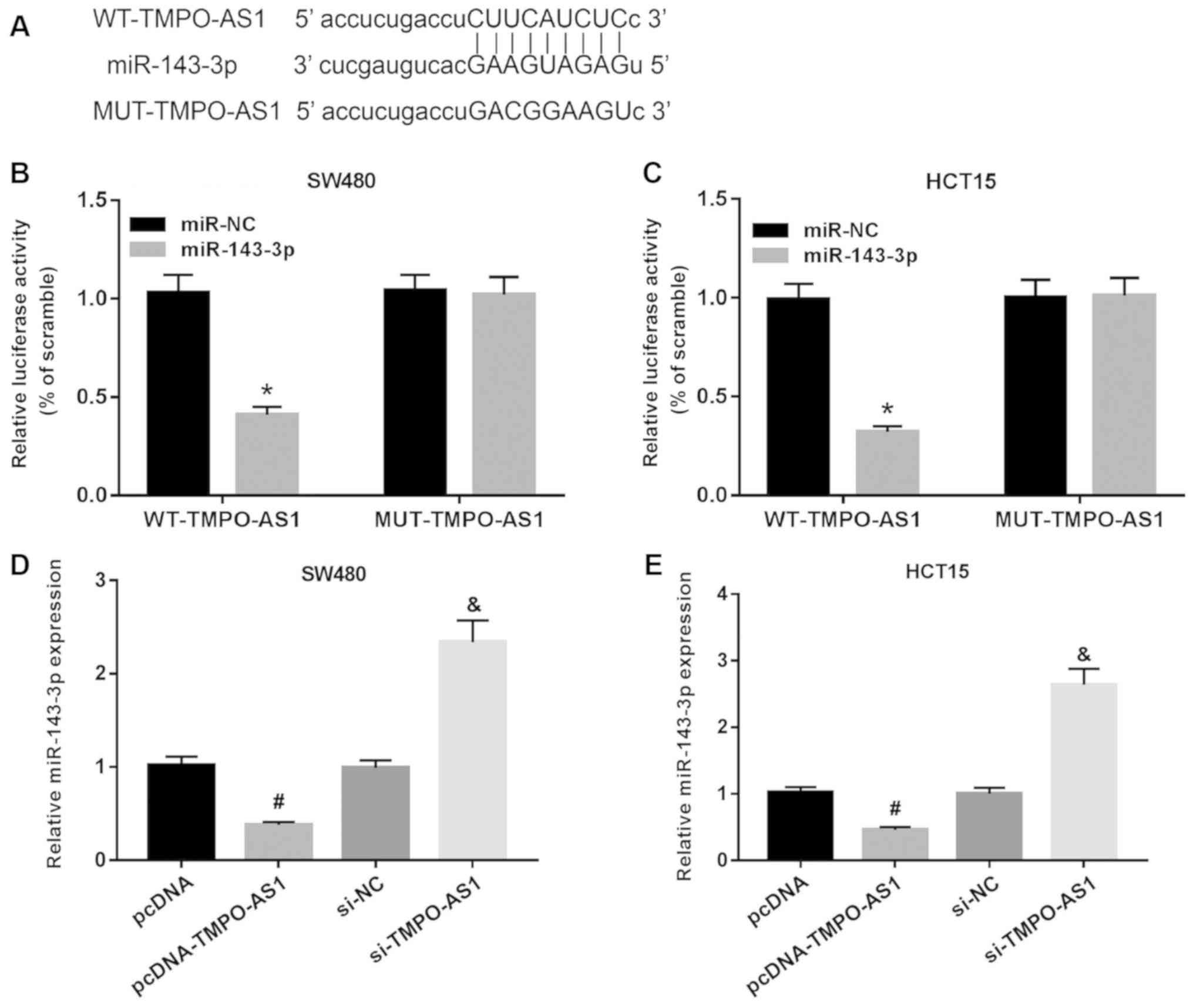
TMPO-AS1 directly targets miR-143-3p
in CRC cells
Bioinformatics analysis predicted that TMPO-AS1 had
complementary binding sites with miR-143-3p (Fig. 5A). A luciferase reporter assay was
performed to assess the relationship between miR-143-3p and
TMPO1-AS, and it was demonstrated that the miR-143-3p mimic
significantly reduced the luciferase activity of WT-TMPO-AS1, while
it had no effect on the luciferase activity of MUT-TMPO-AS1 in
SW480 or HCT15 cells (Fig. 5B and
C). As shown in Fig. S1A,
pcDNA-TMPO-AS1 could significantly upregulate the expression of
pcDNA-TMPO-AS1 in both SW480 and HCT15 cells. Moreover, miR-143-3p
expression was inhibited by TMPO-AS1 overexpression but promoted by
TMPO-AS1 knockdown in SW480 and HCT15 cells (Fig. 5D and E). These results suggested
that miR-143-3p was a target of TMPO-AS1 and negatively regulated
by TMPO-AS1 in CRC cells.
Knockdown of miR-143-3p reverses the
suppressive effects of si-TMPO-AS1 on cell proliferation, migration
and invasion in CRC cells
To further investigate the underlying regulatory
mechanism between miR-143-3p and TMPO-AS1, si-NC, si-TMPO-AS1,
si-TMPO-AS1 + anti-miR-NC or si-TMPO-AS1 + anti-miR-143-3p were
transfected into SW480 and HCT15 cells. The data demonstrated a
successful knockdown efficiency of anti-miR-143-3p in SW480 and
HCT15 cells (Fig. S1B).
RT-qPCR results indicated that miR-143-3p was
enhanced by TMPO-AS1 knockdown, which was significantly eliminated
by miR-143-3p inhibitor in SW480 and HCT15 cells (Fig. 6A). MTT assay results demonstrated
that cell proliferation inhibited by TMPO-AS1 knockdown was
partially blocked by miR-143-3p inhibitor (Fig. 6B and C). Moreover, the inhibitory
effects of TMPO-AS1 knockdown on cell migration and invasion were
eliminated by anti-miR-143-3p in SW480 and HCT15 cells (Fig. 6D-G). The decrease of cyclin D1
protein expression and the increase of p21 protein expression
caused by TMPO-AS1 knockdown were partially restored by
anti-miR-143-3p transfection in SW480 and HCT15 cells (Fig. 6H-J). In addition, the protein
expression levels of MMP-2 and MMP-9 were decreased by the
transfection of si-TMPO-AS1, but alleviated by anti-miR-143-3p in
SW480 and HCT15 cells (Fig. 6K-M).
Collectively, these results suggested that inhibition of TMPO-AS1
reduced cell proliferation, migration and invasion by targeting
miR-143-3p in CRC cells.
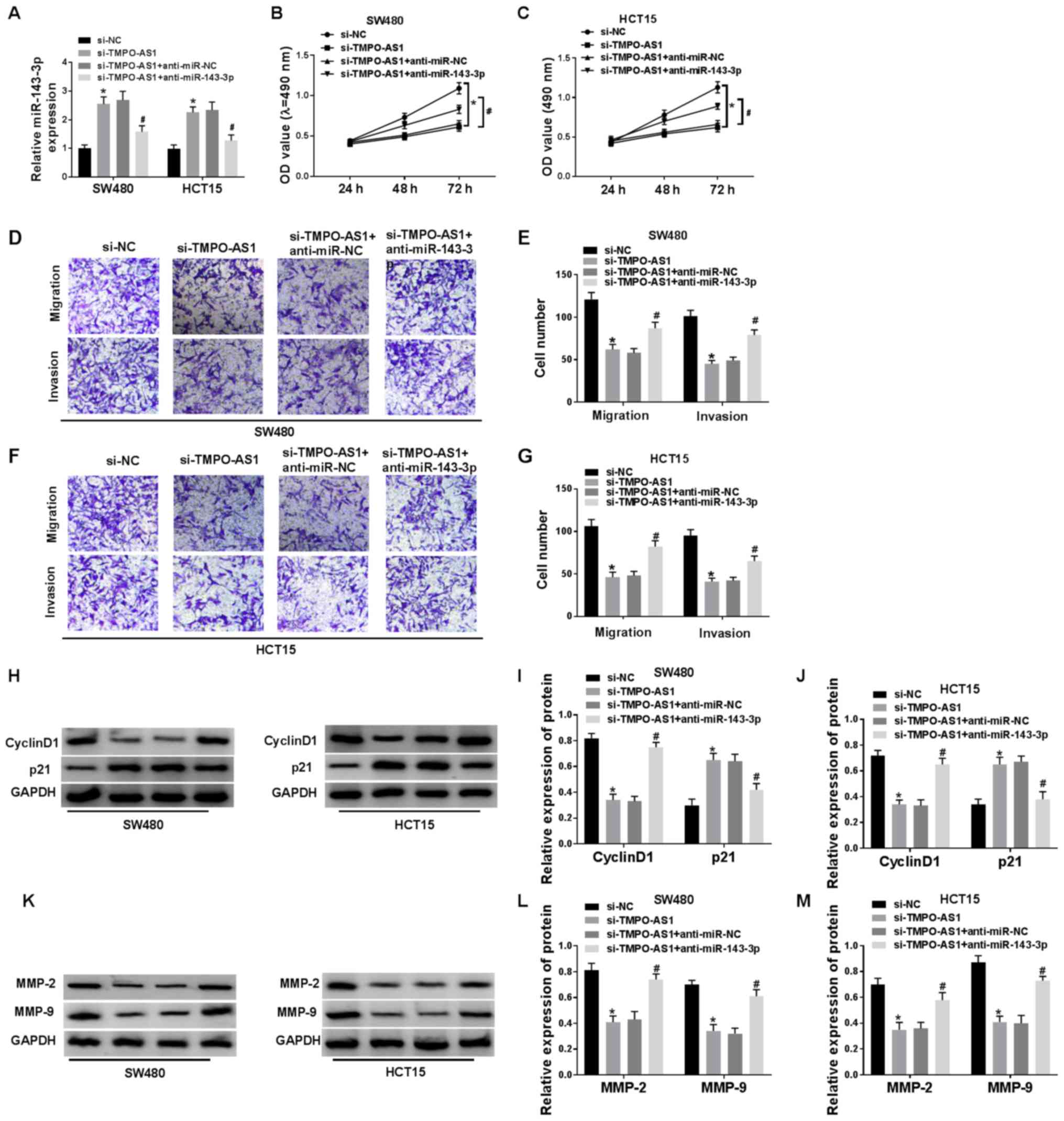 | Figure 6.Downregulation of miR-143-3p reverses
the suppressive effects of si-TMPO-AS1 on cell proliferation,
migration and invasion in CRC cells. (A) Expression of miR-143-3p
was detected in SW480 and HCT15 cells transfected with si-NC,
si-TMPO-AS1, si-TMPO-AS1 + anti-miR-NC and si-TMPO-AS1 +
anti-miR-143-3p via reverse transcription-quantitative PCR. Cell
proliferation was measured in si-NC, si-TMPO-AS1, si-TMPO-AS1 +
anti-miR-NC and si-TMPO-AS1 + anti-miR-143-3p groups in (B) SW480
and (C) HCT15 cells using a MTT assay. (D) Cell migration and
invasion were detected in si-NC, si-TMPO-AS1, si-TMPO-AS1 +
anti-miR-NC and si-TMPO-AS1 + anti-miR-143-3p groups in (E) SW480,
(F) as well as in and (G) HCT15 cells using Transwell assays. (H)
Western blotting results of cyclin D1 and p21 expression levels in
(I) SW480 and (J) HCT15 cells transfected with si-NC, si-TMPO-AS1,
si-TMPO-AS1 + anti-miR-NC and si-TMPO-AS1 + anti-miR-143-3p. (K)
Western blotting results of MMP-2 and MMP-9 in (L) SW480 and (M)
HCT15 cells transfected with si-NC, si-TMPO-AS1, si-TMPO-AS1 +
anti-miR-NC and si-TMPO-AS1 + anti-miR-143-3p. *P<0.05 vs.
si-NC; #P<0.05 vs. si-TMPO-AS1 + anti-miR-NC. miR,
microRNA; si, short interfering; CRC, colorectal cancer; NC,
negative control; MMP, matrix metalloproteinase; AS, antisense; OD,
optical density. |
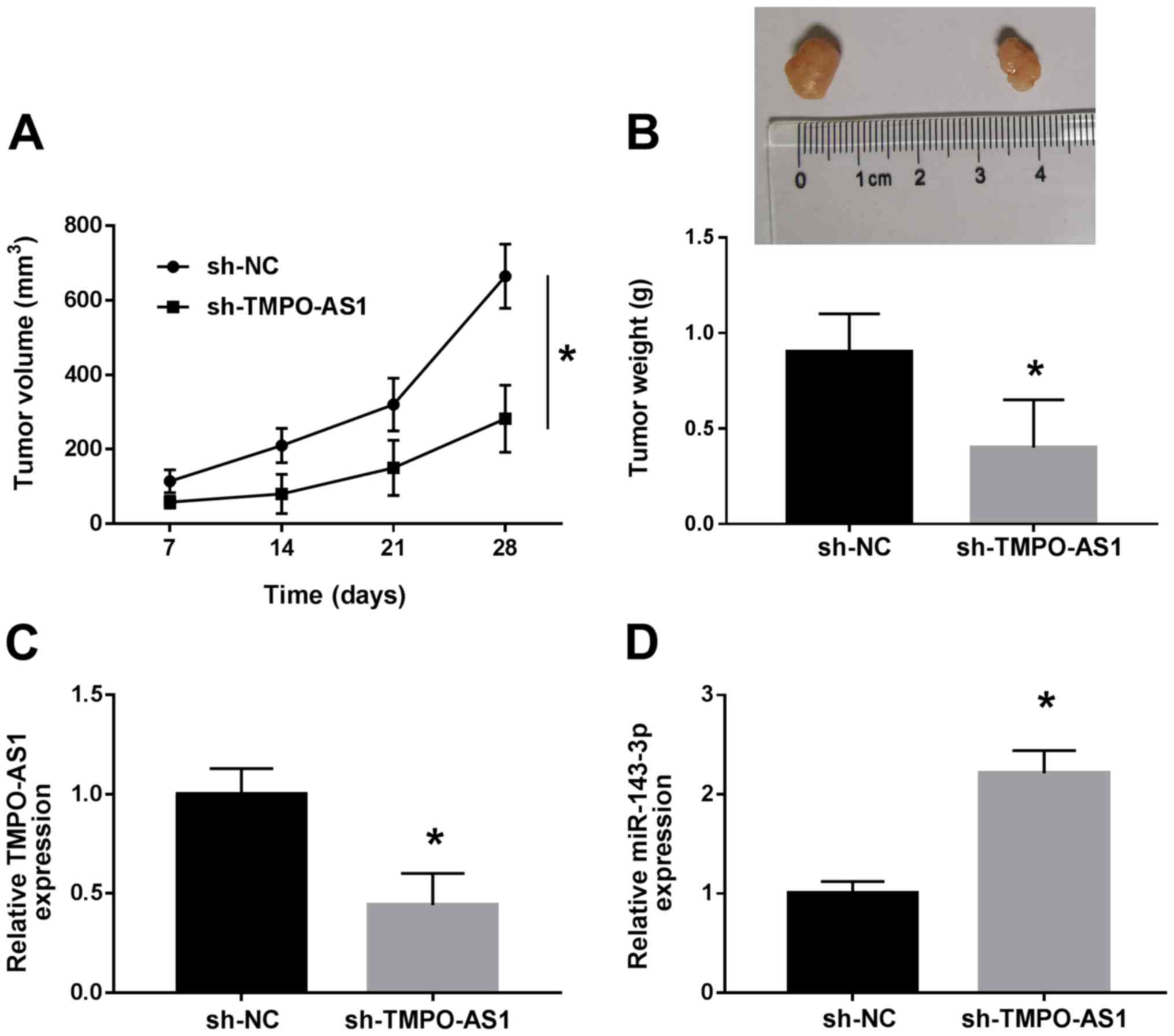
Knockdown of TMPO-AS1 suppresses tumor
growth in CRC in vivo
To further evaluate the effect of TMPO-AS1 on CRC
progression in vivo, animal experiments were performed. As
shown in Fig. 7A, the tumor volume
of sh-TMPO-AS1 group was significantly inhibited compared with the
sh-NC group. In addition, tumor weight was markedly decreased in
sh-TMPO-AS1 group compared with sh-NC group (Fig. 7B). Then, the expression levels of
TMPO-AS1 and miR-143-3p were detected in tumor tissues. The results
demonstrated that sh-TMPO-AS1 transfection inhibited TMPO-AS1
expression and promoted miR-143-3p expression (Fig. 7C and D). Thus, knockdown of
TMPO-AS1 could inhibit tumor growth in CRC in vivo.
Discussion
lncRNAs serve important roles in the development and
progression of multiple types of cancer (29). As a regulatory factor, lncRNA can
inhibit or promote the development of cancer cell and tumor
formation (30–33). For instance, upregulated lncRNA
DLX6-AS1 enhances cell proliferation and invasion by regulating
miR-181b in pancreatic cancer (34), while lncRNA ATB is involved in cell
progression in CRC (35). In
addition, certain lncRNAs are closely associated with the prognosis
of CRC and could serve as biomarkers (36–38).
However, the function of TMPO-AS1 in CRC progression remains
unknown. A previous study reported that TMPO-AS1 was upregulated in
prostate cancer tissues, and that overexpression of TMPO-AS1
promotes the proliferation and migration of prostate cancer cells
(39). Moreover, TMPO-AS1
possesses a tumor-promoting action in osteosarcoma (24) Consistent with these studies, the
present results demonstrated the promoting effect of TMPO-AS1 in
CRC, as evidenced by inhibition on cell proliferation, migration
and invasion in CRC cells after TMPO-AS1 knockdown.
lncRNA can target miRNAs to participate in the
regulation of cell processes and metabolism (13). The present study found that
TMPO-AS1 directly targeted miR-143-3p. Accumulating evidence
indicates that miR-143-3p is involved in cell proliferation and
apoptosis in various of types of cancer, including breast cancer,
hepatocellular carcinoma and esophageal squamous cell carcinoma
(40–42). Furthermore, miR-143-3p is
downregulated in cervical cancer, colorectal cancer and breast
cancer. It has also been shown that overexpression of miR-143-3p
inhibits cell progression (40,43,44).
In concurrence, the present study found that miR-143-3p was
decreased in CRC cells, and that overexpression of miR-143-3p
significantly suppressed cell proliferation, migration and invasion
in CRC cells.
The results of the present recovery experiments
indicated that TMPO-AS1 affected cell proliferation, migration and
invasion by targeting miR-143-3p. Therefore, the
TMPO-AS1/miR-143-3p axis may be an important regulatory mechanism
for CRC cell proliferation and metastasis. Additionally, it was
demonstrated that TMPO-AS1 knockdown repressed tumor growth via
regulating miR-143-3p in vivo.
In conclusion, the current study demonstrated the
function and regulatory network of TMPO-AS1 in CRC. The results
suggested that TMPO-AS1 expression levels were increased in CRC
cells. Knockdown of TMPO-AS1 impeded cell proliferation, migration
and invasion in CRC cells, and inhibited tumor growth in CRC in
vivo, by regulating miR-143-3p. These findings improved the
understanding of the regulatory mechanism of CRC and may provide a
new target for the treatment of CRC.
Supplementary Material
Supporting Data
Acknowledgements
Not applicable.
Funding
No funding was received.
Availability of data and materials
All data generated or analyzed during this study are
included in this published article.
Authors' contributions
LZ and YL designed the study and wrote the
manuscript. LZ, YL and ALS performed the experiments and analyzed
the data. LZ edited the manuscript. All authors read and approved
the final manuscript.
Ethics approval and consent to
participate
All animal experiments were approved by the Animal
Ethics Committee of Lanzhou University Second Hospital.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Baena R and Salinas P: Diet and colorectal
cancer. Maturitas. 80:3245–264. 2015. View Article : Google Scholar
|
|
2
|
Bray F, Ferlay J, Soerjomataram I, Siegel
RL, Torre LA and Jemal A: Global cancer statistics 2018: GLOBOCAN
estimates of incidence and mortality worldwide for 36 cancers in
185 countries. CA Cancer J Clin. 68:394–424. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Wang XC, Du LQ, Tian LL, Wu HL, Jiang XY,
Zhang H, Li DG, Wang YY, Wu HY, She Y, et al: Expression and
function of miRNA in postoperative radiotherapy sensitive and
resistant patients of non-small cell lung cancer. Lung Cancer.
72:92–99. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Qi W, Liang W, Jiang H and Waye MM: The
function of miRNA in hepatic cancer stem cell. Biomed Res Int.
2013:3589022013. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Bao AD, Liu CQ, Hong-Mei WU, Liu S, Guan
WJ and Yue-Hui MA: Association analysis between function of miRNA
and formation of cancer. Chin Anim Husbandry Veterinary Medicine.
2008.
|
|
6
|
Dhamija S and Diederichs S: From junk to
master regulators of invasion: LncRNA functions in migration, EMT
and metastasis. Int J Cancer. 139:269–280. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Lian Y, Xia-Yu LI, Tang YY, Yang LT,
Xiao-Ling LI, Xiong W, Gui-Yuan LI and Zeng ZY: Long non-coding
RNAs function as competing endogenous RNAs to regulate cancer
progression. Prog Biochem Biophys. 2016.
|
|
8
|
Fang Q, Chen XY and Zhi XT: Long
non-coding RNA (LncRNA) urothelial carcinoma associated 1 (UCA1)
increases multi-drug resistance of gastric cancer via
downregulating miR-27b. Med Sci Monit. 22:3506–3513. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Liu H, Yang Z, Ma J and Fan D: Function of
miRNA in controlling drug resistance of human cancers. Curr Drug
Targets. 14:1118–1127. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Dou J, Ni Y, He X, Wu D, Li M, Wu S, Zhang
R, Guo M and Zhao F: Decreasing lncRNA HOTAIR expression inhibits
human colorectal cancer stem cells. Am J Transl Res. 8:98–108.
2016.PubMed/NCBI
|
|
11
|
Han P, Li JW, Zhang BM, Lv JC, Li YM, Gu
XY, Yu ZW, Jia YH, Bai XF, Li L, et al: The lncRNA CRNDE promotes
colorectal cancer cell proliferation and chemoresistance via
miR-181a-5p-mediated regulation of Wnt/β-catenin signaling. Mol
Cancer. 16:92017. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Peng W, Wang Z and Fan H: LncRNA NEAT1
impacts cell proliferation and apoptosis of colorectal cancer via
regulation of Akt signaling. Pathol Oncol Res. 23:651–656. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Zhang W, Yuan W, Song J, Wang S and Gu X:
LncRna CPS1-IT1 suppresses cell proliferation, invasion and
metastasis in colorectal cancer. Cell Physiol Biochem. 44:567–580.
2017. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Chen Y, Han X, Yin X, Zhou Y and Wu T:
Decreased expression of miR-132 in CRC tissues and its inhibitory
function on tumor progression. Open Life Sci. 11:130–135. 2016.
View Article : Google Scholar
|
|
15
|
Guo H, Hu X, Ge S, Qian G and Zhang J:
Regulation of RAP1B by miR-139 suppresses human colorectal
carcinoma cell proliferation. Int J Biochem Cell Biol.
44:1465–1472. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Ma K, Pan X, Fan P, He Y, Gu J, Wang W,
Zhang T, Li Z and Luo X: Loss of miR-638 in vitro promotes cell
invasion and a mesenchymal-like transition by influencing SOX2
expression in colorectal carcinoma cells. Mol Cancer. 13:1182014.
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Luo H, Zou J, Dong Z, Zeng Q, Wu D and Liu
L: Up-regulated miR-17 promotes cell proliferation, tumour growth
and cell cycle progression by targeting the RND3 tumour suppressor
gene in colorectal carcinoma. Biochem J. 442:311–321. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Gao Y, Meng H, Liu S, Hu J, Zhang Y, Jiao
T, Liu Y, Ou J, Wang D, Yao L, et al: LncRNA-HOST2 regulates cell
biological behaviors in epithelial ovarian cancer through a
mechanism involving microRNA let-7b. Hum Mol Genet. 24:841–852.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Paraskevopoulou MD and Hatzigeorgiou AG:
Analyzing MiRNA-LncRNA interactions. Methods Mol Biol.
1402:271–286. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Jalali S, Bhartiya D, Lalwani MK,
Sivasubbu S and Scaria V: Systematic transcriptome wide analysis of
lncRNA-miRNA interactions. PLoS One. 8:e538232013. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Ye S, Yang L, Zhao X, Song W, Wang W and
Zheng S: Bioinformatics method to predict two regulation mechanism:
TF-miRNA-mRNA and lncRNA-miRNA-mRNA in pancreatic cancer. Cell
Biochem Biophys. 70:1849–1858. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Zhao L, Kong H, Sun H, Chen Z, Chen B and
Zhou M: LncRNA-PVT1 promotes pancreatic cancer cells proliferation
and migration through acting as a molecular sponge to regulate
miR-448. J Cell Physiol. 233:4044–4055. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Mohammadrezakhani H, Baradaran B,
Shanehbandi D, Asadi M, Hashemzadeh S, Hajiasgharzadeh K and
Safaralizadeh R: Overexpression and clinicopathological correlation
of long noncoding RNA TMPO-AS1 in colorectal cancer patients. J
Gastrointest Cancer. Nov 25–2019.(Epub ahead of print).
|
|
24
|
Cui H and Zhao J: LncRNA TMPO-AS1 serves
as a ceRNA to promote osteosarcoma tumorigenesis by regulating
miR-199a-5p/WNT7B axis. J Cell Biochem. 121:2284–2293. 2020.
View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Saluzzo J, Hallman KM, Aleck K, Dwyer B,
Quigley M, Mladenovik V, Siebert AE and Dinda S: The regulation of
tumor suppressor protein, p53, and estrogen receptor (ERα) by
resveratrol in breast cancer cells. Genes Cancer. 7:414–425.
2016.PubMed/NCBI
|
|
27
|
Chen R, Li WX, Sun Y, Duan Y, Li Q, Zhang
AX, Hu JL, Wang YM and Gao YD: Comprehensive analysis of lncRNA and
mRNA expression profiles in lung cancer. Clin Lab. 63:313–320.
2017. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Wang X, Hu Y, Cui J, Zhou Y and Chen L:
Coordinated targeting of MMP-2/MMP-9 by miR-296-3p/FOXCUT exerts
tumor-suppressing effects in choroidal malignant melanoma. Mol Cell
Biochem. 445:25–33. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Bhan A, Soleimani M and Mandal SS: Long
noncoding RNA and cancer: A new paradigm. Cancer Res. 77:3965–3981.
2017. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Cui Y, Zhang F, Zhu C, Geng L, Tian T and
Liu H: Upregulated lncRNA SNHG1 contributes to progression of
non-small cell lung cancer through inhibition of miR-101-3p and
activation of Wnt/β-catenin signaling pathway. Oncotarget.
8:17785–17794. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Zhang G, Li S, Lu J, Ge Y, Wang Q, Ma G,
Zhao Q, Wu D, Gong W, Du M, et al: LncRNA MT1JP functions as a
ceRNA in regulating FBXW7 through competitively binding to
miR-92a-3p in gastric cancer. Mol Cancer. 17:872018. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Zhang J, Li XY, Hu P and Ding YS: LncRNA
NORAD contributes to colorectal cancer progression by inhibition of
miR-202-5p. Oncol Res. Feb 22–2018 (Epub ahead of print).
View Article : Google Scholar
|
|
33
|
Zhang M, Wu WB, Wang ZW and Wang XH:
LncRNA NEAT1 is closely related with progression of breast cancer
via promoting proliferation and EMT. Eur Rev Med Pharmacol Sci.
21:1020–1026. 2017.PubMed/NCBI
|
|
34
|
An Y, Chen XM, Yang Y, Mo F, Jiang Y, Sun
DL and Cai HH: LncRNA DLX6-AS1 promoted cancer cell proliferation
and invasion by attenuating the endogenous function of miR-181b in
pancreatic cancer. Cancer Cell Int. 18:1432018. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Iguchi T, Uchi R, Nambara S, Saito T,
Komatsu H, Hirata H, Ueda M, Sakimura S, Takano Y, Kurashige J, et
al: A long noncoding RNA, lncRNA-ATB, is involved in the
progression and prognosis of colorectal cancer. Anticancer Res.
35:1385–1388. 2015.PubMed/NCBI
|
|
36
|
Peng Q, Lei D, Lin W, Zhou X and Xiang D:
Abstract B42: A two-lncRNA signature in serous exosomes serves as a
new biomarker for colorectal cancer diagnosis. Cancer Res. 76 (6
Suppl):B422016.
|
|
37
|
Shi D, Zheng H, Zhuo C, Peng J, Li D, Xu
Y, Li X, Cai G and Cai S: Low expression of novel lncRNA
RP11-462C24.1 suggests a biomarker of poor prognosis in colorectal
cancer. Med Oncol. 31:312014. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Zheng HT, Shi DB, Wang YW, Li XX, Xu Y,
Tripathi P, Gu WL, Cai GX and Cai SJ: High expression of lncRNA
MALAT1 suggests a biomarker of poor prognosis in colorectal cancer.
Int J Clin Exp Pathol. 7:3174–3181. 2014.PubMed/NCBI
|
|
39
|
Huang W, Su X, Yan W, Kong Z, Wang D,
Huang Y, Zhai Q, Zhang X, Wu H, Li Y, et al: Overexpression of
AR-regulated lncRNA TMPO-AS1 correlates with tumor progression and
poor prognosis in prostate cancer. Prostate. 78:1248–1261. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Li D, Hu J, Song H, Xu H, Wu C, Zhao B,
Xie D, Wu T, Zhao J and Fang L: miR-143-3p targeting LIM domain
kinase 1 suppresses the progression of triple-negative breast
cancer cells. Am J Transl Res. 9:2276–2285. 2017.PubMed/NCBI
|
|
41
|
Chen L, Yao H, Wang K and Liu X: Long
non-coding RNA MALAT1 regulates ZEB1 expression by sponging
miR-143-3p and promotes hepatocellular carcinoma progression. J
Cell Biochem. 118:4836–4843. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
He Z, Yi J, Liu X, Chen J, Han S, Jin L,
Chen L and Song H: MiR-143-3p functions as a tumor suppressor by
regulating cell proliferation, invasion and epithelial-mesenchymal
transition by targeting QKI-5 in esophageal squamous cell
carcinoma. Mol Cancer. 15:512016. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Liu M, Jia J, Wang X, Liu Y, Wang C and
Fan R: Long non-coding RNA HOTAIR promotes cervical cancer
progression through regulating BCL2 via targeting miR-143-3p.
Cancer Biol Ther. 19:391–399. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Ding X, Du J, Mao K, Wang X, Ding Y and
Wang F: MicroRNA-143-3p suppresses tumorigenesis by targeting
catenin-δ1 in colorectal cancer. OncoTargets Ther. 12:3255–3265.
2019. View Article : Google Scholar
|