Introduction
Melanoma is the second most dangerous malignant
tumor type and ranks second only to pancreatic cancer (1). Moreover, melanoma is characterized by
early metastasis and high mortality rates. In recent years, the
incidence of numerous malignant tumors has decreased from 40 to 20%
worldwide, but the incidence of malignant melanoma remains high
(2). Epithelial-mesenchymal
transition (EMT) has a critical role in the process of cell
remodeling during tumorigenesis (3). Furthermore, previous studies have
reported that EMT plays a key role in the metastasis of malignant
tumors (4–6). However, as the metastasis of melanoma
has not been effectively controlled (7), it is important to identify the
mechanism via which the EMT process is promoted.
Cell adhesion molecule 1 (CADM1), which is regarded
as a suppressor gene in the tumorigenesis of non-small cell lung
cancer, has been identified as a diagnostic marker for adult T-cell
leukemia and lymphoma (8). In
addition, it has been previously reported that CADM1 plays an
inhibitory role in the progression, migration and invasion of
cancer, such as cervical cancer and liver cancer (9–11),
indicating that CADM1 is an inhibitor of tumor cell malignant
features. You et al (12)
showed that CADM1 was differentially expressed in melanoma, but its
function in melanoma remains unknown. Moreover, microRNAs (miRNAs)
act as imperfect sequence guides to recruit a ribonucleoprotein
complex to the complementary RNA (13). Furthermore, miRNA expression
profiles differ between different stages of cancer and healthy
tissues, and previous studies have used miRNAs as diagnostic
markers, either alone or in combination with other known biomarkers
(14–16). Preliminary studies examining miRNA
expression also used tissues to determine the functional and
diagnostic roles of miRNAs (13,17,18),
which have been reported to regulate the development and
progression of cancer (19,20).
It has also been revealed that miRNAs (miRs) can promote or inhibit
the EMT process in cancer types (21–23).
For instance, miR-214 is a member of the miRNA family, and it can
induce immune suppression and promote the occurrence of cancer
(24), thus demonstrating that
miR-214 plays a key role in tumorigenesis. In addition, Mirzaei
et al (25) revealed that
miR-214 is differentially expressed in melanoma. However, the
relationship between CADM1 and miR-214 is not fully understood.
Based on these previous findings, the aims of the
present study were to investigate the mechanism via which miR-214
regulates the EMT process in melanoma, and to identify the
relationship between CADM1 and miR-214. Thus, the present results
may provide a potential novel target for treating melanoma.
Materials and methods
Cell lines and cell culture
The PIG-1, WM-266-4, A2058 and A375 cell lines were
purchased from the Cell Bank of Chinese Academy of Science, and
cultured in 90% RPMI-1640 (Thermo Fisher Scientific, Inc.) with 10%
FBS (Thermo Fisher Scientific, Inc.) and maintained at 37°C in a
humidified incubator containing 5% CO2. The BR-V-108
plasmid vector (Shanghai Biosciences Co., Ltd.) and competent TOP10
E. coli cells (Shanghai Biosciences Co., Ltd.) were
used.
Vector construction and cell
transfection
RNA interference target sequences were designed
using the CADM1 gene as a template to construct a target
gene RNA interference 5′-CCACAGGACAATGCTGAACTT-3′ lentiviral vector
(Shanghai Biosciences Co., Ltd.). A single-stranded DNA oligo
containing the interference sequence was synthesized, and the
obtained lyophilized powder was dissolved in an annealing buffer
(final concentration, 100 M; Beyotime Institute of Biotechnology)
and incubated in a water bath at 90°C for 15 min. After naturally
cooling to room temperature, a double-stranded oligo with overhang
ends was formed, which was then directly ligated into the digested
lentiviral vector through restriction sites at both ends. A 50 µl
reaction system was prepared according to the NEB (New England
Biolabs, Inc.) instructions, the BR-V-108 vector was linearized by
double digestion and the ligation product was introduced into
prepared TOP10 E. coli (Tiangen Biotech Co., Ltd.) competent
cells. The sequencing results were compared with those of the
correct clones and the plasmid was isolated.
miR-214 mimics (upregulation of miR-214, a
chemically modified single-stranded RNA that can mimic and enhance
the function of endogenous miR-214) and miR-214 inhibitor
(downregulation of miR-214, a chemically modified single-stranded
RNA that can specifically target miR-214 in melanoma cells), as
well as their corresponding negative controls (NCs), were designed
by Shanghai GenePharma Co., Ltd. For cell transfection, melanoma
cells (1.5×105/well) in a 6-well plate were transfected
with 200 pmol CADM1 short hairpin RNA (shRNA), or 100 pmol miR-214
mimics, inhibitor or their NCs using Lipofectamine® 2000
reagent (Thermo Fisher Scientific, Inc.) for 48 h according to the
manufacturer's instructions. Transfected cells were incubated for
an additional 24 h at 4°C and then used to test the cell viability.
The sequences were as follows: miR-214 mimics,
5′-UGCCUGUCUACACUUGCUGUGC-3′; miR-214 inhibitor,
5′-ACUGCCUGUCUGUGCCUGCUGU-3′; CADM1 siRNA sense,
5′-CACCGCAGATGACTTATCCTCTACAATTCAAGAGATTGTAGAGGATAAGTCATCTGTTTTTTG-3′
and anti-sense,
5′-GATCCAAAAAACAGATGACTTATCCTCTACAATCTCTTGAATTGTAGAGGATAAGTCATCTGC-3′.
Reverse transcription-quantitative PCR
(RT-qPCR)
Total RNA was extracted from melanoma cell lines
using TRIzol® reagent (Thermo Fisher Scientific, Inc.)
according to the manufacturer's protocol. First-strand cDNA was
synthesized using the PrimeScript RT reagent kit (Takara Bio, Inc.)
according to the manufacturer's protocol. RT-qPCR was performed in
an ABI7500 system (Thermo Fisher Scientific, Inc.) using SYBR-Green
(Takara Bio, Inc.) methods. RT-qPCR was performed in triplicate
using the following protocol: Initial denaturation for 2 min at
94°C, followed by 35 cycles for 30 sec at 94°C and 45 sec at 55°C.
The primers for all the genes were obtained from Shanghai
GenePharma Co., Ltd. Relative gene expression was quantified using
the 2−ΔΔCq method (26). The primer sequences are listed in
Table I.
 | Table I.Primer sequences used in the present
study. |
Table I.
Primer sequences used in the present
study.
| Gene | Sequence |
|---|
| miR-214 | F:
5′-AGCACAGCAGGCACAGACA-3′ |
|
| R:
5′-CAGTGCGTGTCGTGGAGT-3′ |
| U6 | F:
5′-CGCTTCGGCAGCACACATATAC-3′ |
|
| R:
5′-CAGGGGCCATGCTAATCTT-3′ |
| CADM1 | F:
5′-GGTGATGGGCAGAATCTGTT-3′ |
|
| R:
5′-AGGCCTGAAGTCCCTGAAAT-3′ |
| E-cadherin | F:
5′-TCACATCCTACACTGCCCAG-3′ |
|
| R:
5′-AGTGTCCCTGTTCCAGTAGC-3′ |
| Vimentin | F:
5′-GGACCAGCTAACCAACGACA-3′ |
|
| R:
5′-AAGGTCAAGACGTGCCAGAG-3′ |
| Slug | F:
5′-ACGCCTCCAAAAAGCCAAAC-3′ |
|
| R:
5′-ACAGTGATGGGGCTGTATGC-3′ |
| GAPDH | F:
5′-CCACCCATGGCAAATTCCATGGCA-3′ |
|
| R:
5′-TCTAGACGGCAGGTCAGGTCCAC-3′ |
Mature miRNAs were quantified using miRNA assays.
The protocol was as following: cDNA was synthesized from 5–10 µg of
total RNA using the microRNA cDNA Synthesis kit (Takara Bio, Inc.).
Then, PCR assays using were performed using SYBR premix Ex Taq II
kit (Takara Bio, Inc.). RT-qPCR conditions used were as follows:
Initial denaturation for 10 min at 95°C, followed by 40 cycles of
15 sec at 95°C and 1 min at 60°C. Relative miRNA and mRNA
expression levels were measured and normalized to U6 and GAPDH
expression, respectively.
Western blot analysis
To investigate EMT-related protein expression
levels, A375 or A2058 cells were collected and lysed using RIPA
lysis buffer (Cell Signaling Technology, Inc.) on ice according to
the manufacturer's instructions. Total cellular proteins (40
µg/lane) were quantified by bicinchoninic protein assay kit
(Beyotime Institute of Biotechnology), and then subjected to 10%
SDS-PAGE for western blotting. Proteins were transferred to PVDF
membranes, the blots were incubated at room temperature with 5% BSA
(Thermo Fisher Scientific, Inc.) in TBST (0.5% Tween-20) for 60 min
and then incubated overnight at 4°C on a rocker with the following
primary antibodies: Anti-CADM1 (1:1,000; cat. no. ab3910; Abcam),
anti-E-cadherin (1:3,000; cat. no. ab194982; Abcam), anti-vimentin
(1:3,000; cat. no. ab92547; Abcam), anti-Slug (1:1,000; cat. no.
ab51772; Abcam) and anti-GAPDH (1:3,000; cat. no. ab181602;
Bioworld Technology, Inc.). After washing three times with TBST for
5 min, the membranes were incubated with horseradish
peroxidase-conjugated goat anti-rabbit IgG polyclonal secondary
antibody (1:3,000; ab191866; Beyotime Institute of Biotechnology)
at room temperature for 1 h. An ECL+ western blotting
system kit (GE Healthcare Life Science) was used for color
development. The density of the bands was measured using ImageJ
software (version 6.0; National Institutes of Health). Signals were
detected with enhanced chemiluminescence using GAPDH as the
internal standard (Kodak).
Dual-luciferase reporter assay
The downstream target of CADM1 was predicted using
Starbase 2.0 (Sun Yat-sen University; http://starbase.sysu.edu.cn/). Both wild-type and
mutant constructs of CADM1 were cloned into the pmirGLO
Dual-Luciferase miRNA target expression vector (Promega
Corporation). A375 or A2058 cells (5×103 cells per well)
were seeded in a 24-well plate and co-transfected with wild-type or
mutant CADM1 3′untranslated region and mimic NC or miR-214 mimics
using Lipofectamine® 3000 (Thermo Fisher Scientific,
Inc.). Transfection and harvest efficiencies were controlled using
the pmirGLO reporter as an internal control. Cells were collected
48 h post-transfection and analyzed using the dual-luciferase
reporter assay system (Promega Corporation). The data were
quantified via normalizing to Renilla luciferase
activity.
MTT assay
An MTT assay was performed to investigate the effect
of miR-214 and CADM1 on melanoma cell viability. A375 and A2058
cells (5×103 cells per well) were seeded on 96-well
plates and incubated at 37°C for 24, 48 and 72 h. Then, 20 µl MTT
solution (Sigma-Aldrich; Merck KGaA) was added to each well and
incubated at room temperature for another 4 h. Then, the culture
medium with MTT was discarded, 150 µl DMSO was added and cells were
agitated for 10 min at room temperature. The absorbance was
measured at 490 nm with a standard microplate reader, and cell
viability was analyzed.
Wound healing assay
A375 and A2058 cells (5×104 cells per
well) were plated in a 6-well plate for culture. When cells were at
90% confluency, the medium was replaced with RPMI-1640 (Thermo
Fisher Scientific, Inc.) for the scratch test, and the slide was
pushed up to form a scratch. Then, 0.5% FBS was added and cells
were imaged using a light microscope (magnification, ×100). Cells
were incubated at 37°C in a 5% CO2 incubator for 48 h
and then assessed using ImageJ software (version 6.0; National
Institutes of Health). The cell migration rate of each group was
calculated based on the images acquired after the scratch.
Transwell assay
For the invasion assay, the upper chamber
(polycarbonic membrane; diameter 6.5 mm; pore size, 8 µm; Corning,
Inc.) was pretreated with 100 µl Matrigel (BD Biosciences) at 37°C
for 30 min, and melanoma cells (1×105 cells) in FBS-free
medium (RPMI-1640) were seeded into the upper chamber. The lower
chamber contained RPMI-1640 supplemented with 10% FBS. Then, cells
were incubated at 37°C for 24 h. The cells that attached to the
underside of the membrane were fixed using 5% glutaraldehyde at 4°C
for 30 min and stained with a 0.5% crystal violet solution at room
temperature for 30 min. Subsequently, images were captured and the
number of invading cells was counted under a light microscope
(magnification, ×100).
Statistical analysis
All assays were performed in ≥3 independent
experiments, and data are presented as the mean ± SD. Comparison
between two groups was analyzed using unpaired Student's t-test.
Comparisons among multiple groups were performed with one-way ANOVA
followed by Tukey's test, using GraphPad Prism 7 (version 7;
GraphPad Software, Inc.). P<0.05 was considered to indicate a
statistically significant difference.
Results
CADM1 has an inhibitory effect in
melanoma
To investigate the role of CADM1 in melanoma cells,
the expression of CADM1 was detected using RT-qPCR. It was found
that the expression of CADM1 was significantly decreased in
melanoma cells (WM-266-4, A2058 and A375) compared with normal
melanocytes cells (PIG1; Fig. 1A).
Western blotting was then used to assess this result, and it was
identified that the protein expression of CADM1 was significantly
inhibited in the melanoma cells compared with PIG1 cells (Fig. 1B). Therefore, these results
indicated that CADM1 had an inhibitory effect in melanoma in
vitro.
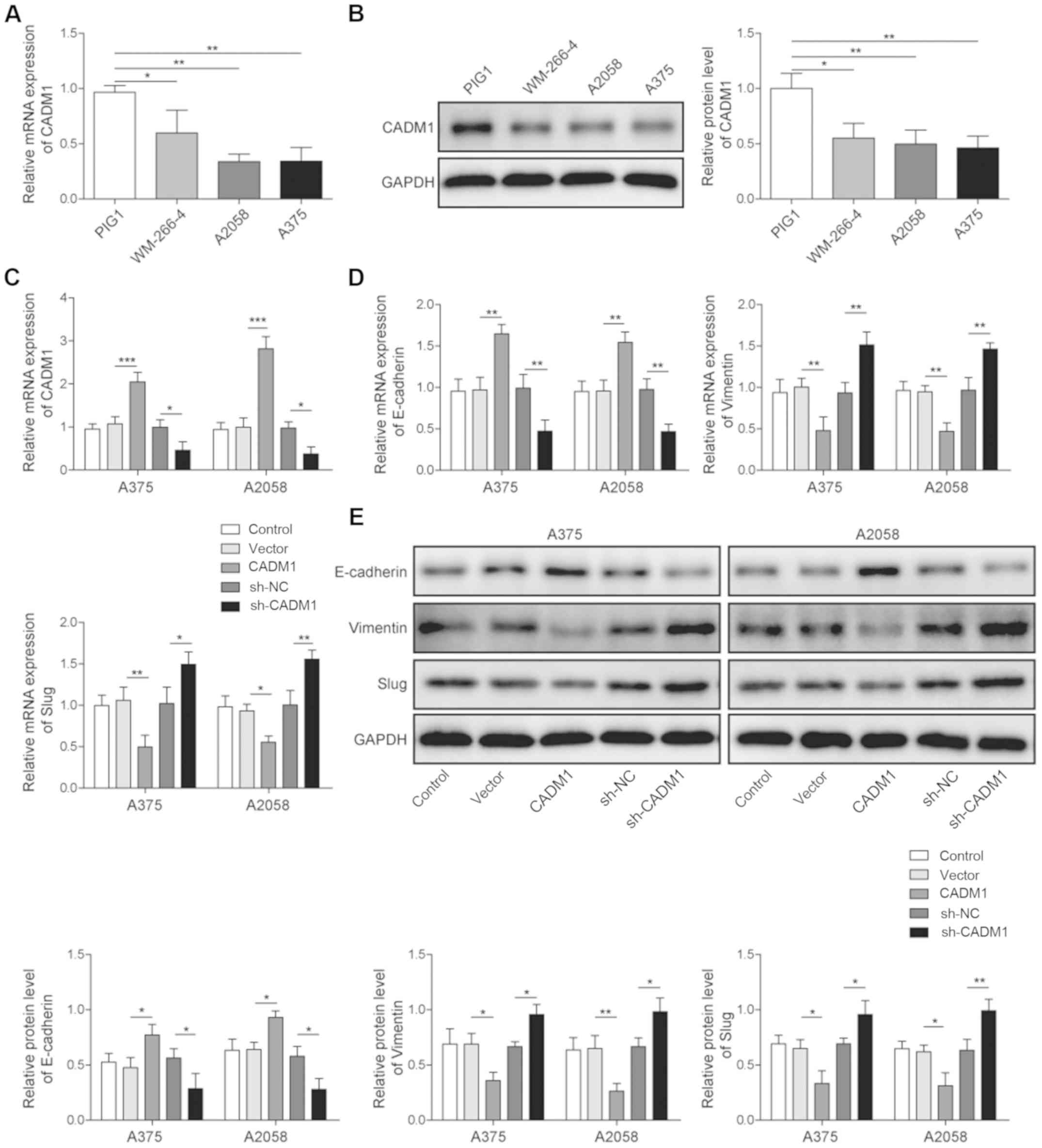 | Figure 1.CADM1 downregulates the EMT process
in melanoma. Relative expression of CADM1 in PIG1, WM-266-4, A2058
and A375 cells was detected by (A) RT-qPCR and (B) western
blotting. (C) A375 or A2058 cells were transfected with
overexpression of CADM1, NC or sh-CADM1 for 24 h. Then, cell
transfection was assessed by RT-qPCR. Relative expression levels of
EMT-related proteins in melanoma cells were detected by (D) RT-qPCR
and (E) western blotting. There were ≥3 independent experiments
performed for each group.*P<0.05, **P<0.01, ***P<0.001.
sh, short hairpin RNA; NC, negative control; miR, microRNA; CADM1,
cell adhesion molecule 1; RT-qPCR, reverse
transcription-quantitative PCR; EMT, epithelial-mesenchymal
transition. |
Next, the efficiency of cell transfection was
detected by RT-qPCR. The results suggested that the expression of
CADM1 in the melanoma cells was significantly upregulated by
overexpression of CADM1, but decreased in the presence of short
hairpin (sh)-CADM1 (Fig. 1C).
Then, to examine the effect of CADM1 on the EMT process of
melanoma, RT-qPCR and western blotting were performed. The results
demonstrated that the relative expression levels of vimentin and
Slug were significantly inhibited by overexpression of CADM1, but
increased by CADM1 knockdown. However, the expression of E-cadherin
in melanoma cells was significantly increased after CADM1
overexpression, while knockdown of CADM1 significantly decreased
the relative expression of E-cadherin (Fig. 1D and E). Collectively, these
results demonstrated that overexpression of CADM1 significantly
downregulated the EMT process in melanoma cells, while reducing the
expression of CADM1 had the opposite effect.
miR-214 significantly upregulates the
progression of melanoma in vitro
RT-qPCR was used to investigate the role of miR-214
in melanoma, and it was identified that the expression of miR-214
was significantly increased in melanoma cells compared with normal
cells (Fig. 2A). In addition,
miR-214 expression was increased to the highest level in A375
cells. Next, RT-qPCR was used to assess the cell transfection and
the results indicated that miR-214 was significantly upregulated in
melanoma cells in the presence of miR-214 mimics. However, the
expression of miR-214 in melanoma cells was significantly decreased
by miR-214 inhibitor (Fig.
2B).
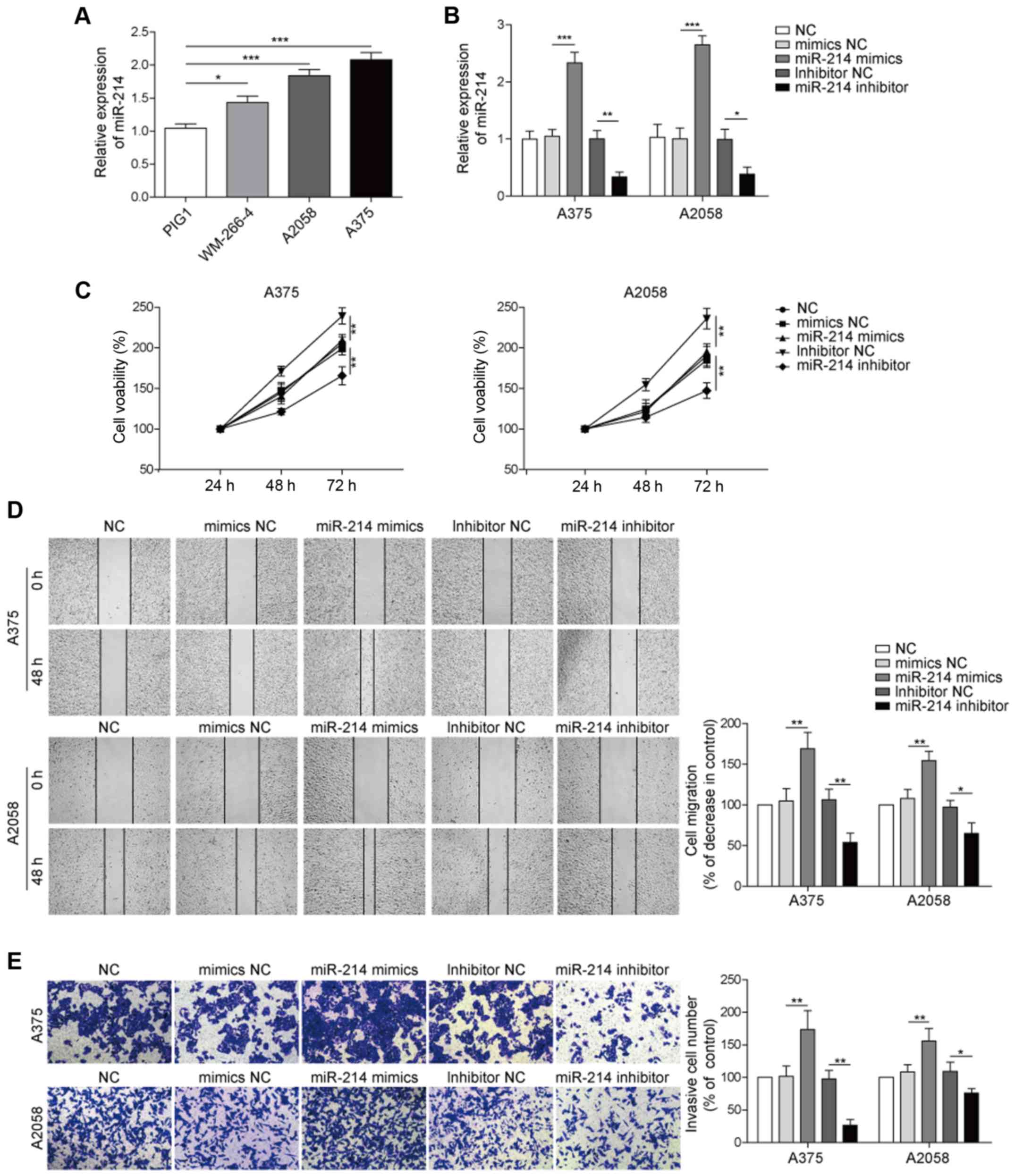 | Figure 2.miR-214 upregulates the tumorigenesis
and progression of melanoma. (A) Relative expression of miR-214 in
PIG1, WM-266-4, A2058 and A375 cells was detected by RT-qPCR. (B)
A375 or A2058 cells were transfected with miR-214 mimics, NC or
miR-214 inhibitor for 24 h. Then, cell transfection was assessed by
RT-qPCR. (C) Cell viability in the NC, mimic NC, inhibitor NC,
miR-214 mimic and miR-214 inhibitor groups was measured after 24,
48 and 72 h using MTT assay. (D) Cell migration in the NC, mimic
NC, inhibitor NC, miR-214 mimic and miR-214 inhibitor groups was
tested by wound healing assay (magnification, ×100). (E) Cell
invasion in the NC, mimic NC, inhibitor NC, miR-214 mimic and
miR-214 inhibitor groups was tested by Transwell assay
(magnification, ×100). NC, negative control. There were ≥3
independent experiments performed for each group. *P<0.05,
**P<0.01, ***P<0.001. NC, negative control; miR, microRNA;
RT-qPCR, reverse transcription-quantitative PCR. |
To further examine the effect of miR-214 on the
tumorigenesis of melanoma, an MTT assay was used to measure the
viability of melanoma cells. The results suggested that the
viability of A375 cells was significantly increased in the presence
of miR-214 mimic, which was reversed by miR-214 inhibitor (Fig. 2C). Moreover, to investigate the
role of miR-214 in the progression of melanoma in vitro,
wound healing and Transwell assays were performed to detect the
migration and invasion of melanoma cells, respectively. It was
found that the miR-214 mimic significantly increased the migration
of melanoma cells, while the miR-214 inhibitor significantly
inhibited the migration of A375 and A2058 cells (Fig. 2D). Furthermore, during the invasion
assay, A375 cells treated with miR-214 mimic exhibited a
significant invasive ability. However, the miR-214 inhibitor
significantly inhibited the invasive ability of melanoma cells
(Fig. 2E). Therefore, the results
demonstrated that miR-214 overexpression significantly promoted the
tumorigenesis and progression of melanoma in vitro, while
downregulation of miR-214 exhibited an inhibitory effect.
miR-214 overexpression significantly
downregulates the expression of CADM1
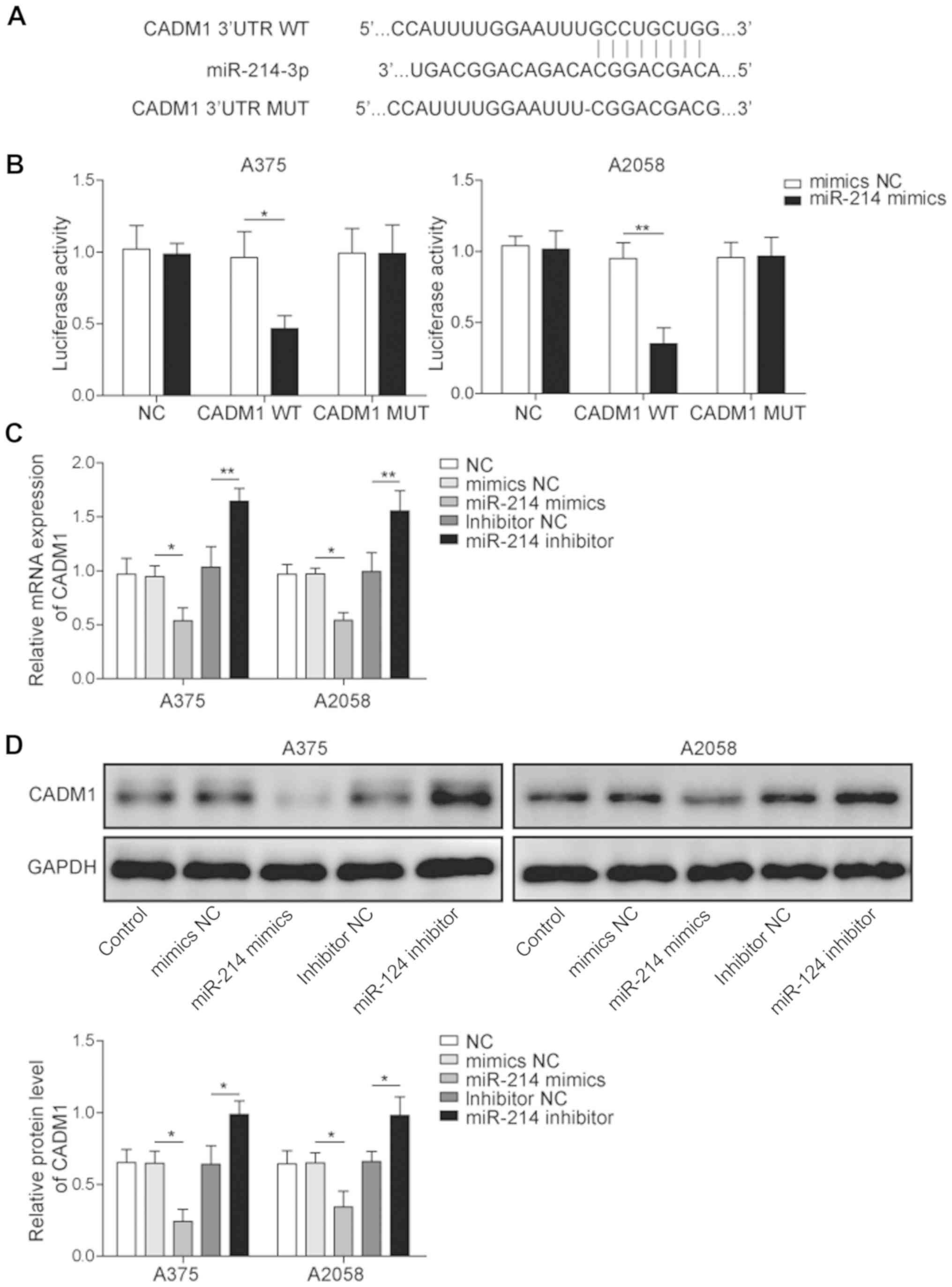
To investigate the relationship between miR-214 and
CADM1, the binding site of CADM1 and miR-214 was analyzed by
bioinformatics prediction software, and a dual-luciferase reporter
assay was performed to verify the specific binding of miR-214 and
CADM1. Luciferase reporter analysis results identified that miR-214
mimic significantly downregulated the activity of the reporter
containing wild-type CADM1; however, mutating the binding site of
miR-214 in CADM1 abolished this suppression (Fig. 3A and B). Thus, the results
suggested that CADM1 may be a direct target regulated by
miR-214.
In addition, RT-qPCR was used to determine the
expression of CADM1 in melanoma cells transfected with miR-214
mimic or inhibitor. It was found that the expression of CADM1 was
significantly enhanced by the miR-214 inhibitor, but was reversed
by miR-214 mimic (Fig. 3C).
Moreover, the results indicated that the protein expression of
CADM1 was significantly inhibited by the presence of miR-214 mimic,
which was reversed by miR-214 inhibitor (Fig. 3D). Therefore, it was suggested that
the overexpression of miR-214 downregulated the expression of CADM1
in melanoma cells, while downregulation of miR-214 increased the
expression of CADM1.
miR-214 promotes EMT in melanoma cells
by downregulating CADM1 expression
To further investigate the mechanism via which
miR-214 regulated the EMT process of melanoma in vitro, the
expression levels of CADM1 and EMT-related proteins were detected
by RT-qPCR and western blotting. It was demonstrated that the
expression of CADM1 was significantly upregulated in the presence
of miR-214 inhibitor (Fig. 4A and
B). Moreover, the miR-214 inhibitor significantly decreased the
relative expression levels of vimentin and Slug, but increased the
relative expression of E-cadherin in melanoma cells, which was
significantly reversed by the knockdown of CADM1.
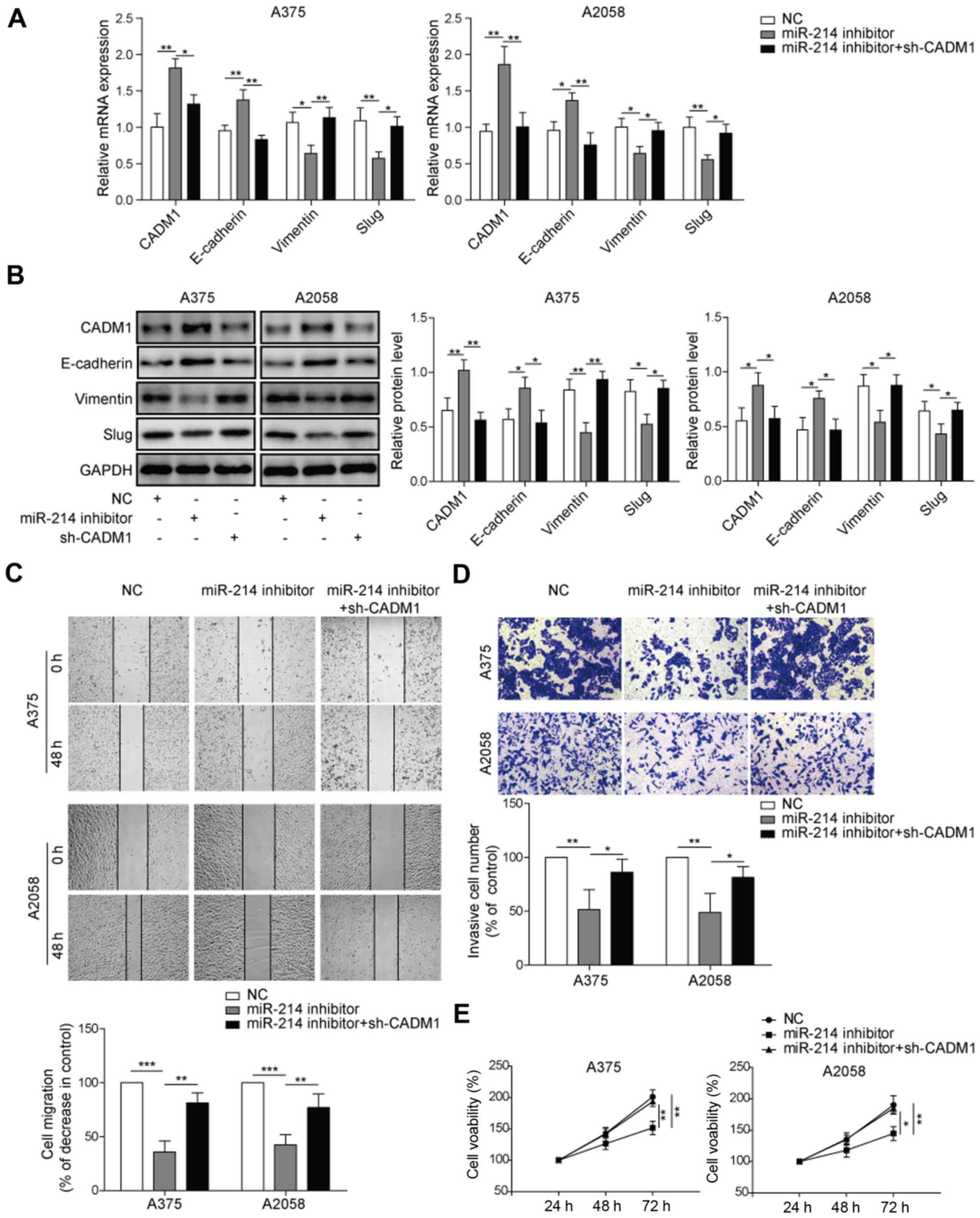 | Figure 4.miR-214 downregulates CADM1 and
upregulates the EMT process in melanoma cells. (A) Relative mRNA
expression levels of E-cadherin, Slug and vimentin in the NC,
miR-214 inhibitor and sh-CADM1 + miR-214 inhibitor groups were
detected by reverse transcription-quantitative PCR. (B) Relative
protein expression levels of EMT-related proteins in the NC,
miR-214 inhibitor and sh-CADM1 + miR-214 inhibitor groups were
detected by western blotting. (C) Cell migration in the NC, miR-214
inhibitor and sh-CADM1 + miR-214 inhibitor groups was assessed by
wound healing assay (magnification, ×100). (D) Cell invasion in the
NC, miR-214 inhibitor and sh-CADM1 + miR-214 inhibitor groups was
measured by Transwell assay (magnification, ×100). (E) Cell
viability in the NC, miR-214 inhibitor and sh-CADM1 + miR-214
inhibitor groups was examined after 24, 48 and 72 h by MTT assay.
There were ≥3 independent experiments performed for each group.
*P<0.05, **P<0.01, ***P<0.001. NC, negative control; miR,
microRNA; WT, wild-type; MUT, mutant; CADM1, cell adhesion molecule
1; sh, short hairpin RNA; EMT, epithelial-mesenchymal
transition. |
Furthermore, to investigate the effect of CADM1 on
melanoma cells transfected with miR-214 inhibitor, wound healing
and Transwell assays were performed. The results demonstrated that
the migration of A375 cells was significantly inhibited by miR-214
inhibitor, which was partially rescued by downregulation of CADM1
(Fig. 4C). In the Transwell assay,
the invasion of A375 and A2058 cells was significantly inhibited in
the presence of miR-214 inhibitor. However, CADM1 downregulation
partially reversed the inhibitory effect of miR-214 inhibitor on
cell invasion in melanoma (Fig.
4D). Moreover, to test the effect of CADM1 on the viability of
A375 or A2058 cells treated with miR-214 inhibitor, an MTT assay
was conducted. It was found that cell viability was decreased by
the miR-214 inhibitor, which was partially reversed by knockdown of
CADM1 (Fig. 4E). Collectively, the
results indicated that miR-214 affected the EMT process of melanoma
by modulating the expression of CADM1.
Discussion
Previous studies have focused on the metastasis of
malignant tumors (27–29); however, the understanding of the
mechanism of melanoma development remains limited (30). It has been reported that the immune
system plays a key role in the treatment of metastasis in advanced
cancer (31–36). Moreover, E-cadherin, vimentin and
Slug are key regulators during the EMT process of malignant tumors
(37,38). Previous studies have also shown
that EMT-related proteins are closely associated with tumor
metastasis (39,40), which is in line with the present
results, thus indicating that EMT is closely related to tumor
metastasis. Furthermore, it is possible that metastatic cells after
EMT are vulnerable to host immunosurveillance (41). Consistent with these findings, the
present results suggested that metastasis in melanoma may be
closely associated with the EMT process, and thus EMT may play a
critical role in the metastasis of melanoma.
It has been reported that CADM1 plays an inhibitory
role in tumorigenesis and progression of cancer. For instance,
overexpression of CADM1 inhibits cell-cycle progression and
invasion of hepatocellular carcinoma (11). Moreover, the present results
indicated that CADM1 acted as a tumor suppressor in melanoma, which
was similar to the results of a previous study (9). Furthermore, overexpression of CADM1
significantly inhibited the EMT process of melanoma. Therefore, it
was speculated that CADM1 may inhibit the tumorigenesis and
progression of melanoma by downregulating the EMT process.
miR-214 functions as a pro-metastatic factor in
malignant tumors (42), and plays
a critical role in the migration and invasion of cancer cells
(43–45). The present study identified a
potential link between miR-214 and CADM1 in the coordination of
metastasis in melanoma. Furthermore, the present results suggested
that CADM1 expression decreased when miR-214 was upregulated in
melanoma cell lines. It was also found that miR-214 significantly
promoted the EMT process of melanoma, while CADM1 inhibited the EMT
process. These results were similar to those of previous studies
(46–48), thus indicating that miR-214
increased the invasion and migration of melanoma by downregulating
CADM1 to promote the EMT process. However, it has also been
revealed that miR-214 promotes cancer progression by activating the
NF-κB signaling pathway (49).
Therefore, future studies should focus on the association between
melanoma and the NF-κB signaling pathway. In addition, the present
study used 0.5% FBS to test the cell migration in wound healing
assay, which may be a limitation. Although 0.5% FBS may enhance the
proliferative ability of cells to a certain extent, the effect of
proliferation on scratches may be negligible due to the low
concentration, however, in order to further eliminate this
interference, follow-up studies will use other methods to detect
cell migration ability.
In conclusion, the present results suggest that
miR-214 promoted the EMT process of melanoma by downregulating
CADM1, which may serve as a potential novel target for the
treatment of melanoma.
Acknowledgements
Not applicable.
Funding
No funding was received.
Availability of data and materials
All data generated or analyzed during this study are
included in this published article.
Authors' contributions
SJW, WWL and TLZ conceived and supervised the study.
SJW and TLZ designed the study. CJW and YLD performed the
experiments and analyzed the data. All authors reviewed the results
and approved the final version of the manuscript.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Pavri SN, Clune J, Ariyan S and Narayan D:
Malignant melanoma: Beyond the basics. Plast Reconstr Surg.
138:330e–340e. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Rastrelli M, Tropea S, Rossi CR and
Alaibac M: Melanoma: Epidemiology, risk factors, pathogenesis,
diagnosis and classification. In Vivo. 28:1005–1011.
2014.PubMed/NCBI
|
|
3
|
Xue YB, Ding MQ, Xue L and Luo JH:
CircAGFG1 sponges miR-203 to promote EMT and metastasis of
non-small-cell lung cancer by upregulating ZNF281 expression.
Thorac Cancer. 10:1692–1701. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Zhao C, Ling X, Li X, Hou X and Zhao D:
MicroRNA-138-5p inhibits cell migration, invasion and EMT in breast
cancer by directly targeting RHBDD1. Breast Cancer. 26:817–825.
2019. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Ribatti D, Tamma R and Annese T:
Epithelial-mesenchymal transition in cancer: A historical overview.
Transl Oncol. 13:1007732020. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Wang F, Xia H and Yao S: Regulatory T
cells are a double-edged sword in pulmonary fibrosis. Int
Immunopharmacol. 84:1064432020. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Mansoori B, Mohammadi A, Naghizadeh S,
Gjerstorff M, Shanehbandi D, Shirjang S, Najafi S, Holmskov U,
Khaze V, Duijf PHG and Baradaran B: miR-330 suppresses EMT and
induces apoptosis by downregulating HMGA2 in human colorectal
cancer. J Cell Physiol. 235:920–931. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Yuki A, Shinkuma S, Hayashi R, Fujikawa H,
Kato T, Homma E, Hamade Y, Onodera O, Matsuoka M, Shimizu H, et al:
CADM1 is a diagnostic marker in early-stage mycosis fungoides:
Multicenter study of 58 cases. J Am Acad Dermatol. 79:1039–1046.
2018. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Hartsough EJ, Weiss MB, Heilman SA, Purwin
TJ, Kugel CH III, Rosenbaum SR, Erkes DA, Tiago M, HooKim K,
Chervoneva I and Aplin AE: CADM1 is a TWIST1-regulated suppressor
of invasion and survival. Cell Death Dis. 10:2812019. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Rong G, Zhang M, Xia W, Li D, Miao J and
Wang H: Plasma CADM1 promoter hypermethylation and D-dimer as novel
metastasis predictors of cervical cancer. J Obstet Gynaecol Res.
45:1251–1259. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Wang F, Qi X, Li Z, Jin S, Xie Y and Zhong
H: lncRNA CADM1-AS1 inhibits cell-cycle progression and invasion
via PTEN/AKT/GSK-3β axis in hepatocellular carcinoma. Cancer Manag
Res. 11:3813–3828. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
You Y, Zhang J, Li Y, Li Y, Shi G, Ma L
and Wei H: CADM1/TSLC1 inhibits melanoma cell line A375 invasion
through the suppression of matrix metalloproteinases. Mol Med Rep.
10:2621–2626. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Mohr AM and Mott JL: Overview of microRNA
biology. Semin Liver Dis. 35:3–11. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Guo J, Jin H, Xi Y, Guo J, Jin Y and Jiang
D: The miR-582/CD1B axis is involved in regulation of dendritic
cells and is associated with clinical outcomes in advanced lung
adenocarcinoma. Biomed Res Int. 2020:43609302020. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Nayak B, Khan N, Garg H, Rustagi Y, Singh
P, Seth A, Dinda AK and Kaushal S: Role of miRNA-182 and miRNA-187
as potential biomarkers in prostate cancer and its correlation with
the staging of prostate cancer. Int Braz J Urol. 46:614–623. 2020.
View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Sun X, Xu W, Zang C and Li N:
miRNA-520c-3p accelerates progression of nasopharyngeal carcinoma
via targeting RAB22A. Oncol Lett. 19:771–776. 2020.PubMed/NCBI
|
|
17
|
Tiwari A, Mukherjee B and Dixit M:
MicroRNA key to angiogenesis regulation: MiRNA biology and therapy.
Curr Cancer Drug Targets. 18:266–277. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Lu TX and Rothenberg ME: MicroRNA. J
Allergy Clin Immunol. 141:1202–1207. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Chen S, Yang C, Sun C, Sun Y, Yang Z,
Cheng S and Zhuge B: miR-21-5p suppressed the sensitivity of
hepatocellular carcinoma cells to cisplatin by targeting FASLG. DNA
Cell Biol. 38:865–873. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Guo HL, Chen LD, Li W, Liang JY, Zhang JC,
Li X, Xie XY, Lu MD, Kuang M and Wang W: Ultrasomics for early
evaluation of the tumor response to microRNA-122 in a nude mouse
hepatocellular carcinoma model. J Ultrasound Med. 39:61–71. 2019.
View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Lin T, Hou PF, Meng S, Chen F, Jiang T, Li
ML, Shi ML, Liu JJ, Zheng JN and Bai J: Emerging roles of p53
related lncRNAs in cancer progression: A systematic review. Int J
Biol Sci. 15:1287–1298. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Chen Z, Zhuang W, Wang Z, Xiao W, Don W,
Li X and Chen X: MicroRNA-450b-3p inhibits cell growth by targeting
phosphoglycerate kinase 1 in hepatocellular carcinoma. J Cell
Biochem. 120:18805–18815. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Chen J, Lin Y, Jia Y, Xu T, Wu F and Jin
Y: LncRNA HAND2-AS1 exerts anti-oncogenic effects on ovarian cancer
via restoration of BCL2L11 as a sponge of microRNA-340-5p. J Cell
Physiol. 234:23421–23436. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Yin Y, Cai X, Chen X, Liang H, Zhang Y, Li
J, Wang Z, Chen X, Zhang W, Yokoyama S, et al: Tumor-secreted
miR-214 induces regulatory T cells: A major link between immune
evasion and tumor growth. Cell Res. 24:1164–1180. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Mirzaei H, Gholamin S, Shahidsales S,
Sahebkar A, Jaafari MR, Mirzaei HR, Hassanian SM and Avan A:
MicroRNAs as potential diagnostic and prognostic biomarkers in
melanoma. Eur J Cancer. 53:25–32. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Uketa S, Shimizu Y, Ogawa K, Utsunomiya N,
Katsushima H, Ishihara M, Hashimoto K and Kanamaru S: A case of
isolated peritoneal metastasis from prostate cancer. Hinyokika
Kiyo. 65:175–179. 2019.(In Japanese). PubMed/NCBI
|
|
28
|
Shi J, Lu P, Shen W, He R, Yang MW, Fang
Y, Sun YW, Niu N and Xue J: CD90 highly expressed population
harbors a stemness signature and creates an immunosuppressive niche
in pancreatic cancer. Cancer Lett. 453:158–169. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Nakano K, Masui T, Yogo A, Uchida Y, Sato
A, Kasai Y, Nagai K, Anazawa T, Kawaguchi Y and Uemoto S:
Chloroquine induces apoptosis in PanNEN via endoplasmic reticulum
stress. Endocr Relat Cancer. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Shen C, Hua H, Gu L, Cao S, Cai H, Yao X
and Chen X: miR-124 functions as a melanoma tumor suppressor by
targeting RACK1. Onco Targets Ther. 12:9975–9986. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Chockley PJ, Chen J, Chen G, Beer DG,
Standiford TJ and Keshamouni VG: Epithelial-mesenchymal transition
leads to NK cell-mediated metastasis-specific immunosurveillance in
lung cancer. J Clin Invest. 128:1384–1396. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Slaney CY, Rautela J and Parker BS: The
emerging role of immunosurveillance in dictating metastatic spread
in breast cancer. Cancer Res. 73:5852–5857. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Hong H, Gu Y, Zhang H, Simon AK, Chen X,
Wu C, Xu XN and Jiang S: Depletion of CD4+CD25+ regulatory T cells
enhances natural killer T cell-mediated anti-tumour immunity in a
murine mammary breast cancer model. Clin Exp Immunol. 159:93–99.
2010. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Olkhanud PB, Baatar D, Bodogai M, Hakim F,
Gress R, Anderson RL, Deng J, Xu M, Briest S and Biragyn A: Breast
cancer lung metastasis requires expression of chemokine receptor
CCR4 and regulatory T cells. Cancer Res. 69:5996–6004. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Paolino M, Choidas A, Wallner S, Pranjic
B, Uribesalgo I, Loeser S, Jamieson AM, Langdon WY, Ikeda F, Fededa
JP, et al: The E3 ligase Cbl-b and TAM receptors regulate cancer
metastasis via natural killer cells. Nature. 507:508–512. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Yang L, Huang J, Ren X, Gorska AE, Chytil
A, Aakre M, Carbone DP, Matrisian LM, Richmond A, Lin PC and Moses
HL: Abrogation of TGF beta signaling in mammary carcinomas recruits
Gr-1+CD11b+ myeloid cells that promote metastasis. Cancer Cell.
13:23–35. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Wei CY, Zhu MX, Yang YW, Zhang PF, Yang X,
Peng R, Gao C, Lu JC, Wang L, Deng XY, et al: Downregulation of
RNF128 activates Wnt/β-catenin signaling to induce cellular EMT and
stemness via CD44 and CTTN ubiquitination in melanoma. J Hematol
Oncol. 12:212019. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Pearlman RL, Montes de Oca MK, Pal HC and
Afaq F: Potential therapeutic targets of epithelial-mesenchymal
transition in melanoma. Cancer Lett. 391:125–140. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Wang C, Li K, Men Y, Ding C, Du J, Liang
T, Ji Z, Chen L, Wang T and Kang Q: Protein 4.1B suppresses tumor
metastasis by regulating epithelial-mesenchymal transition
progression in melanoma cells. Int J Med Sci. 16:529–536. 2019.
View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Wels C, Joshi S, Koefinger P, Bergler H
and Schaider H: Transcriptional activation of ZEB1 by Slug leads to
cooperative regulation of the epithelial-mesenchymal
transition-like phenotype in melanoma. J Invest Dermatol.
131:1877–1885. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Bie Y, Ge W, Yang Z, Cheng X, Zhao Z, Li
S, Wang W, Wang Y, Zhao X, Yin Z and Li Y: The crucial role of
CXCL8 and its receptors in colorectal liver metastasis. Dis
Markers. 2019:80234602019. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Penna E, Orso F, Cimino D, Vercellino I,
Grassi E, Quaglino E, Turco E and Taverna D: miR-214 coordinates
melanoma progression by upregulating ALCAM through TFAP2 and
miR-148b downmodulation. Cancer Res. 73:4098–4111. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Maity S, Das F, Ghosh-Choudhury N,
Kasinath BS and Ghosh Choudhury G: High glucose increases miR-214
to power a feedback loop involving PTEN and the Akt/mTORC1
signaling axis. FEBS Lett. 593:2261–2272. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Gong L, Xu H, Zhang X, Zhang T, Shi J and
Chang H: Oridonin relieves hypoxia-evoked apoptosis and autophagy
via modulating microRNA-214 in H9c2 cells. Artif Cells Nanomed
Biotechnol. 47:2585–2592. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Eguchi S, Takefuji M, Sakaguchi T,
Ishihama S, Mori Y, Tsuda T, Takikawa T, Yoshida T, Ohashi K,
Shimizu Y, et al: Cardiomyocytes capture stem cell-derived,
anti-apoptotic microRNA-214 via clathrin-mediated endocytosis in
acute myocardial infarction. J Biol Chem. 294:11665–11674. 2019.
View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Sun R, Liu Z, Han L, Yang Y, Wu F, Jiang
Q, Zhang H, Ma R, Miao J, He K, et al: miR-22 and miR-214 targeting
BCL9L inhibit proliferation, metastasis, and epithelial-mesenchymal
transition by down-regulating Wnt signaling in colon cancer. FASEB
J. 33:5411–5424. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Okazaki Y, Chew SH, Nagai H, Yamashita Y,
Ohara H, Jiang L, Akatsuka S, Takahashi T and Toyokuni S:
Overexpression of miR-199/214 is a distinctive feature of iron- and
asbestos-induced sarcomatoid mesothelioma in rats. Cancer Sci.
2020.(Epub ahead of print). View Article : Google Scholar
|
|
48
|
Kong L, Liu P, Zheng M, Wang Z, Gao Y,
Liang K, Wang H and Tan X: The miR-1224-5p/ELF3 axis regulates
malignant behaviors of pancreatic cancer via PI3K/AKT/Notch
signaling pathways. Onco Targets Ther. 13:3449–3466. 2020.
View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Duan Y, Tan Z, Yang M, Li J, Liu C, Wang
C, Zhang F, Jin Y, Wang Y and Zhu L: PC-3-derived exosomes inhibit
osteoclast differentiation by downregulating miR-214 and blocking
NF-κB signaling pathway. Biomed Res Int. 2019:86508462019.
View Article : Google Scholar : PubMed/NCBI
|