Introduction
Neural stem cells (NSCs) are able to self-renew and
differentiate into neurons, oligodendrocytes and astrocytes under
appropriate inducing conditions (1); therefore, NSCs are the most commonly
used stem cells in repairing neuronal injuries (2). In neurogenesis, NSCs exist in the
developing brain and serve an important role in brain plasticity
throughout life (3,4). Transplanting NSCs into the host brain
is an efficient strategy to repair neuronal injury and treat
neurodegenerative diseases (5).
Mesenchymal stem cells (MSCs) have the potential of
multidirectional differentiation and neuroprotective effects
(6). Notably, it may be the
secretory products rather than the stem cells themselves, which
serve a dominant role in neuroprotection (7–10).
Extracellular vesicles (EVs) of MSCs have advantages, including
being more efficient when used in the treatment of diseases
compared with MSCs (11–13). EVs carry a variety of proteins,
lipids, RNAs, microRNAs (miRs) and other molecules, and have a high
ability to enter into target cells (14,15).
Among the cargo of EVs, miRs are critical posttranscriptional
factors that regulate biological and physiological functions
(16). A large number of different
miRs are expressed sequentially and participate in the process of
nerve repair or promote neuron regeneration following peripheral
nerve injury (17). miR-210-3p is
a key regulator that determines neuron survival under hypoxic
conditions (18). miR-210-3p
inhibits apoptosis and promotes cell survival under oxygen-glucose
deprivation and oxidative stress conditions, and repairs neurons
following spinal cord injury (19). In addition, miR-210-3p is present
in EVs, which indicates a possible physiological function for
miR-210-3p (20).
Cobalt chloride is widely used to produce in
vitro hypoxic injury models in NSCs (21,22)
as it triggers NSC apoptosis via the nuclear factor erythroid
2-related factor 2 (Nrf2)/antioxidant response elements (ARE)
signaling pathway (21). The
present study aimed to examine the potential function of miR-210-3p
in the protective effects of MSC-EVs against NSC hypoxic injury
regarding NSC apoptosis.
Materials and methods
Animals
A total of 5 pregnant Sprague-Dawley (SD) rats (350
g, aged 3 months) were purchased from Hunan Slake Jingda Laboratory
Animal Co., Ltd. (license no. SCXK (Hunan) 2016-0002). The animals
were housed in a specific pathogen-free environment at 23±2°C with
45–65% humidity, 12-h light/dark cycles and free access to food and
water. All experimental protocols were approved by the Ethics
Committee of Kunming Medical University.
Preparation of EVs from MSCs
MSCs were purchased from BeNa Culture Collection
(cat no. BNCC 340370; Suzhou Bena Chuanglian Biotechnology Co.
Ltd.). After 48-h starvation using fetal bovine serum (FBS)-free
Cellartis MSC Xeno-Free Culture Medium (cat. no. Y50200, Cellartis;
Takara Bio Europe AB), EVs were prepared using an Exosome
Extraction kit (cat no. E1310, Beijing Weihui Biotechnology Co.,
Ltd.) following the manufacturer's protocol. Cells and cell debris
were removed by centrifugation at 2,000 × g at 4°C for 10 min.
Subsequently, the EVs were fixed in 2.5% glutaraldehyde for 2 h at
room temperature. Following embedding in epoxy-based resins at room
temperature for 3 h, the samples were sectioned into 70-nm slices
and stained with 3% uranium acetate and 3% lead citrate at room
temperature for 5 min. The slides were observed via transmission
electron microscopy (JEM-1230; JEOL, Ltd.) at 80KV. The particle
sizes of exosomes were measured using a Laser Particle Size
Analyzer (LA-960; HORIBA Scientific).
Preparation of NSCs
A total of 5 pregnant rats (age, 15 days; weight,
300 g) were anesthetized with isoflurane (5%) and then decapitated.
The fetal rats were sterilized with 75% ethanol for 30 min at room
temperature. The hippocampus tissue was separated and immersed in
Hanks' Balanced Salt solution (Thermo Fisher Scientific, Inc.).
After being fully shredded, the hippocampus tissue was digested
with 0.125% trypsin for 10 min and the reaction was terminated
using Dulbecco's Minimum Essential Medium (Gibco; Thermo Fisher
Scientific, Inc.). The cells were collected and suspended in fresh
medium containing 10% FBS (Hyclone; Cytivia) and cultured in a 5%
CO2 incubator at 37°C.
Immunohistochemistry
The collected cells were fixed in 4%
paraformaldehyde for 30 min at room temperature. Endogenous
peroxidase activity was blocked with 3% (v/v)
H2O2 for 5 min at room temperature. The
slides were blocked in 5% bovine serum albumin (Hyclone, Thermo
Fisher Scientific, Inc.) at room temperature for 2 h and incubated
with a monoclonal anti-nestin primary antibody (1:200; cat no.
OM264981; Omnimabs) overnight at 4°C, followed by incubation with a
horseradish peroxidase (HRP)-labeled goat anti-rabbit IgG secondary
antibody (1:10,000; cat. no. A16104SAMPLE; Thermo Fisher
Scientific, Inc.) for 30 min at room temperature.
Immunohistochemical staining was visualized using
3,3′-diaminobenzidine chromogen for 3 min at room temperature.
Images were captured using a routine light microscope
(magnification: 200×; BX51; Olympus Corporation) and at ≥5 fields
in each slide were selected at random.
Experimental groups
Following culture for 24 h, the hypoxic injury model
was established using 200 µM cobalt chloride (purity: 97%;
Sigma-Aldrich; Merck KGaA), as previously described (21). The NSCs were divided into three
groups: i) Normal NSCs group (Control); ii) NSCs hypoxic injury
group (Model); and iii) NSCs hypoxic injury + MSCs-EVs group.
Following treatment with cobalt chloride for 24 h at 37°C, MSCs-EVs
(20 µg/ml) were added to the cells (3×105/ml) for
a further 24 h at 37°C. To evaluate the effect of miR-210-3p, an
miR-210-3p inhibitor was designed and transfected into NSCs. The
NSCs were divided into four groups: i) Control group; ii) hypoxic
injury group (Model); iii) hypoxic injury + miR-210-3p NC group
(miR-210-3p NC); and iv) hypoxic injury + miR-210-3p inhibitor
group (miR-210-3p inhibitor).
Cell transfection
At 70% confluence, NSCs were transfected. The
transfection solution consisted of 125 µl Opti-MEM (Gibco, Thermo
Fisher Scientific, Inc.), 5 µl Lipofectamine® 3000
(Thermo Fisher Scientific, Inc.) and 12.5 µl RNA inhibitor (RNA
inhibitor powder dissolved in diethyl pyrocarbonate; 40 µg/125 µl).
The transfection solution was added to the corresponding wells in
the 6-well plate for 4 h and the final concentration of miR-210-3p
inhibitor was 1 µg/ml. Subsequently, complete medium containing 20%
FBS was added to the 6-well plate at 37°C. Then, 48 h later at
37°C, the transfections were verified by reverse
transcription-quantitative PCR (RT-qPCR). miR-210-3p inhibitor was
synthesized by Anhui General Bioengineering Co., Ltd. The sequences
of miR-210-3p inhibitor and NC were UCAGCCGCUGUCACACGCACAG (5′-3′)
and UCUACUCUUUCUAGGAGGUUGUGA (5′-3′), respectively.
TUNEL
NSC apoptosis was detected by performing a TUNEL
assay (cat no. C1082; Beyotime Institute of Biotechnology)
following the manufacturer's protocol. Briefly, terminal
deoxynucleotidyl transferase (2 µl) and fluorescent reagent (DAPI,
48 µl) were mixed and applied to the cells at 37°C for 60 min.
Following washing by 0.1 M phosphate buffer saline, the slides were
covered with anti-fading reagent and observed under a fluorescence
microscope (magnification: 200×; BX51; Olympus Corporation) and ≥5
fields in each slide were selected randomly.
Reverse transcription-quantitative
PCR
Following treatment, total RNA was extracted from
cells using an Ultrapure RNA extraction kit (CoWin Biosciences).
The purity of RNA was assessed by measuring the optical density of
each sample at a wavelength of 280/260 nm by a spectrophotometer
(ShanghaiPuyuan Instrument Co., Ltd.). RNA (1 µg) was reverse
transcribed into cDNA using an Avian Myeloblastosis Virus
Reverse-Transcriptase kit (cat. no. CW2141S; CoWin Biosciences).
The qPCR reaction system included 7 µl RNase-Free dH2O, 1 µl
cDNA/DNA, 2 µl primers and 10 µl 2XUltraSYBR Mixture (cat. no.
00081405; CoWin Biosciences). qPCR was performed using the
following thermocycling conditions: Predenaturation for 10 min; 40
cycles of denaturation at 95°C for 10 sec, annealing at 58°C for 30
sec and extension at 72°C for 30 sec. The following primers were
used for qPCR: U6 forward, 5′-GCTTCGGCAGCACATATACTAAAAT-3′ and
reverse, 5′-CGCTTCACGAATTTGCGTGTCAT-3′; and forward,
rno-miR-210-3p, 5′-CTGTGCGTGTGACAGCGGCTGA-3′; reverse,
5′-GCACTGCCTGCGGGACATACC−3′. The target gene expression was
normalized to U6 using the 2−ΔΔCq method (23).
Western blotting
Following treatment, total protein was extracted
from cells using the TriplePrep kit (cat. no. 28-9425-44;
ReadyPrep; Cytiva). Total protein was quantified using a
bicinchoninic acid protein assay kit (Beyotime Institute of
Biotechnology). A total of 25 µg/lane protein was separated via
SDS-PAGE (10% gel) and transferred onto nitrocellulose membranes,
as previously described (24). The
membranes were blocked in 5% skimmed milk at room temperature for 2
h and incubated with the following primary antibodies overnight at
4°C: Rabbit polyclonal anti-Bcl-2 19 kDa interacting protein
(BNIP3; 1:1,000; cat. no. ab109362; Abcam), mouse polyclonal
anti-apoptosis-inducing factor (AIF; 1:1,000; cat. no. BF0591;
Affinity Biosciences) and mouse monoclonal anti-β-actin (1:2,000;
cat. no. TA890010; OriGene Technologies, Inc.). The membranes were
washed three times with 1X phosphate-buffered saline with 0.05%
Tween (PBST) and incubated with secondary antibodies (HRP-labeled
goat anti-rabbit IgG; cat. no. 65-6120; Thermo Fisher Scientific,
Inc.; HRP-labeled goat anti-mouse IgG; cat. no. 31430; Thermo
Fisher Scientific, Inc.) at 4°C for 2 h. Protein bands were
visualized using an electrochemiluminescence kit (Thermo Fisher
Scientific, Inc.). Protein expression was quantified using the
Quantity One software (v4.62; Bio-Rad Laboratories, Inc.) with
β-actin as the loading control.
Statistical analysis
Data are presented as the mean ± standard deviation
with five experimental repeats in each group. One-way ANOVA
followed by the Newman-Keuls post hoc test was used to analyze
comparisons between multiple groups. Statistical analyses were
performed using SPSS software (version 19.0; IBM Corp.). P<0.05
was considered to indicate a statistically significant
difference.
Results
Identification of EVs and hippocampal
cells
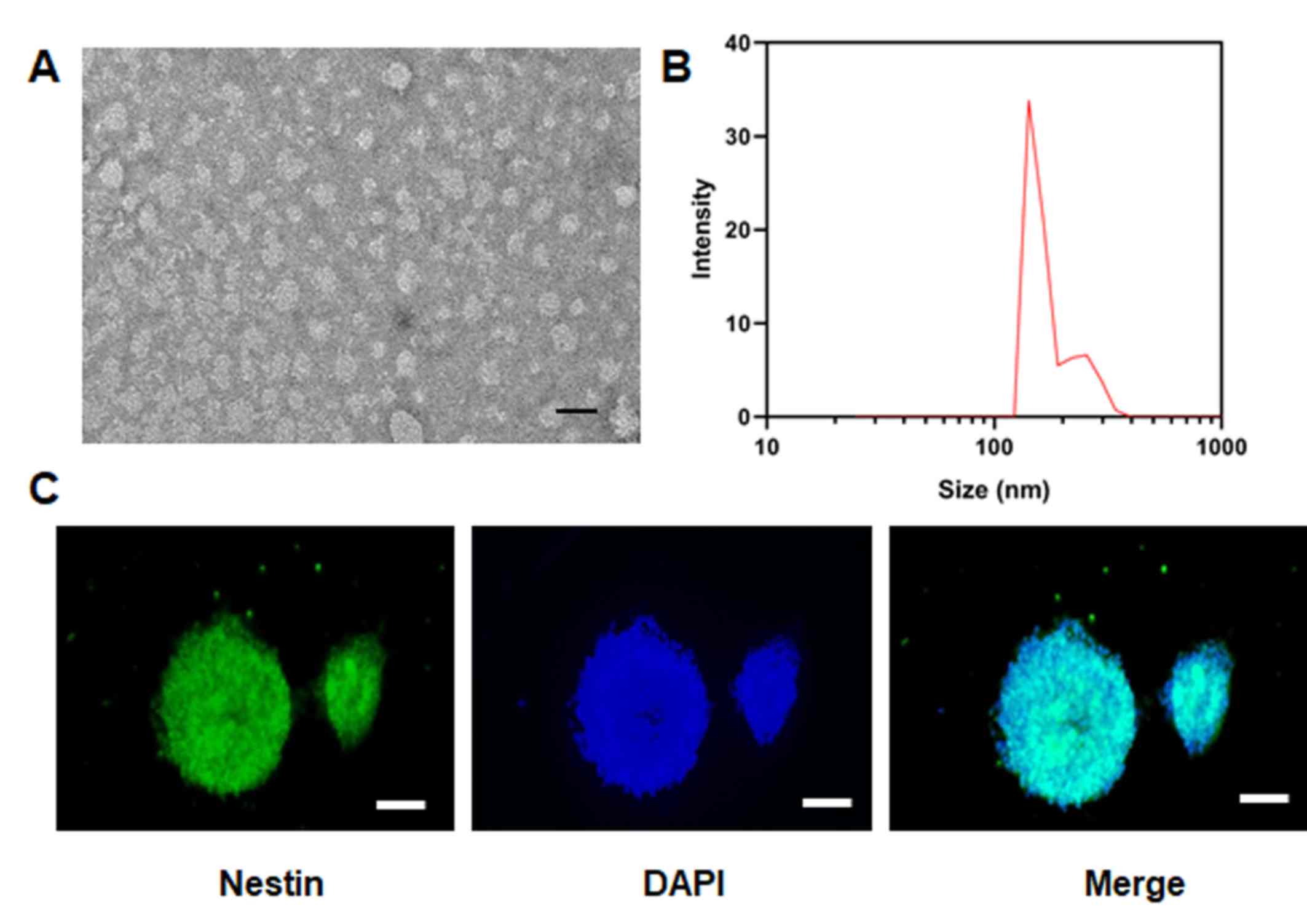
EVs were imaged via transmission electron microscopy
(Fig. 1A) and the results
indicated that the particle size of EVs was ~150-200 nm (Fig. 1B). The positive rate of nestin, a
specific marker of NSCs, in the isolated cells was >98% based on
immunohistochemical staining (Fig.
1C), indicating the successful isolation of NSCs.
MSC-EVs reduce hypoxic injury-induced
NSC apoptosis, and AIF and BNIP3 expression levels, but promote
miR-210-3p expression
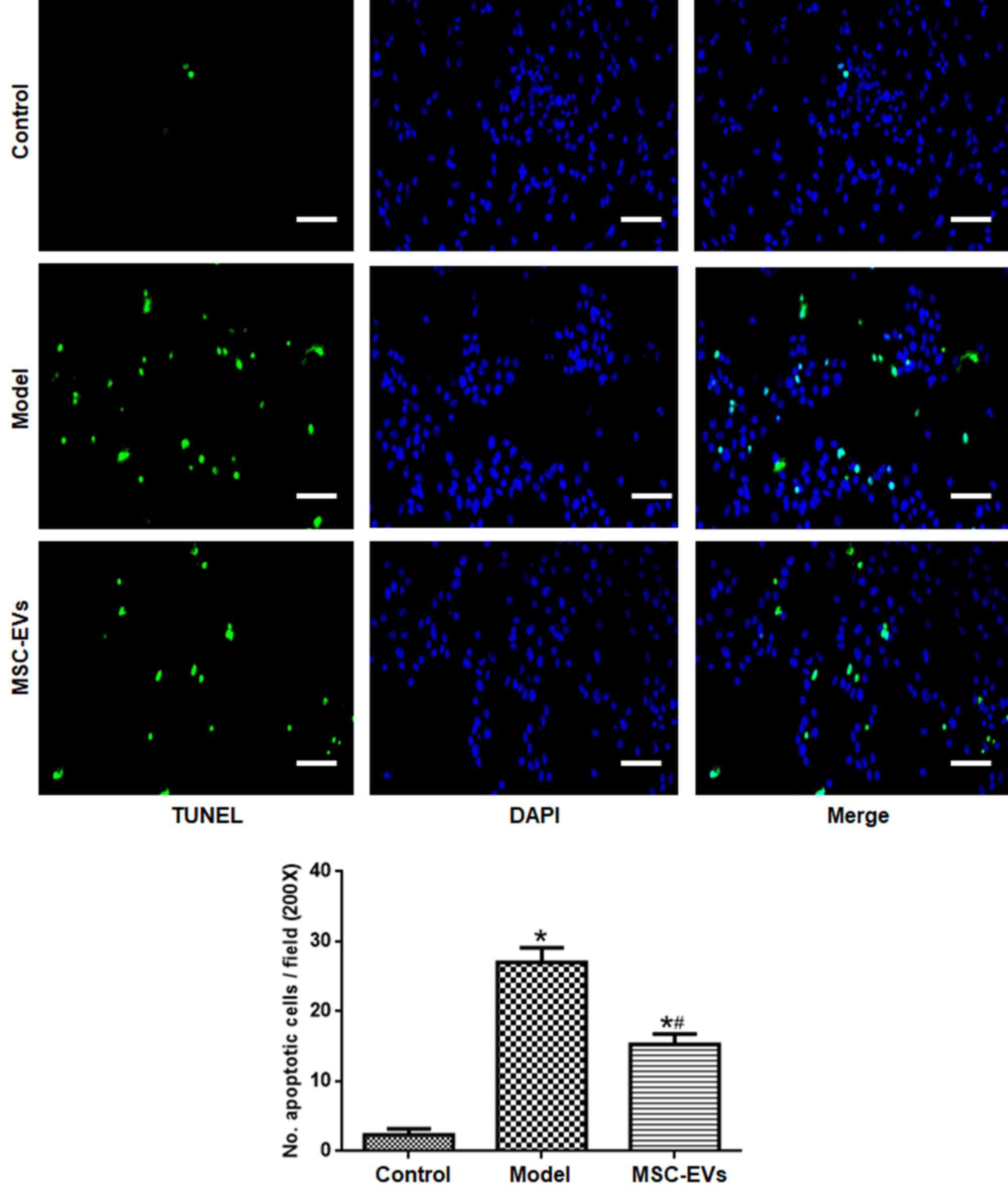
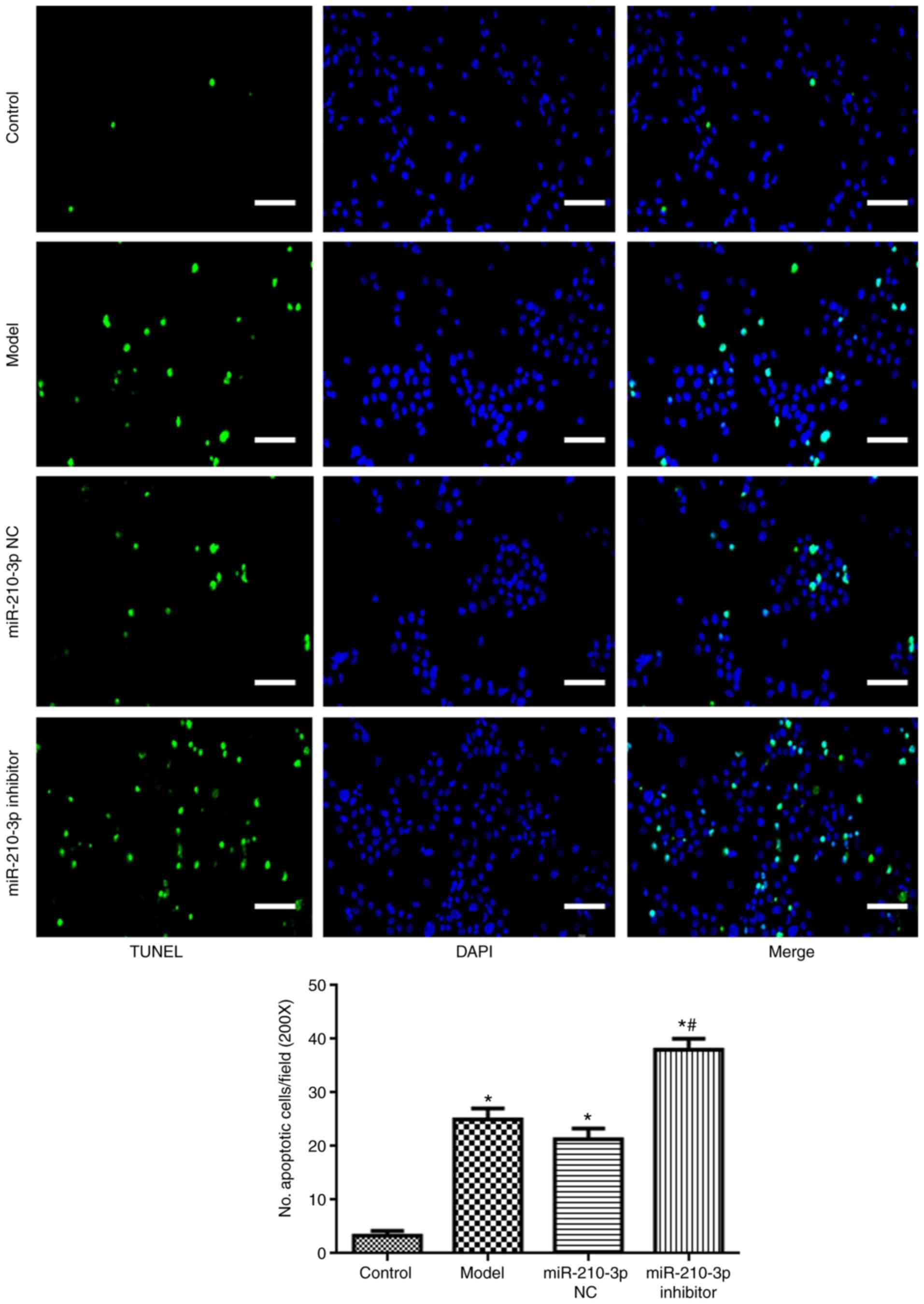
Compared with the control group, NSC apoptosis in
the model group was significantly higher (Fig. 2). By contrast, NSC apoptosis was
significantly reduced in the MSC-EVs group compared with the model
group (P<0.05).
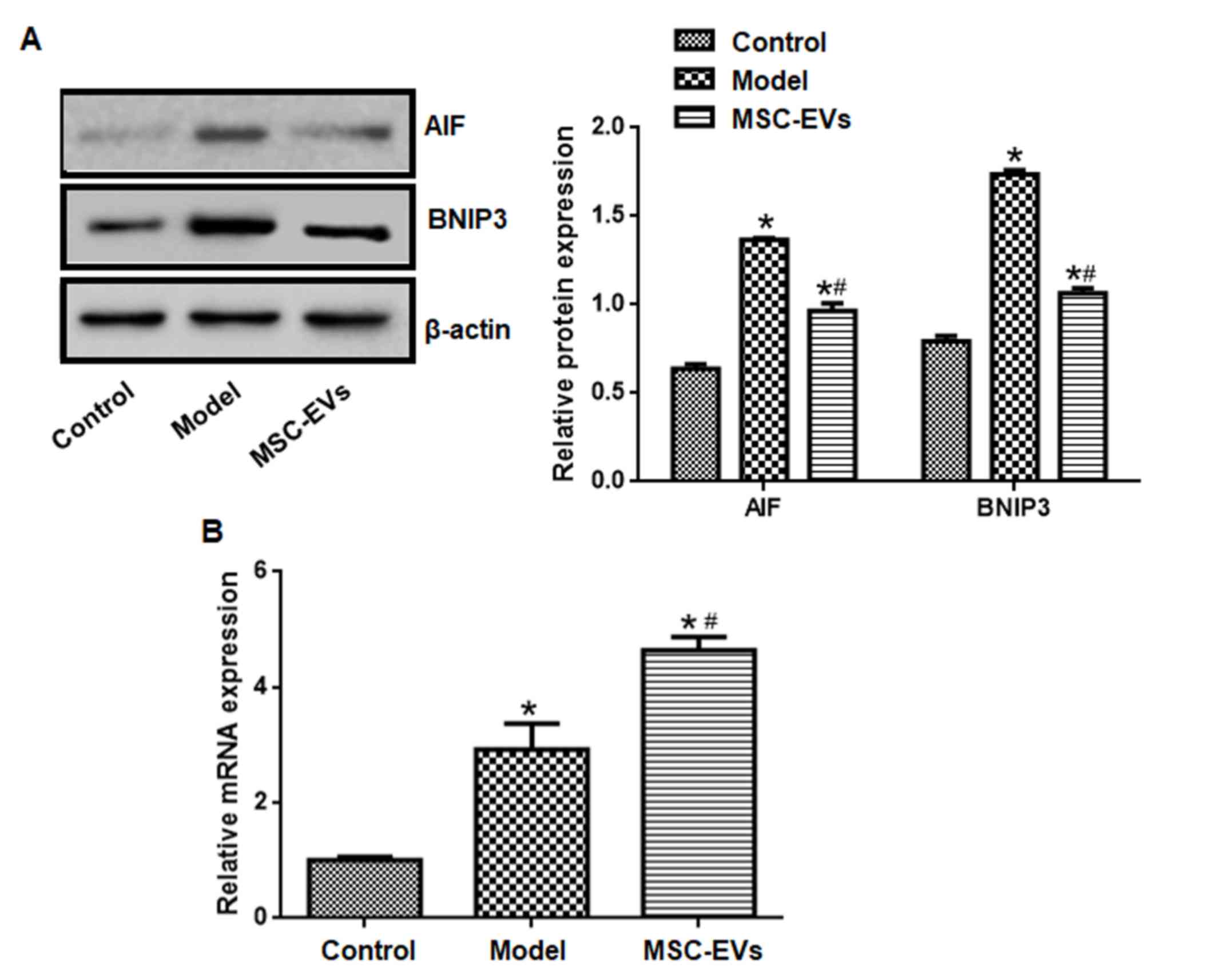
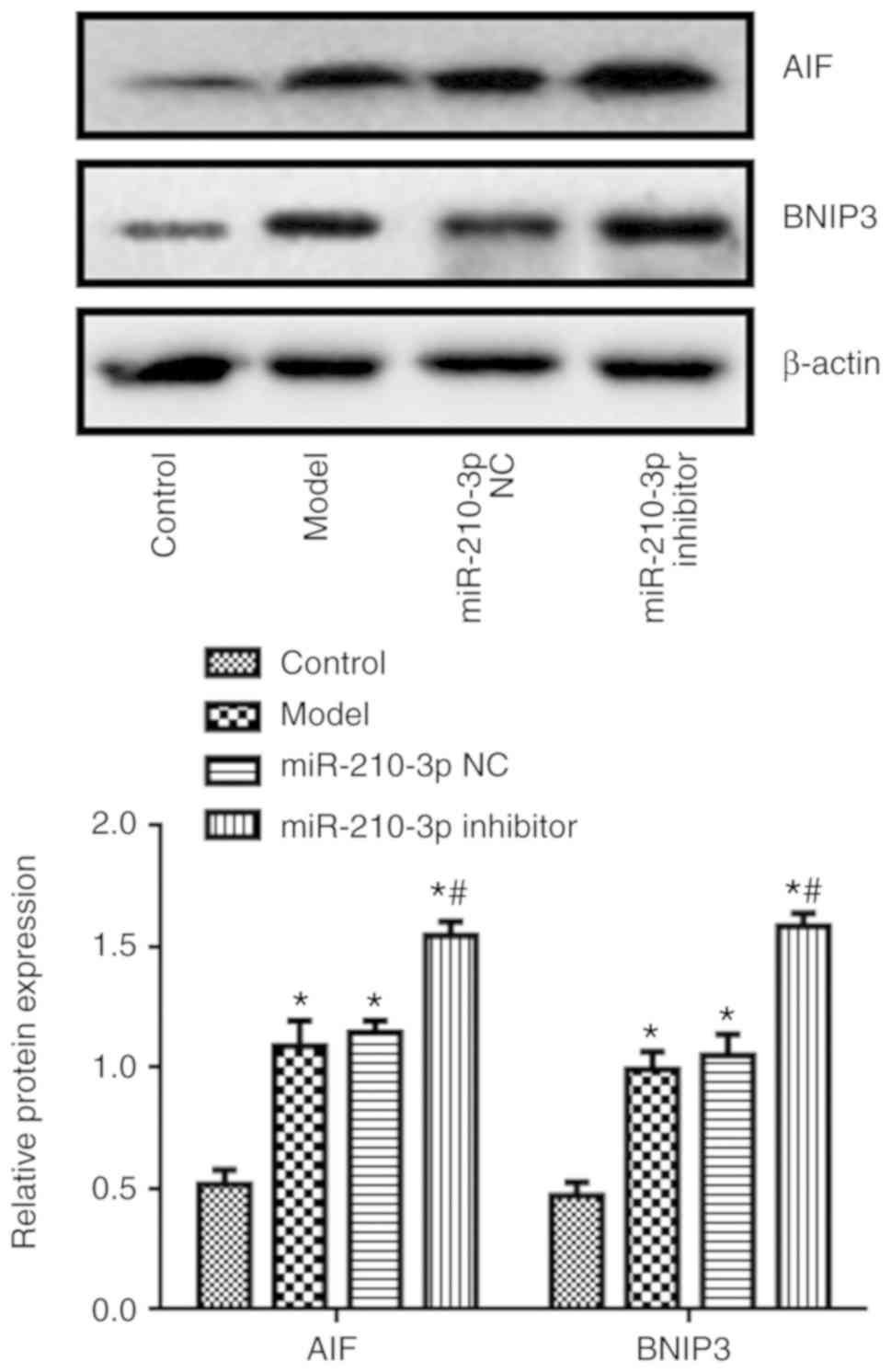
The expression levels of AIF and BNIP3 in the model
group were significantly higher compared with the control group,
whereas the expression levels of AIF and BNIP3 in the MSC-EVs group
were significantly lower compared with the model group (P<0.05;
Fig. 3A).
Compared with the control group, miR-210-3p
expression levels in the model group were significantly higher. In
addition, treatment with MSC-EVs further increased miR-210-3p
expression compared with the model group (P<0.05; Fig. 3B).
miR-210-3p inhibitor promotes hypoxic
injury-induced NSC apoptosis, and AIF and BNIP3 expression
levels
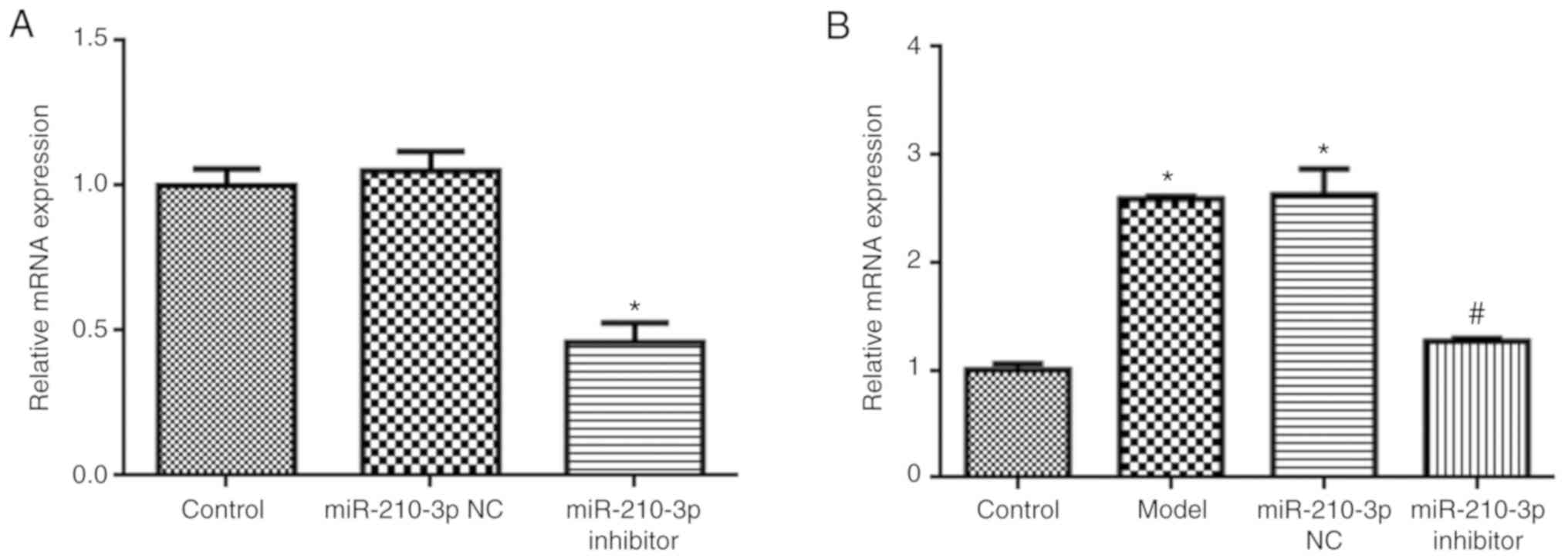
Compared with the control group, miR-210-3p
expression was significantly reduced in miR-210-3p
inhibitor-treated NSCs (P<0.05; Fig. 4A). Additionally, miR-210-3p
inhibitor also reduced miR-210-3p expression levels compared with
the model group (P<0.05; Fig.
4B). In addition, compared with the model group, miR-210-3p
inhibitor increased NSC apoptosis (P<0.05; Fig. 5), and the expression levels of AIF
and BNIP3 (P<0.05; Fig. 6).
Discussion
The present study demonstrated that MSC-EVs reduced
hypoxia injury-induced NSC apoptosis and promoted miR-210-3p
expression. miR-210-3p inhibitor also promoted hypoxia
injury-induced NSC apoptosis. Collectively, the results indicated
that MSC-EVs possibly reduced apoptosis by modulating the
miR-210-3p/AIF/BNIP3 signaling pathway.
Cobalt chloride triggered NSC apoptosis and caused
NSC hypoxic injury (21,22). The signaling pathways involved in
NSC apoptosis have been investigated; Mfat-1 transgene
protects NSCs against cobalt chloride-mediated hypoxic injury by
activating Nrf2/ARE signaling pathways (21). The present study demonstrated that
MSC-EVs also protected NSCs against cobalt chloride-induced NSC
apoptosis. MSCs are beneficial for a variety of diseases including
stroke and infection diseases, primarily via self-differentiation
and secretory vesicles (25),
especially EVs (26,27). EVs reach the injured area more
easily than MSCs, as they can pass tissue barriers, such as the
blood-spinal cord barrier, with ease (28). In addition, EVs are not susceptible
to degradation in comparison with other biological products
(29–31); therefore, EVs may serve as
therapeutic agents for a range of diseases.
In the present study, EVs were extracted from MSCs,
and the transmission electron microscopy and particle size analysis
results indicated that the EVs were >100 nm in size, which
indicated the concentration and the purity of the extracted
exosomes. NSCs were extracted from the hippocampus and cultured
in vitro. The results suggested that the positive rate of
nestin, a specific protein of neural stem cells, was >98% in the
extracted cells, which indicated that the isolation of NSCs from
the hippocampus of SD rats was successful. A previous study
indicated that exosome therapy can reduce the permeability of the
blood-spinal cord barrier following spinal cord injury (32). The results of the present study
also suggested that NSC apoptosis was increased in the hypoxic
injury model group compared with the control group, but decreased
following treatment with MSC-EVs.
The present study indicated that miR-210-3p
expression was upregulated under hypoxic conditions compared with
control conditions. miR-210-3p is an important post-transcriptional
regulator that can inhibit MAX network transcriptional repressor
expression, which is closely related to hypoxia (33). miR-210-3p can be activated by
hypoxia inducible factor-1α to promote cell survival (34). Reducing miR-210-3p expression
antagonizes oxygen-glucose deprivation-induced injury (35). miR-210-3p is also an important
regulator of cell apoptosis and proliferation (36). The present study revealed that
MSC-EVs further promoted miR-210-3p expression in hypoxia injured
NSCs and elevated miR-210-3p expression was beneficial for cell
survival. miR-210-3p expression levels were decreased following
treatment with miR-210-3p inhibitor in normal NSCs, as well as in
hypoxia injured NSCs compared with control NSCs. Moreover,
miR-210-3p inhibitor promoted hypoxic injury-induced NSC
apoptosis.
The Bcl-2 family is involved in the process of cell
injury (37). BNIP3 protein is the
most sensitive apoptotic protein to hypoxia (38). BNIP3-induced cell death is
characterized by increased plasma membrane permeability and
mitochondrial damage in the early stages, but is not accompanied by
caspase activation and cytochrome c release (39). AIF is normally located in the
mitochondrial membrane gap, but during the stress reaction, AIF is
released from mitochondria to cytoplasm and then transferred to
nucleus (40). In the nucleus, AIF
activates endogenous nucleic acid endonuclease (41). AIF-dependent cell death cannot be
reduced by caspase inhibitors, suggesting that AIF does not depend
on caspases (42). Under hypoxic
conditions, the expression of miR-210-3p in NSCs is upregulated
(41). miR-210-3p overexpression
can promote NSC survival and proliferation and its mechanism may be
related to BNIP3 (43). The
present study revealed that the expression levels of BNIP3 and AIF
in NSCs cells were decreased following treatment with MSC-EVs, but
increased by miR-210-3p inhibitor compared with the model group.
miR-210 can target hypoxia-inducible factor 1α to protect renal
cells against hypoxia-induced apoptosis (34). miR-210-3p regulates non-small cell
lung cancer cell proliferation and apoptosis by targeting paired
amphipathic helix protein SIN3 transcription regulator family
member A (44). miR-210-3p also
targets repulsive guidance molecule A to enhance the angiogenic
functions of endothelial progenitor cells under hypoxic conditions
(45). However, whether miR-210-3p
directly regulates BNIP3 and AIF is not completely understood.
The present study had a number of limitations.
First, surface markers, such as cluster of differentiation (CD)9,
CD81 and tumor susceptibility 101 were not detectable in the
MSC-EVs. In the present study, the sizes of the prepared MSC-EVs
were >100 nm, suggesting that these EVs are microvesicles
(46), which typically contain
CD40 as well as cholesterol, sphingomyelin, and ceramide.
Therefore, these surface markers should be detected using sensitive
assay methods in subsequent studies. Secondly, the present study
suggested that MSC-EVs enhanced miR-210-3p expression, but whether
MSC-EVs directly altered miR-210-3p expression was not
investigated.
In summary, MSC-EVs prevented NSC hypoxic injury by
promoting miR-210-3p expression and reducing apoptosis. Therefore,
MSC-EVs may serve as an important treatment strategy for NSC
hypoxic injury.
Acknowledgements
Not applicable.
Funding
The present study was supported by the Basic and
Application Research in Yunnan Province [grant no.
2017FE468(−251)].
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
FL, JZ, RL, YD, LT and YX performed the experiments
and analyzed the data. JZ and AC designed the study and wrote the
manuscript. All authors read and approved the final manuscript.
Ethics approval and consent to
participate
All experimental protocols were approved by the
Ethics Committee of Kunming Medical University (approval no.
KMMU2020180).
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Ma DK, Bonaguidi MA, Ming GL and Song H:
Adult neural stem cells in the mammalian central nervous system.
Cell Res. 19:672–682. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Vishwakarma SK, Bardia A, Tiwari SK,
Paspala SA and Khan AA: Current concept in neural regeneration
research: NSCs isolation, characterization and transplantation in
various neurodegenerative diseases and stroke: A review. J Adv Res.
5:277–294. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Babenko VA, Silachev DN, Popkov VA, Zorova
LD, Pevzner IB, Plotnikov EY, Sukhikh GT and Zorov DB: Miro1
enhances mitochondria transfer from multipotent mesenchymal stem
cells (mmsc) to neural cells and improves the efficacy of cell
recovery. Molecules. 23:6872018. View Article : Google Scholar
|
|
4
|
Zahr SK, Yang G, Kazan H, Borrett MJ,
Yuzwa SA, Voronova A, Kaplan DR and Miller FD: A Translational
repression complex in developing mammalian neural stem cells that
regulates neuronal specification. Neuron. 97:520–537 e6. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Xu X, Warrington AE, Bieber AJ and
Rodriguez M: Enhancing CNS repair in neurological disease:
Challenges arising from neurodegeneration and rewiring of the
network. CNS Drugs. 25:555–573. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Ullah I, Subbarao RB and Rho GJ: Human
mesenchymal stem cells-current trends and future prospective.
Biosci Rep. 35:e001912015. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Zhang X, Sai B, Wang F, Wang L, Wang Y,
Zheng L, Li G, Tang J and Xiang J: Hypoxic BMSC-derived exosomal
miRNAs promote metastasis of lung cancer cells via STAT3-induced
EMT. Mol Cancer. 18:402019. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Lee KW, Kim DH, Lee JH and Youn YN: The
effect of pulsatile flow on bmsc-derived endothelial-like cells in
a small-sized artificial vessel made by 3-dimensional bioprinting.
Stem Cells Int. 2018:78238302018. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Chen L, Lu FB, Chen DZ, Wu JL, Hu ED, Xu
LM, Zheng MH, Li H, Huang Y, Jin XY, et al: BMSCs-derived
miR-223-containing exosomes contribute to liver protection in
experimental autoimmune hepatitis. Mol Immunol. 93:38–46. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Wang Y, Zhao R, Liu D, Deng W, Xu G, Liu
W, Rong J, Long X, Ge J and Shi B: Exosomes derived from
miR-214-enriched bone marrow-derived mesenchymal stem cells
regulate oxidative damage in cardiac stem cells by targeting
caMKII. Oxid Med Cell Longev. 2018:49712612018. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Baglio SR, Rooijers K, Koppers-Lalic D,
Verweij FJ, Pérez Lanzón M, Zini N, Naaijkens B, Perut F, Niessen
HW, Baldini N and Pegtel DM: Human bone marrow- and
adipose-mesenchymal stem cells secrete exosomes enriched in
distinctive miRNA and tRNA species. Stem Cell Res Ther. 6:1272015.
View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Su T, Xiao Y, Xiao Y, Guo Q, Li C, Huang
Y, Deng Q, Wen J, Zhou F and Luo XH: Bone amesenchymal stem
cells-derived exosomal MiR-29b-3p regulates Aging-associated
insulin resistance. ACS Nano. 13:2450–2462. 2019.PubMed/NCBI
|
|
13
|
Mead B and Tomarev S: Bone marrow-derived
mesenchymal stem cells-derived exosomes promote survival of retinal
ganglion cells through miRNA-Dependent mechanisms. Stem cells
Transl Med. 6:1273–1285. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Santos JC, Lima NDS, Sarian LO, Matheu A,
Ribeiro ML and Derchain SFM: Exosome-mediated breast cancer
chemoresistance via miR-155 transfer. Sci Rep. 8:8292018.
View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Lin Y, Wu J, Gu W, Huang Y, Tong Z, Huang
L and Tan J: Exosome-Liposome hybrid nanoparticles deliver
CRISPR/Cas9 System in MSCs. Adv Sci (Weinh). 5:17006112018.
View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Michlewski G and Cáceres JF:
Post-transcriptional control of miRNA biogenesis. RNA. 25:1–16.
2019. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Yang S, Mi X, Chen Y, Feng C, Hou Z, Hui R
and Zhang W: MicroRNA-216a induces endothelial senescence and
inflammation via Smad3/IkappaBα pathway. J Cell Mol Med.
22:2739–2749. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Huang X, Le QT and Giaccia AJ:
MiR-210-micromanager of the hypoxia pathway. Trends Mol Med.
16:230–237. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Ma X, Wang J, Li J, Ma C, Chen S, Lei W,
Yang Y, Liu S, Bihl J and Chen C: Loading MiR-210 in endothelial
progenitor cells derived exosomes boosts their beneficial effects
on hypoxia/reoxygeneation-injured human endothelial cells via
protecting mitochondrial function. Cell Physiol Biochem.
46:664–675. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Bavelloni A, Ramazzotti G, Poli A, Piazzi
M, Focaccia E, Blalock W and Faenza I: MiRNA-210: A current
overview. Anticancer Res. 37:6511–6521. 2017.PubMed/NCBI
|
|
21
|
Yu J, Yang H, Fang B, Zhang Z, Wang Y and
Dai Y: Mfat-1 transgene protects cultured adult neural stem cells
against cobalt chloride-mediated hypoxic injury by activating
Nrf2/ARE pathways. J Neurosci Res. 96:87–102. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Walls KC, Ghosh AP, Ballestas ME, Klocke
BJ and Roth KA: Bcl-2/Adenovirus E1B 19-kd interacting protein 3
(BNIP3) regulates hypoxia-induced neural precursor cell death. J
Neuropathol Exp Neurol. 68:1326–1338. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Song Z, Chen H, Xu W, Wu S and Zhu G:
Basolateral amygdala calpain is required for extinction of
contextual fear-memory. Neurobiol Learn Mem. 155:180–188. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Zuo R, Liu M, Wang Y, Li J, Wang W, Wu J,
Sun C, Li B, Wang Z, Lan W, et al: BM-MSC-derived exosomes
alleviate radiation-induced bone loss by restoring the function of
recipient BM-MSCs and activating Wnt/β-catenin signaling. Stem Cell
Res Ther. 10:302019. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Xu T, Xu M, Bai J, Lin J, Yu B, Liu Y, Guo
X, Shen J, Sun H, Hao Y and Geng D: Tenocyte-derived exosomes
induce the tenogenic differentiation of mesenchymal stem cells
through TGF-β. Cytotechnology. 71:57–65. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Yu B, Shao H, Su C, Jiang Y, Xiteng Y, Bai
L, Zhang Y, Li Q, Zhang X and Li X: Exosomes derived from MSCs
ameliorate retinal laser injury partially by inhibition of MCP-1.
Sci Rep. 6:345622016. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Lu Y, Zhou Y, Zhang R, Wen L, Wu K, Li Y,
Yao Y, Duan R and Jia Y: Bone mesenchymal stem cell-derived
extracellular vesicles promote recovery following spinal cord
injury via improvement of the integrity of the blood-spinal cord
barrier. Front Neurosci. 13:2092019. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Gilligan KE and Dwyer RM: Engineering
exosomes for cancer therapy. Int J Mol Sci. 18:11222017. View Article : Google Scholar
|
|
30
|
Lou G, Song X, Yang F, Wu S, Wang J, Chen
Z and Liu Y: Exosomes derived from miR-122-modified adipose
tissue-derived MSCs increase chemosensitivity of hepatocellular
carcinoma. J Hematol Oncol. 8:1222015. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Li X, Wang S, Zhu R, Li H, Han Q and Zhao
RC: Lung tumor exosomes induce a Pro-inflammatory phenotype in
mesenchymal stem cells via NFκB-TLR signaling pathway. J Hematol
Oncol. 9:422016. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Kong FL, Wang XP, Li YN and Wang HX: The
role of exosomes derived from cerebrospinal fluid of spinal cord
injury in neuron proliferation in vitro. Artif Cells Nanomed
Biotechnol. 46:200–205. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Wang Z, Deng M, Liu Z and Wu S:
Hypoxia-induced miR-210 promoter demethylation enhances
proliferation, autophagy and angiogenesis of schwannoma cells.
Oncol Rep. 37:3010–3018. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Liu LL, Li D, He YL, Zhou YZ, Gong SH, Wu
LY, Zhao YQ, Huang X, Zhao T, Xu L, et al: miR-210 protects renal
cell against hypoxia-induced apoptosis by targeting HIF-1 alpha.
Mol Med. 23:258–271. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Costales MG, Haga CL, Velagapudi SP,
Childs-Disney JL, Phinney DG and Disney MD: Small Molecule
inhibition of microRNA-210 reprograms an oncogenic hypoxic circuit.
J Am Chem Soc. 139:3446–3455. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Sun LL, Li WD, Lei FR and Li XQ: The
regulatory role of microRNAs in angiogenesis-related diseases. J
Cell Mol Med. 22:4568–4587. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Tsujimoto Y: Role of Bcl-2 family proteins
in apoptosis: Apoptosomes or mitochondria? Genes Cells. 3:697–707.
1998. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Burton TR and Gibson SB: The role of Bcl-2
family member BNIP3 in cell death and disease: NIPping at the heels
of cell death. Cell Death Differ. 16:515–523. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Jin Q, Li R, Hu N, Xin T, Zhu P, Hu S, Ma
S, Zhu H, Ren J and Zhou H: DUSP1 alleviates cardiac
ischemia/reperfusion injury by suppressing the Mff-required
mitochondrial fission and Bnip3-related mitophagy via the JNK
pathways. Redox Biol. 14:576–587. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Bano D and Prehn JHM: Apoptosis-Inducing
factor (AIF) in physiology and disease: The tale of a repented
natural born killer. EBioMedicine. 30:29–37. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Liu J, Yuan C, Pu L and Wang J: Nutrient
deprivation induces apoptosis of nucleus pulposus cells via
activation of the BNIP3/AIF signalling pathway. Mol Med Rep.
16:7253–7260. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Benitez-Guzman A, Arriaga-Pizano L, Moran
J and Gutierrez-Pabello JA: Endonuclease G takes part in
AIF-mediated caspase-independent apoptosis in Mycobacterium
bovis-infected bovine macrophages. Veterinary Res. 49:692018.
View Article : Google Scholar
|
|
43
|
Luan Y, Zhang X, Zhang Y and Dong Y:
MicroRNA-210 protects PC-12 cells against hypoxia-induced injury by
targeting BNIP3. Front Cell Neurosci. 11:2852017. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Ren J, Li X, Dong H, Suo L and Zhang J,
Zhang L and Zhang J: miR-210-3p regulates the proliferation and
apoptosis of non-small cell lung cancer cells by targeting SIN3A.
Exp Ther Med. 18:2565–2573. 2019.PubMed/NCBI
|
|
45
|
Lu WJ, Liang HB, Li YF, Tu XQ, He JR, Ding
KQ, Yang GY, Xin XY and Zeng LL: MicroRNA-210-3p targets RGMA to
enhance the angiogenic functions of endothelial progenitor cells
under hypoxic conditions. Front Cell Neurosci. 13:2232019.
View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Keshtkar S, Azarpira N and Ghahremani MH:
Mesenchymal stem cell-derived extracellular vesicles: Novel
frontiers in regenerative medicine. Stem Cell Res Ther. 9:632018.
View Article : Google Scholar : PubMed/NCBI
|