Introduction
Schizophrenia (SCZ) is a complex and severe
psychiatric disorder that affects ~1% of the general population
worldwide, placing a heavy burden on the patients, the patients'
family and society (1). Although
administration of antipsychotics remains the major therapeutic
strategy for SCZ, ~ one-third of patients are insensitive to them
(2). Patients who only have a
minor or no response to at least two non-clozapine drugs at
adequate doses and treatment cycles are diagnosed with
treatment-resistant SCZ (TRS) according to the guidelines of the
American Psychiatric Association (3). At present, frequent rehospitalization
and repeated episodes of symptoms in patients with TRS increase the
burden on patients and their families (4). It is currently thought that a
combination of environmental and genetic factors is most likely to
be the major cause of SCZ. However, the precise diagnosis of SCZ
still only depends on clinical symptoms, without the help of
sensitive and objective molecular markers. It is therefore urgent
and necessary to identify sensitive biomarkers for the early and
accurate diagnosis of SCZ, so that earlier and effective
therapeutic intervention may be provided (5–7).
Nowadays, strong evidence from psychiatric clinical studies has
indicated that environmental factors may result in aberrant gene
expression in the central nervous system (CNS), which may be the
major cause of symptom onset (8,9). A
study has indicated that ≥50% of changes in gene expression in the
CNS are involved in the changes in the expression of the same genes
in peripheral tissues (10). Since
brain tissues, several of which have deeply located and complex
structures with difficult access, are challenging to directly
investigate, the present study attempted to examine and elucidate
potential biomarkers for SCZ via a number of blood-derived gene
expression analyses (10–14). By using this non-invasive method,
effective peripheral blood biomarkers may be discovered that may be
considerably helpful in promoting earlier TRS diagnosis, as well as
enabling the concise assessment of treatment response and even
precisely predicting prognosis.
Recently, increasing attention has been paid to the
regulatory mechanisms of microRNAs (miRNAs/miRs) (15–17),
which are considered to be in charge of gene expression monitoring
through the inhibition of mRNA translation (18). Previous studies have indicated that
changes in the expression of certain mRNAs reflect parallel changes
in the CNS (19) and peripheral
blood (20,21). The notion that certain miRNAs may
be sensitive biomarkers for SCZ has gained much research interest
(22–27), since alterations in their
expression in peripheral blood mononuclear cells (PBMCs) may
provide valuable clues for the earlier and more precise diagnosis
and more effective treatment of SCZ (28). It is worth noting that abnormally
expressed miRNAs have been detected in the PBMCs and plasma of
patients with SCZ. In a recent study, Lai et al (28) indicated that 7 miRNAs, namely
miR-31, miR-431, miR-433, miR-107, miR-134, miR-99b and miR-487b,
were downregulated in the PBMCs derived from 112 patients with
mental illness, as compared with those of a control group of 76
non-psychotic patients. Of note, the imprinted delta-like homologue
1 and the type III iodothyronine deiodinase (DLK1-DIO3) region of
14q32 from PBMCs was considered an SCZ-associated miRNA signature
(28). In that study, a total of 7
differentially expressed miRNAs [Homo sapiens (hsa)-miR-34a,
miR-449a, miR-564, miR-432, miR-548d, miR-572 and miR-652] were
identified to be closely associated with emotional retardation,
restricted language and other neurocognitive dysfunctions in 30
patients with SCZ, as compared with 30 controls (28). In another noteworthy study,
specific miRNA30e in the plasma was identified as a biomarker of
SCZ diagnosis from the expressed profiles of 9 SCZ-associated
miRNAs in PBMCs by means of reverse transcription-quantitative PCR
(RT-qPCR) (29). Subsequently,
miR-130b and miR-193a-3p were identified as biomarkers for the
diagnosis of SCZ by high-throughput sequencing and RT-qPCR
(30). The early growth response
protein-1 (EGR1)/miR-30a-5p/neurogenic differentiation 1 (NEUROD1)
axis was proposed as a promising biomarker for the diagnosis and
treatment monitoring of patients with SCZ with acute mental states
based on RT-qPCR technology (31).
miRNA analysis may therefore be used to effectively
elucidate the etiology of SCZ and identify potential differences
among mental illnesses with similar but unclear clinical symptoms
(32). The present study aimed to
identify novel and specific biomarkers to earlier and more
precisely diagnose patients with TRS and implement effective
therapeutic strategies. Second-generation high-throughput
sequencing technology and bioinformatics were used to screen out
and analyze the top three abnormally expressed miRNAs in PBMCs from
6 patients with TRS and 6 healthy controls. Next, RT-qPCR was used
to validate the significantly differentially expressed miRNAs in a
larger sample of cases and healthy controls. The newly identified
target genes of the two confirmed differentially expressed miRNAs
were predicted with the online software TargetScan and Diana-micro
T, and bioinformatics methods further indicated that these target
genes were closely associated with functions of the nervous system.
Finally, these two miRNAs were proposed as diagnostic biomarkers
for TRS, thereby providing a novel, simple and convenient
diagnostic tool for this condition.
Materials and methods
Screening and collection of cases
The subjects of the present study were divided into
two groups, the case group and the control group. For the case
group, a total of 34 patients with TRS (17 males and 17 females)
were recruited from the Psychiatric inpatient ward of Honghe Second
People's Hospital and Yuxi Second People's Hospital (Yunnan,
China). The inclusion criteria were as follows: i) All patients met
the mental illness criteria outlined in the Diagnostic and
Statistical Manual Fifth Edition Criteria for SCZ (DSM-5; American
Psychiatric Association; 2013) (33); ii) Complete clinical data and iii)
Voluntary participation in the study. These 34 patients were
screened by 2 independent experienced deputy chief physicians,
according to the inclusion criteria. The exclusion criteria were
conditions such as physical or cerebral organic disease, other
types of mental illness, drug abuse prior to hospitalization,
patients who received modified electroconvulsive therapy within 3
months from enrollment, and patients with a history of blood
transfusion within the last month.
The control group included 31 healthy individuals
with no family history of major mental illness recruited at the
Physical Examination Center of the Sixth Affiliated Hospital of
Kunming Medical University (Kunming, China). Of the 31 control
individuals, 16 were males and 15 females, all of whom had a normal
physical examination and psychological examination to exclude the
presence of SCZ, none of whom had suffered any recent adverse life
events. The age and sex ratio of patients and controls was 2:1.
All included subjects were older than 18 years.
According to the requirements of the ethics committee, written
informed consent for the case group was required not only from the
patient but also one family member/guardian, while for the healthy
control group, informed consent was provided by each subject
themselves. The present study was approved by the ethics committees
of Honghe Second People's Hospital (Honghe, Yunnan; approval no.
20190013) and The Sixth Affiliated Hospital of Kunming Medical
University (Yuxi, Yunnan; approval no. 2020kmykdx6h58).
Extraction of PBMCs
Approximately 5 ml peripheral venous blood was
collected by quick blood sampling from 65 cases and processed
within 1 h. This procedure was carried out according to the
manufacturer's protocol (Dakwei Biological Engineering). In brief,
fresh whole blood was collected in EDTA-K2 anti-coagulation tubes
and then diluted with an equal volume of isotonic solution (normal
saline). Subsequently, 15 ml separation liquid was added to the
centrifuge tube. The volume ratio of the separation liquid,
anticoagulated undiluted whole blood and isotonic solution
(physiological saline) was 1:1:1. Next, the sample was centrifuged
at 700–800 × g for 20–30 min at room temperature. Between the
plasma and separation layers was a thin and dense white film. The
white film layer was carefully pipetted into another centrifuge
tube. The horizontal rotor was then centrifuged at 250 × g at room
temperature for 10 min and the supernatant was discarded. Washing
was repeated 1–2 times. Finally, 1 ml TRIzol®
(Invitrogen; Thermo Fisher Scientific, Inc.) was added and the
sample was mixed. A total of 1.5 ml was added to the labeled
cryotube. The cryotube was tightly sealed with parafilm and stored
at −80°C.
RNA extraction and miRNA
sequencing
RNA samples were harvested from 6 patients with TRS
(3 males, 3 females) and 6 healthy controls (3 males, 3 females).
The patients in the experimental groups were age-matched. Samples
were delivered to Guangzhou Gidio Biotechnology Co., Ltd. for the
extraction of small RNA and the construction of small RNA
libraries. In brief, RNA was extracted using the TRIzol method,
followed by detection of RNA purity with a NanoDrop 2000 (Thermo
Fisher Scientific, Inc.) and RNA integrity with an Agilent 2100
Bioanalyzer (Agilent Technologies GmbH). Small RNAs were obtained
by gel electrophoresis from total RNA, and 5′ and 3′ ends were
joined. RT-qPCR was performed to construct small RNA libraries.
Subsequently, the constructed libraries were tested for quality and
yield using an Agilent 2100 and ABI StepOnePlus Real-Time PCR
System (Thermo Fisher Scientific, Inc.). An Illumina HiSeq2000
(Illumina, Inc.) was used for sequencing. All procedures were in
strict accordance with the manufacturer's protocols (Gidio
Biotechnology Co., Ltd.)
RT-qPCR
RT-qPCR was used to validate the top three
aberrantly expressed miRNAs in 34 patients with TRS and 31 healthy
controls. In brief, after total RNA was isolated from PBMCs,
complementary (c)DNA was synthetized using the miRcute Plus miRNA
First-Strand cDNA kit and miRcute Plus miRNA qPCR kit (SYBR Green)
according to the manufacturer's protocol. RT-qPCR was performed in
the Quantitative PCR Reaction Amplifier (CFX96 Touch; Bio-Rad
Laboratories, Inc.), with the reaction system including 2X miRcute
Plus miRNA Premix (SYBR and ROX), PCR forward primer and PCR
reverse primer. The primers for three miRNAs are summarized in
Table SI. Detailed experimental
instruments and reagents are included in Tables SII and SIII. h-actin for miRNA was applied to
normalize the results. The expression levels of miRNAs were
calculated using the 2−ΔΔCq method (34). All RT-qPCR validation procedures
were performed by Wuhan Genecreate Biological Engineering Co.,
Ltd.
miRNA target gene prediction and
bioinformatics analysis
TargetScan release 7.2 (http://www.targetscan.org/) (35) and Diana-microT v5.0 (http://diana.imis.athena-innovation.gr/DianaTools/index.php?r=microT_CDS/index)
(36), which are two major and
widely accepted open databases, were used to predict the target
genes of the significantly differentially expressed miRNAs.
Subsequently, the target genes obtained from the two databases were
intersected and underwent further GO and Kyoto Encyclopedia of
Genes and Genomes (KEGG) pathway enrichment analysis. GO annotation
and KEGG enrichment analysis of the target genes were performed
using the OmicShare tools (37), a
free online platform for data analysis (http://www.omicshare.com/tools). The GO project covers
three classical domains, namely biological process (BP), cellular
component and molecular function. The KEGG is a database that
integrates genome, chemistry, and system function information, and
aims to reveal the genetic and chemical blueprint of biological
phenomena. P<0.05 was considered to indicate a statistically
significant result. The data were analyzed and obtained using the
OmicShare cloud platform (https://www.omicshare.com/).
Statistical analysis
SPSS 22 (IBM Corp.) and GraphPad Prism 7.0 (GraphPad
Software, Inc.) were used for data analysis. The χ2 test
for the qualitative variables and the t-test for the quantitative
variables were respectively used for the demographic analysis of
the case and control groups. The Mann-Whitney U test was used to
compare the differential expression levels of miRNAs. Receiver
operating characteristic (ROC) curve analysis was used to assess
the area under the curve (AUC). P<0.05 was considered to
indicate a statistically significant difference.
Results
Demographic data of the subjects
All subjects were Han Chinese. The mean age (mean ±
standard deviation) of TRS cases and controls was 41.82±11.414 and
37.06±11.363 years, respectively. There were no significant
differences in terms of age or sex between the cases and healthy
controls (Table I).
 | Table I.Demographic data of the cases and
healthy controls. |
Table I.
Demographic data of the cases and
healthy controls.
| Item | Cases (n=34) | Controls
(n=31) |
P-valuea |
|---|
| Sex |
|
| 0.897 |
|
Male | 17 (50.0) | 16 (51.6) |
|
|
Female | 17 (50.0) | 15 (48.4) |
|
| Age (years) | 41.82±11.414 | 37.06±11.363 | 0.956 |
miRNA expression profile analysis
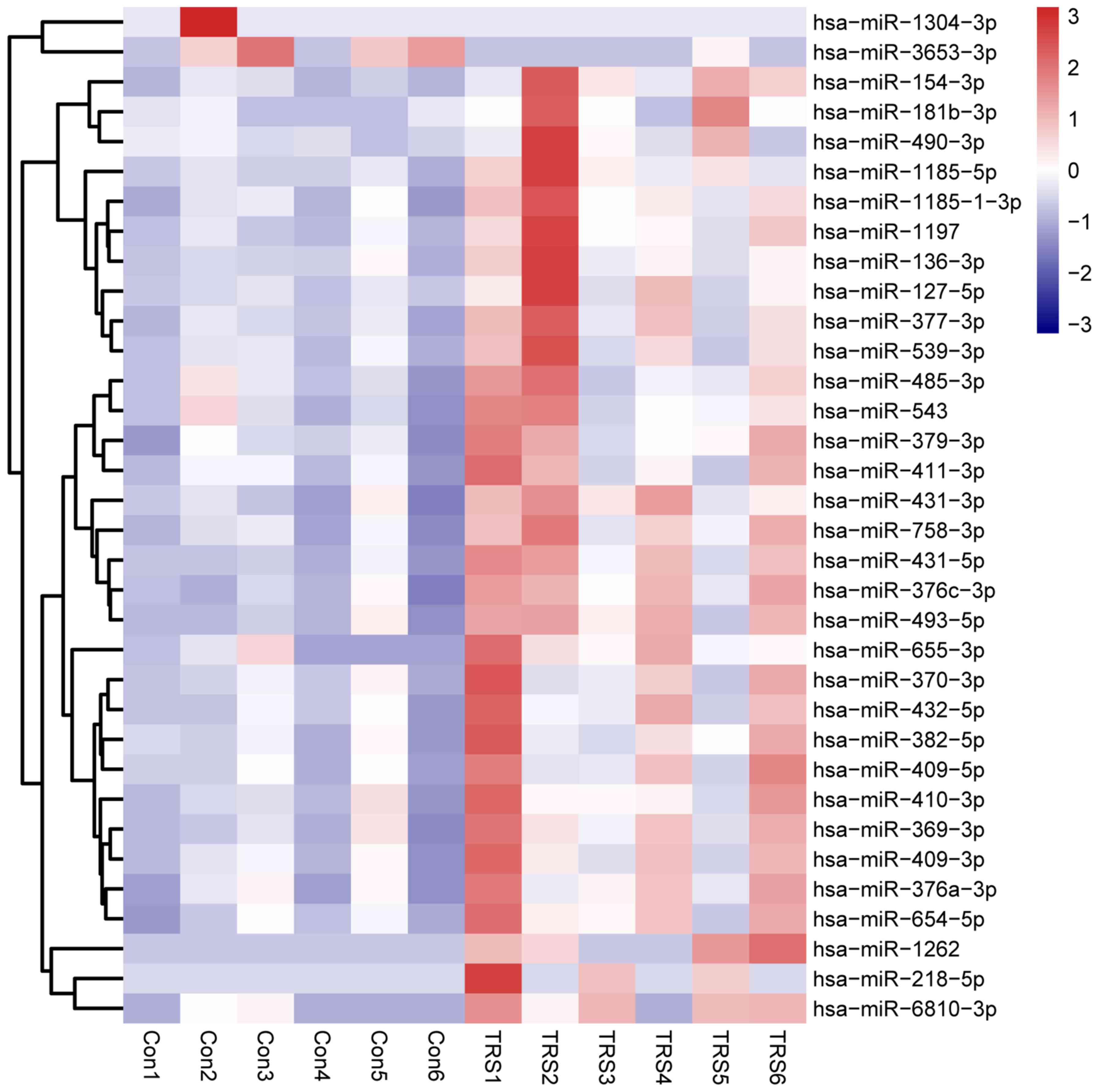
Analysis of the miRNA sequencing data revealed that
a total of 34 miRNAs were significantly differentially expressed in
the 6 cases of TRS as compared with the 6 controls (|log2 (fold
change)|>1; P<0.05). Among the six miRNAs with significantly
differential expression, two were markedly downregulated and four
were upregulated (Table II). A
heat map was constructed to display the differential hierarchical
clustering distribution between TRS and control samples (Fig. 1).
| Table II.Differentially expressed miRNAs in
peripheral blood mononuclear cells from cases vs. healthy
controls. |
Table II.
Differentially expressed miRNAs in
peripheral blood mononuclear cells from cases vs. healthy
controls.
| miRNA | Log2FC | P-value | Direction of
change |
|---|
|
hsa-miR-1185-1-3p | 1.38018786 | 0.014888805 | Up |
|
hsa-miR-1185-5p | 2.055813817 | 0.01030376 | Up |
| hsa-miR-1197 | 1.669823494 | 0.003827819 | Up |
| hsa-miR-1262 | 4.449671416 | 0.01862468 | Up |
| hsa-miR-127-5p | 1.663957827 | 0.010180048 | Up |
|
hsa-miR-1304-3p | −7.019942788 | 0.004047489 | Down |
| hsa-miR-136-3p | 1.536408035 | 0.010754284 | Up |
| hsa-miR-154-3p | 2.635532322 | 0.001485503 | Up |
|
hsa-miR-181b-3p | 2.399752284 | 0.037210273 | Up |
| hsa-miR-218-5p | 4.289711531 | 0.046989353 | Up |
|
hsa-miR-3653-3p | −3.106775941 | 0.031498569 | Down |
| hsa-miR-369-3p | 1.219176104 | 0.035538469 | Up |
| hsa-miR-370-3p | 1.383232956 | 0.034880938 | Up |
|
hsa-miR-376a-3p | 1.265825554 | 0.041422801 | Up |
|
hsa-miR-376c-3p | 1.281059616 | 0.019343549 | Up |
| hsa-miR-377-3p | 1.68135408 | 0.004843865 | Up |
| hsa-miR-379-3p | 1.33354957 | 0.02582764 | Up |
| hsa-miR-382-5p | 1.164588341 | 0.042177088 | Up |
| hsa-miR-409-3p | 1.215682697 | 0.031732848 | Up |
| hsa-miR-409-5p | 1.33800193 | 0.02587475 | Up |
| hsa-miR-410-3p | 1.144646395 | 0.04797714 | Up |
| hsa-miR-411-3p | 1.188991694 | 0.049323611 | Up |
| hsa-miR-431-3p | 1.433665135 | 0.037350442 | Up |
| hsa-miR-431-5p | 1.703040789 | 0.003189292 | Up |
| hsa-miR-432-5p | 1.265157654 | 0.035781474 | Up |
| hsa-miR-485-3p | 1.118394154 | 0.046032191 | Up |
| hsa-miR-490-3p | 1.792169025 | 0.040858315 | Up |
| hsa-miR-493-5p | 1.499534768 | 0.008400669 | Up |
| hsa-miR-539-3p | 1.558249279 | 0.010618161 | Up |
| hsa-miR-543 | 1.112920365 | 0.044806101 | Up |
| hsa-miR-654-5p | 1.433656533 | 0.014340331 | Up |
| hsa-miR-655-3p | 1.943235022 | 0.013931082 | Up |
|
hsa-miR-6810-3p | 2.183414515 | 0.042813132 | Up |
| hsa-miR-758-3p | 1.339956277 | 0.01416857 | Up |
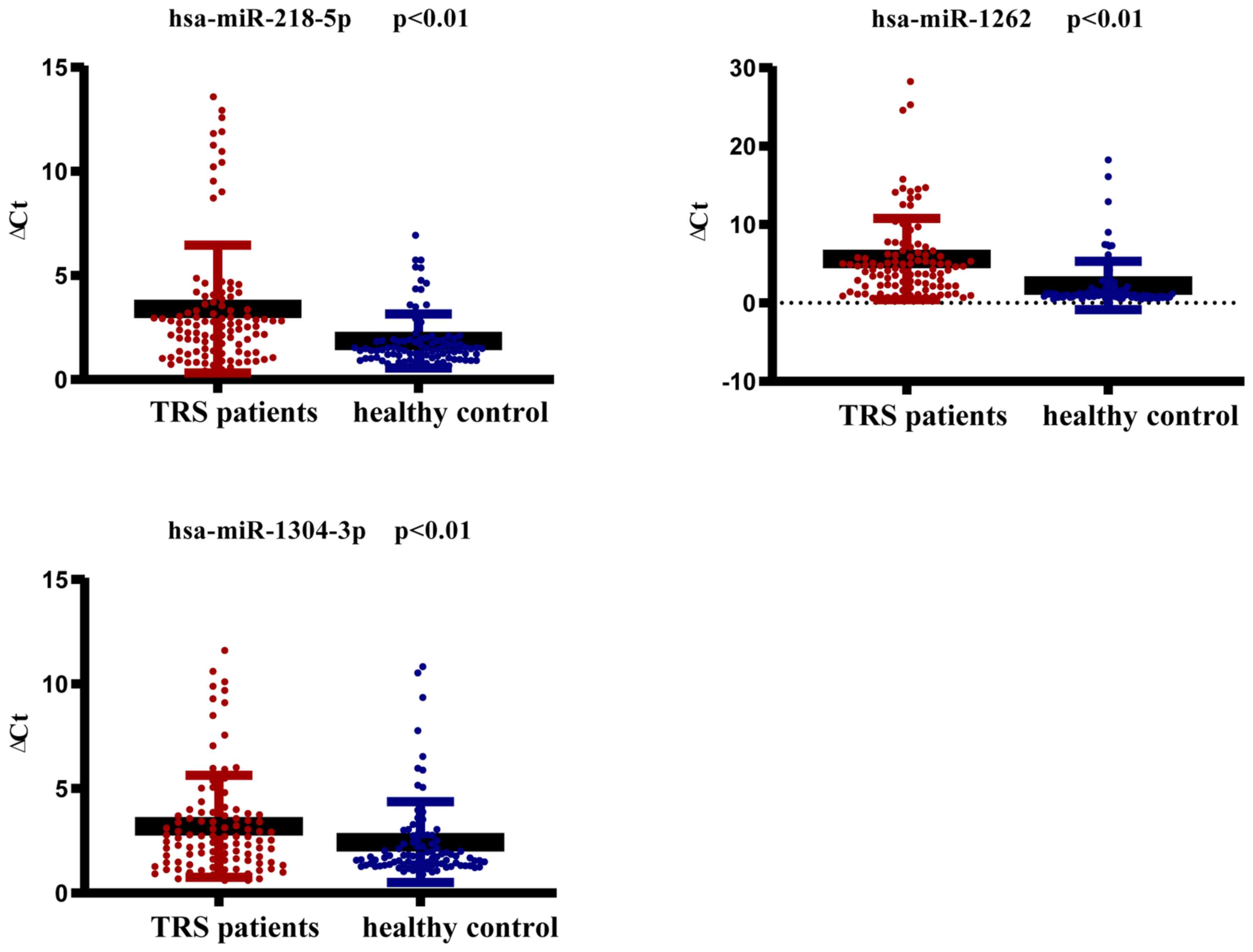
RT-qPCR validation
The three miRNAs (hsa-miR-1262, hsa-miR-1304-3p and
hsa-miR-218-5p) with the highest log2FC values
(|log2FC|>4) were verified in 34 TRS cases and 31
controls by RT-qPCR. Of these three miRNAs, hsa-miR-218-5p and
hsa-miR-1262 were markedly upregulated according to the
next-generation sequencing results, while hsa-miR-1304-3p was
markedly downregulated. The RT-qPCR results indicated that the
expression of hsa-miR-218-5p and hsa-miR-1262 in the TRS cases was
markedly higher than that in the controls, which was consistent
with the sequencing results (P<0.01). However, the expression of
hsa-miR-1304-3p in the TRS cases was also markedly higher than that
in the controls, which was inconsistent with the sequencing results
(P<0.01; Fig. 2).
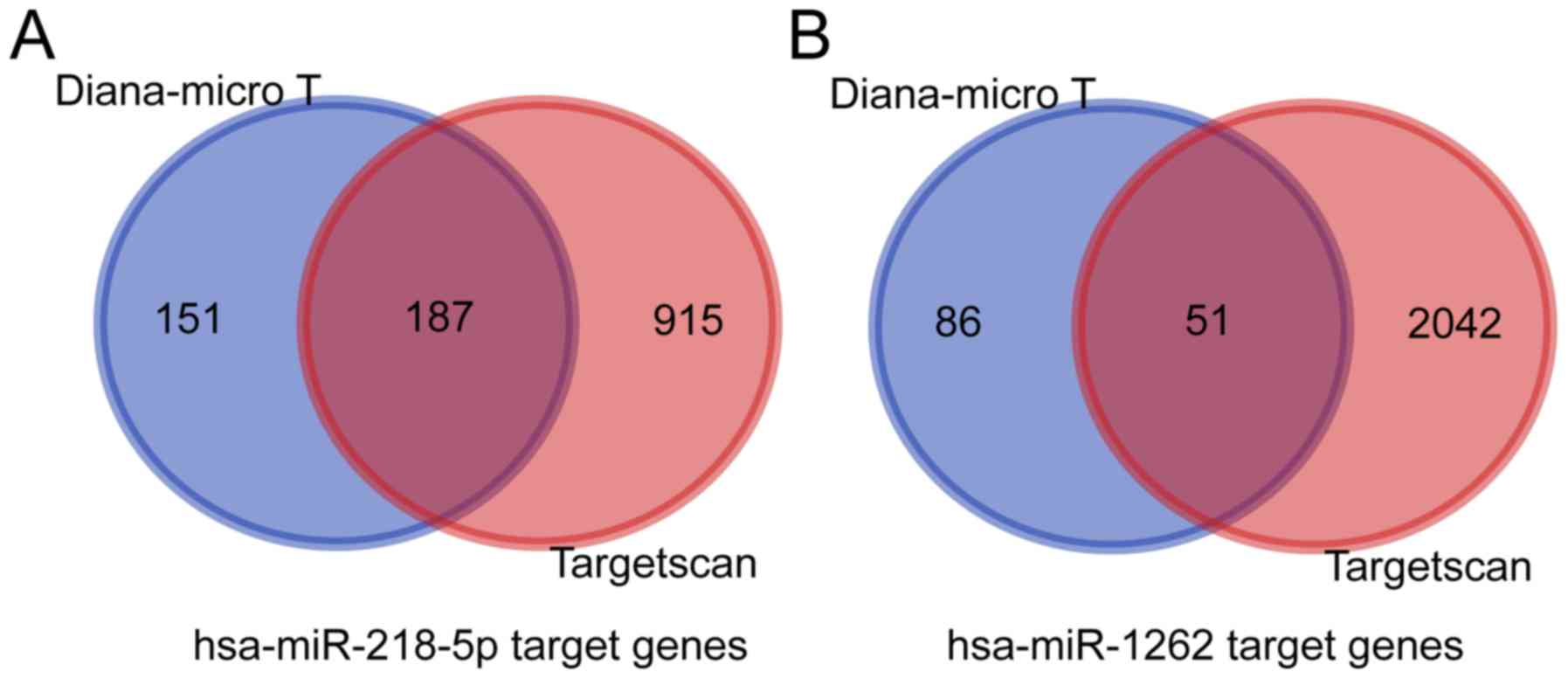
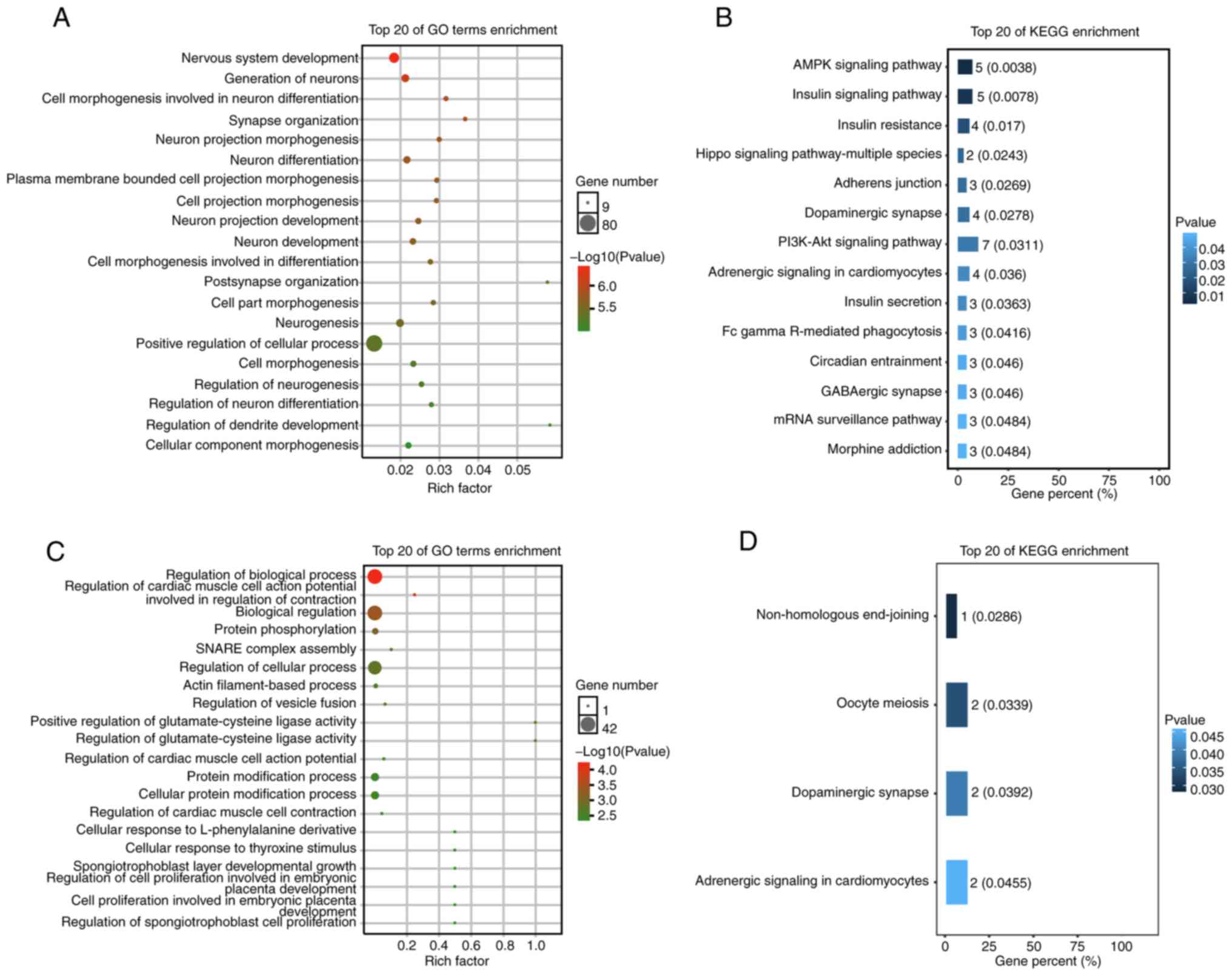
Prediction of miRNA target genes
By using two publicly available and widely accepted
miRNA target gene prediction databases, TargetScan and Diana-micro
T, the target genes of hsa-miR-218-5p and hsa-miR-1262 were
predicted (Fig. 3A and B,
respectively). For hsa-miR-218-5p, 187 target genes were predicted,
while for hsa-miR-1262 had only 51 target genes were obtained.
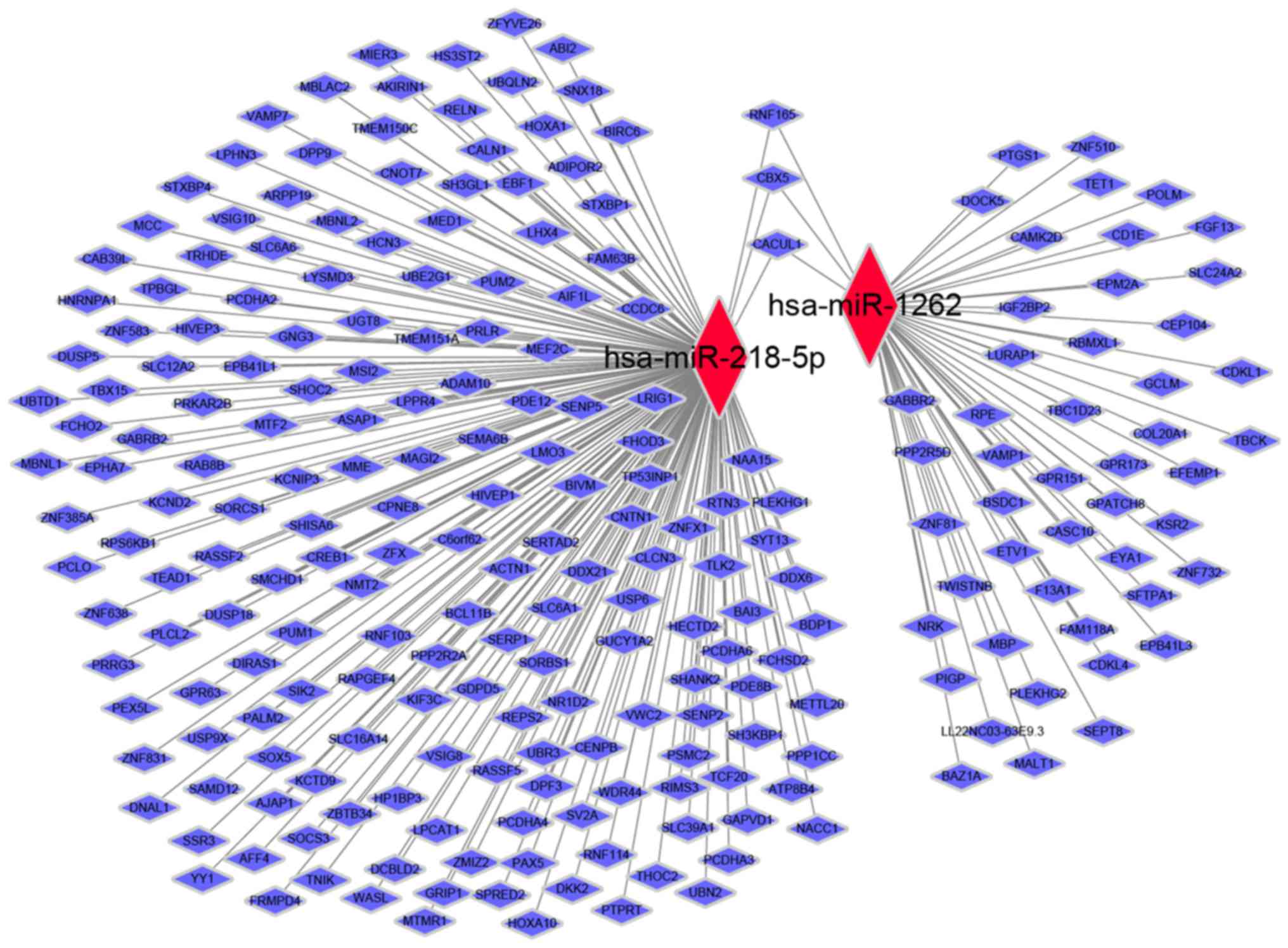
Next, a miRNA-gene co-expression network was generated to summarize
the interaction between miRNAs and target genes (Fig. 4).
GO and pathway enrichment
analysis
Target genes of these two miRNAs were enriched by GO
annotation and KEGG analysis. The results for the top 20 GO terms
enriched by the target genes of hsa-miR-218-5p are presented in
Fig. 5A. In the GO category BP,
the target genes of hsa-miR-218-5p were prominently involved in
various physiological and pathophysiological processes, including
nervous system development, generation of neurons, cell
morphogenesis regarding neuron differentiation, synapse
organization, neuron projection morphogenesis and neuron
differentiation. BP terms associated with brain development and
function are presented in Table
III. KEGG pathway analysis for the target genes of
hsa-miR-218-5p indicated a significant enrichment in several
important pathways including the PI3K/Akt, dopaminergic synapse and
AMPK signaling pathways, as well as the Hippo signaling
pathway-multiple species (Fig.
5B).
| Table III.Top 20 enriched GO Biological Process
terms associated with brain development and functions of
hsa-miR-218-5p target genes. |
Table III.
Top 20 enriched GO Biological Process
terms associated with brain development and functions of
hsa-miR-218-5p target genes.
| GO ID | GO term | Number of enriched
genes | P-value |
|---|
| GO:0007399 | Nervous system
development | 46 |
3.79×10−7 |
| GO:0048699 | Generation of
neurons | 34 |
7.75×10−7 |
| GO:0048667 | Cell morphogenesis
involved in neuron differentiation | 19 |
1.15×10−6 |
| GO:0050808 | Synapse
organization | 16 |
1.34×10−6 |
| GO:0048812 | Neuron projection
morphogenesis | 20 |
1.40×10−6 |
| GO:0030182 | Neuron
differentiation | 31 |
1.73×10−6 |
| GO:0031175 | Neuron projection
development | 25 |
2.26×10−6 |
| GO:0048666 | Neuron
development | 27 |
2.52×10−6 |
| GO:0099173 | Postsynapse
organization | 10 |
2.78×10−6 |
| GO:0022008 | Neurogenesis | 34 |
3.36×10−6 |
| GO:0050767 | Regulation of
neurogenesis | 22 |
5.84×10−6 |
| GO:0045664 | Regulation of
neuron differentiation | 19 |
7.09×10−6 |
| GO:0050773 | Regulation of
dendrite development | 9 |
8.09×10−6 |
| GO:0048523 | Negative regulation
of cellular process | 72 |
1.06×10−5 |
| GO:0016358 | Dendrite
development | 11 |
1.08×10−5 |
| GO:0060996 | Dendritic spine
development | 7 |
2.48×10−5 |
| GO:0050807 | Regulation of
synapse organization | 10 |
4.98×10−5 |
| GO:0060998 | Regulation of
dendritic spine development | 6 |
5.14×10−5 |
| GO:0099504 | Synaptic vesicle
cycle | 9 |
6.40×10−5 |
| GO:0050803 | Regulation of
synapse structure or activity | 10 |
6.76×10−5 |
In the category BP, the GO analysis for the target
genes of hsa-miR-1262 revealed their close association with BP
regulation, biological regulation, cellular process regulation and
cellular protein modification process (Fig. 5C). BP terms closely associated with
brain development and function are presented in Table IV. KEGG pathway analysis for the
target genes of hsa-miR-1262 indicated an obvious enrichment in
crucial pathways, such as nonhomologous end-joining dopaminergic
synapse oocyte meiosis and adrenergic signaling in cardiomyocytes
(Fig. 5D).
| Table IV.Top 20 enriched GO Biological Process
terms associated with brain development and functions of
hsa-miR-1262 target genes. |
Table IV.
Top 20 enriched GO Biological Process
terms associated with brain development and functions of
hsa-miR-1262 target genes.
| GO ID | GO term | Number of enriched
genes | P-value |
|---|
| GO:0045196 | Establishment or
maintenance of neuroblast polarity | 1 | 0.006 |
| GO:0045200 | Establishment of
neuroblast polarity | 1 | 0.007 |
| GO:0055059 | Asymmetric
neuroblast division | 1 | 0.014 |
| GO:0022008 | Neurogenesis | 9 | 0.014 |
| GO:0048812 | Neuron projection
morphogenesis | 5 | 0.018 |
| GO:0031630 | Regulation of
synaptic vesicle fusion to presynaptic active zone membrane | 1 | 0.018 |
| GO:2000465 | Regulation of
glycogen (starch) synthase activity | 1 | 0.021 |
| GO:0007399 | Nervous system
development | 11 | 0.024 |
| GO:0007611 | Learning or
memory | 3 | 0.025 |
| GO:2001223 | Negative regulation
of neuron migration | 1 | 0.027 |
| GO:0048699 | Generation of
neurons | 8 | 0.028 |
| GO:0031175 | Neuron projection
development | 6 | 0.029 |
| GO:1990403 | Embryonic brain
development | 1 | 0.030 |
| GO:0036445 | Neuronal stem cell
division | 1 | 0.032 |
| GO:0055057 | Neuroblast
division | 1 | 0.032 |
| GO:0007613 | Memory | 2 | 0.033 |
| GO:0007272 | Ensheathment of
neurons | 2 | 0.038 |
| GO:0050890 | Cognition | 3 | 0.039 |
| GO:0016082 | Synaptic vesicle
priming | 1 | 0.043 |
| GO:0030182 | Neuron
differentiation | 7 | 0.043 |
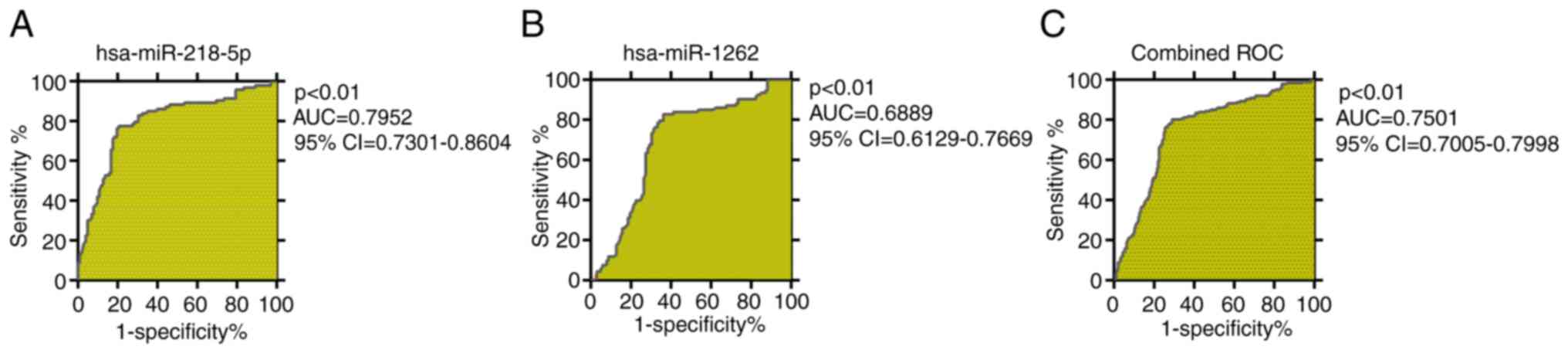
Diagnostic evaluation of
hsa-miR-218-5p and hsa-miR-1262
ROC curve analysis was performed to demonstrate the
potential of the expression levels of the 2 miRNAs confirmed by
RT-qPCR as biomarkers for distinguishing patients with TRS from
healthy controls. As displayed in Fig.
6, the AUC for hsa-miR-218-5p was 0.80 (95% CI, 0.7301–0.8604;
P<0.01; Fig. 6A), the best
cutoff value was 0.62 (specificity, 0.826; sensitivity, 0.794). The
AUC for hsa-miR-1262 was 0.69 (95% CI, 0.6129–0.7669; P<0.01;
Fig. 6B), the best cutoff value
was 0.47 (specificity, 0.828; sensitivity, 0.637). The AUC for the
combination of the 2 miRNAs was able to clearly distinguish
patients with TRS from healthy controls with an AUC of 0.7501 (95%
CI, 0.7005–0.7998; P<0.01; Fig.
6C), the best cutoff value was 0.51 (specificity, 0.801;
sensitivity, 0.711).
Discussion
TRS accounts for 30% of all cases of SCZ (38). Due to its refractory and complex
nature, TRS is a unique and severe phenotype of SCZ and places a
burden on the patients' family and society (39). Therefore, precision medicine for
this particular type of SCZ requires to be urgently implemented in
the clinic. Although precision medicine is a novel healthcare
discipline (40) that gives
priority to the unique characteristics of specific patients
(41), its use in psychiatry is
still lagging compared with other medical fields, since it relies
on neuroimaging and/or biological technology to provide a benefit
(42,43). Several studies have made
outstanding contributions to the development of precise psychiatric
medical treatments. The ENIGMA-Diffusion Tensor Imaging working
group confirmed that the normal variation of human white matter is
genetically associated with mental, neurological and developmental
disorders (44,45). The ENIGMA consortium performed a
meta-analysis of 760 patients with SCZ and 957 healthy participants
and revealed that the cognitive function of patients with SCZ is
associated with the connectivity of the overall brain structure
(46). Another study indicated
that antipsychotic drugs resulted in an acute extrapyramidal
response in the body by acting on two single nucleotide
polymorphisms (SNPs; rs938112 and rs2987902) of two genes (LSMAP
and ABL1) and destroying the binding sites of key transcription
factors (47), LSAMP is an
adhesion molecule of neuronal cells that participates in
Neurogenesis, neurite outgrowth and plasticity (48), Abelson non-receptor tyrosine kinase
(ABL1) is related to neurodegenerative diseases such as Parkinson's
disease (48); the schizophrenia
genomics platform is able to further elucidate the pathogenesis of
SCZ (49) and provide therapeutic
prospects for personalized therapy (50).
It is well known that the therapeutic effect of
antipsychotic drugs is mainly exerted through the blocking of
dopamine D2 receptor and/or the serotonin type 2A receptor to
reduce dopaminergic neurotransmission in the limbic midbrain.
However, even though the molecular targets of psychotropic drugs
have remained to be fully elucidated, several psychotropic drugs
used in the clinic still comprise the cornerstone of psychosis
treatment. Furthermore, clozapine is the only evidence-based
treatment for TRS (51,52), with 60–70% of patients reporting
relief of their symptoms following this treatment (53); however, at present, there are no
evidence-based pharmacotherapies for the remaining 30% of TRS
patients who are not responsive to clozapine (54). Therefore, it is important to
maximize the effectiveness of the drug through personalized
treatment (55). The aim of
pharmacogenomics (PGx) is to study how genes control the
pharmacokinetics and pharmacodynamics of drugs (56). The concept may be used
interchangeably with pharmacogenetics: A specific gene or group of
genes may affect the patient's response to drugs. Ongoing
randomized controlled trials have evaluated the clinical utility of
PGx testing on antipsychotics in patients with SCZ to probe the
association between genes and drugs on SCZ (trial registration nos.
NCT02573168 and NCT02566057). Furthermore, the feasibility of
genome-wide association studies, determination of polygenic risk
scores and epigenetics of DNA methylation may render PGx more
suitable for clinical research (57).
Based on the above concepts of precision medicine
and PGx, even the application of epigenetics in SCZ research
(58), combined with the fact that
TRS is a highly heritable mental disorder and its heritability is
markedly stronger than that of drug-responsive SCZ (59) the aim of the present study was to
explore more sensitive treatment targets for TRS. Multiple studies
have indicated that changes in cells and molecules in the brain may
be relatively reflected by changes in the transcription in PBMCs
(60–62), which may possess conjunct miRNA
expression patterns with the brain (63). A previous study suggested that
overexpressed miR-223 directly acts on mRNAs in plasma by the
method of RT-qPCR and luciferase assay, revealing that it may be
involved in the pathogenesis of SCZ (64). miR-30e, miR-181b, miR-34a, miR-346
and miR-7 in plasma may be useful non-invasive biomarkers for the
diagnosis of SCZ, miR-132, miR-181b, miR-30e and miR-432 in plasma
may be the improvement of symptoms in patients with SCZ, revealing
potential indicators of their prognosis (65). Therefore, we can speculate that the
abnormal expression levels of certain miRNAs in plasma may directly
reflect the gene expression in the brain. Whether the expression of
miRNA in PBMC is indirectly related to the expression of genes in
the brain and the permeability of the blood-brain barrier of miRNAs
need to be further studied. Fortunately, certain studies have
confirmed the value of abnormally expressed miRNAs in the
peripheral blood as biomarkers for SCZ (66,67).
Therefore, this novel, simple and simultaneously much cheaper
strategy was explored by miRNA sequencing data analysis in the MCs
derived from the peripheral blood of patients with TRS. With the
assistance of bioinformatics analysis and RT-qPCR for validation,
two biomarkers, hsa-miR-1262 and hsa-miR-218-5p, were first
identified for the diagnosis of TRS, which exhibited significant
alterations in patients with TRS, as compared with healthy
controls. As there are no reports of these 2 miRNAs being
associated with SCZ or other psychiatric diseases, hsa-miR-218-5p
and hsa-miR-1262 underwent target gene predictions to provide
further insight into the function of these two miRNAs using not
only the TargetScan and Diana-microT online databases, but also GO
and KEGG pathway analysis along with bioinformatics methods. The
functions of these two miRNAs were indicated to be closely
associated with brain development and function, including processes
such as nervous system development, neuronal generation,
neurogenesis and synapse function, and they were also mainly
involved in dopaminergic synapses, Hippo signaling pathway-multiple
species and GABAergic synapses, as demonstrated by KEGG analysis.
Of note, certain studies on TRS candidate genes have focused on the
dopaminergic and serotoninergic systems (68–72).
SCZ is a complex disease that may be of neurodevelopmental origin
and is characterized by abnormal synaptic plasticity at the
cortical and subcortical levels, abnormal regulation of
neurotransmitters (including dopamine, glutamate and serotonin) and
abnormal signaling (73–75). Therefore, the ROC curve analysis
reflected improved ability to differentiate patients with TRS from
healthy controls (AUC value, 0.7501; 95% CI, 0.7005–0.7998;
specificity, 0.801; sensitivity, 0.711) in the present study,
hsa-miR-218-5p and hsa-miR-1262, which have a critical role in the
pathophysiological changes of TRS, were indicated to be promising
sensitive biomarkers for TRS diagnosis, therapy and prediction.
The miRNA-gene network analysis of the present study
indicated that there were three genes, namely, RNF165, CBX5, and
CACUL1 targeted by the 2 miRNAs. Among these three genes, the
identification of ring finger protein RNF165 was in accordance with
previous studies, in which it was identified as a good candidate
for SCZ diagnosis. In the study by Pickard et al (76), five key genes, including TTMA,
TCF4, DLGAP1, ARKL1 and ARKL2, were confirmed as biomarkers for SCZ
diagnosis. Along with the development of SCZ gene diagnosis, more
biomarkers were gradually identified, including CBX5 (also termed
HP1), which serves as a platform for other repressive proteins,
such as DNA methyltransferases, and has been indicated to have a
role in the functioning of dopamine neurons and the construction of
memory as a member of heterochromatin in the brain. In addition,
recently identified CACUL1, also known as CAC1 and C10orf46, is a
novel gene of the cullin family that was reported to promote cell
proliferation in a gastric cancer cell line (77), serves as a potential antitumor
target in prostate cancer (78),
promotes the proliferation of human lung cancer (79) and has an important role in
protecting cells from Aβ toxicity and oxidative stress in the
hippocampus of patients with Alzheimer's disease (80). The Pharmacogenomics Knowledgebase
(PharmGKB) may provide candidate antipsychotic drug target genes
for research (81). A drug-gene
interaction study indicated that the rs6313 position of the
serotonin receptor gene HTR2A has a regulatory mechanism involving
risperidone and olanzapine in SCZ (82). Another study indicated that gene
variation of GABRB1 may be significantly associated with the
antipsychotic drug dose (83). A
strong connection has been reported between genetic polymorphisms
(in the HSPG2, CNR1, DPP6 and SLC18A2 genes) and antipsychotic
drugs, and even the side effects of these drugs (84). Another study identified 12 SNPs
from 143 genes of 79 Han Chinese patients with SCZ via HaloPlex
technology, with 2/12 SNPs determined to be significantly
associated with the response to olanzapine, using the MassARRAY
platform in two independent cohorts (85). Other studies confirmed that the
effectiveness of antipsychotic drugs is closely associated with the
variation of drug target genes in patients with TRS (86–89).
Although these three genes (RNF165, CBX5 and CACUL1) have not been
reported in the PharmGKB to interact with medicines, the above
information may be useful for future drug research.
In conclusion, in the present study, a simple, fast
and effective strategy, mainly based on the detection of the
expression of the top three miRNAs in peripheral blood MCs, was
applied. hsa-miR-218-5p and hsa-miR-1262 were proposed as two major
biomarkers for earlier and more precise diagnosis of TRS, shedding
new light on the development of both gene-based diagnosis and
treatment. However, the present study had certain limitations.
First, the sample size of the study was small. Furthermore,
limitations regarding the diagnostic accuracy may lead to
misdiagnosis of certain samples. For example, the AUC value in ROC
analysis was not very high, and there may be false positives and
false negatives. Another limitation of this study is that the
inclusion criteria were not designed to tell apart patients with
normal schizophrenia that may respond to treatment from those with
TRS. Finally, although the two target genes were confirmed in a
relatively large cohort, there was a lack of functional
verification, such as gene knockout experiments. In the future,
studies with larger samples and a multi-center design are required
to further verify the present results.
Supplementary Material
Supporting Data
Acknowledgements
Not applicable.
Funding
The current study was supported by The National
Natural Science Fund (grant no. 81760253).
Availability of data and materials
The datasets used and/or analyzed in this study are
available from the corresponding author on reasonable request.
Authors' contributions
YZ and ZWT designed and supervised the study. XY,
YQZ and QL were responsible for analyzing the data, writing the
manuscript and revising it, and designing the figures. ZJL, XM,
ZXL, WY, ZF and WYZ contributed to the statistical analysis of the
data and text correction. All authors read and approved the final
version.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Kahn RS, Sommer IE, Murray RM,
Meyer-Lindenberg A, Weinberger DR, Cannon TD, O'Donovan M, Correll
CU, Kane JM, van Os J and Insel TR: Schizophrenia. Nat Rev Dis
Primers. 1:150672015. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Gillespie AL, Samanaite R, Mill J, Egerton
A and MacCabe JH: Is treatment-resistant schizophrenia
categorically distinct from treatment-responsive schizophrenia? A
systematic review. BMC Psychiatry. 17:122017. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Lehman AF, Lieberman JA, Dixon LB,
McGlashan TH, Miller AL, Perkins DO and Kreyenbuhl J; American
Psychiatric Association; Steering Committee on Practice Guidelines,
: Practice guideline for the treatment of patients with
schizophrenia, second edition. Am J Psychiatry. 161:1–56.
2004.PubMed/NCBI
|
|
4
|
Samara MT, Dold M, Gianatsi M,
Nikolakopoulou A, Helfer B, Salanti G and Leucht S: Efficacy,
acceptability, and tolerability of antipsychotics in
treatment-resistant schizophrenia: A network meta-analysis. JAMA
Psychiatry. 73:199–210. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Lakhan SE and Kramer A: Schizophrenia
genomics and proteomics: Are we any closer to biomarker discovery?
Behav Brain Funct. 5:22009. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Schwarz E and Bahn S: Biomarker discovery
in psychiatric disorders. Electrophoresis. 29:2884–2890.
2008.PubMed/NCBI
|
|
7
|
Schwarz E and Bahn S: The utility of
biomarker discovery approaches for the detection of disease
mechanisms in psychiatric disorders. Br J Pharmacol. 153:S133–S136.
2008. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Qureshi IA and Mehler MF: Emerging roles
of non-coding RNAs in brain evolution, development, plasticity and
disease. Nat Rev Neurosci. 13:528–541. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Tsankova N, Renthal W, Kumar A and Nestler
EJ: Epigenetic regulation in psychiatric disorders. Nat Rev
Neurosci. 8:355–367. 2007. View
Article : Google Scholar : PubMed/NCBI
|
|
10
|
Sullivan PF, Fan C and Perou CM:
Evaluating the comparability of gene expression in blood and brain.
Am J Med Genet B Neuropsychiatr Genet. 141:261–268. 2006.
View Article : Google Scholar
|
|
11
|
Vawter MP, Ferran E, Galke B, Cooper K,
Bunney WE and Byerley W: Microarray screening of lymphocyte gene
expression differences in a multiplex schizophrenia pedigree.
Schizophr Res. 67:41–52. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Glatt SJ, Everall IP, Kremen WS, Corbeil
J, Sásik R, Khanlou N, Han M, Liew CC and Tsuang MT: Comparative
gene expression analysis of blood and brain provides concurrent
validation of SELENBP1 up-regulation in schizophrenia. Proc Natl
Acad Sci USA. 102:15533–15538. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Kuzman MR, Medved V, Terzic J and Krainc
D: Genome-wide expression analysis of peripheral blood identifies
candidate biomarkers for schizophrenia. J Psychiatr Res.
43:1073–1077. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Maffioletti E, Tardito D, Gennarelli M and
Bocchio-Chiavetto L: Micro spies from the brain to the periphery:
New clues from studies on microRNAs in neuropsychiatric disorders.
Front Cell Neurosci. 8:752014. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Yan Y, Chen X, Wang X, Zhao Z, Hu W, Zeng
S, Wei J, Yang X, Qian L, Zhou S, et al: The effects and the
mechanisms of autophagy on the cancer-associated fibroblasts in
cancer. J Exp Clin Cancer Res. 38:1712019. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Li H, Zhou DS, Chang H, Wang L, Liu W, Dai
SX, Zhang C, Cai J, Liu W, Li X, et al: Interactome Analyses
implicated CAMK2A in the genetic predisposition and pharmacological
mechanism of bipolar disorder. J Psychiatr Res. 115:165–175. 2019.
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Yan Y, Gong Z and Xu Z: Commentary: Lico A
causes ER stress and apoptosis via up-regulating miR-144-3p in
human lung cancer cell line H292. Biomed J. 41:391–392. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Ambros V: The functions of animal
microRNAs. Nature. 431:350–355. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Xu Z, Yan Y, Zeng S, Dai S, Chen X, Wei J
and Gong Z: Circular RNAs: Clinical relevance in cancer.
Oncotarget. 9:1444–1460. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Perkins DO, Jeffries C and Sullivan P:
Expanding the ‘central dogma’: The regulatory role of nonprotein
coding genes and implications for the genetic liability to
schizophrenia. Mol Psychiatry. 10:69–78. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Cheng HY, Papp JW, Varlamova O, Dziema H,
Russell B, Curfman JP, Nakazawa T, Shimizu K, Okamura H, Impey S
and Obrietan K: MicroRNA modulation of circadian-clock period and
entrainment. Neuron. 54:813–829. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Kocerha J, Faghihi MA, Lopez-Toledano MA,
Huang J, Ramsey AJ, Caron MG, Sales N, Willoughby D, Elmen J,
Hansen HF, et al: MicroRNA-219 modulates NMDA receptor-mediated
neurobehavioral dysfunction. Proc Natl Acad Sci USA. 106:3507–3512.
2009. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Coyle JT: MicroRNAs suggest a new
mechanism for altered brain gene expression in schizophrenia. Proc
Natl Acad Sci USA. 106:2975–2976. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Miller BH and Wahlestedt C: MicroRNA
dysregulation in psychiatric disease. Brain Res. 1338:89–99. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Gladkevich A, Kauffman HF and Korf J:
Lymphocytes as a neural probe: Potential for studying psychiatric
disorders. Prog Neuropsychopharmacol Biol Psychiatry. 28:559–576.
2004. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Marques-Deak A, Cizza G and Sternberg E:
Brain-Immune interactions and disease susceptibility. Mol
Psychiatry. 10:239–250. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Marazziti D, Catena Dell'osso M, Baroni S,
Masala I, Dell'Osso B, Consoli G, Giannaccini G, Betti L and
Lucacchini A: Alterations of the dopamine transporter in resting
lymphocytes of patients with different psychotic disorders.
Psychiatry Res. 175:54–57. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Lai CY, Yu SL, Hsieh MH, Chen CH, Chen HY,
Wen CC, Huang YH, Hsiao PC, Hsiao CK, Liu CM, et al: MicroRNA
expression aberration as potential peripheral blood biomarkers for
schizophrenia. PLoS One. 6:e216352011. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Sun Xy, Lu J, Zhang L, Song Ht, Zhao L,
Fan Hm, Zhong Af, Niu W, Guo Zm, Dai Yh, et al: Aberrant microRNA
expression in peripheral plasma and mononuclear cells as specific
blood-based biomarkers in schizophrenia patients. J Clin Neurosci.
22:570–574. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Wei H, Yuan Y, Liu S, Wang C, Yang F, Lu
Z, Wang C, Deng H, Zhao J, Shen Y, et al: Detection of circulating
miRNA levels in schizophrenia. Am J Psychiatry. 172:1141–1147.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Liu S, Zhang F, Shugart YY, Yang L, Li X,
Liu Z, Sun N, Yang C, Guo X, Shi J, et al: The early growth
response protein 1-miR-30a-5p-neurogenic differentiation factor 1
axis as a novel biomarker for schizophrenia diagnosis and treatment
monitoring. Transl Psychiatry. 7:e9982017. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Kalia M and Silva JC: Biomarkers of
psychiatric diseases: Current status and future prospects.
Metabolism. 64:S11–S15. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Tandon R, Gaebel W, Barch DM, Bustillo J,
Gur RE, Heckers S, Malaspina D, Owen MJ, Schultz S, Tsuang M, et
al: Definition and description of schizophrenia in the DSM-5.
Schizophr Res. 150:3–10. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Rao X, Huang X, Zhou Z and Lin X: An
improvement of the 2^(-delta delta CT) method for quantitative
real-time polymerase chain reaction data analysis. Biostat
Bioinforma Biomath. 3:71–85. 2013.PubMed/NCBI
|
|
35
|
Agarwal V, Bell GW, Nam JW and Bartel DP:
Predicting effective microRNA target sites in mammalian mRNAs.
ELife. 4:e050052015. View Article : Google Scholar
|
|
36
|
Paraskevopoulou MD, Georgakilas G,
Kostoulas N, Vlachos IS, Vergoulis T, Reczko M, Filippidis C,
Dalamagas T and Hatzigeorgiou AG: DIANA-microT web server v5.0:
Service integration into miRNA functional analysis workflows.
Nucleic Acids Res. 41:W169–W173. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Wang X, Su R, Guo Q, Liu J, Ruan B and
Wang G: Competing endogenous RNA (ceRNA) hypothetic model based on
comprehensive analysis of long non-coding RNA expression in lung
adenocarcinoma. Peer J. 7:e80242019. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Meltzer HY: Treatment-resistant
schizophrenia-the role of clozapine. Curr Med Res Opin. 14:1–20.
1997. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Wimberley T, Gasse C, Meier SM, Agerbo E,
MacCabe JH and Horsdal HT: Polygenic risk score for schizophrenia
and treatment-resistant schizophrenia. Schizophr Bull.
43:1064–1069. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Ashley EA: Towards precision medicine. Nat
Rev Genet. 17:507–522. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Tonelli MR and Shirts BH: Knowledge for
precision medicine: Mechanistic reasoning and methodological
pluralism. JAMA. 318:1649–1650. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Alda M: Personalized psychiatry: Many
questions, fewer answers. J Psychiatry Neurosci. 38:363–365. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Manchia M, Pisanu C, Squassina A and
Carpiniello B: Challenges and future prospects of precision
medicine in psychiatry. Pharmgenomics Pers Med. 13:127–140.
2020.PubMed/NCBI
|
|
44
|
Kochunov P, Hong LE, Dennis EL, Morey RA,
Tate DF, Wilde EA, Logue M, Kelly S, Donohoe G, Favre P, et al:
ENIGMA-DTI: Translating reproducible white matter deficits into
personalized vulnerability metrics in cross-diagnostic psychiatric
research. Human Brain Mapp. 16:249982020. View Article : Google Scholar
|
|
45
|
Thompson PM, Jahanshad N, Ching CR,
Salminen LE, Thomopoulos SI, Bright J, Baune BT, Bertolín S,
Bralten J, Bruin WB, et al: ENIGMA and global neuroscience: A
decade of large-scale studies of the brain in health and disease
across more than 40 countries. Transl Psychiatry. 10:1002020.
View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Holleran L, Kelly S, Alloza C, Agartz I,
Andreassen OA, Arango C, Banaj N, Calhoun V, Cannon D, Carr V, et
al: The relationship between white matter microstructure and
general cognitive ability in patients with schizophrenia and
healthy participants in the ENIGMA consortium. Am J Psychiatry.
177:537–547. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Boloc D, Rodríguez N, Torres T,
García-Cerro S, Parellada M, Saiz-Ruiz J, Cuesta MJ, Bernardo M,
Gassó P, Lafuente A, et al: Identifying key transcription factors
for pharmacogenetic studies of antipsychotics induced
extrapyramidal symptoms. Psychopharmacology (Berl). 237:2151–2159.
2020. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Heinla I, Leidmaa E, Kongi K, Pennert A,
Innos J, Nurk K, Tekko T, Singh K, Vanaveski T, Reimets R, et al:
Gene expression patterns and environmental enrichment-induced
effects in the hippocampi of mice suggest importance of lsamp in
plasticity. Front Neurosci. 9:2052015. View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Rees E and Owen MJ: Translating insights
from neuropsychiatric genetics and genomics for precision
psychiatry. Genome Med. 12:432020. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Birnbaum R and Weinberger DR: Special
article: Translational science update. Pharmacological implications
of emerging schizophrenia genetics: Can the bridge from ‘Genomics’
to ‘Therapeutics’ be defined and traversed? J Clin Psychopharmacol.
40:323–329. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Chakos M, Lieberman J, Hoffman E, Bradford
D and Sheitman B: Effectiveness of second-generation antipsychotics
in patients with treatment-resistant schizophrenia: A review and
meta-analysis of randomized trials. Am J Psychiatry. 158:518–526.
2001. View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Wahlbeck K, Cheine M, Essali A and Adams
C: Evidence of clozapine's effectiveness in schizophrenia: A
systematic review and meta-analysis of randomized trials. Am J
Psychiatry. 156:990–999. 1999.PubMed/NCBI
|
|
53
|
Meltzer HY: Treatment of the
neuroleptic-nonresponsive schizophrenic patient. Schizophr Bull.
18:515–542. 1992. View Article : Google Scholar : PubMed/NCBI
|
|
54
|
Siskind DJ, Lee M, Ravindran A, Zhang Q,
Ma E, Motamarri B and Kisely S: Augmentation strategies for
clozapine refractory schizophrenia: A systematic review and
meta-analysis. Aust N Z J Psychiatry. 52:751–767. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
55
|
van Westrhenen R, Aitchison KJ,
Ingelman-Sundberg M and Jukić MM: Pharmacogenomics of
antidepressant and antipsychotic treatment: How far have we got and
where are we going? Front Psychiatry. 11:942020. View Article : Google Scholar : PubMed/NCBI
|
|
56
|
Relling MV and Evans WE: Pharmacogenomics
in the clinic. Nature. 526:343–350. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
57
|
Zhang JP and Malhotra AK: Recent progress
in pharmacogenomics of antipsychotic drug response. Curr Psychiatry
Rep. 20:242018. View Article : Google Scholar : PubMed/NCBI
|
|
58
|
Feinberg AP and Fallin MD: Epigenetics at
the crossroads of genes and the environment. JAMA. 314:1129–1130.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
59
|
Nucifora FC, Woznica E, Lee BJ, Cascella N
and Sawa A: Treatment resistant schizophrenia: Clinical,
biological, and therapeutic perspectives. Neurobiol Dis.
131:1042572019. View Article : Google Scholar : PubMed/NCBI
|
|
60
|
Fisar Z and Raboch J: Depression,
antidepressants, and peripheral blood components. Neuro Endocrinol
Lett. 29:17–28. 2008.PubMed/NCBI
|
|
61
|
Cattaneo A, Sesta A, Calabrese F, Nielsen
G, Riva MA and Gennarelli M: The expression of VGF is reduced in
leukocytes of depressed patients and it is restored by effective
antidepressant treatment. Neuropsychopharmacology. 35:1423–1428.
2010. View Article : Google Scholar : PubMed/NCBI
|
|
62
|
Chen SD, Sun XY, Niu W, Kong LM, He MJ,
Fan HM, Li WS, Zhong AF, Zhang LY and Lu J: A preliminary analysis
of microRNA-21 expression alteration after antipsychotic treatment
in patients with schizophrenia. Psychiatry Res. 244:324–332. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
63
|
Liang Y, Ridzon D, Wong L and Chen C:
Characterization of microRNA expression profiles in normal human
tissues. BMC Genomics. 8:1662007. View Article : Google Scholar : PubMed/NCBI
|
|
64
|
Zhao Z, Jinde S, Koike S, Tada M, Satomura
Y, Yoshikawa A, Nishimura Y, Takizawa R, Kinoshita A, Sakakibara E,
et al: Altered expression of microRNA-223 in the plasma of patients
with first-episode schizophrenia and its possible relation to
neuronal migration-related genes. Transl Psychiatry. 9:2892019.
View Article : Google Scholar : PubMed/NCBI
|
|
65
|
Sun Xy, Zhang J, Niu W, Guo W, Song Ht, Li
Hy, Fan Hm, Zhao L, Zhong Af, Dai Yh, et al: A preliminary analysis
of microRNA as potential clinical biomarker for schizophrenia. Am J
Med Genet B Neuropsychiatr Genet. 168:170–178. 2015. View Article : Google Scholar
|
|
66
|
He K, Guo C, He L and Shi Y: miRNAs of
peripheral blood as the biomarker of schizophrenia. Hereditas.
155:92018. View Article : Google Scholar : PubMed/NCBI
|
|
67
|
Liu S, Zhang F, Wang X, Shugart YY, Zhao
Y, Li X, Liu Z, Sun N, Yang C, Zhang K, et al: Diagnostic value of
blood-derived microRNAs for schizophrenia: Results of a
meta-analysis and validation. Sci Rep. 7:153282017. View Article : Google Scholar : PubMed/NCBI
|
|
68
|
Inada T, Nakamura A and Iijima Y:
Relationship between catechol-O-methyltransferase polymorphism and
treatment-resistant schizophrenia. Am J Med Genet B Neuropsychiatr
Genet. 120:35–39. 2003. View Article : Google Scholar
|
|
69
|
Ji X, Takahashi N, Saito S, Ishihara R,
Maeno N, Inada T and Ozaki N: Relationship between three serotonin
receptor subtypes (HTR3A, HTR2A and HTR4) and treatment-resistant
schizophrenia in the Japanese population. Neurosci Lett. 435:95–98.
2008. View Article : Google Scholar : PubMed/NCBI
|
|
70
|
Kohlrausch FB, Gama CS, Lobato MI,
Belmonte-de-Abreu P, Callegari-Jacques SM, Gesteira A, Barros F,
Carracedo A and Hutz MH: Naturalistic pharmacogenetic study of
treatment resistance to typical neuroleptics in European-Brazilian
schizophrenics. Pharmacogenet Genomics. 18:599–609. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
71
|
Bilic P, Jukic V, Vilibic M, Savic A and
Bozina N: Treatment- Resistant schizophrenia and DAT and SERT
polymorphisms. Gene. 543:125–132. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
72
|
Terzić T, Kastelic M, Dolžan V and
Plesničar BK: Influence of 5-HT1A and 5-HTTLPR genetic variants on
the schizophrenia symptoms and occurrence of treatment-resistant
schizophrenia. Neuropsychiatr Dis Treat. 11:453–459.
2015.PubMed/NCBI
|
|
73
|
Catts VS, Fung SJ, Long LE, Joshi D,
Vercammen A, Allen KM, Fillman SG, Rothmond DA, Sinclair D, Tiwari
Y, et al: Rethinking schizophrenia in the context of normal
neurodevelopment. Front Cell Neurosci. 7:602013. View Article : Google Scholar : PubMed/NCBI
|
|
74
|
Alexander-Bloch AF, Reiss PT, Rapoport J,
McAdams H, Giedd JN, Bullmore ET and Gogtay N: Abnormal cortical
growth in schizophrenia targets normative modules of synchronized
development. Biol Psychiatry. 76:438–446. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
75
|
Horváth S and Mirnics K: Schizophrenia as
a disorder of molecular pathways. Biol Psychiatry. 77:22–28. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
76
|
Pickard BS, Malloy MP, Clark L, Lehellard
S, Ewald HL, Mors O, Porteous DJ, Blackwood DH and Muir WJ:
Candidate psychiatric illness genes identified in patients with
pericentric inversions of chromosome 18. Psychiatr Genet. 15:37–44.
2005. View Article : Google Scholar : PubMed/NCBI
|
|
77
|
Zheng Q, Zhao LY, Kong Y, Nan KJ, Yao Y
and Liao ZJ: CDK-Associated cullin 1 can promote cell proliferation
and inhibit cisplatin-induced apoptosis in the AGS gastric cancer
cell line. World J Surg Oncol. 11:52013. View Article : Google Scholar : PubMed/NCBI
|
|
78
|
Choi H, Lee SH, Um SJ and Kim EJ: CACUL1
functions as a negative regulator of androgen receptor in prostate
cancer cells. Cancer Lett. 376:360–366. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
79
|
Chen TJ, Gao F, Yang T, Thakur A, Ren H,
Li Y, Zhang S, Wang T and Chen MW: CDK-Associated cullin 1 promotes
cell proliferation with activation of ERK1/2 in human lung cancer
A549 cells. Biochem Biophys Res Commun. 437:108–113. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
80
|
Kong Y, Bai Ps, Sun H and Nan Kj:
Expression of the newly identified gene CAC1 in the hippocampus of
alzheimer's disease patients. J Mol Neurosci. 47:207–218. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
81
|
Drögemöller BI, Wright GE, Niehaus DJ,
Emsley R and Warnich L: Next-generation sequencing of
pharmacogenes: A critical analysis focusing on schizophrenia
treatment. Pharmacogenet Genomics. 23:666–674. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
82
|
Maffioletti E, Valsecchi P, Minelli A,
Magri C, Bonvicini C, Barlati S, Sacchetti E, Vita A and Gennarelli
M: Association study between HTR2A rs6313 polymorphism and early
response to risperidone and olanzapine in schizophrenia patients.
Drug Dev Res. 27:doi:10. 2020.
|
|
83
|
Hettige NC, de Morae GH, Kennedy JL and De
Luca V: Candidate gene analysis of pharmacodynamic targets for
antipsychotic dosage. Pharmacogenomics. 17:199–208. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
84
|
Zai CC, Tiwari AK, Zai GC, Maes MS and
Kennedy JL: New findings in pharmacogenetics of schizophrenia. Curr
Opin Psychiatry. 31:200–212. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
85
|
Zhou W, Xu Y, Lv Q, Sheng YH, Chen L, Li
M, Shen L, Huai C, Yi Z, Cui D and Qin S: Genetic association of
olanzapine treatment response in Han Chinese schizophrenia
patients. Front Pharmacol. 10:1772019. View Article : Google Scholar : PubMed/NCBI
|
|
86
|
Ruderfer DM, Charney AW, Readhead B, Kidd
BA, Kähler AK, Kenny PJ, Keiser MJ, Moran JL, Hultman CM, Scott SA,
et al: Polygenic overlap between schizophrenia risk and
antipsychotic response: A genomic medicine approach. Lancet
Psychiatry. 3:350–357. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
87
|
Gressier F, Porcelli S, Calati R and
Serretti A: Pharmacogenetics of clozapine response and induced
weight gain: A comprehensive review and meta-analysis. Eur
Neuropsychopharmacol. 26:163–185. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
88
|
Zhang JP and Malhotra AK: Pharmacogenetics
and antipsychotics: Therapeutic efficacy and side effects
prediction. Exp Opin Drug Metab Toxicol. 7:9–37. 2011. View Article : Google Scholar
|
|
89
|
Sriretnakumar V, Huang E and Müller DJ:
Pharmacogenetics of clozapine treatment response and side-effects
in schizophrenia: An update. Exp Opin Drug Metab Toxicol.
11:1709–1731. 2015. View Article : Google Scholar
|