Introduction
Congenital heart defects (CHDs), are the most common
type of birth defect, affecting ~ eight in 1,000 livebirths
(1,2). Among these congenital conditions,
tetralogy of Fallot (TOF) is the most common cyanotic heart
malformation and accounts for approximately 10% of all CHD cases
(3). TOF results in anatomic
defects, such as obstruction of the right ventricular outflow tract
(RVOT), ventricular septal defect, aortic dextroposition and right
ventricular hypertrophy (4). The
pathogenesis of TOF is not completely understood. Previous studies
have demonstrated that cardiac tissue-specific transcription
factors, genes and associated signalling pathways, such as the
Notch signalling pathway, play important roles in normal cardiac
development, and that abnormal expression of such genes can
contribute to TOF (5–7). In addition to DNA sequence variants,
environmental factors, such as opioid exposure during early
pregnancy and maternal pre-pregnancy obesity (8), may also be associated with the
aetiology of TOF, as indicated by epidemiological data (9).
Notch is a highly conserved signalling pathway that
regulates cell specification, differentiation, and organ formation
and morphogenesis involved in development (10). Moreover, tissue-specific
endocardial Notch signalling regulates cardiac morphogenesis
through interactions with multiple myocardial, epicardial and
neural crest-derived signals (11). Mutations in Notch signalling
elements result in CHD in humans and mice, demonstrating its
important role in normal cardiac development (12–14).
NOTCH4 serves as a membrane-bound receptor that regulates cell fate
(15). Notch1-deficient embryos
display severe vascular developmental defects, which are
exacerbated in Notch1/Notch4 double-mutant embryos (16). Constitutive activation of
Notch4 in the embryonic vasculature also leads to defects in
vascular remodelling (17,18). In addition, a previous study have
demonstrated that Notch4 activation in endothelial cells causes
trans-differentiation to a mesenchymal phenotype, suggesting that
implicates Jagged1-Notch interactions promote
epithelial-mesenchymal transition, which is required for normal
endocardial cushion differentiation and vascular smooth muscle cell
development (19). Furthermore, a
previous study has demonstrated that the NOTCH4 gene was
downregulated and had low frequency of genetic variants in the
NOTCH4 coding region in patients with TOF (20).
DNA methylation is an epigenetic modification that
is vital for embryonic development, and changes in methylation may
be associated with the development of cardiovascular diseases
(21). The interactions between
DNA and transcription factors could be influenced directly or
indirectly by DNA methylation, due to recruitment of
methyl-CpG-binding proteins (22).
Sheng et al (23,24) suggested that aberrant methylation
levels at the promoter CpG island shore of the ZFPM2 and
HAND1 genes may be responsible for gene transcription
regulation in patients with TOF. Gong et al (25) demonstrated that demethylation in
the TBX20 promoter region may be associated with
overexpression in the cardiac tissue of patients with TOF. However,
a recent study on severe anxiety has reported that a single CpG
site located in the promoter of the Asb1 gene may be
responsible for a methylation increase of 48.5% (26). Methylation at specific loci in
genes, such as tissue factor F3, interleukin-6 and toll-like
receptor 2, is influenced by exposure to air pollution, thus
leading to several adverse health effects (27–29).
Therefore it may be hypothesised that changes in DNA methylation
contribute to downregulation of NOTCH4 in patients with TOF.
NOTCH4 has been demonstrated to be a ETS
factor- regulated gene, and ETS may activate the expression of
Notch signalling components to initiate Notch signalling in the
early artery (30). The ETS1
oncogene belongs to a large family within the ETS domain family of
transcription factors (31). It is
widely expressed in the developing embryo, and could be detectable
in the day-15 embryos of murine (32). Previous studies have demonstrated
that ETS1 plays an important role in human heart development and
was associated with the development of CHD (33,34).
ETS proteins can function as transcriptional activators by binding
to specific DNA sequences of target genes, thereby increasing (such
as DLL4, NOTCH4 and NKp46) or decreasing (such as
MMP1 and BCL2) gene transcription in response to
various stimuli, including changes of genetics, histone
modification or DNA methylation (30,35–37).
The aim of the present study was to examine the
epigenetic mechanisms that regulate the NOTCH4 gene and
their effect on NOTCH4 protein expression in patients with TOF. The
findings of this study may provide insight into the aetiology of
TOF.
Materials and methods
Clinical samples
The present study was approved by The Ethics
Committee of The Children's Hospital of Fudan University (approval
no. 2015) (26). Written informed
consent was obtained from the parents or relatives of all study
participants. Patients with TOF were recruited from the Children's
Hospital of Fudan University between January 2016 to July 2018. TOF
was diagnosed using an echocardiogram and confirmed by surgery. The
control samples from autopsy specimens were provided by the
Department of Forensic Medicine of Soochow University. In total, 24
patients with TOF were enrolled, including 14 (58.3%) males and 10
(41.7%) females. Patient age ranged from 1 month to14 years (mean ±
SD, 2.54 ± 0.86 years). Control samples were obtained from five
male subjects, aged 1 day to 7 months (mean ± SD, 0.35 ± 0.19
years). All samples were taken from RVOT myocardial tissue and
immediately stored at −80°C in RNAlater solution (Ambion; Thermo
Fisher Scientific, Inc.). Patient characteristics are summarized in
Table I.
 | Table I.Clinical characteristics of patients
with TOF and healthy controls. |
Table I.
Clinical characteristics of patients
with TOF and healthy controls.
| Clinical
variable | Patients with
TOF | Controls |
P-valuea |
|---|
| Age, years, mean ±
SD | 2.54±0.86 | 0.35±0.19 |
|
| Age category,
years, n (%) |
|
|
|
|
<1 | 15 (62.5) | 4 (80.0) | 0.796a |
|
1-2 | 4
(16.7) | 1 (20.0) |
|
|
>2 | 5
(20.8) | 0 (0) |
|
| Sex, n (%) |
|
|
|
|
Male | 14 (58.3) | 5 (100) | 0.134a |
|
Female | 10 (41.7) | 0 (0) |
|
Immunohistochemistry
RVOT myocardial tissues were fixed in 10% neutral
buffered formalin at room temperature for 48 h and embedded in
paraffin. The sections were then cut into 4-µm sections and dried
overnight at 56°C. After deparaffination, hydration and antigen
retrieval with citric acid buffer (0.01 mol/l; pH 6.0), endogenous
peroxidase activity was blocked by incubating the sections with 3%
hydrogen peroxide (H2O2) at room temperature
for 20 min and blocking with 5% bovine serum (cat. no. 143183; Bio
Forxx) for 1 h at room temperature. The slides were incubated with
a rabbit anti-human monoclonal antibody against NOTCH4 (dilution
1:200, cat. no. ab184742; Abcam) overnight at 4°C. After primary
antibody incubation, the slides were incubated with goat
anti-rabbit/mouse IgG (cat. no. GK500710; Gene Tech Co., Ltd.) for
30 min at room temperature, then washed in TBST three times for 5
min each time. Finally, the slides were stained with
3,3-diaminobenzidine (DAB) for 40 sec and counterstained with
haematoxylin for 1 min. After dehydration, the sections were
mounted with mounting medium. For each sample, three visual fields
were randomly chosen and examined under a light microscope at ×200
magnification. Images of cardiomyocytes were quantified using
ImageJ (version 1.48; National Institutes of Health). To quantify
the intensity of NOTCH4 protein expression, images were captured
with a light microscope. Three randomly selected fields (×200
magnification) per tissue section were scanned and analyzed using
ImageJ software (version 1.48). The integrated optical density
(IOD) sum of each image was measured after the optical density was
adjusted with the segmentation set at a level to allow for
detection of positive immunostaining. For statistical analysis, the
mean value of the total three counted fields was calculated.
DNA extraction and bisulphite
sequencing PCR (BSP)
Genomic DNA was extracted from RVOT myocardial
tissue samples from patients with TOF and control subjects using a
QIAamp DNA Mini kit (Qiagen GmbH) according to the manufacturer's
instructions. For each sample, bisulphite treatment of genomic DNA
from RVOT myocardial tissues was performed using the EZ DNA
Methylation-Gold kit (Zymo Research Corp.) according to the
manufacturer's protocol. The bisulphite-treated DNA samples were
then used as templates for BSP. The primers used to amplify the
NOTCH4_R region (from position −240 to +113 bp, containing
five CpG sites) were designed using Methyl Primer Express v1.0
software (Applied Biosystem; Thermo Fisher Scientific, Inc.).
Primer sequences were as follows: NOTCH4-BSP-F,
5′-AATAGTAGGGTTGGGATTGTTTAGG-3′ and NOTCH4-BSP-R,
5′-ACAAAAACCACCTCCTCTACTCC-3′. For sequencing, the PCR products
were purified using the AxyPrep DNA Gel Extraction kit (Axygen;
Corning, Inc.) according to the manufacturer's protocol. The
purified PCR products were then cloned into the pGEM-T easy vector
system (Promega Corporation) and transformed into DH5α competent
cells (Tiangen Biotech Co., Ltd.). After a 12-h incubation at 37°C,
blue/white and ampicillin screening was carried out. For each
p-GEM-T vector, 10 clones were determined the methylation status
using Sanger sequencing The BIQ Analyser software (https://biq-analyzer.bioinf.mpi-inf.mpg.de/index.php)
was used to analyse the sequencing results. The percent methylation
of each CpG site in the samples was calculated as the number of
methylated CpG sites relative to the total number of observed
sequenced clones, which represents the overall methylation status
of a specific region in a screened sample.
Reporter plasmid construction and in
vitro methylation
pGL3-basic lacks eukaryotic promoter and enhancer
sequences, and serves as a negative control. The region [position
−179 to −53 bp, relative to the transcription start site (TSS)] of
the human NOTCH4 gene was amplified with Premix PrimeSTAR HS
DNA polymerase (cat. no. R040A; Takara Bio, Inc.), using human
genomic DNA isolated from tissues of normal controls as a template.
The PCR primers were synthesized by Generay Biotech Co., Ltd., as
follows: NOTCH4-KpnI-F, 5′-ATAGGTACCAGATTCCTTCTCCCCTCCTA-3′
and NOTCH4-XhoI-R, 5′-GATCTCGAGGAGGAAGAGTGGAGGAACAC-3′. The
amplified products were digested with KpnI–XhoI (cat. no. R3142V
and R0146S; New England BioLabs, Inc.) and then inserted into the
pGL3-promoter vector (Promega Corporation). The products were
transformed into DH5α-competent cells (Tiangen Biotech Co., Ltd.),
smeared on a lysogeny broth (LB) ampicillin (AMP) plate and
cultured at 37°C overnight. An LB culture containing AMP was used
to screen for the objective colonies. The fidelity of the plasmids
was verified by Sanger sequencing. This construct is referred to as
pGL3-NOTCH4_-179/-53 plasmid thereafter.
In addition, The Mut-pGL3-NOTCH4_-179/-53
plasmid was constructed by mutating a C base in the CpG site 2 to a
T base to simulate its hypermethylation status which might abrogate
binding of the target to NOTCH4_R. The
Mut-pGL3-NOTCH4_-179/-53 plasmid was constructed using the
KOD-plus-mutagenesis kit (cat. no. SMK-101; Toyobo Life Science)
according to the manufacturer's protocol. The primers used for
mutagenesis were synthesized by Generay Biotech Co., Ltd., as
follows: Mut-NOTCH4-F, 5′-TGTCCTACTTCCCCCTACTTCCCCA-3′ and
Mut-NOTCH4-R, 5′-GTGTGCCTGGAGGGCAGGTGATAGG-3′.
Lastly, the Me-pGL3-NOTCH4_-179/-53 plasmid
was generated by incubating the pGL3-NOTCH4_-179/-53 plasmid
with the M.SssI CpG methyltransferase (cat. no. M0226V; New England
BioLabs) for 4 h at 37°C. The methylation status was confirmed by
BSP, as aforementioned.
Transfections and dual luciferase
reporter assays
HeLa and HL-1 cells were purchased from the Cell
Bank of the Type Culture Collection of the Chinese Academy of
Sciences, which were cultured in Dulbecco's modified Eagle's medium
(Gibco; Thermo Fisher Scientific, Inc.) supplemented with 10%
foetal bovine serum (Gibco; Thermo Fisher Scientific, Inc.) at 37°C
with 5% CO2. For transfection, the cells were plated in
96-well plates 12 h before transfection at a density of
1–4×105 cells/well, then separately transfected with 100
ng of pGL3-basic (negative control), pGL3-promoter (empty vector),
pGL3-NOTCH4_-179/-53 or Me-pGL3-NOTCH4_-179/-53
vector using Lipofectamine® 3000 (Invitrogen; Thermo
Fisher Scientific, Inc.).
In order to study the mechanism underlying the
effect of the NOTCH4_R region on gene transcription
activity, the NOTCH4_R region sequence was analysed by a TF
search (http://www.cbrc.jp/research/db/TFSEARCH.html) and the
JASPAR database (http://jaspar.binf.ku.dk/cgi-bin/jaspar_db.pl?-rm=browse&db=core&tax_group=vertebrates),
and the results demonstrated the potential ETS1 transcription
factor binding sites, and the CpG site 2 was within the ETS1
binding site. For dual luciferase reporter assays, 100 ng of
pGL3-NOTCH4_-179/-53 (unmethylated),
Mut-pGL3-NOTCH4_-179/-53 (mutated) and
Me-pGL3-NOTCH4_-179/-53 (methylated) were co-transfected
with 100 ng of ETS1 transcription factor expression vector which
was constructed by cloning the entire human ETS1 cDNA (accession
no. NM_001143820) into a pcDNA3.1 (+) expression vector (cat. no.
G105592; YouBio). The pGL3-basic and pGL3-promoter vectors were
also used as controls. The pRL-TK plasmid (Promega Corporation) was
co-transfected with these plasmids to normalize luciferase activity
48 h following transient transfection. Three independent
experiments were performed.
Electrophoretic mobility shift assay
(EMSA) and shift western blotting
293T cells originally purchased from the Cell Bank
of the Type Culture Collection of the Chinese Academy of Sciences.
The cells were plated in 6-well plates 12 h before transfection at
a density of 1–2×106 cells/well. The cells were
transfected with 2.5 µg pcDNA3.1-ETS1 expression plasmid, and the
nuclear extracts were obtained using cytoplasmic extraction reagent
and nuclear extraction reagent (Thermo Fisher Scientific, Inc.)
according to the manufacturer's protocol. The protein concentration
was measured using a bicinchoninic acid protein assay kit (Takara
Biotechnology Co., Ltd.) according to the manufacturer's protocol.
Subsequently, ETS1 overexpression was confirmed by western
blotting. Protein samples (~10 µg) were separated by 10% SDS-PAGE
gels, transferred to a nitrocellulose membrane (Whatman plc; GE
Healthcare Life Sciences), and blocked with phosphate-buffered
saline (PBS) with 1% Tween-20 (PBST) containing 5% BSA for 2 h at
room temperature. Next, the membrane was probed with primary
antibodies against ETS1 (1:1,000; cat. no. ab220361; Abcam) and
GAPDH (1:3,000; cat. no. ab9482; Abcam) at 4°C overnight, then
incubated with horseradish peroxidase (HRP)-conjugated anti-rabbit
secondary antibody (1:3,000; cat. no. M21002; Abmart) and
HRP-conjugated anti-mouse secondary antibody (1:3,000; cat. no.
M21001; Abmart). The blots were detected by enhanced
chemiluminescence (Thermo Fisher Scientific, Inc.), and were
visualized using GE ImageQuant LAS4000 mini (GE Healthcare Life
Sciences). GAPDH was used as a loading control.
Biotin-labelled oligonucleotide probes
(Biotin-probe) specific for the ETS1-binding site of the
NOTCH4 gene (5′-CGTCCT-3′; position-138 to −133) were
synthesized by Generay Biotech Co., Ltd. and the oligonucleotide
probes was annealed into double strands by heating it to 95°C, then
cooling to room temperature. The mutant biotin-labelled
oligonucleotide probes (Biotin-Mut-probe) were used to confirm the
ETS1 binding specificity. An unlabelled oligonucleotide probe
(Competitor-WT), a mutant unlabelled oligonucleotide probe
(Competitor-Mut) and an methylated oligonucleotide probe
(Competitor-Met) were used as competitors. The sequences of these
oligonucleotide probes are listed in Table II.
| Table II.Sequences of oligonucleotide probes
used for electrophoretic mobility shift assay. |
Table II.
Sequences of oligonucleotide probes
used for electrophoretic mobility shift assay.
| Probe | Sequence
(5′-3′) |
|---|
| Biotin-probe-F |
GCCCTCCAGGCACACCGTCCTACTTCCC |
| Biotin-probe-R |
GGGAAGTAGGACGGTGTGCCTGGAGGGC |
|
Biotin-Mut-probe-F |
GCCCTCCAGGCACACATAACACTAGCCC |
|
Biotin-Mut-probe-R |
GGGCTAGTGTTATGTGTGCCTGGAGGGC |
|
Competitor-WT-F |
GCCCTCCAGGCACACCGTCCTACTTCCC |
|
Competitor-WT-R |
GGGAAGTAGGACGGTGTGCCTGGAGGGC |
|
Competitor-Mut-F |
GCCCTCCAGGCACACATAACACTAGCCC |
|
Competitor-Mut-R |
GGGCTAGTGTTATGTGTGCCTGGAGGGC |
|
Competitor-Met-F |
GCCCTCCAGGCACACCGTCCTACTTCCC |
|
Competitor-Met-R |
GGGAAGTAGGACGGTGTGCCTGGAGGGC |
The DNA-binding ability of ETS1 to the NOTCH4
gene was detected by EMSA using a LightShift™ EMSA kit (cat. no.
20148; Thermo Fisher Scientific, Inc.) according to the
manufacturer's instructions. Specifically, 10 µg nuclear extract
were incubated with 20 fmol biotin-labelled probes in binding
buffer at room temperature for 30 min, and a 200-fold excess of
unlabelled/methylated probes was added to the reaction as
competitors. The protein-DNA complexes were separated from the free
probes in a 6% polyacrylamide gel at 100 V for 50 min, then
transferred onto a nylon membrane at 380 mA for 30 min.
Subsequently, the membrane was analysed using a Fujifilm Las3000
Luminescent Image Analyzer (FUJIFILM Wako Pure Chemical
Corporation). Shift-western blotting was performed by transferring
the protein-DNA complexes from the polyacrylamide gel to a PVDF
membrane in 0.5X Tris-borate-EDTA (TBE) buffer for 30 min. For
protein detection, the membrane was blocked with blocking buffer
(PBS with 1% Tween-20 and 5% BSA) for 2 h at room temperature, then
incubated with a primary antibody against ETS1 (dilution 1:1,000;
cat. no. ab220361; Abcam) at 4°C overnight. Following primary
antibody incubation, the membrane was incubated with a
HRP-conjugated anti-rabbit secondary antibody (dilution 1:3,000;
cat. no. M21002; Abmart) at room temperature for 2 h. The blots
were visualized using a enhanced chemiluminescence (Thermo Fisher
Scientific, Inc.) according to the manufacturer's protocol, and
were visualized using GE ImageQuant LAS4000 mini (GE Healthcare
Life Sciences).
Statistical analysis
All data are presented as the mean ± SD of three
independent experiments. Statistical analysis was performed using
SPSS software v20.0 (IBM Corp.). Differences between two groups
were analysed by the Mann-Whitney test. Differences of luciferase
activity assays between multiple groups were analysed using one-way
ANOVA, followed by the Least Significant Difference post hoc test.
Pearson's correlation analysis was performed to analyse the
relationship between the immunohistochemistry data and bisulphite
sequencing data. P<0.05 was considered to indicate a
statistically significant difference.
Results
Expression of the NOTCH4 protein in
patients with TOF and controls
Immunohistochemistry was carried out to detect
NOTCH4 protein expression in RVOT myocardial tissue from 24
patients with TOF and five controls. NOTCH4 protein was detectable
by yellow or brown staining in the nucleus or cytoplasm. The
staining intensity of cardiomyocytes from patients with TOF was
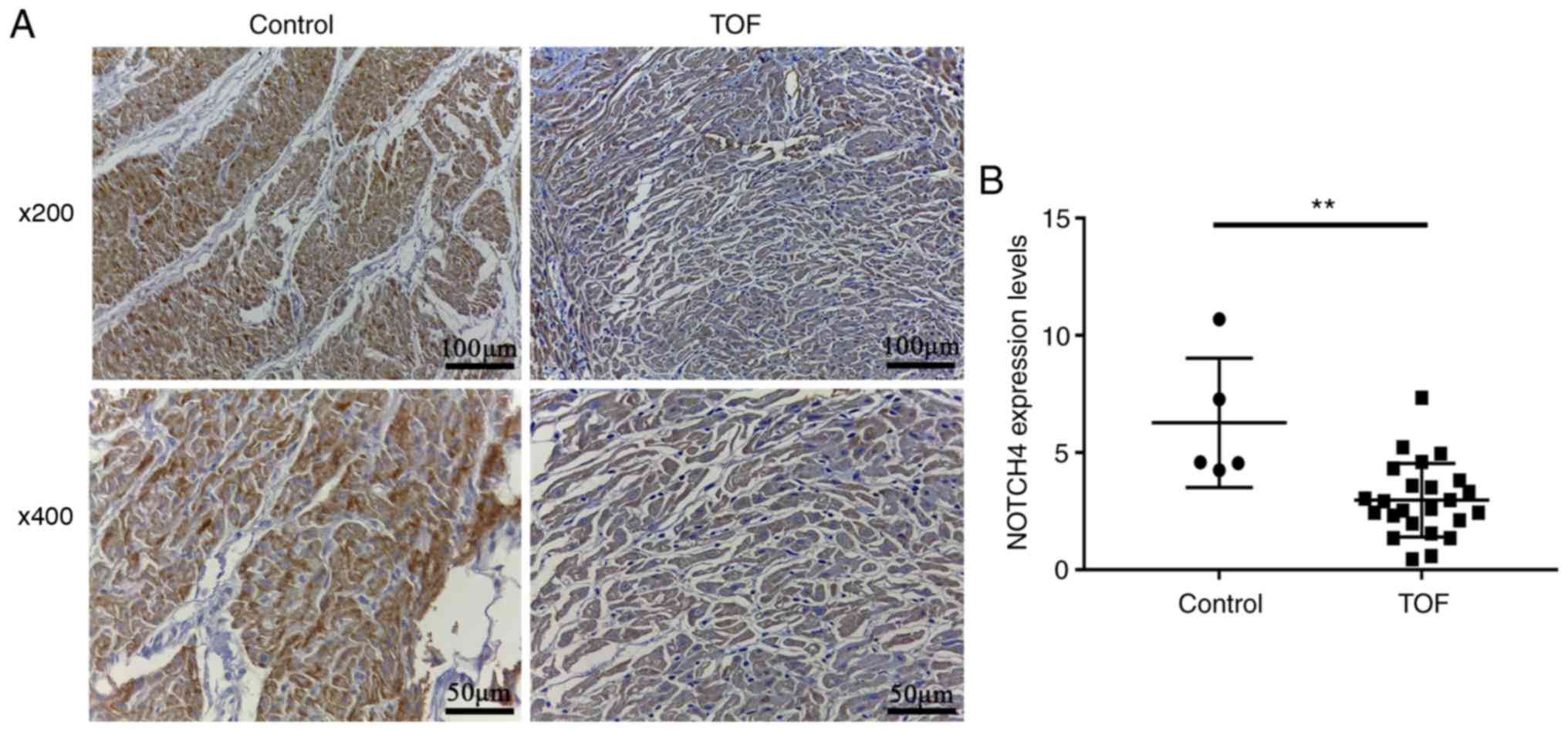
weaker than that in the control subjects (Fig. 1A). Statistical analysis confirmed
that NOTCH4 expression was significantly lower in the patients,
compared with the controls (3.0 ± 0.3 vs. 6.3 ± 1.2; P=0.0055;
Fig. 1B).
NOTCH4 promoter methylation status is
associated with protein expression
BSP was carried out in order to determine whether
reduced NOTCH4 expression was caused by changes in its epigenetic
regulation, BSP was performed on the NOTCH4 promoter using
tissue samples collected from patients with TOF and controls.
Considering that there is no CpG island in the promoter region
(−2,000 to 200 bp), only the region that was targeted for
sequencing (NOTCH4_R: −240 to +113 bp, containing 5 CpG
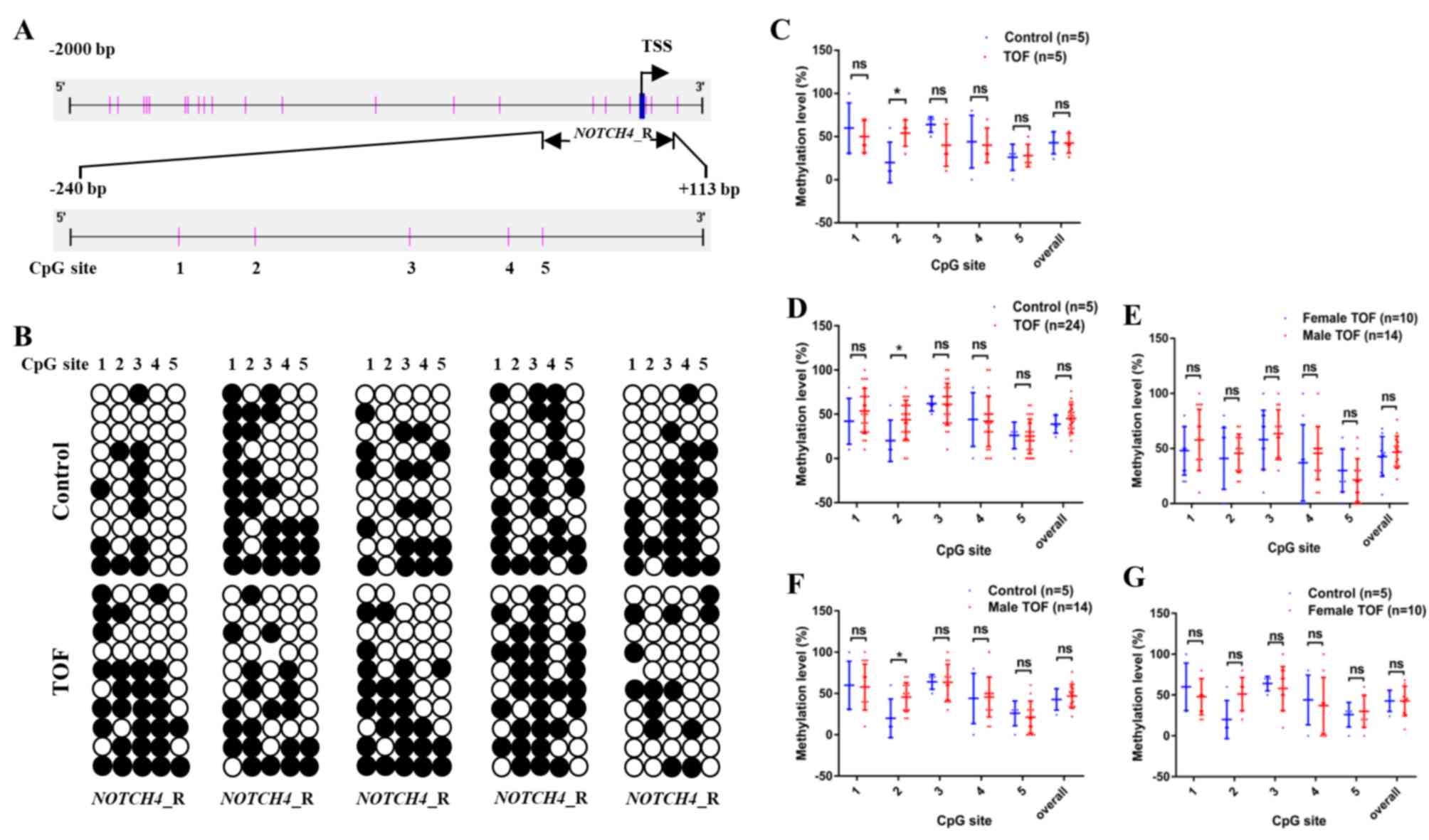
sites; Fig. 2A) was studied.
In an initial screen, the methylation levels in
NOTCH4_R were analysed in five patients with TOF and five
control subjects (Fig. 2B). The
overall methylation levels of NOTCH4_R (CpG sites 1–5) in
patients with TOF did not significantly differ from the controls
(42.4 ± 5.1 vs. 42.8 ± 5.8; Fig.
2C). Interestingly, only one CpG site (CpG site 2) in
NOTCH4_R exhibited significantly higher methylation levels,
compared with the controls (54.0 ± 6.8 vs. 20.0 ± 10.5;
P=0.0476).
Following this initial screen, the methylation
status of NOTCH4_R was determined in another 19 TOF patients
and the data for the combined cohort was analysed (n=24; Fig. 2D). The methylation levels of CpG
site 2 were significantly higher in 24 patients with TOF, compared
with the controls (43.8 ± 4.5 vs. 20.0 ± 10.5; P=0.0459; Fig. 2D).
In addition, the methylation levels of all five CpG
sites, as well as overall NOTCH4_R promoter methylation,
were compared between male (n=14) and female (n=10) patients with
TOF. None of the five CpG sites significantly differed between male
and female patients with respect to methylation levels (Fig. 2E). Furthermore, the methylation
levels of the five CpG sites were also compared between male
patients with and normal controls. Consistent with the
aforementioned results, only CpG site 2 in NOTCH4_R
displayed significantly higher methylation levels in male patients
with TOF, compared with controls (45.7 ± 4.7 vs. 20.0 ± 10.5;
P=0.0263; Fig. 2F). In addition,
the methylation levels of the five CpG sites were also compared
between female patients with and normal controls. No significant
difference was observed between female and normal controls (male)
with respect to methylation levels (45.0 ± 9.2 vs. 20.0 ± 10.5;
P=0.1758; Fig. 2G). After
excluding two extreme values, CpG site 2 in NOTCH4_R showed
higher methylation levels in female patients with TOF, compared
with controls (56.2 ± 6.8 vs. 20.0 ± 10.5; P=0.0111). Thus, it can
be concluded that no sex differences exist in methylation levels in
the TOF group and the normal control group.
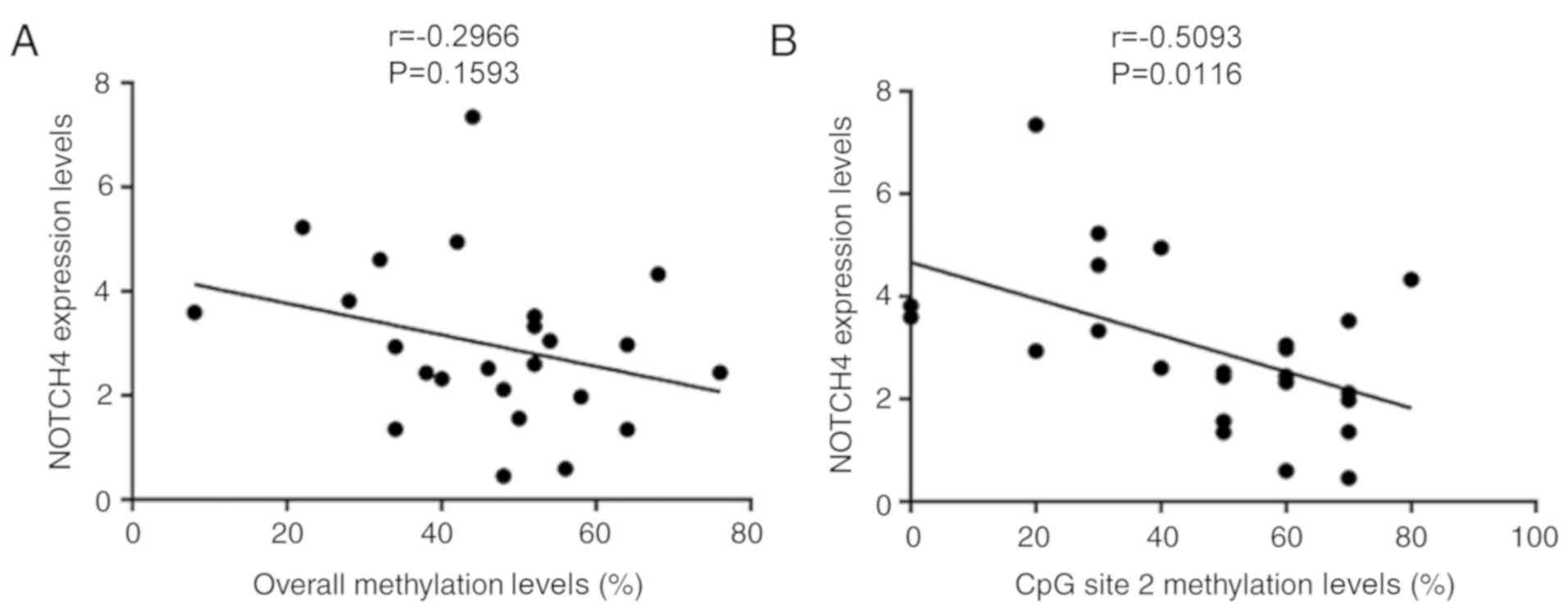
Pearson's correlation was carried out to determine
whether NOTCH4_R methylation status was associated with
NOTCH4 protein expression. There was no correlation between NOTCH4
protein expression and the overall methylation levels of
NOTCH4_R (r=−0.2966; P=0.1593; n=24; Fig. 3A) in patients with TOF. However, a
significant negative association was observed between NOTCH4
expression and the methylation status of the CpG site 2 (r=−0.5063;
P=0.0116; n=24; Fig. 3B) in
patients with TOF.
Effect of NOTCH4 CpG site 2
methylation on gene transcription in vitro
A dual-luciferase assay was carried out in HeLa and
HL-1 cells to determine whether single CpG site 2 methylation could
influence NOTCH4 gene transcription activity. The
pGL3-NOTCH4_-179/-53 plasmid was constructed by cloning the
NOTCH4 promoter region (−179 to −53 bp, containing only the
CpG site 2) into the pGL3-promoter vector. In addition, a
methylated plasmid was generated using M.SssI treatment. BSP was
used to determine the methylation status of the unmethylated and
methylated plasmids (Fig. 4A, B).
As indicated in Fig. 4C, the
luciferase activity of pGL3-NOTCH4_-179/-53 was
significantly higher than that of pGL3-promoter (HeLa: P=0.0022;
HL-1: P=0.0022). Moreover, the luciferase activity of
me-pGL3-NOTCH4_-179/-53 decreased nearly six-fold, compared
with pGL3-NOTCH4_-179/-53. These results indicated that
methylation in the NOTCH4_R region regulated gene
transcription and that a methylation of a single CpG site could
inhibit transcription of the NOTCH4 gene.
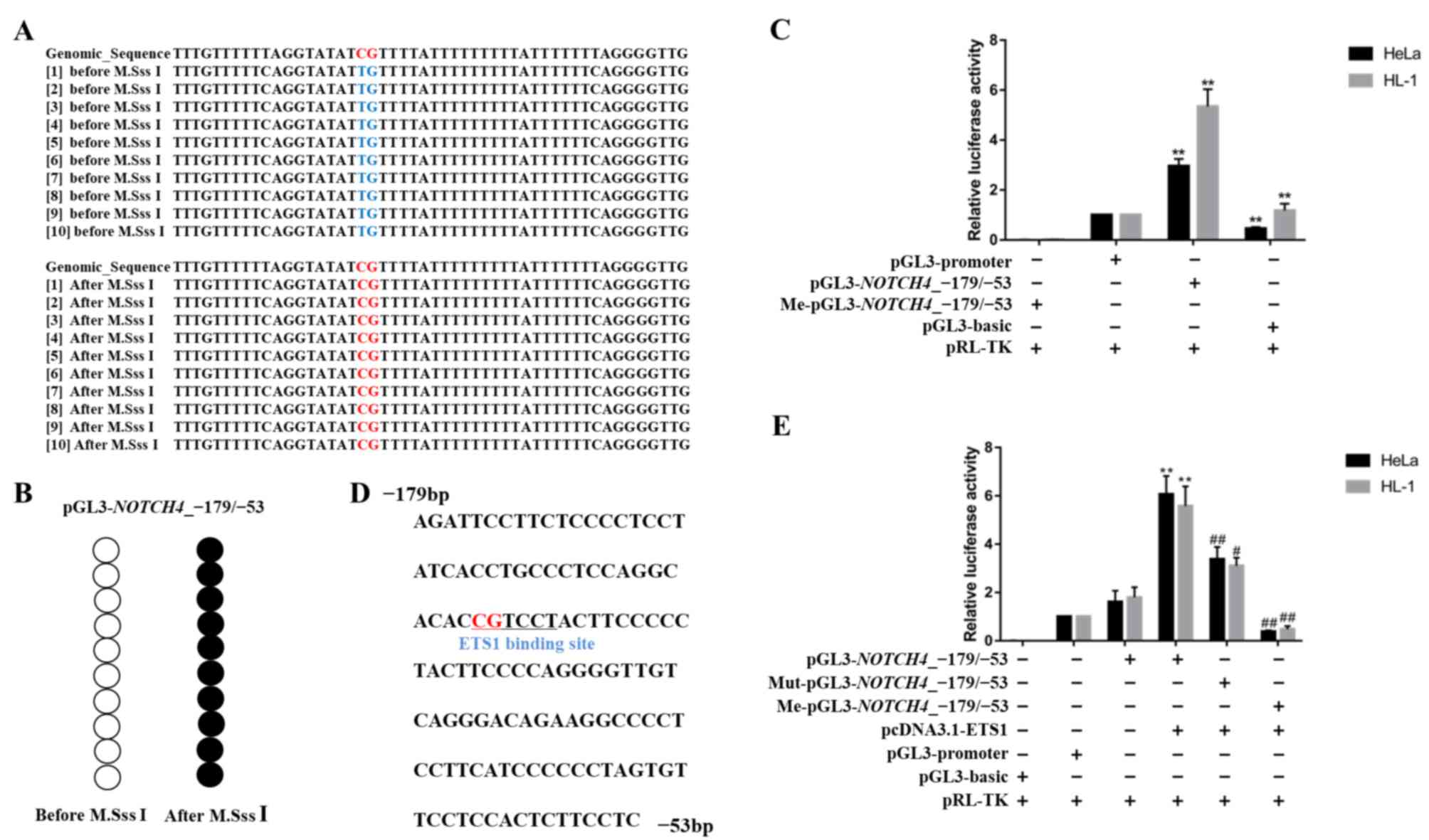 | Figure 4.Effect of NOTCH4_R CpG site 2
methylation on gene transcription activity. (A and B) Bisulphite
sequencing results for pGL3-NOTCH4_-179/-53. After M.SssI
treatment, all CpG sites were methylated. (C) Luciferase assay for
pGL3-basic, pGL3-promoter, pGL3-NOTCH4_-179/-53 and
Me-pGL3-NOTCH4_-179/-53 in HeLa and HL-1 cell lines. (D)
Sequence of the ETS1 transcription factor binding sites predicted
in the NOTCH4 promoter (−179 to −53 bp, relative to TSS).
The underlined sequence represents the ETS1 binding sites,
containing only CpG site 2. (E) Luciferase assay for pGL3-basic,
pGL3-promoter, pGL3-NOTCH4_-179/-53,
Me-pGL3-NOTCH4_-179/-53 and Mut-pGL3-NOTCH4_-179/-53
co-transfected with pcDNA3.1-ETS1 in HeLa and HL-1 cell lines. Data
are presented as the mean ± SD. **P<0.01 vs. pGL3-promoter;
#P<0.05 and ##P<0.01 vs. pGL3
NOTCH4_-179/-53. One-way ANOVA and Least Significant
Difference test. Me, methylated, Mut, mutant; TSS, transcription
start site. |
NOTCH4 expression is regulated by the
ETS1 transcription factor binding to its promoter region
To clarify the mechanism through which
NOTCH4_R methylation affects gene transcription, the
NOTCH4_R sequence was analysed for potential binding sites
for the ETS1 transcription factor using an online TF search and the
JASPAR database (38,39) (Fig.
4D). To evaluate the effect of ETS1 on NOTCH4
transcriptional activity, we co-transfected ETS1-overexpressing
plasmids with pGL3-NOTCH4_-179/-53,
Mut-pGL3-NOTCH4_-179/-53 and Me-pGL3-NOTCH4_-179-/53,
respectively, into HeLa and HL-1 cells (Fig. 4E). Following co-transfection with
pGL3-NOTCH4_-179/-53, luciferase activity was significantly
increased, compared with pGL3-basic. In addition, a significant
increase in luciferase activity following co-transfection with
pGL3-NOTCH4_-179/-53 and ETS1 transcription factor was
observed. However, the luciferase gene activity driven by
Mut-pGL3-NOTCH4_-179/-53 was significantly reduced in the
presence of ETS1, compared with pGL3-NOTCH4_-179/-53 (HeLa:
P=0.0022; HL-1: P=0.0159). Moreover, me-pGL3-NOTCH4_-179/-53
resulted in significantly reduced luciferase activity when
co-transfected with the ETS1-overexpression plasmid (HeLa:
P=0.0022; HL-1: P=0.0095).
Altogether, these findings demonstrated that the
ETS1 transcription factor could bind to the NOTCH4 promoter
region and promote gene expression. This interaction was inhibited
by methylation changes in the ETS1 binding site.
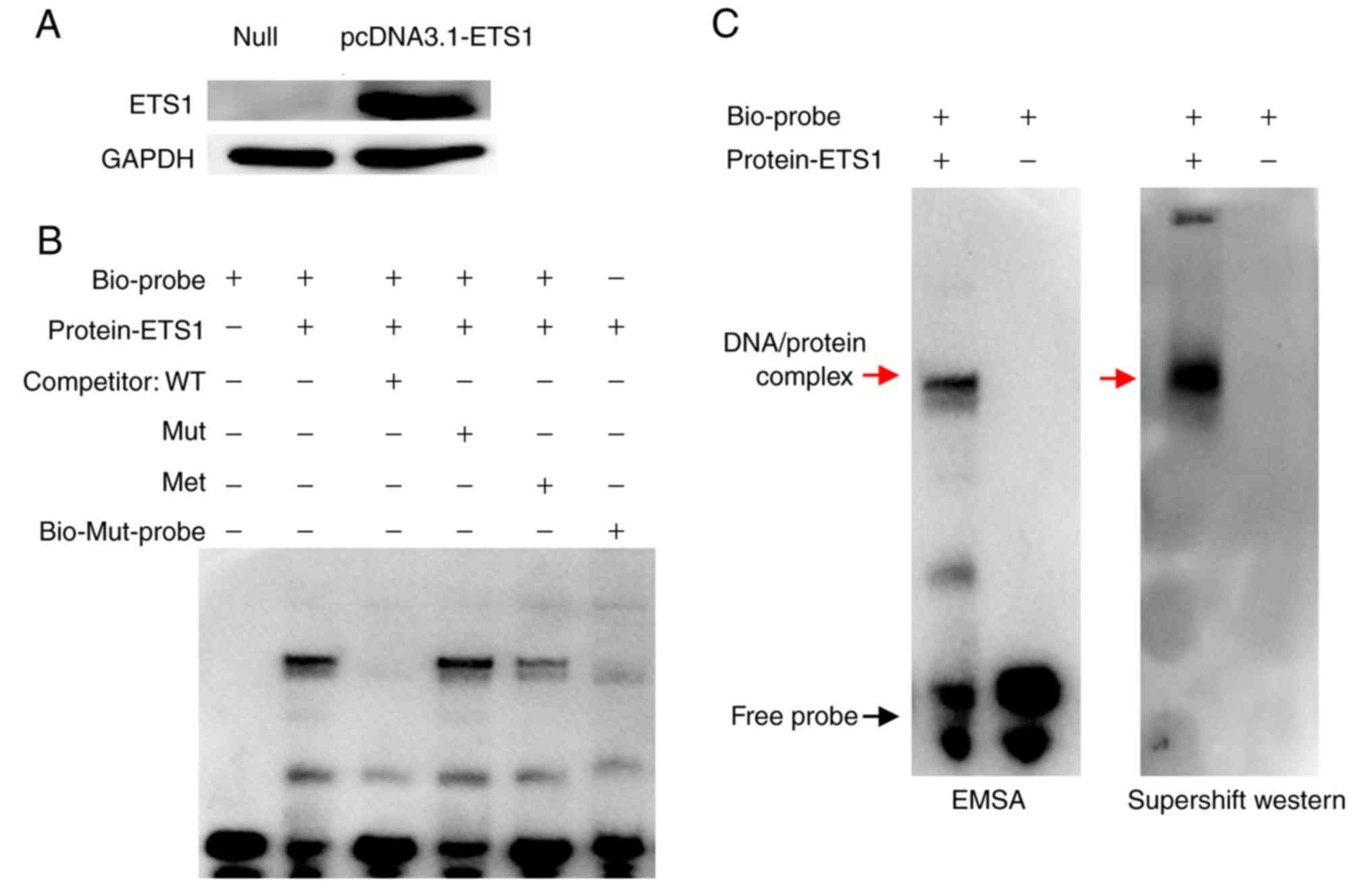
Impact of single CpG site 2
methylation on ETS1 binding affinity
The ETS1 transcription factor was overexpressed in
the 293T cell line (Fig. 5A), and
an EMSA was carried out to further confirm the effect of single CpG
site 2 methylation on ETS1 binding affinity to the NOTCH4
promoter region. The biotinylated probe (Biotin-probe) could bind
to the ETS1 transcription factor in the nuclear protein extract
from transfected cells, forming a visible DNA/protein complex
(Fig. 5B, lane 2). However, when
the unlabelled probe (Competitor-WT) was added as a competitor, the
band corresponding to the DNA/protein complex was not observed
(Fig. 5B, lane 3). However, with
the mutant unlabelled competition probe (Competitor-Mut), the
binding of ETS1 to the biotinylated probe was not affected
(Fig. 5B, lane 4). Moreover, with
the addition of an unlabelled methylated probe (Competitor-Met),
the band of the DNA/protein complex in lane 5 was lighter than that
in lane 2, but heavier than that in lane 3 (Fig. 5B, lane 5). In addition, the complex
was not detected when the mutated biotinylated probe
(Biotin-Mut-probe) was added (Fig.
5B, lane 6). A super-shift western blot was conducted to
confirm that the DNA/protein banding was caused by the presence of
ETS1 protein. The specific DNA/protein complexes are indicated by
red arrows. In addition, the results also demonstrated that the
biotinylated probe could bind to the ETS1 transcription factor, and
the DNA/protein complex was induced by ETS1 antibody, indicating
that the complex consists of ETS1 (Fig. 5C).
Collectively, the present findings indicated that
the ETS1 transcription factor could directly bind to the
NOTCH4_R region and was affected by methylation of the CpG 2
site.
Discussion
The Notch signalling pathway is evolutionarily
conserved and plays a critical role in the growth and development
of diverse organisms, including in normal cardiac development
(16–18). In vertebrates, Notch4 is
distributed in the aorta, the endocardium and the endothelial cells
of arteries, including pulmonary and cardiac vessels (40,41).
In the present study, immunohistochemical staining demonstrated
that NOTCH4 expression in RVOT myocardial tissues was significantly
decreased in patients with TOF, compared with controls, suggesting
that the developmental defects leading to TOF are associated with
distinct changes in NOTCH4 expression. Considering the low
frequency of genetic variants in the NOTCH4 coding region in
patients with TOF (20), it was
hypothesized that epigenetic changes might be at play in the
abnormal expression of this gene.
Several studies have demonstrated that both genetic
and epigenetic mechanisms control the expression of cardiac genes
in a spatiotemporal manner during cardiac development (42,43).
DNA methylation is one of the major epigenetic mechanisms
controlling gene expression, which can increase or decrease the
levels of gene transcription based on the methylation status of the
target gene (44,45). Abnormal methylation can change the
normal expression of genes and lead to different diseases. Abnormal
DNA methylation has been associated with the pathogenesis of
tetralogy of Fallot and can exacerbate defects, such as RVOT and
ventricular septal defect, leading to pathogenic cardiac
remodelling (46,47). Our previous study demonstrated that
altered expression of NK2 homeobox 5, heart and neural crest
derivatives expressed 1 and long interspersed nuclear element-1 may
be associated with epigenetic regulation and involved in the
development of TOF (22,48). Moreover, a single CpG site-based
methylation change was also demonstrated to be responsible for
changes in gene expression in different diseases (49,50).
In the current study, the methylation status of the NOTCH4
promoter region and its association with gene expression was
assessed. Considering that there is no CpG island in the
NOTCH4 promoter, an important regulatory region near the TSS
containing five CpG sites was selected for methylation analysis.
Overall, no significant difference was observed in the overall
methylation level between the patients with TOF and controls.
However, when the individual methylation status of each CpG site
was analysed separately, CpG site 2 exhibited significantly higher
methylation levels in the cardiac tissue of patients with TOF,
compared with the controls. Interestingly, in the patients, a
significant negative correlation was also observed between NOTCH4
expression and the methylation levels at the CpG site 2. These
findings indicated that CpG site 2 methylation changes may affect
NOTCH4 expression.
To determine the effect of CpG site 2 methylation
changes in the NOTCH4 promoter on gene expression and the
underlying molecular mechanism, a dual-luciferase assay combined
with an in vitro methylation assay was carried out.
Following in vitro methylation, decreased transcriptional
activity of pGL3-NOTCH4_-179/-53 was observed, suggesting
that increased methylation of the CpG site 2 had a negative impact
on transcriptional regulatory activity. Several studies have
suggested that CpG site-specific regulation of DNA methylation
could be mediated by transcription factors binding to specific gene
promoter regions (51,52). Therefore, using online prediction
software was used to identify a potential transcriptional factor
that could represent a target for the NOTCH4 promoter
region. A potential binding site for the ETS1 transcription factor
was identified in in the NOTCH4 promoter that contained CpG
site 2. The ETS1 transcription factor plays an important role in
normal cardiac development (32),
and can regulate transcriptional activity of target genes (such as
DLL4, NOTCH4 and MMP1) by binding to specific sites
(30,34–36).
In the present study, in vitro experiments
demonstrated that NOTCH4 expression in patients with TOF was
regulated by binding of ETS1 to NOTCH4_R. The present
findings suggested that, although ETS1 could bind to the
NOTCH4 promoter region and promote gene expression, binding
affinity could be influenced when a single CpG site 2 was
methylated, which might lead to a marked reduction of gene
expression. However, CpG site 2 hypermethylation could result in
reduced gene expression in vivo was not evaluated in this
study and remains unclear. Thus, the direct function of single CpG
site 2 methylation and its role in the regulation of NOTCH4
expression require further study.
Previous studies have demonstrated that DNA
methylation occurs not only on CpG islands, but also on CpG island
shores. Abnormal methylation in these regions may affect gene
expression by altering the chromosomal structure (53,54).
However, there is increasing evidence for the importance of the
methylation status of individual CpG sites in the regulation of
gene expression. For example, the methylation levels of a single
CpG site was inversely correlated with oestrogen receptor α
positivity in breast cancer specimens in a previous study (49). CpG site-specific methylation was
demonstrated to alter the binding affinities of specific
transcription factors that can activate or repress transcription
(55,56). In the present study, CpG site 2 was
differentially methylated in patients with TOF and controls, and
the influence of sex and age was excluded through statistical
analysis. In addition, abnormal methylation of CpG site 2 affected
the binding affinity of the ETS1 transcription factor to the
NOTCH4 gene and downregulate NOTCH4 expression. Furthermore,
the causes of TOF development are complex, and involve many other
factors, such as environmental, genetic and maternal factors such
as age of the mother, radiation and drugs used by mother.
Therefore, other potential interfering factors in the development
of TOF cannot be excluded and should be evaluated in future
studies.
A limitation of this study was the restricted sample
size, due to difficulties in obtaining sufficient matched cardiac
tissue from patients with TOF and controls. Further studies with a
larger number of samples are required to confirm the present
findings. In addition, it is uncertain whether the methylation
patterns observed in the samples were effects or causes of TOF,
since the onset of TOF preceded the measurement of methylation. The
exact mechanism underlying NOTCH4 promoter methylation in
the onset of TOF should also be explored in a large and prospective
cohort, as well as animal models.
In conclusion, the present study suggested that
single CpG site 2 in the NOTCH4 promoter region was
hypermethylated in RVOT myocardial tissue from patients with TOF,
which may lead to decreased NOTCH4 expression. Specifically, NOTCH4
expression may be regulated by an epigenetic mechanism, in which
single CpG site methylation at the binding site of the ETS1
transcription factor in NOTCH4_R decreases ETS1 binding
affinity and downregulates NOTCH4 expression. This, in turn, could
contribute to the development of TOF.
Acknowledgements
Not applicable.
Funding
The current study was supported by The National Key
Research and Development Program of China (grant no.
2016YFC1000500), The National Natural Science Foundation of China
(grant nos. 81570282, 81570283, 81873482 and 81873483) and The
Youth Program of National Natural Science Foundation of China
(grant no. 81800282).
Availability of the data and materials
The datasets used and/or analysed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
WS and GH made major contributions to the conception
and design of this study. MY and HX collected samples and
communicated with the patients' families. YZ performed the
experiments and wrote the manuscript. RG, MC, XL, and XM helped
collect and analyse the data. WS and GH supervised the study and
edited the manuscript. All authors read and approved the final
manuscript.
Ethics approval and consent to
participate
The present study was approved by the Ethics
Committee of The Children's Hospital of Fudan University [approval
no. 2015 (26)]. Written informed
consent was obtained from the parents or relatives of all study
participants.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
van der Linde D, Konings EE, Slager MA,
Witsenburg M, Helbing WA, Takkenberg JJ and Roos-Hesselink JW:
Birth prevalence of congenital heart disease worldwide: A
systematic review and meta-analysis. J Am Coll Cardiol.
58:2241–2247. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Zhao QM, Ma XJ, Ge XL, Liu F, Yan WL, Wu
L, Ye M, Liang XC, Zhang J, Gao Y, et al: Pulse oximetry with
clinical assessment to screen for congenital heart disease in
neonates in China: A prospective study. Lancet. 384:747–754. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Bedard E, Mccarthy KP, Dimopoulos K,
Giannakoulas G, Gatzoulis MA and Ho SY: Structural abnormalities of
the pulmonary trunk in tetralogy of Fallot and potential clinical
implications: A morphological study. J Am Coll Cardiol.
54:1883–1890. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Ho S, Mccarthy KP, Josen M and Rigby ML:
Anatomic- echocardiographic correlates: An introduction to normal
and congenitally malformed hearts. Heart. 86 (Suppl 2):II3–II11.
2001.PubMed/NCBI
|
|
5
|
Kathiriya IS, Nora EP and Bruneau BG:
Investigating the transcriptional control of cardiovascular
development. Circ Res. 116:700–714. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Kaynak B, von Heydebreck A, Mebus S,
Seelow D, Hennig S, Vogel J, Sperling HP, Pregla R,
Alexi-Meskishvili V, Hetzer R, et al: Genome-wide array analysis of
normal and malformed human hearts. Circulation. 107:2467–2474.
2003. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Nemer M: Genetic insights into normal and
abnormal heart development. Cardiovasc Pathol. 17:48–54. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Simeone RM, Tinker SC, Gilboa SM, Agopian
AJ, Oster ME, Devine OJ and Honein MA; National Birth Defects
Prevention Study, : Proportion of selected congenital heart defects
attributable to recognized risk factors. Ann Epidemiol. 26:838–845.
2016. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Pierpont ME, Basson CT, Benson DW Jr, Gelb
BD, Giglia TM, Goldmuntz E, McGee G, Sable CA, Srivastava D, Webb
CL, et al: Genetic basis for congenital heart defects: Current
knowledge: A scientific statement from the American heart
association congenital cardiac defects committee, council on
cardiovascular disease in the young: Endorsed by the American
academy of pediatrics. Circulation. 115:3015–3038. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Artavanis-Tsakonas S, Rand MD and Lake RJ:
Notch signaling: Cell fate control and signal integration in
developmen. Science. 284:770–776. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Luxán G, D'amato G, Macgrogan D and de la
Pompa JL: Endocardial Notch signaling in cardiac development and
disease. Circ Res. 118:e1–e18. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Mccright B, Lozier J and Gridley T: A
mouse model of Alagille syndrome: Notch2 as a genetic modifier of
Jag1 haploinsufficiency. Development. 129:1075–1082.
2002.PubMed/NCBI
|
|
13
|
Digilio MC, Luca AD, Lepri F, Guida V,
Ferese R, Dentici ML, Angioni A, Marino B and Dallapiccola B: JAG1
mutation in a patient with deletion 22q11.2 syndrome and tetralogy
of Fallot. Am J Med Genet A 161A. 3133–3136. 2013. View Article : Google Scholar
|
|
14
|
Garg V: Notch signaling in aortic valve
development and disease. Nakanishi T, Markwald RR, Baldwin HS,
Keller BB, Srivastava D and Yamagishi H: Etiology and morphogenesis
of congenital heart disease: From gene function and cellular
interaction to morphology [internet]. Tokyo: Springer; 2016,
Chapter 53. Jun 25. 2016, View Article : Google Scholar
|
|
15
|
Meester J, Verstraeten A, Alaerts M,
Schepers D, Van Laer L and Loeys BL: Overlapping but distinct roles
for NOTCH receptors in human cardiovascular disease. Clin Genet.
95:85–94. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Krebs LT, Xue Y, Norton CR, Shutter JR,
Maguire M, Sundberg JP, Gallahan D, Closson V, Kitajewski J,
Callahan R, et al: Notch signaling is essential for vascular
morphogenesis in mice. Genes Dev. 14:1343–1352. 2000.PubMed/NCBI
|
|
17
|
Leong KG, Hu X, Li L, Noseda M, Larrivée
B, Hull C, Hood L, Wong F and Karsan A: Activated Notch4 inhibits
angiogenesis: Role of beta 1-integrin activation. Mol Cell Biol.
22:2830–2841. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Uyttendaele H, Ho J, Rossant J and
Kitajewski J: Vascular patterning defects associated with
expression of activated Notch4 in embryonic endothelium. Proc Natl
Acad Sci USA. 98:5643–5648. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Noseda M, Mclean G, Niessen K, Chang L,
Pollet I, Montpetit R, Shahidi R, Dorovini-Zis K, Li L, Beckstead
B, et al: Notch activation results in phenotypic and functional
changes consistent with endothelial-to-mesenchymal transformation.
Circ Res. 94:910–917. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Page DJ, Miossec MJ, Williams SG, Monaghan
RM, Fotiou E, Cordell H, Sutcliffe L, Topf A, Bourgey M, Bourque G,
et al: Whole exome sequencing reveals the major genetic
contributors to nonsyndromic tetralogy of Fallot. Circ Res.
124:553–563. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Shames DS, Minna JD and Gazdar AF: DNA
methylation in health, disease, and cancer. Curr Mol Med. 7:85–102.
2007. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Boyes J and Bird A: DNA methylation
inhibits transcription indirectly via a methyl-CpG binding protein.
Cell. 64:1123–1134. 1991. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Sheng W, Chen L, Wang H, Ma X, Ma D and
Huang G: CpG island shore methylation of ZFPM2 is identified in
tetralogy of Fallot samples. Pediatr Res. 80:151–158. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Sheng W, Qian Y, Wang H, Ma X, Zhang P,
Diao L, An Q, Chen L, Ma D and Huang G: DNA methylation status of
NKX2-5, GATA4 and HAND1 in patients with tetralogy of Fallot. BMC
Med Genomics. 6:462013. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Gong J, Sheng W, Ma D, Huang G and Liu F:
DNA methylation status of TBX20 in patients with tetralogy of
Fallot. BMC Med Genomics. 12:752019. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Emeny RT, Baumert J, Zannas AS, Kunze S,
Wahl S, Iurato S, Arloth J, Erhardt A, Balsevich G, Schmidt MV, et
al: Anxiety associated increased CpG methylation in the promoter of
Asb1: A translational approach evidenced by epidemiological and
clinical studies and a murine model. Neuropsychopharmacology.
43:342–353. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Bind MA, Coull BA, Peters A, Baccarelli
AA, Tarantini L, Cantone L and Schwartz JD, Vokonas PS, Koutrakis P
and Schwartz JD: Beyond the mean: Quantile regression to explore
the association of air pollution with gene-specific methylation in
the normative aging study. Environ Health Perspect. 123:759–765.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Somineni HK, Zhang X, Biagini MJ, Kovacic
MB, Ulm A, Jurcak N, Ryan PH, Hershey GKK and Ji H: Ten-eleven
translocation 1 (TET1) methylation is associated with childhood
asthma and traffic-related air pollution. J Allergy Clin Immunol.
137:797–805.e5. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Plusquin M, Guida F, Polidoro S, Vermeulen
R, Raaschou-Nielsen O, Campanella G, Hoek G, Kyrtopoulos SA,
Georgiadis P, Naccarati A, et al: DNA methylation and exposure to
ambient air pollution in two prospective cohorts. Environ Int.
108:127–136. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Wythe JD, Dang LT, Devine WP, Boudreau E,
Artap ST, He D, Schachterle W, Stainier DYR, Oettgen P, Black BL,
et al: ETS factors regulate Vegf-dependent arterial specification.
Dev Cell. 26:45–58. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Oikawa T and Yamada T: Molecular biology
of the Ets family of transcription factors. Gene. 303:11–34. 2003.
View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Kola I, Brookes S, Green AR, Garber R,
Tymms M, Papas TS and Seth A: The Ets1 transcription factor is
widely expressed during murine embryo development and is associated
with mesodermal cells involved in morphogenetic processes such as
organ formation. Proc Natl Acad Sci USA. 90:7588–7592. 1993.
View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Ye M, Coldren C, Liang X, Mattina T,
Goldmuntz E, Benson DW, Ivy D, Perryman MB, Garrett-Sinha LA and
Grossfeld P: Deletion of ETS-1, a gene in the Jacobsen syndrome
critical region, causes ventricular septal defects and abnormal
ventricular morphology in mice. Hum Mol Genet. 19:648–656. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Gao Z, Kim GH, Mackinnon AC, Flagg AE,
Bassett B, Earley JU and Svensson EC: Ets1 is required for proper
migration and differentiation of the cardiac neural crest.
Development. 137:1543–1551. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Ramirez K, Chandler KJ, Spaulding C, Zandi
S, Sigvardsson M, Graves BJ and Kee BL: Gene deregulation and
chronic activation in natural killer cells deficient in the
transcription factor ETS1. Immunity. 36:921–932. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Li RZ, Pei HP, Watson DK and Papas TS:
EAP1/Daxx interacts with ETS1 and represses transcriptional
activation of ETS1 target genes. Oncogene. 19:745–753. 2000.
View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Pham VN, Lawson ND, Mugford JW, Dye L,
Castranova D, Lo B and Weinstein BM: Combinatorial function of ETS
transcription factors in the developing vasculature. Dev Biol.
303:772–783. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
TFSEARCH, . http://www.cbrc.jp/research/db/TFSEARCH.htmlJune
13–2020
|
|
39
|
JASPAR database, . http://jaspar.binf.ku.dk/cgi-bin/jaspar_db.pl?rm=browse&db=core&tax_group=vertebratesJune
13–2020
|
|
40
|
Iso T, Hamamori Y and Kedes L: Notch
signaling in vascular development. Arterioscler Thromb Vasc Biol.
23:543–553. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Villa N, Walker L, Lindsell CE, Gasson J,
Iruela-Arispe ML and Weinmaster G: Vascular expression of Notch
pathway receptors and ligands is restricted to arterial vessels.
Mech Dev. 108:161–164. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Wu B, Zhang Z, Lui W, Chen X, Wang Y,
Chamberlain AA, Moreno-Rodriguez RA, Markwald RR, O'Rourke BP,
Sharp DJ, et al: Endocardial cells form the coronary arteries by
angiogenesis through myocardial-endocardial VEGF signaling. Cell.
151:1083–1096. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Wu B, Wang Y, Lui W, Langworthy M,
Tompkins KL, Hatzopoulos AK, Baldwin HS and Zhou B: Nfatc1
coordinates valve endocardial cell lineage development required for
heart valve formation. Circ Res. 109:183–192. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Borgel J, Guibert S, Li Y, Chiba H,
Schübeler D, Sasaki H, Forné T and Weber M: Targets and dynamics of
promoter DNA methylation during early mouse development. Nat Genet.
42:1093–1100. 2010. View
Article : Google Scholar : PubMed/NCBI
|
|
45
|
Jones PA and Takai D: The role of DNA
methylation in mammalian epigenetics. Science. 293:1068–1070. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Chamberlain AA, Lin M, Lister RL, Maslov
AA, Wang Y, Suzuki M, Wu B, Greally JM, Zheng D and Zhou B: DNA
methylation is developmentally regulated for genes essential for
cardiogenesis. J Am Heart Assoc. 3:e9762014. View Article : Google Scholar
|
|
47
|
Feinberg AP: The key role of epigenetics
in human disease prevention and mitigation. N Engl J Med.
378:1323–1334. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Sheng W, Wang H, Ma X, Qian Y, Zhang P, Wu
Y, Zheng F, Chen L, Huang G and Ma D: LINE-1 methylation status and
its association with tetralogy of fallot in infants. BMC Med
Genomics. 5:202012. View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Tsuboi K, Nagatomo T, Gohno T, Higuchi T,
Sasaki S, Fujiki N, Kurosumi M, Takei H, Yamaguchi Y, Niwa T and
Hayashi SI: Single CpG site methylation controls estrogen receptor
gene transcription and correlates with hormone therapy resistance.
J Steroid Biochem Mol Biol. 171:209–217. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Fürst RW, Kliem H, Meyer HH and Ulbrich
SE: A differentially methylated single CpG-site is correlated with
estrogen receptor alpha transcription. J Steroid Biochem Mol Biol.
130:96–104. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Bartels SJ, Spruijt CG, Brinkman AB,
Jansen PW, Vermeulen M and Stunnenberg HG: A SILAC-based screen for
Methyl-CpG binding proteins identifies RBP-J as a DNA methylation
and sequence-specific binding protein. PLoS One. 6:e258842011.
View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Mann IK, Chatterjee R, Zhao J, He X,
Weirauch MT, Hughes TR and Vinson C: CG methylated microarrays
identify a novel methylated sequence bound by the CEBPB|ATF4
heterodimer that is active in vivo. Genome Res. 23:988–997. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
53
|
Irizarry RA, Ladd-Acosta C, Wen B, Wu ZJ,
Montano C, Onyango P, Cui HM, Gabo K, Rongione M, Webster M, et al:
The human colon cancer methylome shows similar hypo- and
hypermethylation at conserved tissue-specific CpG island shores.
Nat Genet. 41:178–186. 2009. View
Article : Google Scholar : PubMed/NCBI
|
|
54
|
Portela A and Esteller M: Epigenetic
modifications and human disease. Nat Biotechnol. 28:1057–1068.
2010. View Article : Google Scholar : PubMed/NCBI
|
|
55
|
Claus R, Lucas DM, Stilgenbauer S, Ruppert
AS, Yu L, Zucknick M, Mertens D, Bühler A, Oakes CC, Larson RA, et
al: Quantitative DNA methylation analysis identifies a single CpG
dinucleotide important for ZAP-70 expression and predictive of
prognosis in chronic lymphocytic leukemia. J Clin Oncol.
30:2483–2491. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
56
|
Ceccarelli V, Racanicchi S, Martelli MP,
Nocentini G, Fettucciari K, Riccardi C, Marconi P, Nardo PD,
Grignani F, Binaglia L and Vecchini A: Eicosapentaenoic acid
demethylates a single CpG that mediates expression of tumor
suppressor CCAAT/enhancer-binding protein delta in U937 leukemia
cells. J Biol Chem. 286:27092–27102. 2011. View Article : Google Scholar : PubMed/NCBI
|