Introduction
Human cathelicidin antimicrobial peptide (CAMP) and
its active product, LL-37, serve important roles in infectious
diseases (including viral, bacterial and fungal infections) and
autoimmune diseases (1–5). Previously, it has been found that
CAMP/LL-37 also has a significant effect on the initiation and
progression of various types of cancer (6–9).
Additionally, it has been indicated that the upregulation of
CAMP/LL-37 expression contributes to its tumorigenic effect in
breast cancer, ovarian cancer, lung cancer, prostate cancer,
pancreatic cancer, malignant melanoma and skin squamous cell
carcinoma (10–16). Moreover, the downregulation of
CAMP/LL-37 expression contributes to its anticancer effects on
colon cancer, gastric cancer and haematological malignancies
(17–21). The LL-37-induced activation of
membrane receptors and subsequent signalling pathways leads to the
alteration of cellular functions (6–9).
Different membrane receptors on various cancer cells appear to be
responsible for the tissue-specific effects of LL-37 (6–9).
hCAP18109-135, a 27 amino acids peptide
in the C-terminal of hCAP18, induces caspase-independent apoptosis
of oral squamous cell carcinoma (OSCC) SAS-H1 cells (22). KI-21-3, a shortened fragment of
LL-37, exhibits considerable oncolytic properties on SCC-4
carcinoma cells via the antiproliferative and caspase-3 dependent
apoptotic pathway (23). A
previous study by the same authors also demonstrated that the
expression levels of CAMP/LL-37 were downregulated significantly in
OSCC tissues (24), suggesting
that CAMP exerts an inhibitory effect on the initiation and
progression of OSCC, although the underlying mechanisms remain
unclear in OSCC.
The aim of the present study was to investigate the
roles and mechanisms of action of human CAMP/LL-37 in OSCC HSC-3
cells (25,26) using a colony-formation, CCK-8,
wound healing and Transwell invasion assays, as well as expression
analysis on the basis of established LL-37 C-terminal deletion
mutants (CDEL) and CAMP stable overexpression in OSCC HSC-3 cell
lines.
To the best of our knowledge, the results of the
present study are the first to determine the potential mechanism
underlying the effects of the overexpression of human CAMP in OSCC
HSC-3 cells.
Materials and methods
Plasmid construction
High-fidelity Herculase enzyme (Agilent
Technologies, Inc.) was used for amplification of cDNA from foetal
brain tissue (Clontech Laboratories) and was used as the template.
The primers used for the amplification of the open reading frame
(ORF) of CAMP were as follows: Forward,
5′-CGGAATTCAATGGGGACCATGAAGACCCAAAGG-3′ and reverse,
5′-CGGGATCCCTAGGACTCTGTCCTGGGTACAAG-3′. The primers used for the
amplification of the ORF of CDEL were as follows: Forward,
5′-CGGAATTCAATGGGGACCATGAAGACCCAAAGG-3′ and reverse,
5′-CGGGATCCCTAGGCAAATCTCTTGTTATCCTTATCACAAC-3′. The amplification
conditions were as follows: Pre-denaturation, 95°C for 2 min;
followed by 40 cycles at 95°C for 20 sec, 55°C for 20 sec and 72°C
for 45 sec; with a final extension step of 72°C for 3 min. PCR
products were purified and were then ligated in to pFlag-CMV4
(Sigma-Aldrich; Merck KGaA). Plasmid constructs were confirmed by
sequencing (Sangon Biotech Co., Ltd.).
Cell culture
Human OSCC HSC-3 cells (ATCC) were cultured in
RPMI-1640 medium (Gibco; Thermo Fisher Scientific, Inc.)
supplemented with 10% FBS (Gemini Bio-Products). KB cells (ATCC)
were maintained in modified Eagle's medium (Gibco; Thermo Fisher
Scientific, Inc.) supplemented with 10% FBS. The cells were
incubated in an incubator at 37°C with 5% CO2.
Generation of CDEL or CAMP stably
expressing cells
HSC-3 cells were grown to 60–80% confluency, and
transfected using Expressfect™ Transfection Reagent (Denville
Scientific). Following 24 h of transfection, the transfected cells
were treated with 500 ng/µl G418 (Life Science). The medium
containing G418 was replaced every 48 h. Cells were subsequently
cultured with 200 ng/µl G418 in the medium to maintain cell
resistance. Single positively transfected cell clones were screened
using the filter paper method (27). The expression of either CDEL or
CAMP was confirmed by western blot analysis and immunofluorescence
(28–30).
Immunofluorescence
Stably transfected HSC-3 cells were fixed with cold
methanol for 5 min at −20°C, then washed with DPBS, and
permeabilized with 0.25% Triton X-100 for 10 min. After blocking
for 1 h with 1% BSA in DPBS/0.1% Tween-20, and rinsing with DPBS
three times, the cells were incubated with rabbit anti-cathelicidin
(cat. no. ab69484; Abcam) or mouse anti-LL37 (cat. no. sc-166770;
Santa Cruz Biotechnology, Inc.) antibodies at room temperature for
1 h, followed by incubation with Alexa Fluor 488-conjugated goat
anti-rabbit IgG or Alexa Fluor 594-conjugated goat anti-mouse IgG
(ProteinTech Group, Inc.) at room temperature for 1 h (27,28).
Nuclei were stained with 300 nM DAPI at room temperature for 1 min,
and the cells were then imaged using an epifluorescence microscope
(Nikon Eclipse Ti; Nikon Corporation; magnification, ×400).
Colony formation assay
Stably transfected HSC-3 cells were adjusted to
1,500 cells per dish. The solution was replaced every 3 days, and
crystal violet staining was performed at room temperature for 3 min
12 days later. Cells were imaged by camera. A group of >50 cells
was considered as one colony. The number of clones was counted
under a light microscope (magnification, ×100) and 5 fields of view
were analyzed.
Cell Counting Kit-8 (CCK-8) assay
A CCK-8 assay (Dojindo Molecular Technologies, Inc.)
was used to measure cell viability. Stably transfected HSC-3 cells
(9×103 cells/well) were seeded into a 24-well plate.
Viability was assessed after 0, 24, 48 and 72 h following complete
adherence. A total of 100 µl CCK-8 reagent was added to each well
of the first 24-well plate, covered with tin foil and placed in a
cell incubator (37°C, 5% CO2) for 2 h, and subsequently
the absorbance was measured at 450 nm using a microplate reader
(BioTek Instruments, Inc.).
Wound healing assay
Stably transfected HSC-3 cells were seeded into
1.2×106 cells/well in a 6-well plate, and cultured for
16 h to form a confluent monolayer. The confluent cultures were
scratched with sterile 200-µl pipette tips. The culture medium was
removed, and washed three times with DPBS. After washing and
removing the cell debris, the cells were cultured with serum-free
RPMI medium, and the images were captured over time using a light
microscope (IX-70; Olympus; magnification, ×40) and analysed using
ImageJ version 1.8.0 software (National Institutes of Health).
Transwell invasion assay
The stably transfected HSC-3 cells
(2.5×105 /ml, 200 µl) were seeded into a Matrigel-coated
Transwell upper chamber (Corning, Inc.) in serum-free RPMI medium,
and ~750 µl RPMI medium containing 20% serum was added to the lower
chambers. The chambers were placed in the 24-well plate, and
cultured in a cell incubator for 64 h and subsequently stained with
haematoxylin at room temperature for 10 min, and observed and
imaged using a light microscope (magnification, ×200).
Western blot analysis
Cells were washed with DPBS and then total protein
was extracted using mammalian cell lysate buffer (Biyuntian
Bio-Technology Co., Ltd.) containing 1 mM phenylmethylsulfonyl
fluoride. The cell extracts were centrifuged at 12,000 × g for 5
min at 4°C, and the supernatants were collected. The protein
concentrations were determined using a bicinchoninic acid protein
concentration detection kit (Biyuntian Bio-Technology, Co., Ltd.).
Total protein lysates (20 µg protein) were separated by 12%
SDS-PAGE, and transferred to 0.22-µM nitrocellulose membranes,
which were blocked in Tris-buffered saline Tween (0.1% Tween-20)
containing 5% non-fat dry milk for 1 h and incubated overnight at
4°C with primary antibodies against caspase-3 (cat. no.
19677-1-AP), poly(ADP-ribose) polymerase (PARP; cat. no.
66520-1-Ig), P53 (cat. no. 60283-1-Ig), BAX (cat. no. 60267-1-Ig),
Bcl-2 (cat. no. 60178-1-Ig), BCL-xL (cat. no. 66020-1-Ig), cyclin
B1 (cat. no. 55004-1-AP), PKR-like ER kinase (PERK; cat. no.
20582-1-AP), Akt (cat. no. 60203-2-Ig), phospho- (p-)Akt (cat. no.
66444-1-Ig), ERK (cat. no. 66192-1-Ig) (all from ProteinTech Group,
Inc.), cleaved-caspase-3 (cat. no. 9661; Cell Signalling
Technology, Inc.), cathelicidin (cat. no. ab69484; Abcam), p-ERK
(cat. no. ab76299; Abcam), LL-37 (cat. no. sc-166770; Santa Cruz
Biotechnology, Inc.) or β-actin (cat. no. TA-09; OriGene
Technologies, Inc.) all at a dilution of 1:2,000. The membranes
were washed and then incubated for 2 h at room temperature with
horseradish peroxidase-conjugated goat anti-rabbit antibodies or
goat anti-mouse antibodies (cat. nos. EM35111-01 and EM35110-01;
EMAR Biotechnology) at a dilution of 1:3,000. The immunoreactions
were visualized using Clarity™ Western ECL substrate (Bio-Rad
Laboratories, Inc.) and exposed to Amersham Hyperfilm ECL film
(Amersham, Cytiva). Protein bands were quantified by Quantity One
version 4.6.3 (Bio-Rad Laboratories, Inc.).
RNA-seq
Total cellular RNA was extracted using
TRIzol® reagent (Thermo Fisher Scientific, Inc.). The
RNA-Seq assay was performed (Novagene) as described previously
(31). Briefly, mRNA-seq libraries
were prepared using standard Illumina protocols, and the mRNA
libraries were then sequenced on an Illumina HiSeq 2000 platform
using a 101-bp paired-end sequencing strategy. A reads per kilobase
transcriptome per million reads method was used to calculate
expression levels of genes (32).
Differential expression analysis of two groups was performed using
the DESeq R package version 1.10.1 (33). KOBAS version 2.0 software
(http://kobas.cbi.pku.edu.cn) was used to
examine the statistical enrichment of differentially expressed
genes in Kyoto Encyclopaedia of Genes and Genomes (KEGG)
pathways.
Statistical analysis
A total of 3–6 independent experiments were
performed in the present study. Statistical analysis was performed
using SPSS version 19.0 software (IBM Corp.). A one-way ANOVA
followed by a Bonferroni's post hoc test was used for multiple
comparisons. P<0.05 was considered to indicate a statistically
significant difference.
Results
Generation of stable expression cell
lines
To explore the roles and mechanisms of human CAMP in
OSCC cells, a full-length CAMP and a LL-37 CDEL were used (Fig. 1A). To generate stably expressing
cell lines, either CDEL or CAMP ORFs were cloned into the
eukaryotic expression vector pFlag-CMV4, and these vectors were
then transfected into HSC-3 cells after selecting for transfected
cells using G418, different monoclonal cells were obtained by
gradient dilution. Monoclonal cells which stably expressed either
CDEL or CAMP were screened by western blot analysis after expanding
the cultures (Fig. 1B and C).
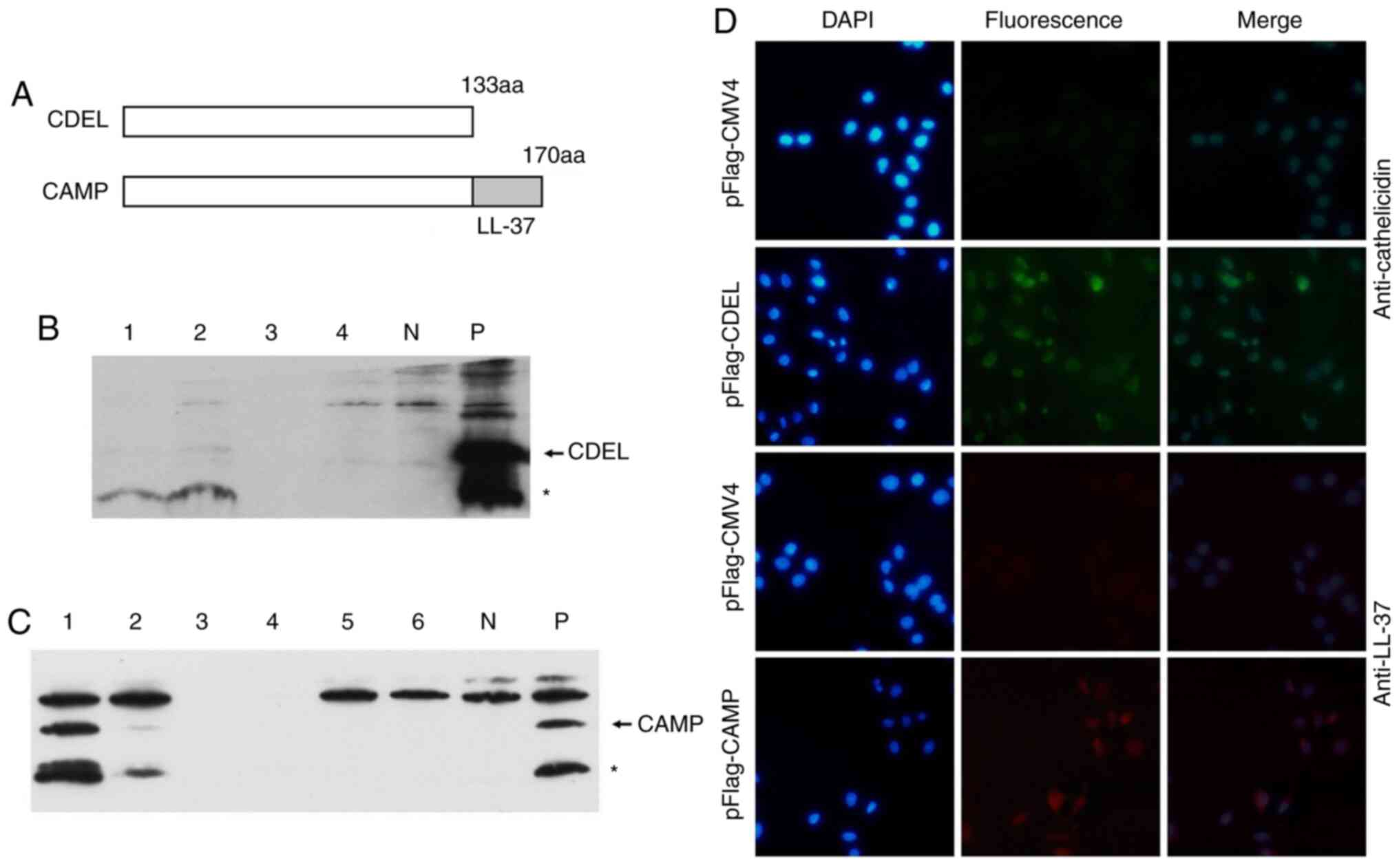 | Figure 1.Generation of CDEL and CAMP stably
transfected cells. (A) Structure diagram of CDEL and CAMP. (B) CDEL
protein expression examined by western blot analysis using
anti-cathelicidin antibody. Lane 1–4, protein extracts from
different clones. KB cells were transiently transfected with
pFlag-CMV4 (N) and pFlag-CDEL(P), respectively. (C) CAMP protein
expression examined by western blot analysis using anti-LL-37
antibody. Lane 1–6, protein extracts from different clones. KB
cells were transiently transfected with pFlag-CMV4 (N) and
pFlag-CAMP (P), respectively. (D) Characterization of CDEL and CAMP
stably transfected cells using immunofluorescence analysis. In the
above two rows of pFlag-CMV4 and pFlag-CDEL, the anti-cathelicidin
antibody was used as the primary antibody, and Alexa Fluor
594-conjugated goat anti-rabbit IgG antibody was used as the
secondary fluorescent antibody. In the following two rows of
pFlag-CMV4 and pFlag-CAMP, anti-LL-37 was used as the primary
antibody, and Alexa Fluor 594-conjugated goat anti-mouse IgG was
used as the secondary fluorescent antibody. Arrows indicate the
target proteins, and asterisks indicate degraded target proteins.
CDEL, LL-37 C-terminal deletion mutant. |
The results of the immunofluorescence analysis
revealed that there was no significant fluorescence of
pFlag-CMV4-transfected cells (negative control), and notable green
fluorescence was observed in the pFlag-CDEL-transfected cells
(Fig. 1D). The results also
illustrated that no significant fluorescence was observed in the
pFlag-CMV4-transfected cells (negative control), whereas red
fluorescence was observed in the pFlag-CAMP-transfected cells
(Fig. 1D). These results indicated
that HSC-3 cells with CDEL and CAMP stable expression were
successfully generated.
CDEL and CAMP stable expression in
HSC-3 cells inhibits colony formation and cell proliferation
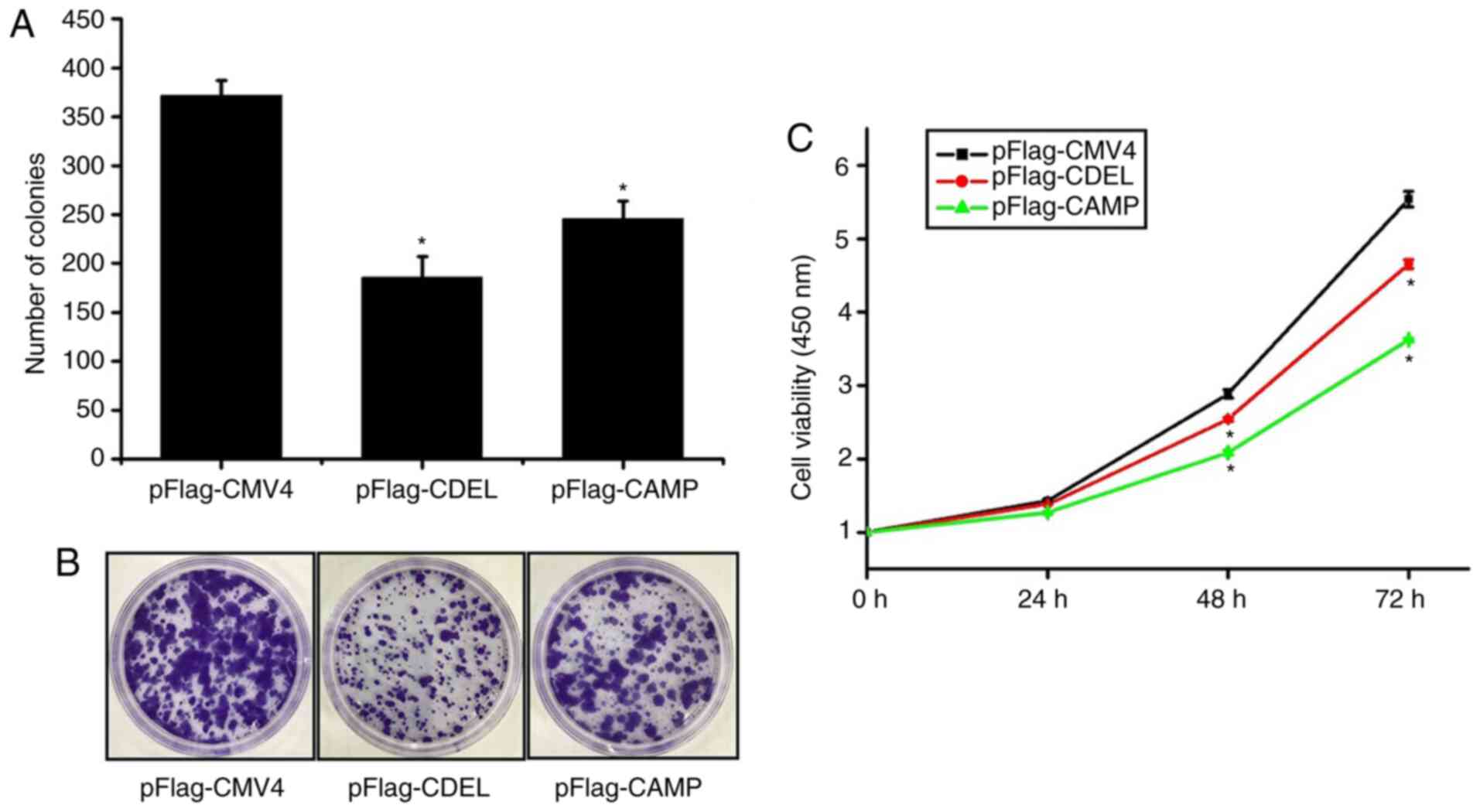
The results of the colony-formation assay revealed
that the number of cell clones of either CDEL or CAMP stably
expressing HSC-3 cells was significantly lower than that of HSC-3
cells stably transfected with the empty vector (Fig. 2A and B). This result demonstrated
that either CDEL or CAMP stable expression in HSC-3 cells inhibited
the colony formation ability compared with the control.
The results of the CCK-8 assay showed that the
viability of either the CDEL or CAMP stably expressing HSC-3 cells
was lower than that of the empty vector-transfected HSC-3 cells
after 24, 48 and 72 h, respectively (Fig. 2C). These results indicated that the
proliferation and viability of either the CDEL or CAMP stably
expressing HSC-3 cells were significantly lower than that of the
controls.
CDEL and CAMP stable expression in
HSC-3 cells inhibits cell migration and invasion
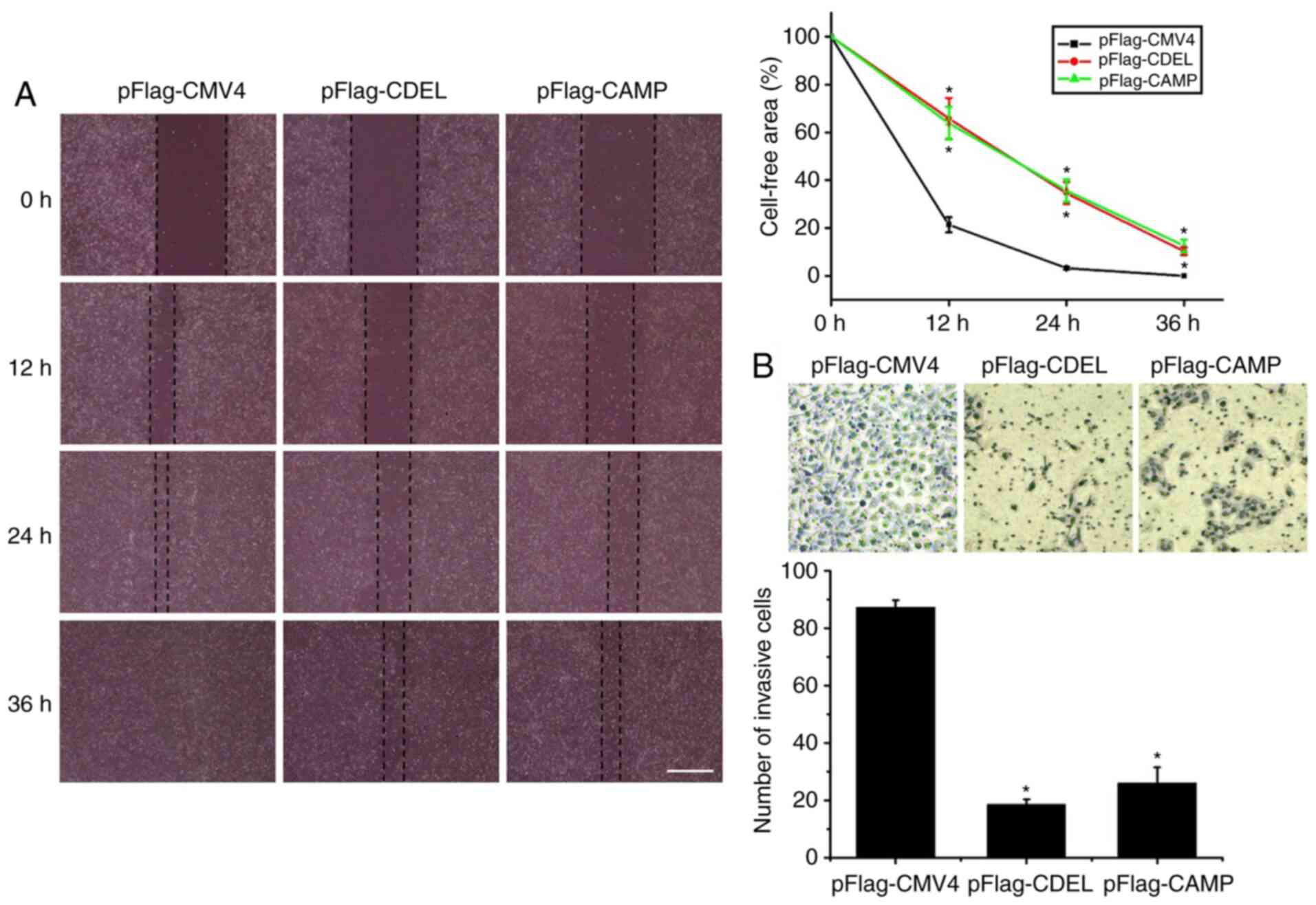
The results of the wound healing assay revealed that
the cell-free area of either the CDEL or CAMP stably expressing
HSC-3 cells was significantly larger than that of the controls at
12, 24 and 36 h, respectively (Fig.
3A). This result indicated that the migratory ability of either
the CDEL or CAMP stably expressing HSC-3 cells was significantly
lower than that of the controls.
The results of the Transwell invasion assay revealed
that the number of invaded CDEL or CAMP stably expressing HSC-3
cells were significantly lower than those of the controls (Fig. 3B). This result indicates that the
invasive ability of either the CDEL or CAMP stably expressing HSC-3
cells is decreased significantly compared with that of the
controls.
Either CDEL or CAMP overexpression
triggers caspase-3 mediated apoptosis of HSC-3 cells
To explore the mechanisms of either CDEL or CAMP in
HSC-3 cells, western blot analysis was performed. The results
revealed that the expression levels of cleaved caspase-3 and
cleaved PARP were significantly upregulated, whereas the expression
levels of caspase-3 were downregulated in both the CDEL- or
CAMP-transfected cells (Fig. 4A).
Moreover, the expression levels of the apoptosis-promoting
proteins, P53 and BAX, in either the CDEL- or CAMP-transfected
cells were significantly upregulated, whereas the expression levels
of the apoptosis-inhibiting proteins, Bcl-2 and Bcl-xL, in either
the CDEL- or CAMP-transfected cells were significantly
downregulated (Fig. 4A). These
results indicated that CDEL or CAMP overexpression induced
caspase-3 mediated apoptosis via the P53-Bcl-2/BAX signalling
pathway in OSCC HSC-3 cells. Taken together, these results
suggested that either CDEL or CAMP overexpression induces caspase-3
mediated apoptosis of HSC-3 cells and exerts a tumour-suppressive
effect on stably transfected HSC-3 cells.
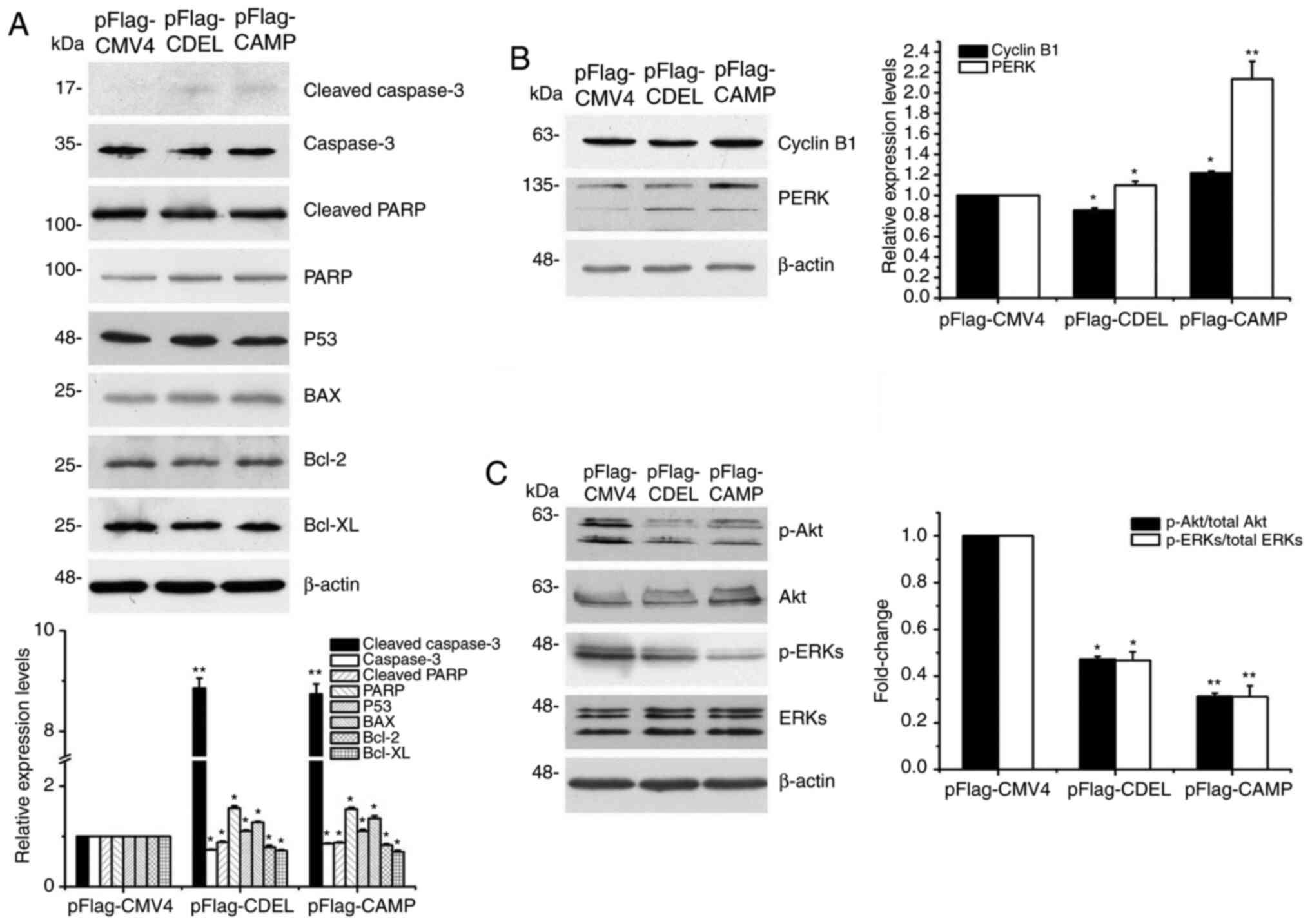 | Figure 4.CDEL, as well as CAMP overexpression
can induce caspase-3 mediated apoptosis. (A) Expression analysis of
apoptosis-related proteins, including cleaved-caspase-3, caspase-3,
cleaved-PARP, p53, BAX, Bcl-2 and Bcl-xL using western blot
analysis. (B) Expression analysis of cell cycle-related proteins,
including Cyclin B1 and PERK using western blot analysis. (C)
Expression analysis of phosphorylated and total Akt and ERK. The
ratio of p-Akt/total Akt and p-ERKs/total ERKs was quantified.
Representative blots of independent experiments are shown. The
bands were quantified relative to β-actin. *P<0.05, **P<0.01
vs. control. CDEL, LL-37 C-terminal deletion mutant; PARP,
poly(ADP-ribose) polymerase; PERK, PKR-like ER kinase; p-,
phospho. |
The results demonstrated that the expression levels
of the cell cycle-related proteins, Cyclin B1 or PERK, were
significantly upregulated in the CAMP-, but not in the
CDEL-transfected cells (Fig. 4B),
which suggested that the possible mechanisms by which CDEL and CAMP
overexpression in cells affected the cell cycle was different.
Moreover, the p-Akt/total Akt and p-ERKs/total ERKs ratios were
notably decreased in both the CDEL- or CAMP-transfected cells
(Fig. 4C).
Comparison of transcriptional
profiles
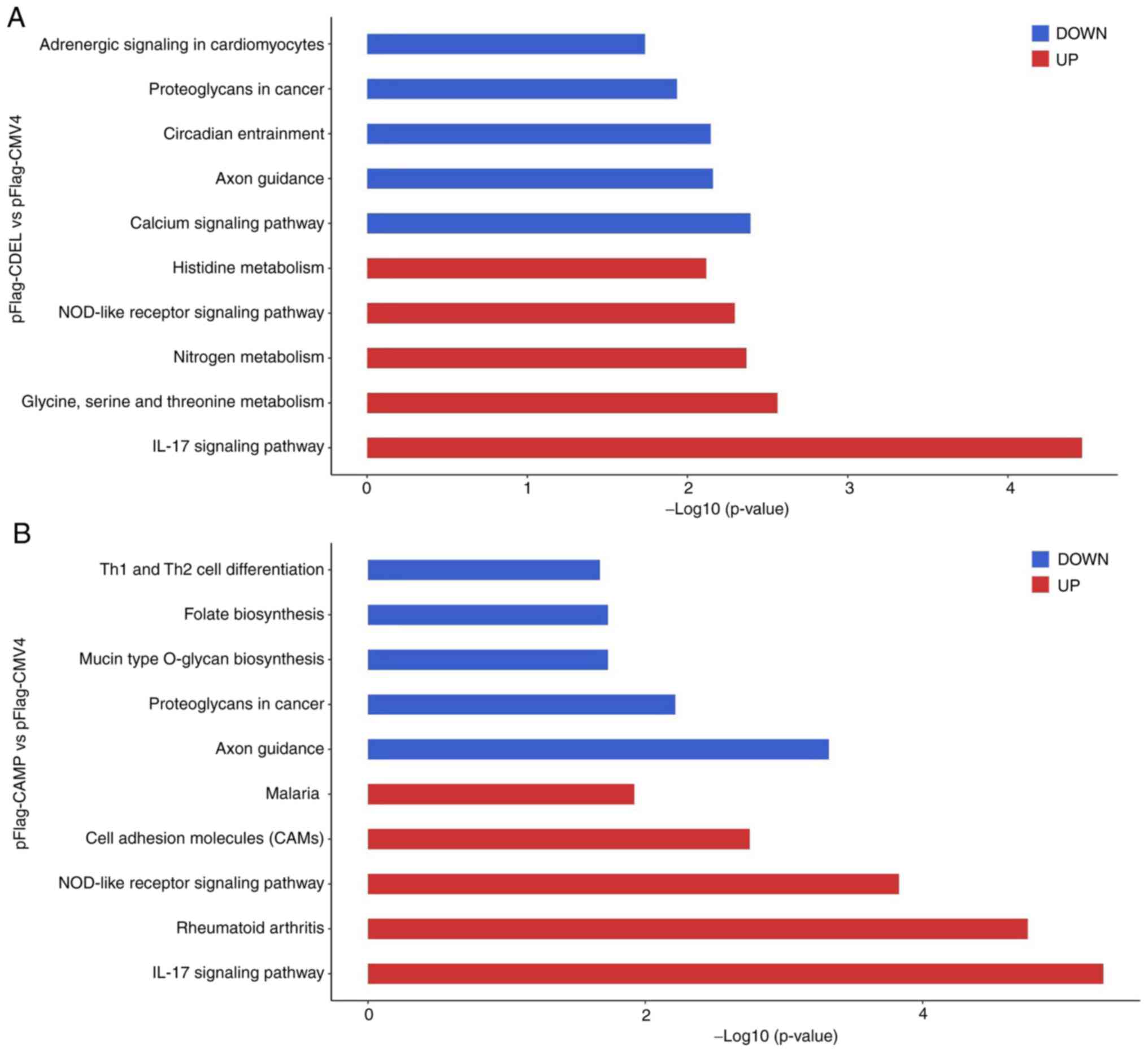
To further explore other possible molecular
mechanisms of either CDEL or CAMP in OSCC HSC-3 cells, the
transcriptomes of three stable cell lines were compared. The
comparison of the CDEL-transfected HSC-3 cells with the controls
revealed that the top upregulated genes were involved in the
enrichment of KEGG processes associated with IL-17 signalling
pathway, glycine, serine and threonine metabolism, nitrogen
metabolism, nucleotide oligomerization domain (NOD)-like receptor
signalling pathway and histidine metabolism; whereas the top
downregulated genes were involved in the calcium signalling
pathway, axon guidance circadian entrainment (Fig. 5A). Similarly, the comparison of the
CAMP-transfected HSC-3 cells with the controls revealed that top
upregulated genes were involved in the IL-17 signalling pathway,
rheumatoid arthritis and NOD-like receptor signalling pathway;
whereas the top downregulated genes were involved in axon guidance,
proteoglycans in cancer and mucin type O-glycan biosynthesis
(Fig. 5B). Each comparison
indicated that the top upregulated genes were involved in
IL-17-dependent and NOD-like receptor signalling pathways, which
are closely associated with the initiation and progression of
cancer (34,35).
Discussion
Previous studies have demonstrated the roles and
mechanisms of human CAMP/LL-37 involvement in various types of
cancer (6–9). A previous study also indicated that
human CAMP/LL-37 was expressed at low levels in OSCC, where it may
exert an inhibitory effect on development, and DNA methylation may
serve a role in OSCC by directly downregulating CAMP gene promoter
activity (24). Human CAMP/LL-37
has multiple roles and mechanisms in various cancer cells (6–21,36).
Both tumorigenic and anti-cancer effects induced by CAMP/LL-37 have
been reported in various types of cancer, and the combined effects
of those involved mechanisms determine the final effect (6,9). The
possible mechanisms of human CAMP/LL-37 in the different OSCC cells
have also been explored previously (22,23).
However, the detailed roles and underlying mechanisms of human
CAMP/LL-37 in OSCC cells remains unknown, to the best of our
knowledge. The present study provides an improved understanding of
the roles and mechanisms of human CAMP/LL-37 in OSCC.
The overexpression of either CDEL or CAMP in OSCC
HSC-3 cells reduced colony-formation, proliferation and viability,
as well as their migratory and invasive ability. The results
revealed that the overexpression of either CDEL or CAMP also
promoted the apoptosis of HSC-3 cells, and induced caspase-3
mediated apoptosis via the P53-Bcl-2/BAX signalling pathway, so as
to exert an anticancer effect on OSCC HSC-3 cells. It has been
previously shown that human CAMP/LL-37 and its analogues induce
caspase-independent apoptosis in colon cancer (18,19)
and haematological malignancies (20). The results of the present study
also showed that the overexpression of CDEL exerted a more potent
inhibitory effect on colony formation, migration and invasion
ability compared with that of CAMP. Therefore, it seems that the
LL-37 fragment may exert an opposing effect to that of the CDEL
fragment. The anti-cancer effects of cathelicidin LL-37 in gastric
cancer has been proposed to involve G0/G1 phase cell cycle arrest
(17). The cell cycle arrest in
G2/M via p21 activation leads to the anti-proliferative effects of
cathelicidin LL-37 in colon cancer cells (19,37).
The results of the present study also demonstrated that the levels
of cell cycle-related proteins, cyclin B1 and PERK, were
significantly upregulated in the CAMP, but not in the
CDEL-transfected cells. These results suggested that the potential
mechanism of anticancer effects of CAMP but not CDEL on OSCC HSC-3
cells may be attributed to cell cycle regulation. The Akt and ERK
signalling pathways are associated with proliferation,
differentiation, migration, invasion and apoptosis in cancer cells
(38,39). The decreased p-Akt/total Akt and
p-ERKs/total ERKs ratios suggested that the Akt and ERK signalling
pathway may be involved in the tumour-suppressive function of
either CDEL or CAMP in HSC-3 cells (Fig. 4C). Conversely, the results also
suggested that the C-terminal of CAMP (LL-37) may exert marked
effects on OSCC cells, and HSC-3 cells stimulated by artificially
synthesized LL-37, thus, small peptides may assist in examining the
effects of LL-37 extracellular treatment and its mechanism of
action.
IL-17-dependent and NOD-like receptor signalling
pathways have significant roles in inflammatory diseases and cancer
(34,35). Transcriptome analysis revealed
differences between CDEL or CAMP-transfected HSC-3 cells when
compared with the control. Upregulated genes involved in the
IL-17-dependent and NOD-like receptor signalling pathways were
observed for each comparison, whereas a number of differentially
expressed genes were also observed for each comparison. However,
the detailed molecular mechanism remains to be further
clarified.
In the present study, determination of the protein
expression of pFlag-CDEL stably-transfected HSC-3 cells using a
Flag antibody was attempted; however, the results were not
satisfactory (data not shown). These results were possibly due to
the fact that Flag at the N-terminal of the protein was cut off
during the expression or preparation of cell-lysed protein samples
in HSC-3 cells. Therefore, various cathelicidin antibodies were
used to detect the expression of CDEL in HSC-3 cells and it was
found that the expression of CDEL in HSC-3 cells could be
effectively detected with an Abcam anti-cathelicidin antibody (the
epitope is located at 50–100 of the CAMP protein). However, it
should be noted that the results of the present study were only
observed in OSCC HSC-3 cells and the effects may differ in other
cells, such as gingival cells. In addition, the results of the
present study are based upon data obtained in vitro and
examination using signal inhibitors should also be performed in
subsequent studies.
In conclusion, the present study successfully
established HSC-3 cell lines stably expressing either CDEL or CAMP,
and examined the roles and mechanisms of CDEL or CAMP in these
cells. To the best of our knowledge, the present study is the first
to show that CAMP/LL-37 may act as a tumour suppressor in OSCC
cells, and the underlying mechanism may include the induction of
caspase-3 dependent apoptosis via the P53-Bcl-2/BAX signalling
pathway. Further studies using other OSCC cells and in vivo
studies will undoubtedly deepen our understanding of its roles and
mechanisms in the initiation and progression of OSCC.
Acknowledgements
Not applicable.
Funding
This study was supported by grants from the National
Natural Science Foundation of China (grant no. 81760490) and
Natural Science Foundation of Guangxi (grant no.
2017GXNSFAA198239).
Availability of data and materials
The datasets used and/or analysed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
XC, SJ, JS, YG and XZo made substantial
contributions to the conception and design of the study. XZh and XW
contributed to data analysis and interpretation. All authors read
and approved the final manuscript.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Alagarasu K, Patil PS, Shil P, Seervi M,
Kakade MB, Tillu H and Salunke A: In-vitro effect of human
cathelicidin antimicrobial peptide LL-37 on dengue virus type 2.
Peptides. 92:23–30. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Majewski K, Żelechowska P and
Brzezińska-Błaszczyk E: Circulating cathelicidin LL-37 in adult
patients with pulmonary infectious diseases. Clin Invest Med.
40:E34–E39. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Tsai PW, Yang CY, Chang HT and Lan CY:
Human antimicrobial peptide LL-37 inhibits adhesion of Candida
albicans by interacting with yeast cell-wall carbohydrates.
PLoS One. 6:e177552011. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Polcyn-Adamczak M and Niemir ZI:
Cathelicidin - Its structure, function and the role in autoimmune
diseases. Adv Cell Biol. 4:83–96. 2014. View Article : Google Scholar
|
|
5
|
Kahlenberg JM and Kaplan MJ: Little
peptide, big effects: The role of LL-37 in inflammation and
autoimmune disease. J Immunol. 191:4895–4901. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Piktel E, Niemirowicz K, Wnorowska U,
Wątek M, Wollny T, Głuszek K, Góźdź S, Levental I and Bucki R: The
role of cathelicidin LL-37 in cancer development. Arch Immunol Ther
Exp (Warsz). 64:33–46. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Wu WK, Wang G, Coffelt SB, Betancourt AM,
Lee CW, Fan D, Wu K, Yu J, Sung JJ and Cho CH: Emerging roles of
the host defense peptide LL-37 in human cancer and its potential
therapeutic applications. Int J Cancer. 127:1741–1747. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Kuroda K, Okumura K, Isogai H and Isogai
E: The human cathelicidin antimicrobial peptide LL-37 and mimics
are potential anticancer drugs. Front Oncol. 5:1442015. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Chen X, Zou X, Qi G, Tang Y, Guo Y, Si J
and Liang L: Roles and mechanism of human cathelicidin LL-37 in
cancer. Cell Physiol Biochem. 47:1060–1073. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Carmona FJ, Montemurro F, Kannan S, Rossi
V, Verma C, Baselga J and Scaltriti M: AKT signaling in
ERBB2-amplified breast cancer. Pharmacol Ther. 158:63–70. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Li D, Wang X, Wu JL, Quan WQ, Ma L, Yang
F, Wu KY and Wan HY: Tumor-produced versican V1 enhances
hCAP18/LL-37 expression in macrophages through activation of TLR2
and vitamin D3 signaling to promote ovarian cancer progression in
vitro. PLoS One. 8:e566162013. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
von Haussen J, Koczulla R, Shaykhiev R,
Herr C, Pinkenburg O, Reimer D, Wiewrodt R, Biesterfeld S, Aigner
A, Czubayko F, et al: The host defence peptide LL-37/hCAP-18 is a
growth factor for lung cancer cells. Lung Cancer. 59:12–23. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Hensel JA, Chanda D, Kumar S, Sawant A,
Grizzle WE, Siegal GP and Ponnazhagan S: LL-37 as a therapeutic
target for late stage prostate cancer. Prostate. 71:659–670. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Sainz B Jr, Alcala S, Garcia E,
Sanchez-Ripoll Y, Azevedo MM, Cioffi M, Tatari M, Miranda-Lorenzo
I, Hidalgo M, Gomez-Lopez G, et al: Microenvironmental
hCAP-18/LL-37 promotes pancreatic ductal adenocarcinoma by
activating its cancer stem cell compartment. Gut. 64:1921–1935.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Muñoz M, Craske M, Severino P, de Lima TM,
Labhart P, Chammas R, Velasco IT, Machado MC, Egan B, Nakaya HI, et
al: Antimicrobial peptide LL-37 participates in the transcriptional
regulation of melanoma cells. J Cancer. 7:2341–2345. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Wang W, Zheng Y, Jia J, Li C, Duan Q, Li
R, Wang X, Shao Y, Chen C and Yan H: Antimicrobial peptide LL-37
promotes the viability and invasion of skin squamous cell carcinoma
by upregulating YB-1. Exp Ther Med. 14:499–506. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
17. Wu WK, Sung JJ, To KF, Yu L, Li HT, Li
ZJ, Chu KM, Yu J and Cho CH: The host defense peptide LL-37
activates the tumor-suppressing bone morphogenetic protein
signaling via inhibition of proteasome in gastric cancer cells. J
Cell Physiol. 223:178–186. 2010.PubMed/NCBI
|
|
18
|
Ren SX, Cheng AS, To KF, Tong JH, Li MS,
Shen J, Wong CC, Zhang L, Chan RL, Wang XJ, et al: Host immune
defense peptide LL-37 activates caspase-independent apoptosis and
suppresses colon cancer. Cancer Res. 72:6512–6523. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Kuroda K, Fukuda T, Krstic-Demonacos M,
Demonacos C, Okumura K, Isogai H, Hayashi M, Saito K and Isogai E:
miR-663a regulates growth of colon cancer cells, after
administration of antimicrobial peptides, by targeting CXCR4-p21
pathway. BMC Cancer. 17:332017. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Mader JS, Mookherjee N, Hancock RE and
Bleackley RC: The human host defense peptide LL-37 induces
apoptosis in a calpain- and apoptosis-inducing factor-dependent
manner involving Bax activity. Mol Cancer Res. 7:689–702. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
21
|
An LL, Ma XT, Yang YH, Lin YM, Song YH and
Wu KF: Marked reduction of LL-37/hCAP-18, an antimicrobial peptide,
in patients with acute myeloid leukemia. Int J Hematol. 81:45–47.
2005. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Okumura K, Itoh A, Isogai E, Hirose K,
Hosokawa Y, Abiko Y, Shibata T, Hirata M and Isogai H: C-terminal
domain of human CAP18 antimicrobial peptide induces apoptosis in
oral squamous cell carcinoma SAS-H1 cells. Cancer Lett.
212:185–194. 2004.PubMed/NCBI
|
|
23
|
Açil Y, Torz K, Gülses A, Wieker H, Gerle
M, Purcz N, Will OM, Eduard Meyer J and Wiltfang J: An experimental
study on antitumoral effects of KI-21-3, a synthetic fragment of
antimicrobial peptide LL-37, on oral squamous cell carcinoma. J
Craniomaxillofac Surg. 46:1586–1592. 2018.PubMed/NCBI
|
|
24
|
Chen X, Qi G, Qin M, Zou Y, Zhong K, Tang
Y, Guo Y, Jiang X, Liang L and Zou X: DNA methylation directly
downregulates human cathelicidin antimicrobial peptide gene (CAMP)
promoter activity. Oncotarget. 8:27943–27952. 2017.PubMed/NCBI
|
|
25
|
Väyrynen O, Åström P, Nyberg P, Alahuhta
I, Pirilä E, Vilen ST, Aikio M, Heljasvaara R, Risteli M, Sutinen
M, et al: Matrix metalloproteinase 9 inhibits the motility of
highly aggressive HSC-3 oral squamous cell carcinoma cells. Exp
Cell Res. 376:18–26. 2019.PubMed/NCBI
|
|
26
|
Chang WL, Cheng FC, Wang SP, Chou ST and
Shih Y: Cinnamomum cassia essential oil and its major constituent
cinnamaldehyde induced cell cycle arrest and apoptosis in human
oral squamous cell carcinoma HSC-3 cells. Environ Toxicol.
32:456–468. 2017.PubMed/NCBI
|
|
27
|
Araújo T, Khayat A, Quintana L, Calcagno
D, Mourão R, Modesto A, Paiva J, Lima A, Moreira F, Oliveira E, et
al: Piwi like RNA-mediated gene silencing 1 gene as a possible
major player in gastric cancer. World J Gastroenterol.
24:5338–5350. 2018.PubMed/NCBI
|
|
28
|
Tang B, Qi G, Tang F, Yuan S, Wang Z,
Liang X, Li B, Yu S, Liu J, Huang Q, et al: Aberrant JMJD3
expression upregulates slug to promote migration, invasion, and
stem cell-like behaviors in hepatocellular carcinoma. Cancer Res.
76:6520–6532. 2016.PubMed/NCBI
|
|
29
|
Qi G, Kudo Y, Tang B, Liu T, Jin S, Liu J,
Zuo X, Mi S, Shao W, Ma X, et al: PARP6 acts as a tumor suppressor
via downregulating Survivin expression in colorectal cancer.
Oncotarget. 7:18812–18824. 2016.PubMed/NCBI
|
|
30
|
Zhao JG, Wang JF, Feng JF, Jin XY and Ye
WL: HHIP overexpression inhibits the proliferation, migration and
invasion of non-small cell lung cancer. PLoS One.
14:e02257552019.PubMed/NCBI
|
|
31
|
Duan Q, Xiao Y, Zhu L, Liu Z, Mao X, Zhou
Z, Liao C, Cai J, Huang F, Liu Z, et al: BET bromodomain is a novel
regulator of TAZ and its activity. Biochim Biophys Acta.
1859:1527–1537. 2016.PubMed/NCBI
|
|
32
|
Mortazavi A, Williams BA, McCue K,
Schaeffer L and Wold B: Mapping and quantifying mammalian
transcriptomes by RNA-Seq. Nat Methods. 5:621–628. 2008.PubMed/NCBI
|
|
33
|
Anders S and Huber W: Differential
expression analysis for sequence count data. Genome Biol.
11:R1062010.PubMed/NCBI
|
|
34
|
Vonderheide RH: Tumor-promoting
inflammatory networks in pancreatic neoplasia: Another reason to
loathe Kras. Cancer Cell. 25:553–554. 2014.PubMed/NCBI
|
|
35
|
Kim YK, Shin JS and Nahm MH: NOD-like
receptors in infection, immunity, and diseases. Yonsei Med J.
57:5–14. 2016.PubMed/NCBI
|
|
36
|
Tuomela JM, Sandholm JA, Kaakinen M,
Hayden KL, Haapasaari KM, Jukkola-Vuorinen A, Kauppila JH,
Lehenkari PP, Harris KW, Graves DE, et al: Telomeric
G-quadruplex-forming DNA fragments induce TLR9-mediated and
LL-37-regulated invasion in breast cancer cells in vitro. Breast
Cancer Res Treat. 155:261–271. 2016.PubMed/NCBI
|
|
37
|
Cheng M, Ho S, Yoo JH, Tran DH, Bakirtzi
K, Su B, Tran DH, Kubota Y, Ichikawa R and Koon HW: Cathelicidin
suppresses colon cancer development by inhibition of cancer
associated fibroblasts. Clin Exp Gastroenterol. 8:13–29.
2014.PubMed/NCBI
|
|
38
|
Martini M, De Santis MC, Braccini L,
Gulluni F and Hirsch E: PI3K/AKT signaling pathway and cancer: An
updated review. Ann Med. 46:372–383. 2014.PubMed/NCBI
|
|
39
|
Sun Y, Liu WZ, Liu T, Feng X, Yang N and
Zhou HF: Signaling pathway of MAPK/ERK in cell proliferation,
differentiation, migration, senescence and apoptosis. J Recept
Signal Transduct Res. 35:600–604. 2015.PubMed/NCBI
|