Introduction
Human papillomavirus (HPV) is the aetiological agent
of cervical cancer (CC) (1). HPV16
is the most prevalent genotype and is responsible for greater than
50% of CC cases worldwide (2,3). The
oncogenic potential of HPV is attributed to the following three
viral proteins: E5, E6 and E7. E6 and E7, the best characterized
viral proteins, promote cell transformation by several mechanisms;
for example, they destabilize and induce the degradation of the
tumour suppressor proteins p53 and pRB, respectively (4). Meanwhile, E5, a transmembrane protein
present in the Golgi apparatus, endoplasmic reticulum and nuclear
envelope (5), has several roles in
cell transformation. For example, E5 can force cells through the
cell cycle and promote the evasion of the immune response (6). Additionally, E5 can modify gene
expression; HPV16 E5 induces the expression of prostaglandin E2
receptor by stimulating the binding of the cAMP-response element
binding protein (CREB) to its promoter, which activates mitogenic
activated protein kinase (MAPK) and increases C-FOS and
C-JUN transcription (7,8).
Moreover, E5 increases the expression of genes related to cell
adhesion, cell motility, and mitogenic signalling, suggesting that
the protein plays an important role in the events associated with
cellular transformation (9).
Altered glycosylation is another characteristic of
cancer cells (10), and it can be
caused by the altered expression of different glycosyltransferases
that can lead to changes in glycan structures. Increased mRNA
levels of some sialyltransferases have been reported in different
cancer types (11). Specifically,
CC and premalignant lesions display increased mRNA levels of the
sialyltransferases ST3GAL3 and ST6GAL1 (12,13),
which are related to increased expression of sialic acid (14,15)
and of sialylated antigens such as sTn and sLe(x) (16–18).
Studies in the HeLa cell line show that E6/E7 HPV18 oncogene
knockdown modified glycogene expression, some of which participate
in the synthesis of O-glycans such as sTn (19). These results suggest that viral
infection could modify glycogene expression and the glycosylation
of the cervical epithelium.
The objective of this work was to identify
glycogenes that displayed modified expression patterns in the
presence of the HPV16 E5 oncoprotein. The results showed that the
HPV16 E5 oncoprotein could increase the expression of the
sialyltransferases ST3GAL3 and ST6GAL1, which have
been reported to be altered in premalignant and malignant cervical
tissues. The network interaction constructed with the altered
glycogenes in the presence of E5 showed that not only the glycan
sialylation but also some glycan structures, such keratan sulphate
and glycosaminoglycans, could also be altered.
Materials and methods
Cell culture
The HaCaT cell line from human skin keratinocytes
stably transfected with the HPV16 E5 oncogene (HaCaT-E5) (20) or the vector pMSG (HaCaT-pMSG)
(kindly donated by Dr. A. Alonso from German Cancer Research
Centre, University of Heidelberg, Germany) was cultured and
maintained in Dulbecco's modified Eagle's medium (DMEM) containing
Earle's salts and L-glutamine (DMEM; Sigma-Aldrich; Merck KGaA) and
supplemented with 10% foetal bovine serum (Biowest), 100 µg/ml
streptomycin (Sigma-Aldrich; Merck KGaA). The CasKi cell line from
a squamous CC (kindly donated by Dr A. Aguilar-Lemarroy from Centro
de Investigación Biomédica de Occidente, IMSS) was cultured and
maintained in RPMI-1640 medium (Sigma-Aldrich; Merck KGaA)
containing L-glutamine and supplemented with 10% foetal bovine
serum, and 100 µg/ml streptomycin (Sigma-Aldrich; Merck KGaA).
Cells were maintained at 37°C with an atmosphere of 5%
CO2. The culture medium was replaced every two days.
Sub-confluent cells were harvested using a mixture of trypsin
(0.025%) and EDTA (0.02%; Sigma-Aldrich; Merck KGaA) and were
washed with phosphate-buffered saline.
Microarray expression assay
The microarray expression assay was performed at the
Cellular Physiology Institute of UNAM. The microarray contained
10,000 gene-specific oligonucleotide probes representing the
best-annotated genes from human. For the probe preparation, total
RNA from HaCaT/E5 and HaCaT/pGSM monocultures was obtained with the
ReliaPrep™ RNA Cell Miniprep System (Promega Corporation) and 10 µg
of each RNA was used for cDNA synthesis incorporating dUTP-Alexa555
or dUTP-Alexa647 and employing the First-Strand cDNA labelling kit
(Invitrogen; Thermo Fisher Scientific, Inc.). Acquisition and
quantification of the array images were performed in GenePix 4100A
with its accompanying software GenePix from Molecular Devices.
Microarray data analysis was performed with the free software
GenArise developed at the Computing Unit of Cellular Physiology
Institute of UNAM (http://www.ifc.unam.mx/genarise/). The software
identifies differentially expressed genes by calculating an
intensity-dependent z-score. The elements with a z-score >2
standard deviations would be the significantly differentially
expressed genes. The analysed data were submitted to the NCBI Gene
Expression Omnibus (access no. GSE118776).
Expression analysis of E5 and
glycogenes
Total RNA from CasKi, HaCaT-E5 and HaCaT-pMSG
monocultures was obtained with the NucleoSpin II RNA kit
(Macherey-Nagel).
To determine the amplification efficiencies of E5,
ST3GAL3 and ST6GAL1, standard curves were performed
with the following RNA concentrations: 10, 1, 0.1, 0.01, and 0.001
ng/µl. HPRT was used as an endogenous gene. Based on these
curves we determined the better concentration to perform the
relative quantification assays, considering that these genes have
different expression levels. cDNA was synthesized using random
primers and the RevertAid First-Strand cDNA Synthesis kit (Thermo
Fisher Scientific, Inc.). qPCR reactions were performed in a final
volume of 10 µl with the following components: 5 µl of 2X Maxima
SYBR-Green/Rox qPCR Master Mix (Thermo Fisher Scientific, Inc.) and
0.5 µl of 10 mM forward and reverse primers (Table I). The reactions were performed
with a StepOne Real-Time PCR System (Applied Biosystems; Thermo
Fisher Scientific, Inc.), and the cycling conditions were as
follows: 95°C for 10 min, followed by 40 cycles of 95°C for 30 sec,
an annealing temperature for 30 sec, and 70°C for 30 sec. For HPV16
E5, the Tms for HPV16 E5, ST3GAL3 and ST6GAL1 were
55°C, 57°C and 60°C, respectively.
 | Table I.Sequences of the oligonucleotides
used in the reverse transcription-quantitative PCR assays. |
Table I.
Sequences of the oligonucleotides
used in the reverse transcription-quantitative PCR assays.
| Primer | Forward (5–3) | Reverse (5–3) | Length (bp)
Product |
|---|
| HPRT |
CCTGGCGTCGTGATTAGTGATGAT |
CGAGCAAGACGTTCAGTCCTGTC | 150 |
| HPV16 E5 |
CGCTGCTTTTGTCTGTGTCT |
GCGTGCATGTGTATGTATTAAAAA | 146 |
| ST3GAL3 |
CATGTGAAGATGGGACTCTTGG |
CCTCCCACTGGAGTAAGTGTAG | 118 |
| ST6GAL1 |
TATCGTAAGCTGCACCCCAATC |
TTAGCAGTGAATGGTCCGGAAG | 372 |
Relative quantification was performed using the
comparative CT method as follows: 2−ΔΔCT. The qPCR
reaction was performed on a StepOne Real-Time PCR System (Applied
Biosystems; Thermo Fisher Scientific, Inc.). The final 10 µl
reaction volume included 1 µl of cDNA template (E5 VPH16-0.5 ng/µl;
ST3GAL3−10 ng/µl, or ST6GAL1−3 ng/µl), 5 µl of 2X
Maxima SYBR-Green/Rox qPCR Master Mix (Thermo Fisher Scientific,
Inc.), 0.5 µl of forward and reverse primers (0.5 µM final
concentration) and 3 µl of RNase free water. qPCR was performed
under with following conditions: 95°C 10 min, followed 40 cycles of
95°C for 30 sec, the annealing temperature for 30 sec (60°C for
ST6GAL1 and 57°C for ST3GAL3) and 70°C for 30 sec. The gene
transcript levels were analysed and normalised to HPRT
expression.
Identification of glycogenes
For the analysis of the glycogenes in the microarray
displaying altered expression, we considered 336 glycogenes
reported to date using the GlycoGene database (http://riodb.ibase.aist.go.jp/rcmg/ggdb/), the
Consortium for Functional Glycomics-CAZy database (http://www.cazy.org/CAZY/) and the published reports
on glycogenes not included in the databases, including
DPY19L1 (21) and
MANBAL (22). We identified
glycogenes displaying altered expression in the microarray as those
with a z-score >2.
Protein-protein interaction
network
The downregulated or upregulated set of glycogenes
in the HaCaT-E5 cells were submitted separately, to the STRING
database (http://string-db.org/). The following
parameter were applied for the analysis: text mining, experiments,
databases, co-expression, neighbourhood, gene fusion and
co-occurrence as interaction sources, no more than 5 interactor,
minimum interaction score of 0.9 as confidence level, and a
protein-protein enrichment P-value at least ≤0.05 and FDR at least
<0.05.
Statistical analysis
Statistical analysis of the qPCR results was
performed using the GraphPad program. The Students t-test was
performed. A P-value <0.05 was considered statistically
significant.
Results
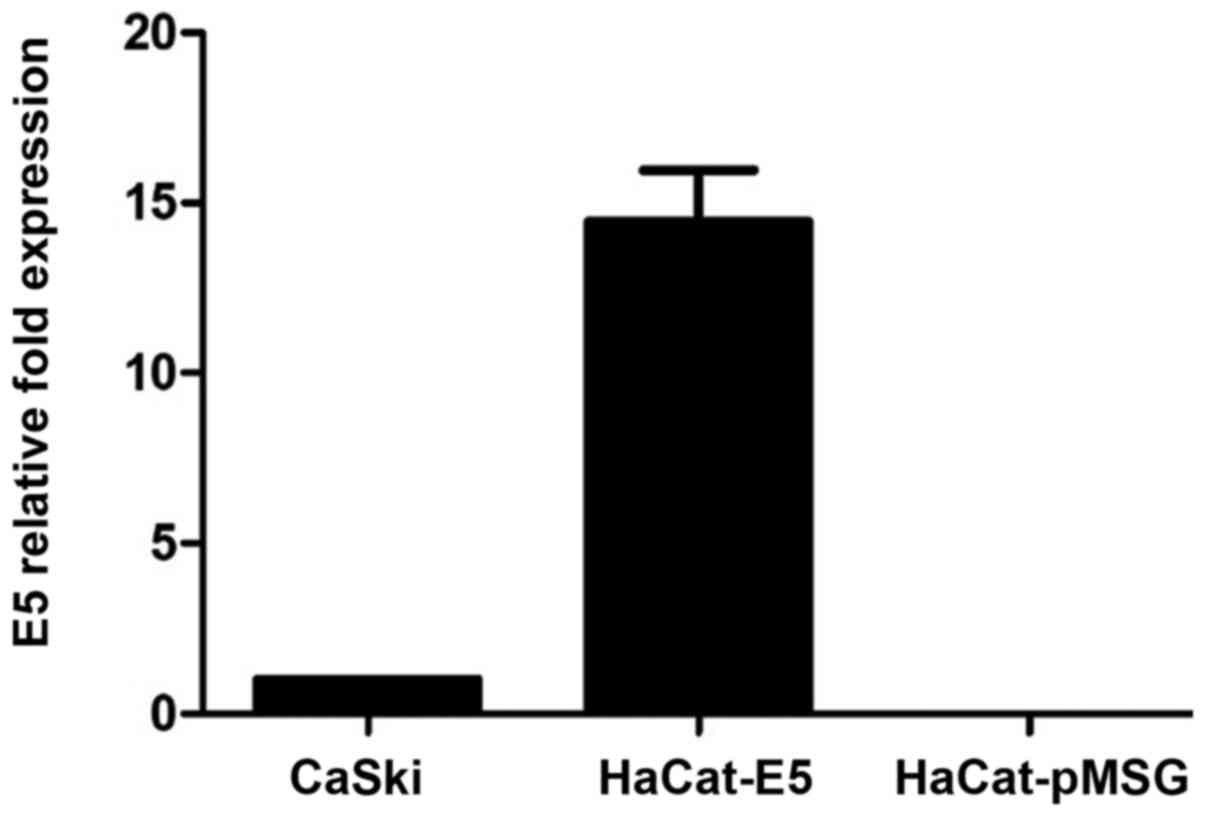
E5 oncogene expression by RT-qPCR
RT-qPCR was performed to quantify E5 mRNA expression
levels from HaCaT-E5 cells, as a positive control it was used mRNA
of CasKi cells (a cell line that contains HPV16-integrated genomes
and expresses E5) and as a negative control was used mRNA of
HaCaT-pMSG. Fig. 1 shows the
relative expression of E5, showing that E5 expression is greater in
HaCaT-E5 than in CasKi cells.
Glycogene expression altered by the E5
oncoprotein
From a total of 336 glycogenes reported to date, we
searched those altered in the HaCaT-E5 microarray with respect to
HaCaT-pGSM. We identified four upregulated glycogenes, including
ST3GAL3, CHST2 and MANBA with a z-score >2 and
ST6GAL1 with a z-score of 1.8. The latter was included
because of its importance in CC. We also identified four
downregulated glycogenes, including GALNT11, NDST2, UGT2B15
and UGT1A10 with a z-score <2. The microarray data
analysed herein are included in the NCBI Gene Expression Omnibus
Database (accession no. GSE118776) and in the article text.
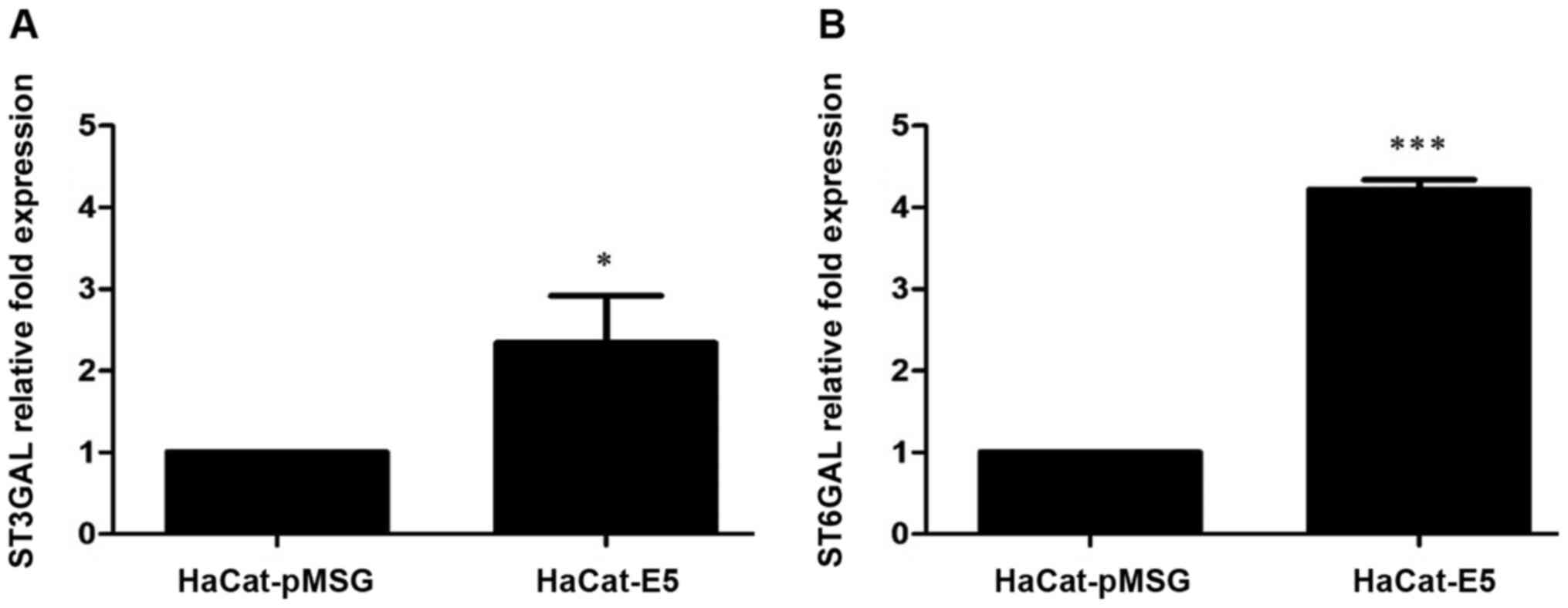
E5 increased the expression of the
sialyltransferases ST3GAL3 and ST6GAL1
Because the microarray results showed that the
presence of E5 can lead to an increase in sialyltransferases
expression, we evaluated the mRNA levels of ST3GAL3 and
ST6GAL1 by RT-qPCR in HaCaT-E5 and HaCaT-pMSG cells.
ST3GAL3 and ST6GAL1 mRNA levels were increased in
HaCaT-E5 cells (Fig. 2).
We next analysed literature data for E5-modified
glycogenes that also have been previously reported as altered in
cancerous tissues [Tables II and
III; (11,12,22–45)]. First, we analysed the upregulated
genes. As previously described, ST3GAL3 and ST6GAL1
were increased in CC and premalignant lesions; in contrast,
CHST2 and MANBA have not been reported in CC, but
display altered expression in other cancer types. The results
showed that the four glycogenes are aberrantly expressed in several
types of cancer and have clinical relevance (Table II).
| Table II.Upregulated glycogenes in HaCaT-E5
cells. |
Table II.
Upregulated glycogenes in HaCaT-E5
cells.
|
|
| Alteration of
mRNA/enzyme in cancer |
|
|---|
|
|
|
|
|
|---|
| Gene/enzyme | Enzyme
function | mRNA | Enzyme | Clinical
relevance |
|---|
| ST3GAL3/ST3
β-galactoside α2,3-sialyltransferase 3 | Transfers sialic
acid from CMP-sialic acid to the structure Galβ1→3/4GlcNAc- and is
involved in the synthesis of sialyl Lewis(a) (NeuAcα2→3Galb1→3
(Fucα1→4) GlcNAc-). | Upregulation in
cervical cancer (12), breast
cancer (23), extrahepatic bile
duct carcinoma (24), colorectal
carcinoma (25), glioblastoma,
renal clear cell carcinoma and lung squamous carcinoma (26). Downregulation in breast invasive
carcinoma, colon adenocarcinoma (26) and ovarian cancers (11). | High levels of
enzyme activity in the tumor tissue correlated with secondary local
tumor recurrence in gastric cancer (27). | In cervical
carcinoma the overexpression of mRNA is associated with lymph node
metastases (12). In breast
cancer, high expression of mRNA is associated with poor prognosis
(23). |
| ST6GAL1/ST6
β-galactoside α-2,6-sialyltransferase 1 | Transfers sialic
acid from CMP-sialic acid to the Galβ1→4GlcNAc structure on
glycoproteins. | Upregulation in
squamous cell carcinoma (12),
breast cancer (23), extrahepatic
bile duct carcinoma (24), ovarian
cancer (11), and colorectal
carcinoma and non-metastasic colorectal tumors (28). | Increased enzyme
expression in colon tumors (28),
ovarian and pancreatic carcinomas (29,30).
Increased enzyme activity in gastric and colorectal cancer
(27,31). | In breast cancer,
high expression of mRNA is associated with poor prognosis (23). In pancreatic ductal adenocarcinoma
promotes chemoresistance to gemcitabine (32). |
|
CHST2/carbohydrate sulfotransferase
2 | Sulfotransferase
that utilizes 3-phospho-5-adenylyl sulfate as donor to catalyze the
transfer of sulfate to position 6 of non-reducing GlcNAc within
keratan-like structures on N-glycans and mucin-associated
glycans. | Upregulation in
breast cancer (33), osteosarcoma
(34) and esophageal cancer
(35). Increased enzyme in uterine
cervical and corpus cancer (36). | Increased enzyme
expression in metastasic osteosarcoma (34). | In osteosarcoma,
weak protein expression is associated with improved survival
(34). |
|
MANBA/β-mannosidase | Exoglycosidase that
cleaves the | Upregulation in
dysplastic esophageal | No reports | Candidate for
molecular |
|
| single β-linked
mannose residue | tissues and
esophageal |
| target for early
detection |
|
| from the
non-reducing end of N-glycoproteins oligosaccharides. | squamous cell
carcinoma (37). |
| of esophageal
cancer (37). |
| Table III.Downregulated glycogenes in HaCaT-E5
cells. |
Table III.
Downregulated glycogenes in HaCaT-E5
cells.
|
|
| Alteration of
mRNA/enzyme in cancer |
|
|---|
|
|
|
|
|
|---|
| Gene/enzyme | Enzyme
function | mRNA | Enzyme | Clinical
relevance |
|---|
|
UGT2B15/UDP | An enzyme of the
glucuronidation | Upregulation in
castration | Upregulation of
expression | In gastric cancer,
patients |
|
glucuronosyltransferase | pathway, involved
in the metabolism | resistant prostate
cancer (38). | in gastric cancer
(39). | with higher UGT2B15
mRNA |
| family 2 member
B15 | and elimination of
toxic compounds. | Upregulation in
gastric | Low level of
expression in aggressive | expression have
poor |
|
| Serves a role in
the regulation of estrogens and androgens. | cancer (39). | prostate tumors and
undetectable expression in prostate cancer with lymph node
metastasis (40). | prognosis (39). |
|
GALNT11/polypeptide | Catalyzes the
initiation of protein | Upregulation in
chronic | No reports. | GALNT11 expression
is |
|
N-acetylgalactosaminyltransferase 11 | O-linked
glycosylation and | lymphocytic
leukemia (41). |
| associated with
prognosis |
|
| is involved in
left/right asymmetry by mediating O-glycosylation of
NOTCH1. |
|
| of chronic
lymphocytic leukemia (41). |
|
NDST2/N-deacetylase | Enzyme with dual
functions: | Moderately
upregulated | No reports. | No reports. |
| and
N-sulfotransferase 2 | Participates in
processing glucosamine | in hepatocellular
cancer (42). |
|
|
|
| and heparin
polymers, including | Decreased levels in
II, III |
|
|
|
| N-deacetylation and
N-sulfation. | and IV stages of
glioma (43). |
|
|
|
UGT1A10/UDP | An enzyme of the
glucuronidation | Upregulated in
stomach cancer (44). | Downregulated in
breast | No reports. |
|
glucuronosyltransferase | pathway that
transforms small | Downregulated in
breast cancer (45). | cancer (45). |
|
| family 1 member
A10 | lipophilic
molecules, such as steroids, bilirubin, hormones and drugs, into
water-soluble, excretable metabolites. |
|
|
|
We next compared the downregulated glycogenes in
HaCaT-E5 cells with those reported in cancerous tissues. The four
glycogenes have been reported in cancer. GALNT11 is
overexpressed in chronic lymphocytic leukaemia (CLL), and
UGT2B15 is downregulated in prostate cancer, but none of the
downregulated glycogenes have been previously reported to be
altered in CC (Table III).
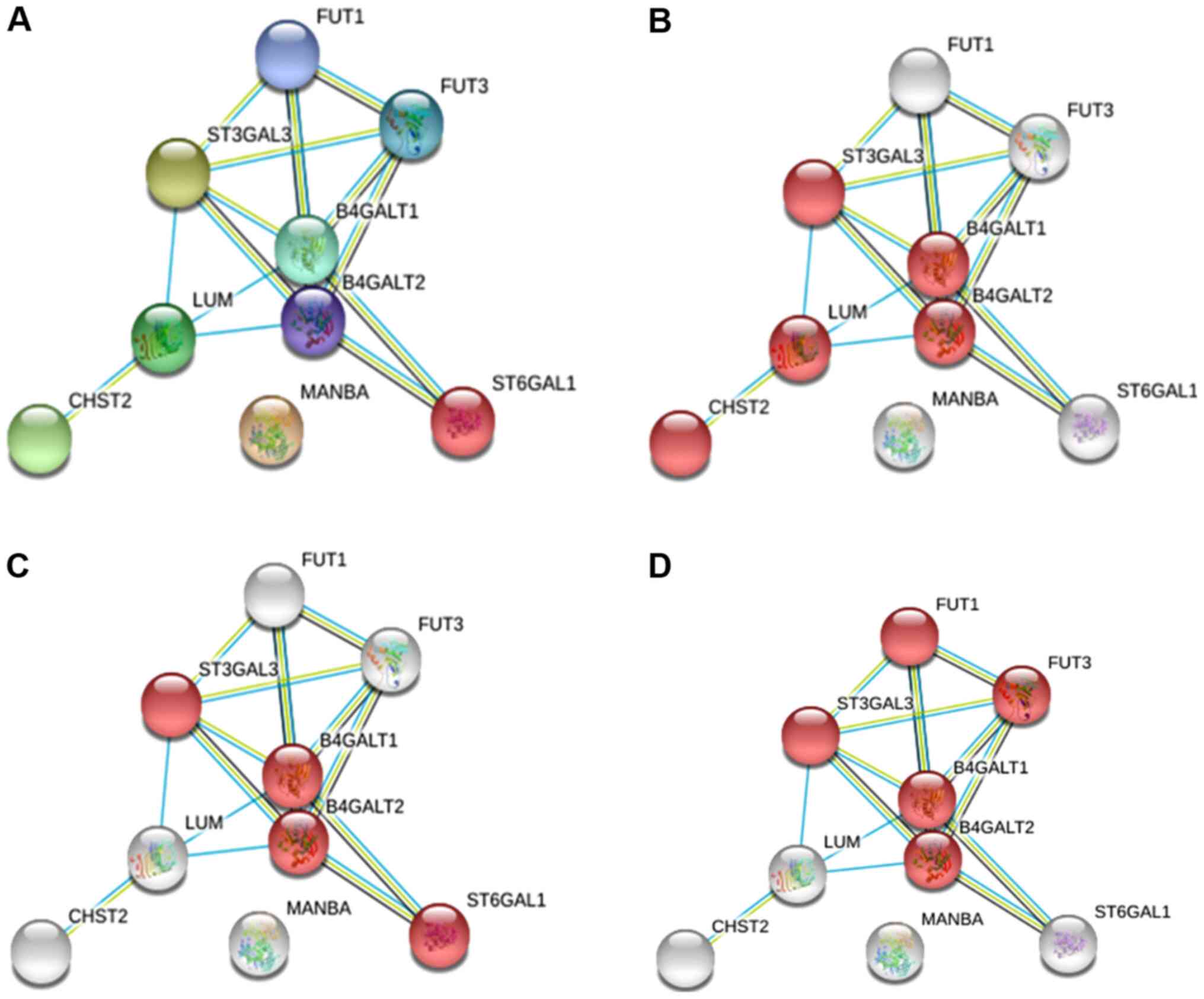
E5 and glycosylation pathways
To identify possible functional associations among
the enzymes identified as altered glycogenes under E5 regulation,
we analysed the data with STRING software to generate predicted
protein-protein interactions with a higher confidence level (0.9).
For the analysis, we considered the altered glycogenes and five
additional proteins with the goal of identifying possible
glycosylation pathways.
For the upregulated glycogenes (ST3GAL3, CHST2,
MANBA and ST6GAL1), the results displayed an interacting
network with eight proteins where MANBA did not interact
with any protein but participates in N-glycosylation
(Fig. 3A). Specifically, we
identified the keratan sulfate biosynthesis pathway, which includes
CHST2 and ST3GAL3 (Fig.
3B); the N-glycosylation pathway, in which both the
sialyltransferase genes are involved in the sialylation of
N-glycans (Fig. 3C); and
the glycosphingolipid biosynthesis pathway, where ST3GAL3
participates with fucosyltransferases and galactosyltransferases
(Fig. 3D).
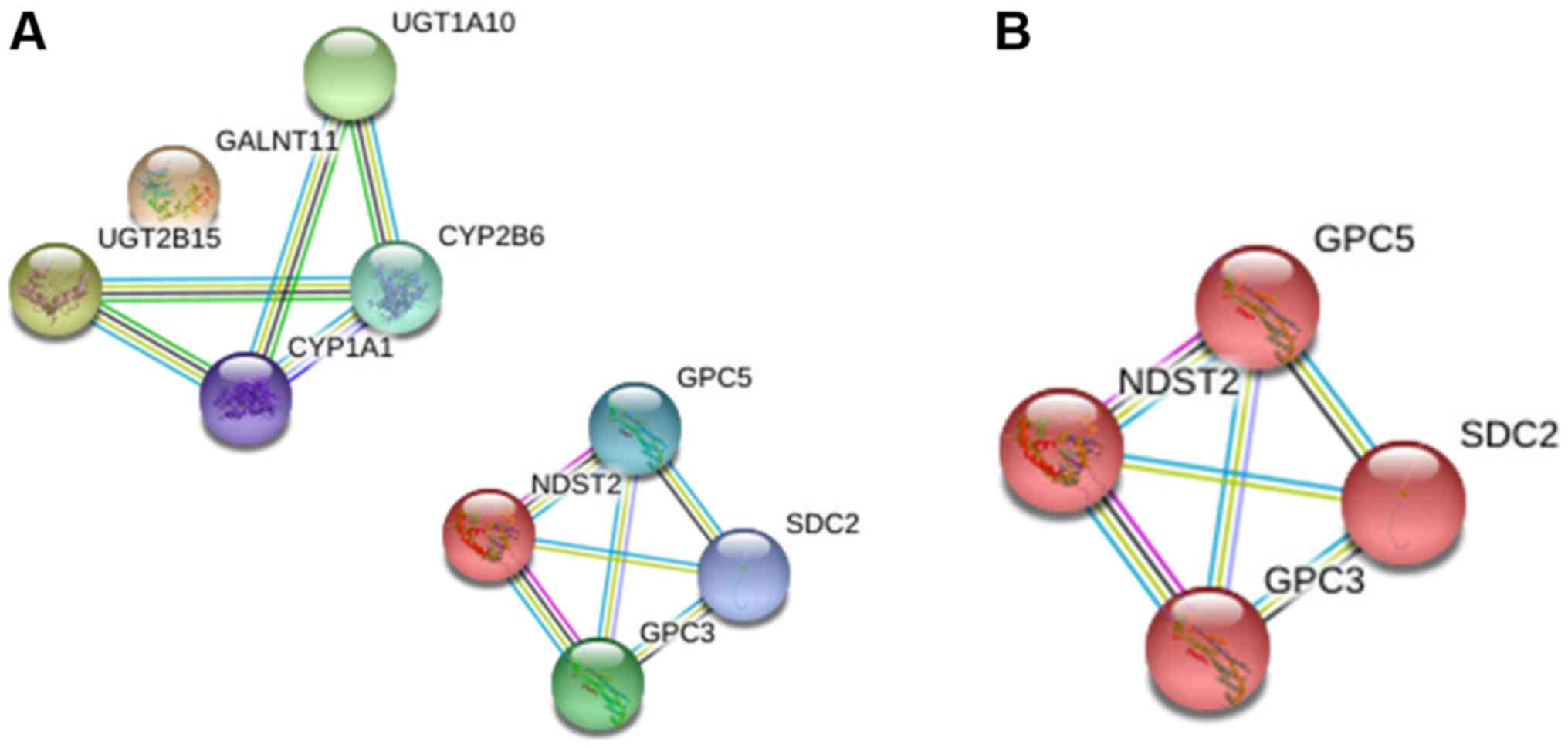
For the downregulated glycogenes (UGT2B15,
GALNT11, NDST2 and UGT1A10), the interaction analysis
showed two different networks, among which GALNT11 was
excluded from the first network and NDST2 establishes a
separate network (Fig. 4A). The
analysis showed that the latter is involved in glycosaminoglycan
biosynthesis (Fig. 4B).
Discussion
Aberrant glycosylation is a characteristic of tumour
cells. Changes in glycan structures in cancer have been related
with altered glycogene expression (46). Increased sialylation is one of the
most frequent alterations in cancer (47). Increased activity of
sialyltransferases, enzymes that transfer sialic acid to
glycoconjugates, has been reported during tumour transformation
(11,48). Increased α2,3 and α2,6 sialic acid
levels have been reported in premalignant lesions from the cervical
epithelium (14). Increased
expression of the sialylated antigens sLe(a) and sLe(x) have also
been reported in CC and premalignant lesions (17,18).
These findings could be the result of enhanced ST3GAL3 and
ST6GAL1 (sialyltransferase genes) mRNA levels, which have
been previously reported during cervical transformation (12,13).
HPV is the aetiological agent of CC, and the HPV genome encodes
three oncoproteins, E5, E6 and E7 (4); however, their roles in altering
glycogene expression have been poorly investigated. Our research
group is interested in the role of HPV infection and its
relationship with glycogene expression changes in the cervical
epithelium. Our group recently reported that the oncoproteins E6
and E7 from HPV18 modify the expression of some glycogenes, some of
which participate in the glycosylation of the Notch receptor and
O-glycosylation type mucin (19). However, similar reports focused on
the E5 oncoprotein do not exist. E5 is a protein expressed during
the early stages of viral infection, and this protein has different
targets in the cell that promote cellular transformation (6,49).
With the aim of determining whether E5 modifies the
expression of some glycogenes, we performed an expression
microarray on the HaCaT cell line stably transfected with the HPV16
E5 oncoprotein. We identified four upregulated glycogenes
(CHST2, MANBA, ST3GAL3 and ST6GAL1) and four
downregulated glycogenes (UGT2B15, GALNT11, NDST2 and
UGT1A10). All these genes have been reported to be altered
in cancer either at the transcript or protein level (Tables II and III).
Increased CHST2 mRNA levels have been
reported in osteosarcoma and breast and oesophageal cancer
(33–35), while increased protein levels have
been reported in ovarian cancer and CC (36). With regards to the glycogene
MANBA, increased mRNA levels have only been reported in
oesophageal cancer (37).
Interestingly, of the four upregulated glycogenes, two correspond
to the sialyltransferases genes ST3GAL3 and ST6GAL1.
ST3GAL3 expression has been reported as altered in different
cancer types, such as gastric, bile, colon, kidney, lung, breast,
and ovarian cancers and glioblastoma (11,23–27).
Increased ST6GAL1 mRNA and protein expression has been
reported in gastric and biliary cancers (24,27),
colon and colorectal cancers (25,29,31),
and ovarian and pancreatic cancers (11,30).
Here, we showed that the mRNA levels of the ST6GAL1 and
ST3GAL3 genes can be increased by the presence of E5;
interestingly, both mRNA levels are increased in premalignant and
malignant tissues in the cervical epithelium (12,13).
However, whether these phenotypes could be a consequence of HPV
infection remains unclear. Additionally, the upregulation of these
genes agrees with the increased levels of sialic acid reported for
CC and with the increased levels of α2,3 and α2,6 sialic acid in
premalignant lesions in the cervix (14). These results suggest that the
sialyltransferase expression changes that occur in the early stages
of cervical cell transformation could be related to HPV infection
and due to E5, but not E6 or E7, activity (19). Nevertheless, this hypothesis
requires more investigation. With respect to E5 and sialylation, a
previous study found no important changes in the sialylation status
of keratinocytes in the presence of HPV16 E5 (50), however, in this study the authors
analysed the expression of different monosaccharaides and
disaccharides, using a panel of seven lectins, but they did not
perfom glycogene expression analysis. Additionally, as they use an
inducible vector, they incubated the transfected cells with
dexamethasone to increase the expression of E5, so the
glycosylation pattern could be influenced by the effect of this
molecule, as has been reported in different studies where the
dexamethasone increase the sialyltransferases expression (51,52).
Regarding the downregulated genes in the presence of
HPV16 E5, GALNT11 has been reported as diminished in breast
cancer (33) and increased in
chronic lymphocytic leukaemia, and it was proposed to be implicated
in O-glycosylation changes in chronic lymphocytic leukaemia
(CLL) cells (41). NDST2
has demonstrated increased mRNA levels in hepatocellular cancer
(42). It was also interesting to
find expression changes in two UGT genes (UGT2B15 and
UGT1A10) that encode uridine
5′-diphospho-glucuronosyltransferases as these enzymes play
important roles in the biotransformation of drugs, xenobiotics, and
toxic compounds (44). Expression
changes in UGT genes could modify the response to some cancer
drugs. UGT2B15 has been reported as diminished in prostate
cancer (40), and UGT1A10
has been reported as downregulated in breast cancer (45) but upregulated in gastric cancer
(44). These UGT genes have
not been previously reported as altered in CC; thus, it could be
interesting to analyse their expression status and the response of
patients to treatment when their expression is altered.
Additionally, we also identified possible
glycosylation pathways altered in the presence of E5 by analysing
protein-protein interactions. We identified keratan sulfate and
glycosphingolipid synthesis as including the involvement of the
glycogenes ST3GAL3, ST6GAL1 and CHST2. Keratan
sulfate belongs to the family of glycosaminoglycans, which
participates in the regulation of cellular functions in epithelial
and mesenchymal tissues (53).
The glycogenes ST3GAL3 and ST6GAL1 are
involved in sialylation of glycosphingolipid. These glycolipids can
be located in the cellular membrane, participate in cell-cell
interactions and modulate transduction pathways, cell growth,
apoptosis, cell proliferation, endocytosis, cell migration,
senescence and embryogenesis (54,55).
The aberrant expression of glycosphingolipids and the enzymes that
participate in their biosynthesis has been reported in different
cancer types (56). The expression
of the ganglioside GM1 has been reported as being increased in
ectocervix cells expressing VPH16 E5; gangliosides are expressed at
high levels on tumour cells and inhibit cytotoxic T lymphocytes
(56). Our results support these
findings, demonstrating that E5 could be modifying ganglioside
synthesis.
The glycogenes present in the N-glycosylation
pathway are ST3GAL3 and ST6GAL1. Changes in sialic
acid expression have been reported for different cancer types
including CC, as was mention previously (11). Increased expression of sialylated
antigens such as sLe(x) and sLe(a) has been reported in
premalignant lesions and CC, and the enzyme ST3Gal III participates
in the synthesis of both of these antigens (17,18,57).
sLe(x) antigens can be modified by the enzyme sulfotransferase
CHST2 to produce 6-sulfosialil Lewis-x (6-sulfo sLex), which can be
a ligand of L-, P- and E-selectin (58). Moreover, ST6Gal I catalyses the
α2,6 sialylation of N-glycans, and its expression is
increased in different types of cancer (59). ST6Gal I had been implicated in the
hypersialylation of the cell membrane protein β1 integrin in tumour
cells, which leads to increased migration capacity (60). ST6Gal I also participates in the
sialylation of epidermal growth factor receptor (EGFR) (61), which showed increased expression
due to HPV16 E5 (62).
For the downregulated genes, the network
interactions implicated in a glycosylation pathway involved only
one glycogene. Fig. 4 shows a
network of four proteins implicated in glycosaminoglycan
biosynthesis, but only one glycogene, NDST2, was present in
this network. The upregulation of NDST2 has been related to
an increase in heparan sulfate (63).
For the glycogenes downregulated in the presence of
E5, there are no reports focused on CC; thus, it would be important
to evaluate their expression and roles in cervical
transformation.
The results of this study provide important
information related to glycogenes that modify their expression in
the presence of the HPV-16 E5 protein. The expression microarray
generated large amounts of information on the genes that were
altered, but the results must be validated. The study present some
limitations, it is important to confirm that the expression of the
gene is related with the expression level of its corresponding
protein, additionally, the study was performed in a cell line, so
the role of the HPV infection and the expression of the other viral
proteins could be participating in the glycosylation changes, so
the effect of E5 could be affected by other factors. Therefore, the
results must be confirmed with more in vitro experiments,
followed by the evaluation of glycogene expression in different
stages in cervical neoplasias positive for HPV16, as some of the
altered glycogenes have not been previously reported in
premalignant and malignant tissues. Also, it could be of interest
to evaluate if the protein E5 from different viral genotypes, plays
a different role in the glycosylation changes. Studying the effects
of E5 of high and low risks HPV, will certainly provide relevant
additional information of the effects of this viral infection.
The expression of the HPV16 E5 protein in HaCaT
cells increased ST3GAL3 and ST6GAL1 mRNA levels,
suggesting that the E5 protein could participate in the
glycosylation changes found during cervical transformation in the
cervical epithelium infected with HPV, especially those related to
increased α2,3 and α2,6 sialylation, which have been reported as
enhanced in premalignant lesions and CC. Additionally, E5 could
participate in UGT gene expression changes implicated in
treatment response. Changes in UGT gene expression have not
been previously reported; thus, it would be interesting to analyse
their levels in CC samples as well as to examine the correlation of
these changes with responses to treatment.
Acknowledgements
The authors would like to thank Dr Lorena Chávez
González, Dr Simón Guzmán León, Dr José Luis Santillán Torres and
Dr Jorge Ramírez for their technical assistance with microarray
determinations. The authors would also like to thank Mr. Gerardo
Coello, Mr. Gustavo Corral and Ms. Ana Patricia Gómez for their
assistance with the genArise software. All the previously mentioned
individuals are affiliated to the Cellular Physiology Institute of
National Autonomous University of Mexico.
Funding
The current study was supported by the Consejo
Nacional de Ciencia y Tecnología (grant no. SALUD-2017-01-290068).
JRL has a fellowship from the Fundación IMSS (grant no. 8066205).
DCR was supported by a Ph.D. fellowship from CONACYT (grant no.
242745) and IMSS (grant no. 98227564). Funding was also provided by
the Programa Cátedras CONACYT 2016 (grant no. 485) and Fondo Redes
Temáticas de Investigación CONACYT (grant no. 253596).
Availability of data and materials
The datasets generated and analysed during the
current study are available in the repository NCBI Gene Expression
Omnibus database, (accession no. GSE118776; http://www.ncbi.nlm.nih.gov/geo/query/acc.cgi?acc=GSE118776).
Authors' contributions
DCR performed molecular biology experiments, in
silico analysis, data analysis and revised the manuscript. YML
participated in data analysis and critically revised the
manuscript. PMM participated in the in silico and data
analyses, and critically revised the manuscript. AAL analysed the
data and critically revised the manuscript. LFJS participated in
the in silico analysis and critically revised the
manuscript. GSL participated in the molecular biology experiments,
analysed the data and revised the manuscript. JRL designed the
current study, analysed the data and critically revised the
manuscript. VVR conceived and designed the current study,
coordinated the study and draft the manuscript. All the authors
read and approved the final manuscript.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
zur Hausen H: Papillomaviruses in the
causation of human cancers - a brief historical account. Virology.
384:260–265. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Smith JS, Lindsay L, Hoots B, Keys J,
Franceschi S, Winer R and Clifford GM: Human papillomavirus type
distribution in invasive cervical cancer and high-grade cervical
lesions: A meta-analysis update. Int J Cancer. 121:621–632. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Aguilar-Lemarroy A, Vallejo-Ruiz V,
Cortés-Gutiérrez EI, Salgado-Bernabé ME, Ramos-González NP,
Ortega-Cervantes L, Arias-Flores R, Medina-Díaz IM, Hernández-Garza
F, Santos-López G, et al IMSS Research Network on HPV, : Human
papillomavirus infections in Mexican women with normal cytology,
precancerous lesions, and cervical cancer: Type-specific prevalence
and HPV coinfections. J Med Virol. 87:871–884. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Münger K and Howley PM: Human
papillomavirus immortalization and transformation functions. Virus
Res. 89:213–228. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Conrad M, Bubb VJ and Schlegel R: The
human papillomavirus type 6 and 16 E5 proteins are
membrane-associated proteins which associate with the 16-kilodalton
pore-forming protein. J Virol. 67:6170–6178. 1993. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Venuti A, Paolini F, Nasir L, Corteggio A,
Roperto S, Campo MS and Borzacchiello G: Papillomavirus E5: The
smallest oncoprotein with many functions. Mol Cancer. 10:1402011.
View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Chen SL, Huang CH, Tsai TC, Lu KY and Tsao
YP: The regulation mechanism of c-jun and junB by human
papillomavirus type 16 E5 oncoprotein. Arch Virol. 141:791–800.
1996. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Chen SL, Lin YK, Li LY, Tsao YP, Lo HY,
Wang WB and Tsai TC: E5 proteins of human papillomavirus types 11
and 16 transactivate the c-fos promoter through the NF1 binding
element. J Virol. 70:8558–8563. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Kivi N, Greco D, Auvinen P and Auvinen E:
Genes involved in cell adhesion, cell motility and mitogenic
signaling are altered due to HPV 16 E5 protein expression.
Oncogene. 27:2532–2541. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Varki A, Kannagi R, Toole B and Stanley P:
Glycosylation Changes in Cancer. Essentials of Glycobiology
[Internet]. 3rd edition. Varki A, Cummings RD, Esko JD, Stanley P,
Hart GW, Aebi M, Darvill AG, Kinoshita T, Packer NH, Prestegard JH,
et al: Cold Spring Harbor Laboratory Press; Cold Spring Harbor, NY:
2017
|
|
11
|
Wang PH: Altered Glycosylation in Cancer:
Sialic Acids and Sialyltransferases. J Cancer Mol. 1:73–81.
2005.
|
|
12
|
Wang PH, Li YF, Juang CM, Lee YR, Chao HT,
Ng HT, Tsai YC and Yuan CC: Expression of sialyltransferase family
members in cervix squamous cell carcinoma correlates with lymph
node metastasis. Gynecol Oncol. 86:45–52. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
López-Morales D, Velázquez-Márquez N,
Valenzuela O, Santos-López G, Reyes-Leyva J and Vallejo-Ruiz V:
Enhanced sialyltransferases transcription in cervical
intraepithelial neoplasia. Invest Clin. 50:45–53. 2009.PubMed/NCBI
|
|
14
|
López-Morales D, Reyes-Leyva J,
Santos-López G, Zenteno E and Vallejo-Ruiz V: Increased expression
of sialic acid in cervical biopsies with squamous intraepithelial
lesions. Diagn Pathol. 5:742010. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Roy A and Chakraborty S: Detection of
cancer cervix by estimation of sialic acid. J Indian Med Assoc.
103:589–590. 2005.PubMed/NCBI
|
|
16
|
Terasawa K, Furumoto H, Kamada M and Aono
T: Expression of Tn and sialyl-Tn antigens in the neoplastic
transformation of uterine cervical epithelial cells. Cancer Res.
56:2229–2232. 1996.PubMed/NCBI
|
|
17
|
Engelstaedter V, Fluegel B, Kunze S, Mayr
D, Friese K, Jeschke U and Bergauer F: Expression of the
carbohydrate tumour marker Sialyl Lewis A, Sialyl Lewis X, Lewis Y
and Thomsen-Friedenreich antigen in normal squamous epithelium of
the uterine cervix, cervical dysplasia and cervical cancer. Histol
Histopathol. 27:507–514. 2012.PubMed/NCBI
|
|
18
|
Velázquez-Márquez N, Santos-López G,
Jiménez-Aranda L, Reyes-Leyva J and Vallejo-Ruiz V: Sialyl Lewis ×
expression in cervical scrapes of premalignant lesions. J Biosci.
37:999–1004. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Aco-Tlachi M, Carreño-López R,
Martínez-Morales PL, Maycotte P, Aguilar-Lemarroy A, Jave-Suárez
LF, Santos-López G, Reyes-Leyva J and Vallejo-Ruiz V: Glycogene
expression profiles based on microarray data from cervical
carcinoma HeLa cells with partially silenced E6 and E7 HPV
oncogenes. Infect Agent Cancer. 13:252018. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Kabsch K and Alonso A: The human
papillomavirus type 16 E5 protein impairs TRAIL- and FasL-mediated
apoptosis in HaCaT cells by different mechanisms. J Virol.
76:12162–12172. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Buettner FF, Ashikov A, Tiemann B, Lehle L
and Bakker H: C. elegans DPY-19 is a C-mannosyltransferase
glycosylating thrombospondin repeats. Mol Cell. 50:295–302. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Milde-Langosch K, Karn T, Schmidt M, zu
Eulenburg C, Oliveira-Ferrer L, Wirtz RM, Schumacher U, Witzel I,
Schütze D and Müller V: Prognostic relevance of
glycosylation-associated genes in breast cancer. Breast Cancer Res
Treat. 145:295–305. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Recchi MA, Hebbar M, Hornez L,
Harduin-Lepers A, Peyrat JP and Delannoy P: Multiplex reverse
transcription polymerase chain reaction assessment of
sialyltransferase expression in human breast cancer. Cancer Res.
58:4066–4070. 1998.PubMed/NCBI
|
|
24
|
Jin XL, Zheng SS, Wang BS and Chen HL:
Correlation of glycosyltransferases mRNA expression in extrahepatic
bile duct carcinoma with clinical pathological characteristics.
Hepatobiliary Pancreat Dis Int. 3:292–295. 2004.PubMed/NCBI
|
|
25
|
Petretti T, Kemmner W, Schulze B and
Schlag PM: Altered mRNA expression of glycosyltransferases in human
colorectal carcinomas and liver metastases. Gut. 46:359–366. 2000.
View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Ashkani J and Naidoo KJ:
Glycosyltransferase gene expression profiles classify cancer types
and propose prognostic subtypes. Sci Rep. 6:264512016. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Gretschel S, Haensch W, Schlag PM and
Kemmner W: Clinical relevance of sialyltransferases ST6GAL-I and
ST3GAL-III in gastric cancer. Oncology. 65:139–145. 2003.
View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Zhang S, Lu J, Xu Z, Zou X, Sun X, Xu Y,
Shan A, Lu J, Yan X, Cui Y, et al: Differential expression of
ST6GAL1 in the tumor progression of colorectal cancer. Biochem
Biophys Res Commun. 486:1090–1096. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Swindall AF, Londoño-Joshi AI, Schultz MJ,
Fineberg N, Buchsbaum DJ and Bellis SL: ST6Gal-I protein expression
is upregulated in human epithelial tumors and correlates with stem
cell markers in normal tissues and colon cancer cell lines. Cancer
Res. 73:2368–2378. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Schultz MJ, Holdbrooks AT, Chakraborty A,
Grizzle WE, Landen CN, Buchsbaum DJ, Conner MG, Arend RC, Yoon KJ,
Klug CA, et al: The tumor-associated glycosyltransferase ST6Gal-I
regulates stem cell transcription factors and confers a cancer stem
cell phenotype. Cancer Res. 76:3978–3988. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Vázquez-Martín C, Gil-Martín E and
Fernández-Briera A: Elevation of ST6Gal I activity in malignant and
transitional tissue in human colorectal cancer. Oncology.
69:436–444. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Chakraborty A, Dorsett KA, Trummell HQ,
Yang ES, Oliver PG, Bonner JA, Buchsbaum DJ and Bellis SL: ST6Gal-I
sialyltransferase promotes chemoresistance in pancreatic ductal
adenocarcinoma by abrogating gemcitabine-mediated DNA damage. J
Biol Chem. 293:984–994. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Potapenko IO, Haakensen VD, Lüders T,
Helland A, Bukholm I, Sørlie T, Kristensen VN, Lingjaerde OC and
Børresen-Dale AL: Glycan gene expression signatures in normal and
malignant breast tissue; possible role in diagnosis and
progression. Mol Oncol. 4:98–118. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Chen X, Yang TT, Qiu XC, Ji ZG, Li CX,
Long H, Zhou Y, Ma BA, Ma Q, Zhang X, et al: Gene expression
profiles of human osteosarcoma cell sublines with different
pulmonary metastatic potentials. Cancer Biol Ther. 11:287–292.
2011. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Warnecke-Eberz U, Metzger R, Hölscher AH,
Drebber U and Bollschweiler E: Diagnostic marker signature for
esophageal cancer from transcriptome analysis. Tumour Biol.
37:6349–6358. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Seko A, Kataoka F, Aoki D, Sakamoto M,
Nakamura T, Hatae M, Yonezawa S and Yamashita K:
N-Acetylglucosamine 6-O-sulfotransferase-2 as a tumor marker for
uterine cervical and corpus cancer. Glycoconj J. 26:1065–1073.
2009. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Sud N, Sharma R, Ray R, Chattopadhyay T
and Ralhan R: Differential expression of beta mannosidase in human
esophageal cancer. Int J Cancer. 112:905–907. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Pfeiffer MJ, Smit FP, Sedelaar JP and
Schalken JA: Steroidogenic enzymes and stem cell markers are
upregulated during androgen deprivation in prostate cancer. Mol
Med. 17:657–664. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Chen X, Li D, Wang N, Yang M, Liao A, Wang
S, Hu G, Zeng B, Yao Y, Liu D, et al: Bioinformatic analysis
suggests that UGT2B15 activates the Hippo YAP signaling pathway
leading to the pathogenesis of gastric cancer. Oncol Rep.
40:1855–1862. 2018.PubMed/NCBI
|
|
40
|
Pâquet S, Fazli L, Grosse L, Verreault M,
Têtu B, Rennie PS, Bélanger A and Barbier O: Differential
expression of the androgen-conjugating UGT2B15 and UGT2B17 enzymes
in prostate tumor cells during cancer progression. J Clin
Endocrinol Metab. 97:E428–E432. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Libisch MG, Casás M, Chiribao M, Moreno P,
Cayota A, Osinaga E, Oppezzo P and Robello C: GALNT11 as a new
molecular marker in chronic lymphocytic leukemia. Gene.
533:270–279. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Tátrai P, Egedi K, Somorácz A, van
Kuppevelt TH, Ten Dam G, Lyon M, Deakin JA, Kiss A, Schaff Z and
Kovalszky I: Quantitative and qualitative alterations of heparan
sulfate in fibrogenic liver diseases and hepatocellular cancer. J
Histochem Cytochem. 58:429–441. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Ushakov VS, Tsidulko AY, de La Bourdonnaye
G, Kazanskaya GM, Volkov AM, Kiselev RS, Kobozev VV, Kostromskaya
DV, Gaytan AS, Krivoshapkin AL, et al: Heparan sulfate biosynthetic
system is inhibited in human glioma due to EXT1/2 and HS6ST1/2
D}down-regulation. Int J Mol Sci. 18:E23012017. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Cengiz B, Yumrutas O, Bozgeyik E, Borazan
E, Igci YZ, Bozgeyik I and Oztuzcu S: Differential expression of
the UGT1A family of genes in stomach cancer tissues. Tumour Biol.
36:5831–5837. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Starlard-Davenport A, Lyn-Cook B and
Radominska-Pandya A: Identification of UDP-glucuronosyltransferase
1A10 in non-malignant and malignant human breast tissues. Steroids.
73:611–620. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Pinho SS and Reis CA: Glycosylation in
cancer: Mechanisms and clinical implications. Nat Rev Cancer.
15:540–555. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Magalhães A, Duarte HO and Reis CA:
Aberrant glycosylation in cancer: A novel molecular mechanism
controlling metastasis. Cancer Cell. 31:733–735. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Harduin-Lepers A, Krzewinski-Recchi MA,
Colomb F, Foulquier F, Groux-Degroote S and Delannoy P:
Sialyltransferases functions in cancers. Front Biosci (Elite Ed).
4:499–515. 2012. View
Article : Google Scholar : PubMed/NCBI
|
|
49
|
Estêvão D, Costa NR, Gil da Costa RM and
Medeiros R: Hallmarks of HPV carcinogenesis: The role of E6, E7 and
E5 oncoproteins in cellular malignancy. Biochim Biophys Acta Gene
Regul Mech. 1862:153–162. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Oetke C, Auvinen E, Pawlita M and Alonso
A: Human papillomavirus type 16 E5 protein localizes to the Golgi
apparatus but does not grossly affect cellular glycosylation. Arch
Virol. 145:2183–2191. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Vandamme V, Pierce A, Verbert A and
Delannoy P: Transcriptional induction of beta-galactoside
alpha-2,6-sialyltransferase in rat fibroblast by dexamethasone. Eur
J Biochem. 211:135–140. 1993. View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Coughlan CM, Burger PG, Berger EG and
Breen KC: The biochemical consequences of alpha2,6(N)
sialyltransferase induction by dexamethasone on sialoglycoprotein
expression in the rat H411e hepatoma cell line. FEBS Lett.
413:389–393. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
53
|
Caterson B and Melrose J: Keratan sulfate,
a complex glycosaminoglycan with unique functional capability.
Glycobiology. 28:182–206. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
54
|
D'Angelo G, Capasso S, Sticco L and Russo
D: Glycosphingolipids: Synthesis and functions. FEBS J.
280:6338–6353. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
55
|
Zhuo D, Li X and Guan F: Biological roles
of aberrantly expressed glycosphingolipids and related enzymes in
human cancer development and progression. Front Physiol. 9:4662018.
View Article : Google Scholar : PubMed/NCBI
|
|
56
|
Suprynowicz FA, Disbrow GL, Krawczyk E,
Simic V, Lantzky K and Schlegel R: HPV-16 E5 oncoprotein
upregulates lipid raft components caveolin-1 and ganglioside GM1 at
the plasma membrane of cervical cells. Oncogene. 27:1071–1078.
2008. View Article : Google Scholar : PubMed/NCBI
|
|
57
|
Harduin-Lepers A, Vallejo-Ruiz V,
Krzewinski-Recchi MA, Samyn-Petit B, Julien S and Delannoy P: The
human sialyltransferase family. Biochimie. 83:727–737. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
58
|
Ohmori K, Kanda K, Mitsuoka C, Kanamori A,
Kurata-Miura K, Sasaki K, Nishi T, Tamatani T and Kannagi R: P- and
E-selectins recognize sialyl 6-sulfo Lewis X, the recently
identified L-selectin ligand. Biochem Biophys Res Commun.
278:90–96. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
59
|
Lu J and Gu J: Significance of
β-galactoside α2,6 sialyltranferase 1 in cancers. Molecules.
20:7509–7527. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
60
|
Seales EC, Jurado GA, Brunson BA,
Wakefield JK, Frost AR and Bellis SL: Hypersialylation of beta1
integrins, observed in colon adenocarcinoma, may contribute to
cancer progression by up-regulating cell motility. Cancer Res.
65:4645–4652. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
61
|
Park JJ, Yi JY, Jin YB, Lee YJ, Lee JS,
Lee YS, Ko YG and Lee M: Sialylation of epidermal growth factor
receptor regulates receptor activity and chemosensitivity to
gefitinib in colon cancer cells. Biochem Pharmacol. 83:849–857.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
62
|
Tomakidi P, Cheng H, Kohl A, Komposch G
and Alonso A: Modulation of the epidermal growth factor receptor by
the human papillomavirus type 16 E5 protein in raft cultures of
human keratinocytes. Eur J Cell Biol. 79:407–412. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
63
|
Deligny A, Dierker T, Dagälv A, Lundequist
A, Eriksson I, Nairn AV, Moremen KW, Merry CLR and Kjellén L: NDST2
(N-Deacetylase/N-Sulfotransferase-2) enzyme regulates heparan
sulfate chain length. J Biol Chem. 291:18600–18607. 2016.
View Article : Google Scholar : PubMed/NCBI
|