Introduction
Patients with cancer, such as leukemia, exhibit bone
marrow suppression, including thrombocytopenia, which is
characterized by subnormal levels of blood platelets following
chemotherapy or radiotherapy (1).
Platelet transfusion (2) and
thrombopoietin (TPO) treatment (3)
are commonly used for the management of thrombocytopenia. However,
aggravation of thrombocytopenia may be induced due to the
subsequent formation of anti-platelet and anti-TPO antibodies
(3,4). Furthermore, repeated platelet
transfusions have been associated with increased risk of blood
infections (5). It has also been
reported that treatment with recombinant human TPO (rhTPO) therapy
results in the formation of TPO antibodies, leading to severe
thrombocytopenia (6,7). Therefore, there is a need for the
development of safer and more effective therapies for
thrombocytopenia.
In ancient China, a variety of Chinese herbal
decoctions have been used to treat blood disorders. For instance,
Danggui Buxue Tang (DBT), a renowned formulation that has been used
for >800 years, consists of two herbs, namely Danggui (extracted
from Radix Angelica Sinensis) and Huangqi (extracted from Radix
Astragali) (8,9). Our previous studies have confirmed the
hematopoietic and thrombopoietic effects of DBT and
polysaccharides, extracted from the Angelica Sinensis root, on
irradiated mice (10,11). These studies indicated that
treatment with DBT significantly increased the recovery of
megakaryocytes, as well as enhanced platelet recovery and the
number of colony forming unit-megakaryocytes (CFU-MK) in
vivo (10,11). Furthermore, the polysaccharide
fraction of Angelica Sinensis increased not only the recovery of
platelets, other blood cells and their progenitor cells, but also
the number of CFUs (10).
However, the role of Radix Astragali, another major
component of DBT, in hematopoiesis and thrombopoiesis is yet to be
fully elucidated. Several compounds have been identified in Radix
Astragali, including various polysaccharides, glycosides,
alkaloids, volatile oils and organic acids (12). Among them, astragalus polysaccharide
(ASPS) from the primary constituent of Radix Astragali is
considered the most important bioactive component, as it is
hypothesized that polysaccharides are the main components in plants
that promote hematopoiesis (13).
In addition, our previous study also confirmed that angelica
polysaccharides promoted hematopoiesis (10). Therefore, the present study aimed to
investigate the effects and mechanisms of ASPS on hematopoiesis and
megakaryocytes.
Materials and methods
Ethics statement
The Animal Research Welfare Committee of Southern
Medical University approved the present experimental protocol. The
principles of the National Institutes of Health Guidelines for
Laboratory Animals (14) were
followed during the entire course of the experiments.
Preparation of herbal materials
Radix Astragali, provided and identified by the
Institute of Chinese Medicine, The Chinese University of Hong Kong,
is currently deposited in the Molecular Chinese Medicine
Laboratory, University of Hong Kong (voucher no. mcm-010) (10,11).
For the preparation of ASPS (15), Radix Astragali was sliced, boiled in
water and the extract was filtered three times. The three filtrates
were pooled and concentrated to 30 ml for 1 h in a rotary
evaporator at 70°C. After removing the protein content using the
Sevag method (16), the solution
was precipitated with 95% ethanol and collected via centrifugation
at 2,500 × g for 10 min at 4°C. Subsequently, the pellet was
dissolved in distilled water, dialyzed for 48 h and then
lyophilized, to eventually obtain the white, powdery
polysaccharide.
Animals
A total of 18 male BALB/c mice (age, 7–8 weeks;
weight, 18–20 g) were purchased from Charles River Laboratories
Japan, Inc., and given free access to food and water. Mice were
housed in 400×332×286-mm cages (6 mice/cage) in
temperature-controlled rooms (temperature, 24±2°C; humidity, 50±5%)
under a 12-h light/dark cycle. Mice were monitored twice daily for
their health status.
Radiation-induced
hemocytopenia/thrombocytopenia mouse model
Mice were randomly divided into the following four
groups: i) Control (radiation-induced, saline treated) group; ii)
ASPS-treated group; iii) rhTPO (TPO)-treated group; and iv) normal
(untreated) group. The hemocytopenia/thrombocytopenia model was
established using 4 Gy irradiation from a 137Cs source
(Gammacell-1000 Elite Irradiator; Nordion, Inc.) as previously
described (10,11,17).
Mice underwent intraperitoneal administration of ASPS (12.5 mg/day)
(10,11) or rhTPO (1 µg/kg/day; 3SBio, Inc.)
daily for 21 days starting from the day following radiotherapy.
Control mice were administered with an intraperitoneal injection of
saline (1.25 ml).
Bone marrow samples were frozen in cyomolds at −74°C
for 2 h, cut into 5-µm thick sections and fixed in 95% ethyl
alcohol at 4°C for 5 min, and then stained with Giemsa for 25 min
at room temperature. A total of 25 random high-power fields from
each bone marrow sample were chosen and visualized using a light
microscope (magnification, ×40) to determine the mean total cell
count (MTC) (18). Additionally,
the mean cell counts of the erythroid, granulocytic and
megakaryocytic cell lineages were recorded.
Blood and bone marrow collection
On days 0, 7, 14 and 21 after irradiation, 0.1 ml
peripheral blood samples were obtained via tail veins, following
administration of isoflurane (3.5% induction for 4 min and 1.5%
maintenance) anesthesia. Blood cell counts were analyzed using a
ProCyte Dx Hematology analyzer (IDEXX Laboratories, Inc.). The body
weight of all mice was also recorded in order to evaluate the
general effects of ASPS or other treatments on well-being of
animals. All mice were euthanized via cervical dislocation on day
21.
For the CFU assays, bone marrow cells were harvested
from the proximal femur by inserting a needle and forcing cell
culture medium (IMDM; Gibco; Thermo Fisher Scientific, Inc.)
supplemented with 10% FCS (HyClone; Cytiva) through the bone shaft.
Murine bone marrow cells (2×105 nucleated cells) were
cultured in 35-mm Petri dishes using the plasma clot culture method
(10,11,19).
The medium comprised 1% deionized BSA (Sigma-Aldrich; Merck KGaA),
0.34 mg CaCl2, 10% citrated bovine plasma
(Sigma-Aldrich; Merck KGaA), 100 units penicillin (Gibco; Thermo
Fisher Scientific, Inc.), 50 µg streptomycin (Gibco; Thermo Fisher
Scientifc, Inc.), 0.1 mM β-mercaptoethanol (Sigma-Aldrich; Merck
KGaA), 3 IU/ml erythropoietin (Cilag AG), 10 ng/ml IL-3 (PeproTech,
Inc.), 50 ng/ml stem cell factor (PeproTech, Inc.) and IMDM in a
total volume of 1 ml. The dishes were incubated at 37°C in a fully
humidified atmosphere with 5% CO2.
CFU-MK assay
Murine bone marrow was exposed by cutting the ends
of the femurs and was extruded by inserting a needle and forcing
cell culture medium with 10% FCS (HyClone; Cytiva) through the bone
shaft. Murine bone marrow cells (2×105 cells) were
cultured in 35-mm Petri dishes using the plasma clot culture method
(10,18). The system contained 1% deionized
BSA, 0.34 mg CaCl2, 10% citrated bovine plasma (all from
Sigma-Aldrich; Merck KGaA), 100 µg penicillin and 50 µg
streptomycin in Iscove's modified Dulbecco's medium (IMDM; Thermo
Fisher Scientific, Inc.) supplemented with different concentrations
of ASPS (0, 50, 100 or 200 µg/ml), TPO (50 ng/ml) in a total volume
of 1 ml. Cells were incubated at 37°C and 5% CO2
atmosphere for 7 days. Following incubation for 7 days, the number
of CFU-MK-derived colonies was counted using the acetylcholine
esterase staining method (incubation for 20–30 min at 37°C)
(10,11). Subsequently, cells were stained with
hematoxylin for 5 min at room temperature to count the
CFU-granulocyte macrophage (CFU-GM)-derived colonies using a light
microscope (magnification, ×10). A CFU-MK colony was defined as a
cluster of ≥3 acetylcholine esterase-positive cells, while a CFU-GM
colony consisted of a cluster of ≥40 cells (10).
Murine bone marrow CFU-fibroblast
(CFU-F) assay
A CFU-F assay was performed as previously described
(10,18). Briefly, murine bone marrow cells
(1×106 cells) from different groups were resuspended in
2 ml IMDM supplemented with 10% FCS in the presence of ASPS (100
µg/ml) or TPO (50 ng/ml), and incubated at 37°C in a fully
humidified atmosphere with 5% CO2 for 9 days. All
procedures were performed in triplicate. A CFU-F colony was defined
as an aggregate containing ≥20 fibroblasts (18). Subsequently, adherent cells were
stained with Giemsa for 25 min at room temperature, and the CFU-F
colonies were counted under an Olympus CKX53 inverted light
microscope (Olympus Corporation) at ×10 magnification.
CFU-GM, burst-forming
unit/CFU-erythroid (BFU/CFU-E) and CFU-granulocyte, erythroid,
monocyte and megakaryocyte (GEMM) assays
To further evaluate the effects of ASPS (100 µg/ml)
on hematologic progenitor cells, CFU-GM, BFU/CFU-E and CFU-GEMM
assays were performed (10,11,20).
Briefly, murine bone marrow cells (2×105 cells/ml) were
cultured in culture medium supplemented with 1% methylcellulose
(Sigma-Aldrich; Merck KGaA), 30% FCS, 1% BSA, 0.1 mM
β-mercaptoethanol (Sigma-Aldrich; Merck KGaA), 3 IU/ml
erythropoietin (Cilag AG), 10 ng/ml IL-3 and 50 ng/ml stem cell
factor (PeproTech, Inc.). Murine bone marrow cells
(2×105 cells/ml) were seeded into 35-mm Petri dishes in
triplicate and incubated for 7 days. Finally, colonies were counted
blindly using a light microscope (magnification, ×10) by two
investigators (10,11).
ELISA assay for TPO
Briefly, 0.1 ml murine peripheral blood samples were
centrifuged (400 × g; room temperature; 10 min) to obtain the
plasma. TPO levels were measured using an ELISA kit (R&D
Systems, Inc.; cat. MTP00), according to the manufacturer's
instructions. The optical density of each well was measured at 450
nm using a microplate reader (BioTek Instruments, Inc.).
Annexin V, caspase-3 and
5,5,6,6-tetrachloro-1,1,3,3-Tetraethylbenzimidazolcarbocyanine
iodide (JC-1) analysis via flow cytometry
Cells were randomly divided into normal (untreated),
control (serum-free, cytokine and serum depleted), TPO-treated,
ASPS-treated, LY294002-treated and ASPS + LY294002-treated groups.
Flow cytometric analysis was performed as described previously.
Briefly, the megakaryoblastic cell line M-07e (American Type
Culture Collection) was maintained in IMDM supplemented with
granulocyte-macrophage colony-stimulating factor (20 ng/ml) and 10%
FCS. Apoptotic cell death was induced via cytokine and serum
depletion. Cell cultures were supplemented with ASPS (100 µg/ml),
TPO (100 ng/ml) or LY294002 (25 µM) and then cells were incubated
at 37°C for 72 h. Apoptotic cell death (percentage of early + late
apoptotic cells) was evaluated using the Annexin V-FITC/PI, active
caspase-3-PE and JC-1 ApoAlert reagent kits (BD Biosciences), as
previously described (18,20,21) or
according to the manufacturer's instructions (for ApoAlert reagent
kit). Apoptotic cells were grouped based on their percentages as
‘early’ (R2; FITC+ and PI−), ‘late’ (R1;
FITC+ and PI+) and total (R1 + R2;
FITC+) apoptotic cells. A total of 10,000 events were
acquired for each sample and were analyzed on the FACSCanto™ Flow
Cytometry system (BD Biosciences) using the Lysis II C32 software
(FACScan; BD Pharmingen; BD Biosciences) (10,18,20).
The dose of ASPS was chosen as previously reported (10).
Western blot analysis
For AKT and phosphorylated (p)-AKT immunodetection,
cells were plated into 35-mm diameter plates at initial density of
5×105 cells, and were serum-starved overnight. When
required, a 30-min pre-incubation step at 37°C with the PI3K
inhibitor LY294002 (Sigma-Aldrich; Merck KGaA) was performed prior
to stimulation. Cells were stimulated for the indicated time-points
at 37°C for 30 min with TPO (100 ng/ml) or ASPS (100 µg/ml).
Subsequently, cells were rinsed rapidly in ice-cold PBS and lysed
in 2% SDS (Sigma-Aldrich; Merck KGaA) buffer containing 125 mM Tris
(pH 6.8). Lysates were then sonicated in an ice water bath for 2
min (5 sec exposure separated by 10 sec intervals), and the protein
concentration was quantified using the DC Protein assay (Bio-Rad
Laboratories, Inc). Cell lysates (40 µg/lane) were resolved using
8% SDS-PAGE. Following protein transfer, membranes were blocked
with 5% non-fat dry milk in TBS-Tween-20 buffer, containing 20 mM
Tris-HCl (pH 7.4), 150 mM NaCl and 0.05% Tween-20, for 1 h at room
temperature. Subsequently, membranes were probed with the
appropriate primary antibodies (1:1,000; all from Santa Cruz
Biotechnology, Inc.; β-actin, cat. no. sc-47778; AKT, cat. no.
sc-5298; p-AKT, cat. no. sc-293125) at 4°C overnight. The next day
membranes were incubated at room temperature for 1 h with a
corresponding peroxidase-conjugated secondary antibody (1:1,000;
anti-mouse IgGκ; cat. no. sc-516102) at the dilutions recommended
by the manufacturers. Blots were visualized using an ECL (Amersham;
Cytiva) western blotting detection system (19).
Statistical analysis
Data are presented as the mean ± SD (n=6).
Statistical analysis was performed using one-way ANOVA, followed by
Tukey's HSD test. P<0.05 was considered to indicate a
statistically significant difference. All statistical analyses were
performed using the SPSS 19.0 software (IBM Corp.).
Results
Effects of ASPS on hematopoiesis in
vitro
The results demonstrated that ASPS at 50, 100 and
200 µg/ml had a significant effect on the formation of CFU-GM and
CFU-MK compared with 0 µg/ml group. ASPS at only 100 µg/ml had a
significant effect on the formation of CFU-F. However, ASPS did not
affect the formation of BFU/CFU-E (Fig.
1).
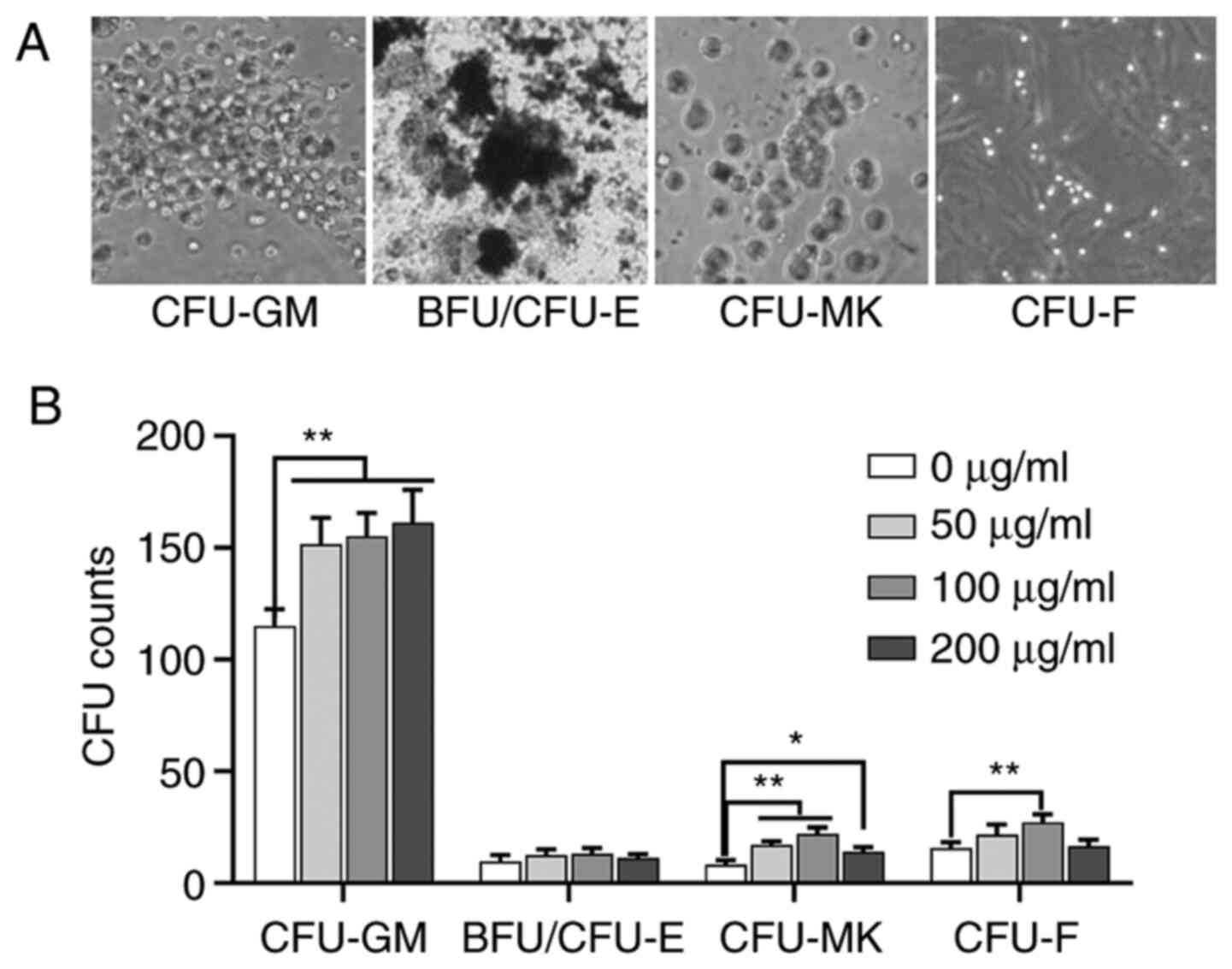 | Figure 1.Effects of different concentrations
of ASPS on the number of CFUs in vitro. (A) Representative
image of CFU-GM, BFU/CFU-E, CFU-MK and CFU-F (magnification, ×10).
(B) Effect of 0, 50, 100 and 200 µg/ml ASPS on the formation of
CFU-GM, BFU/CFU-E, CFU-MK and CFU-F. A concentration of 100 µg/ml
ASPS exhibited the maximum effect. *P<0.05, **P<0.01. ASPS,
Astragalus polysaccharide; CFU-MK, colony-forming
unit-megakaryocyte; CFU-GM, colony-forming unit-granulocyte
macrophage; BFU/CFU-E, burst-forming unit/colony-forming
unit-erythroid; CFU-GEMM, colony-forming unit- granulocyte,
erythroid, monocyte and megakaryocyte; CFU-F, colony-forming
unit-fibroblast. |
Effects of ASPS on a radiation-induced
hemocytopenia/thrombocytopenia mouse model
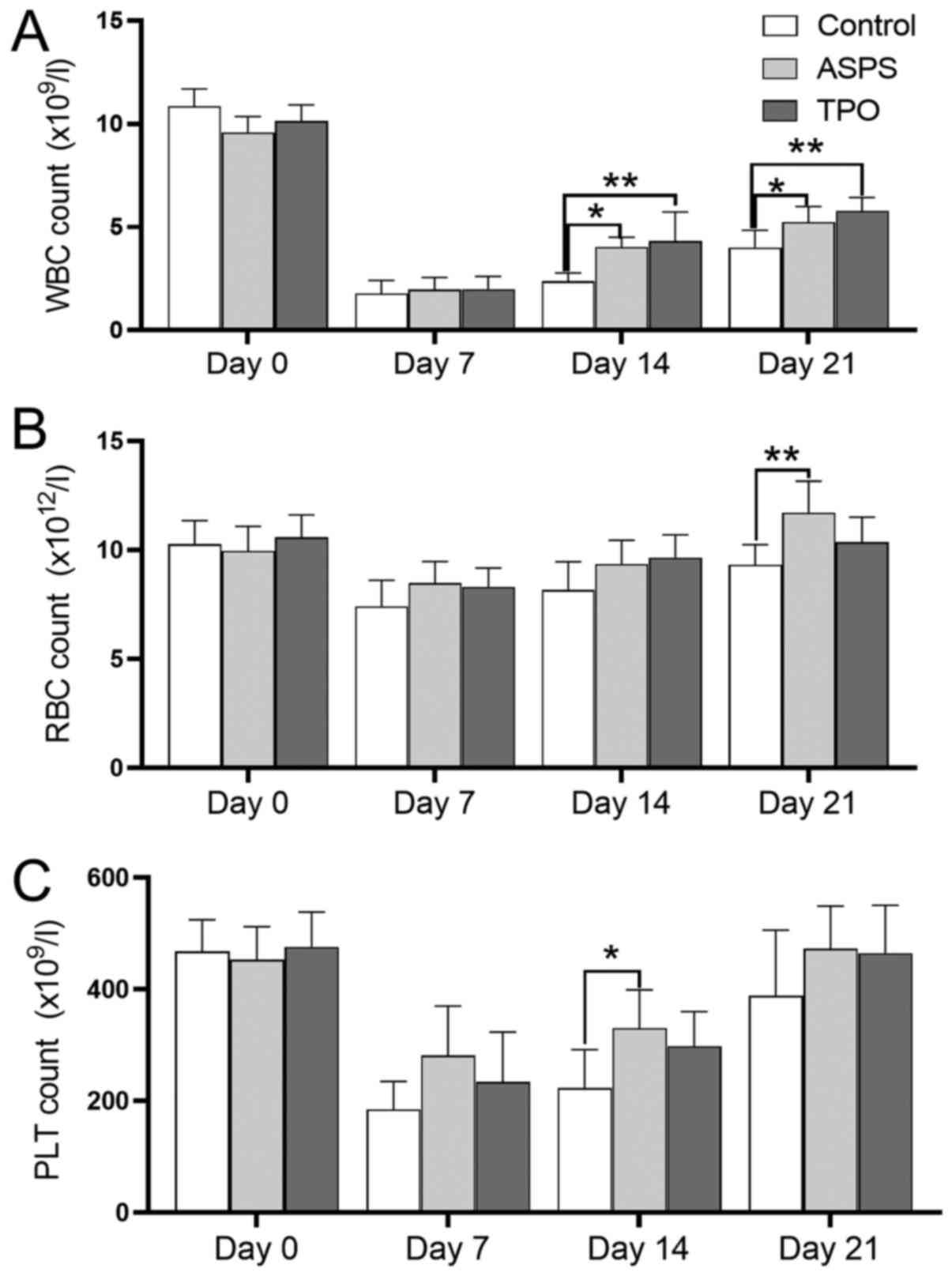
ASPS increases blood cell counts in
the hemocytopenia/thrombocytopenia mouse model
The changes in the counts of different blood cell
subpopulations are presented in Fig.
2. The number of white blood cells (WBC) in each group was
decreased to the lowest level on day 7 following exposure to
irradiation and was gradually increased thereafter. On days 14 and
21, the WBC count in the ASPS-treated group was significantly
higher compared with that to the control group. Similar results
were observed in the WBC counts between the TPO-treated and control
groups (Fig. 2A).
Following ASPS and TPO administration, the red blood
cell (RBC) counts in the ASPS- and TPO-treated groups were elevated
after day 7 (Fig. 2B). The RBC
counts did not demonstrate a statistically significant difference
between the ASPS-treated and control groups on day 7 and day 14.
However, the RBC counts in the ASPS-treated group were
significantly increased compared with the control group on day 21.
There was no significant difference in the RBC counts between the
TPO-treated and control groups from day 7 to day 21.
The myelosuppressed mice exhibited a platelet nadir
(<300×109/l) on day 7. Platelet counts were gradually
increased from day 7 to day 21 (Fig.
2C). Following ASPS (12.5 mg/day) administration, the number of
platelets was significantly elevated compared with the control
group at day 14.
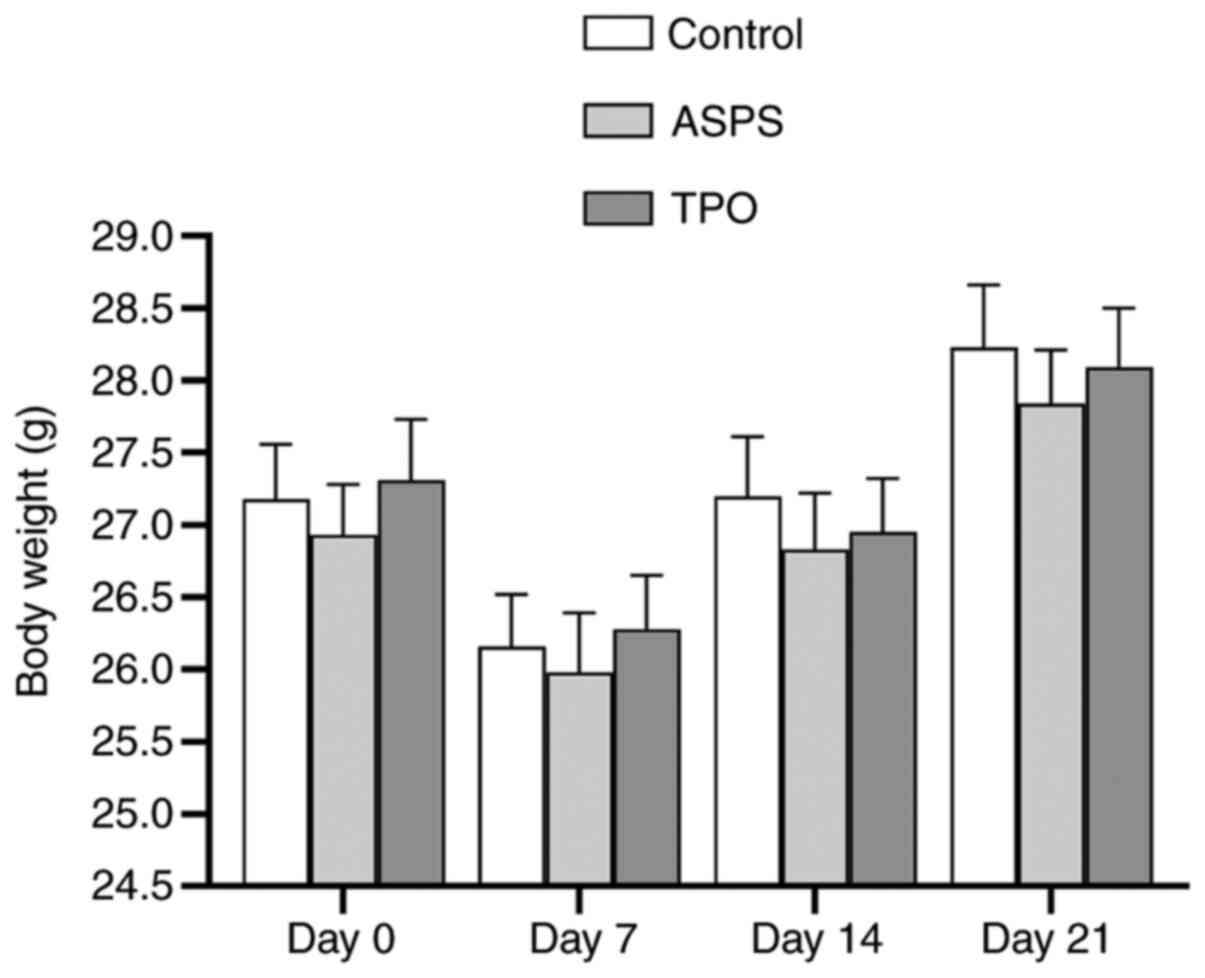
ASPS has no effect on the total body
weight in the hemocytopenia/thrombocytopenia mouse model
The total body weights in the control, ASPS- and
TPO-treated groups were lowest on day 7 and were gradually
increased thereafter. However, there was no statistically
significant difference in the body weight of all mice in the
ASPS-treated group compared with the control and TPO-treated groups
on days 14 and 21 (Fig. 3).
ASPS significantly enhances the CFU
values of bone marrow in the hemocytopenia/thrombocytopenia mouse
model
Bone marrow cells in the ASPS-, TPO-treated and
control mice were collected and cultured for CFU assays. Treatment
with ASPS significantly increased the number of CFU-GM, BFU/CFU-E,
CFU-GEMM, CFU-MK and CFU-F colonies (n=6; Fig. 4). Furthermore, TPO significantly
increased the CFU counts in all cell lineages compared with the
control group. It was found that treatment with ASPS and TPO
exhibited similar effects in CFU-GM, CFU-GEMM and CFU-MK counts.
Additionally, TPO had a much stronger effect on the number of
BFU/CFU-E colonies compared with ASPS (n=6).
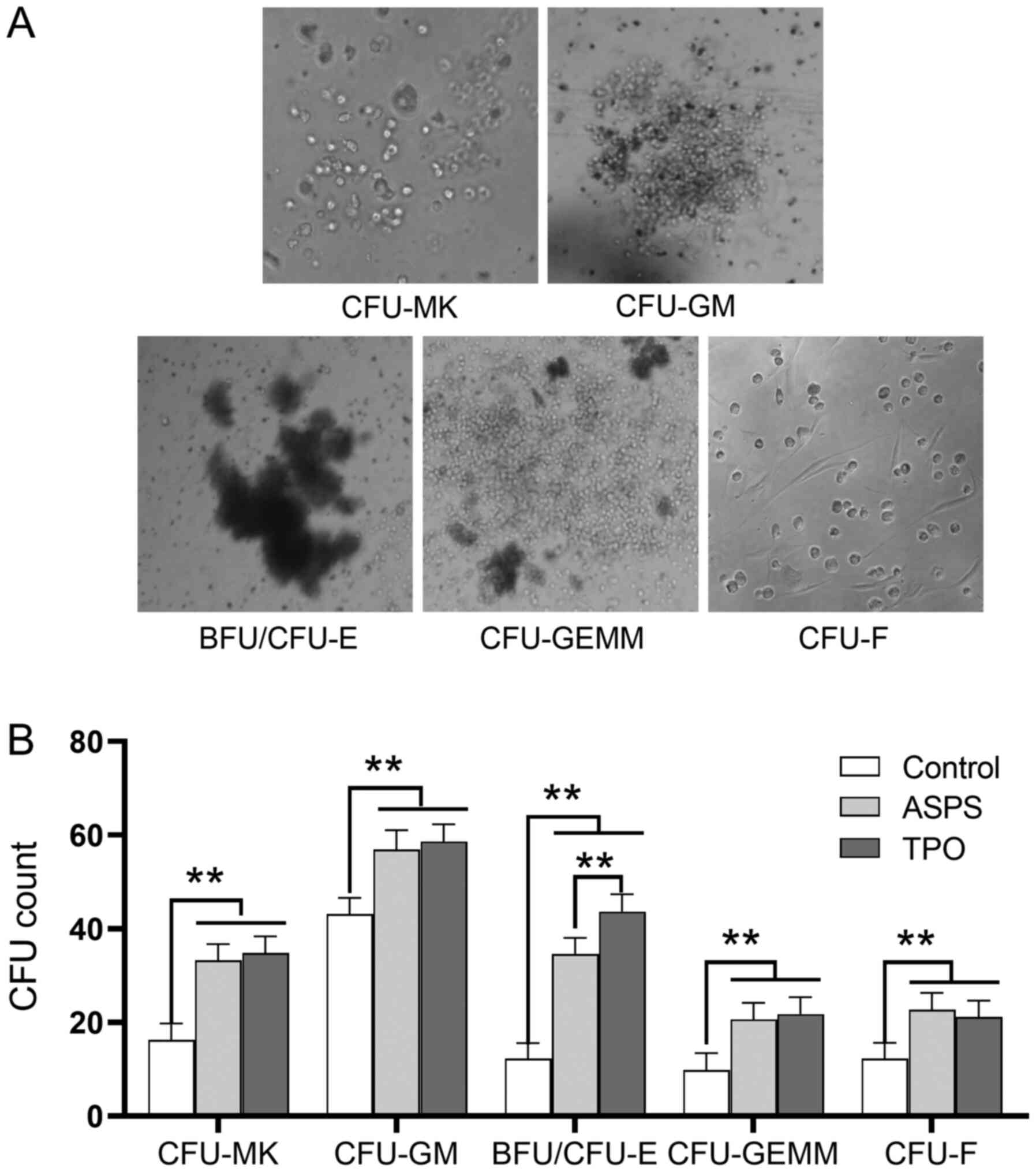 | Figure 4.ASPS promotes the formation of
CFU-GM, BFU/CFU-E, CFU-GEMM, CFU-MK and CFU-F cells in vivo.
The colony counts for various CFU, with or without treatment with
ASPS are presented. (A) Representative image of CFU-MK, CFU-GM,
BFU/CFU-E, CFU-GEMM and CFU-F (magnification, ×10). (B) Statistical
analyses were performed between the colony numbers of ASPS and
TPO-treated samples, and those of the control samples (one-way
ANOVA). **P<0.01; n=6. ASPS, Astragalus polysaccharide; TPO,
thrombopoietin; CFU-MK, colony-forming unit-megakaryocyte; CFU-GM,
colony-forming unit-granulocyte macrophage; BFU/CFU-E,
burst-forming unit/colony-forming unit-erythroid; CFU-GEMM,
colony-forming unit-granulocyte, erythroid, monocyte and
megakaryocyte; CFU-F, colony-forming unit-fibroblast. |
ASPS protects megakaryocytic cells in
bone marrow. In the present study the morphology of bone marrow
cells was also investigated using the Giemsa staining (Fig. 5)
Compared with the control group, the tri-lineage
hematopoiesis was preserved in the ASPS- and TPO-treated groups.
This effect was particularly prominent in the megakaryocytic and
granulocytic cells, but not in the erythroid one. The number of
megakaryocytic and granulocytic cells was notable elevated in the
ASPS- and TPO-treated groups, compared with that in the control
group, indicating an improved recovery of these cells compared with
those of the erythroid series. Furthermore, in the ASPS-treated
group, overall cellularity (MTC/area) was similar to that in the
TPO-treated group, mainly due to a prominent granulocytic
expansion. Finally, the recovery of the megakaryocytes was markedly
increased in the TPO-treated group compared with the control group
by day 21.
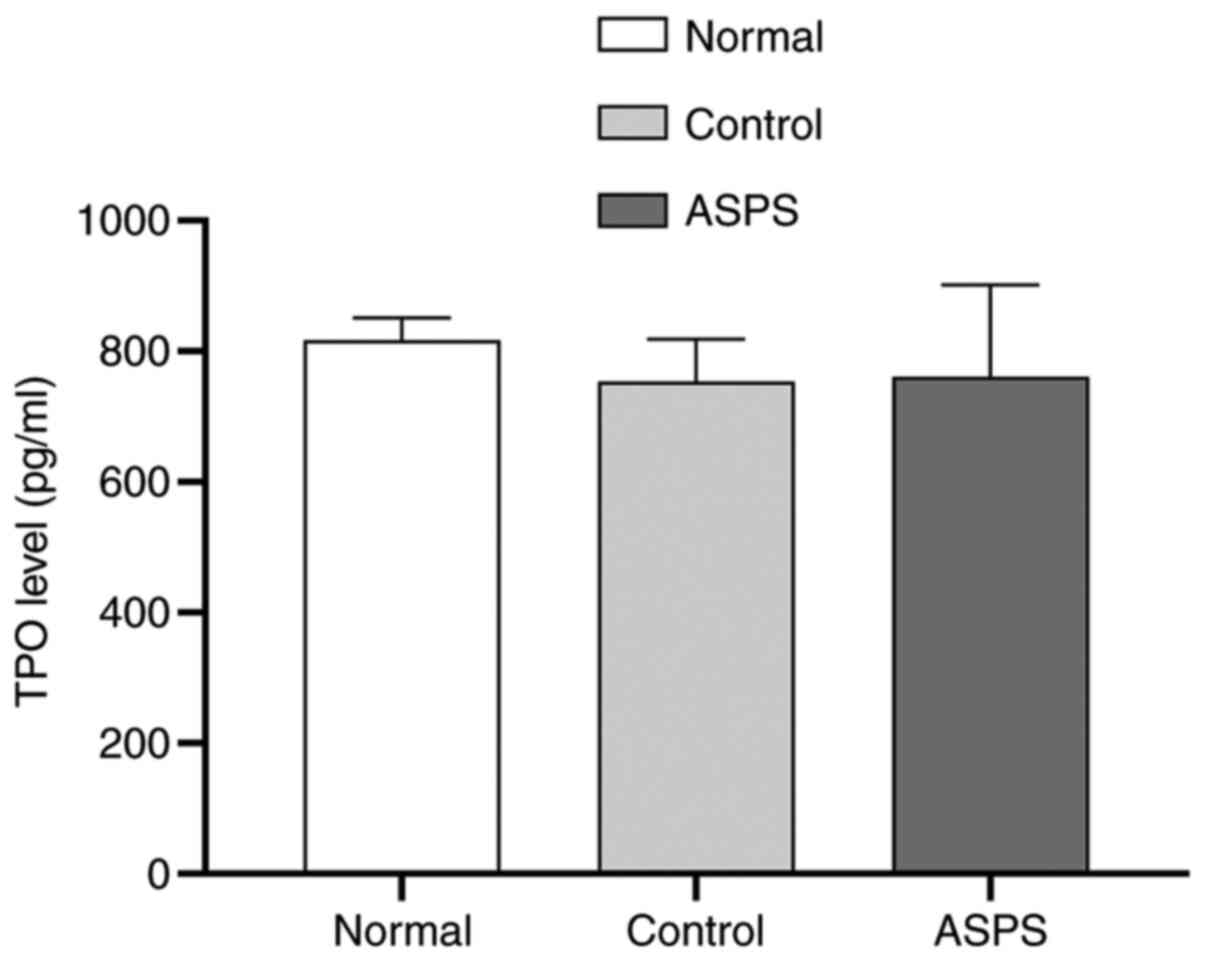
Effects of ASPS may be mediated via a
TPO-independent pathway
To determine whether ASPS could alter the expression
of TPO, ELISA was performed to measure the plasma concentration of
TPO in different mice groups. There were no significant differences
in the plasma TPO levels between the normal, control and
ASPS-treated mice (Fig. 6).
Effects of ASPS on M-07e cells in
vitro
ASPS attenuates apoptosis in M-07e
cells
Cell apoptosis was evaluated in different treatment
groups using the Annexin V assay (Fig.
7A). Apoptotic cells were grouped based on their percentages as
‘early’ (R2; FITC+ and PI−), ‘late’ (R1;
FITC+ and PI+) and total (R1 + R2;
FITC+) apoptotic cells. The lowest percentage of
apoptotic cells was obtained in normal samples and the highest one
in the LY294002-treated samples, whereas an intermediate percentage
of apoptotic cells was observed in the control samples. Treatment
with ASPS and TPO significantly decreased the early (R2), late
(R1), and total (R1 + R2) apoptotic cells compared with the control
group, and their effects were not significantly different between
each other. Compared with treatment with LY294002 alone,
co-treatment with ASPS decreased the percentage of late apoptotic
cells (R1) from 27.75 to 15.88%, early apoptotic (R2) from 18.18 to
13.10% and total apoptotic (R1 + R2) cells from 45.93 to
28.98%.
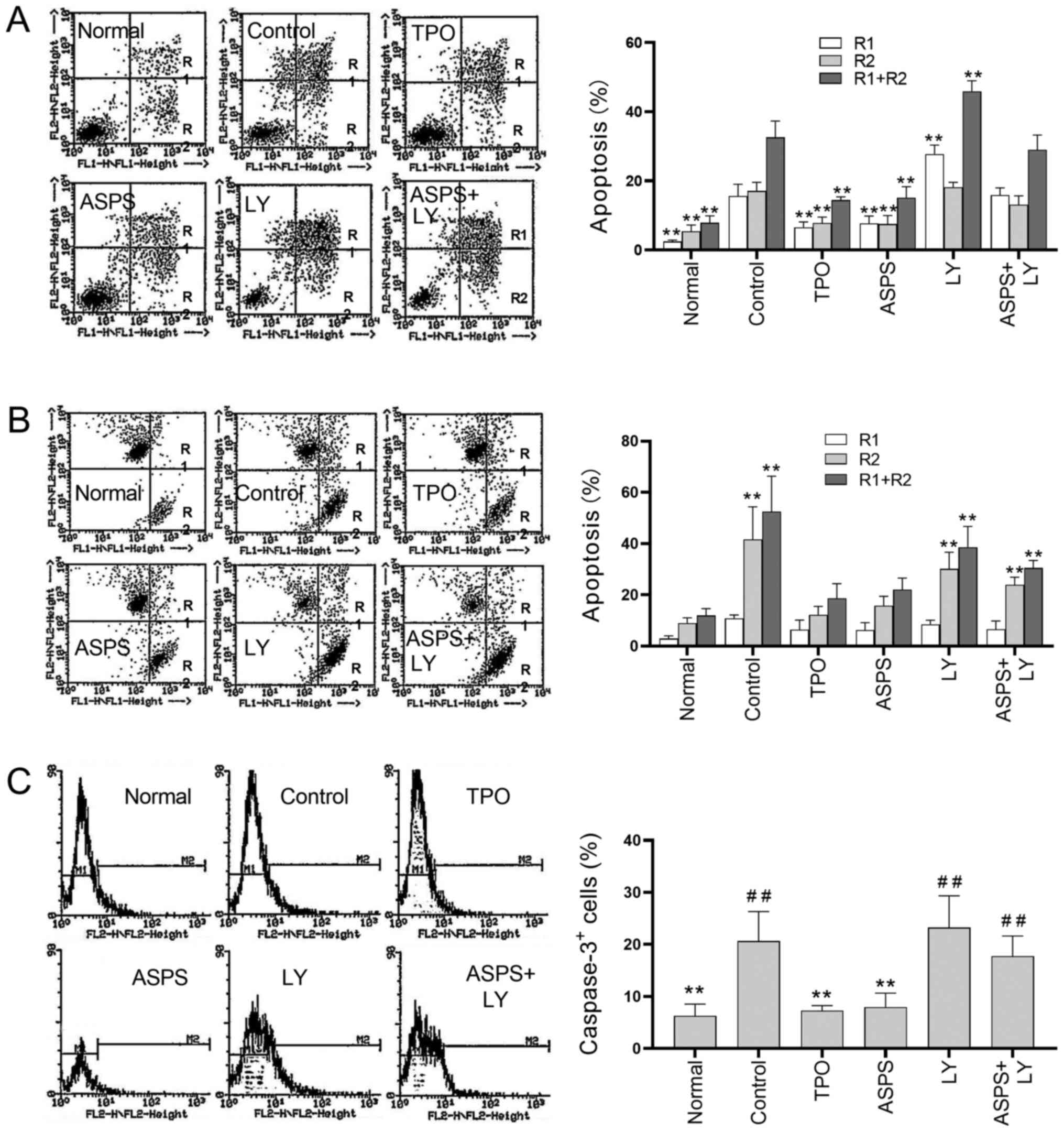 | Figure 7.Effects of ASPS on cell apoptosis
analyzed using Annexin V, JC-1 and caspase-3 assays. Apoptosis was
induced via serum depletion (control). Cells were treated with
ASPS, TPO or LY for 72 h. (A) Dot plots, quantification and
statistical analyses under various treatments were performed using
the Annexin V assay. **P<0.01 vs. control group. (B) Dot plots,
quantification and statistical analyses under various treatments
were conducted using the JC-1 assay. **P<0.01 vs. normal group.
(C) Dot plots, quantification and statistical analyses under
various treatments were performed using the caspase-3 assay. R1,
early apoptotic cells; R2, late apoptotic cells; R1 + R2, total
apoptotic cells. **P<0.01 vs. control group;
##P<0.01 vs. normal group (n=4). ASPS, Astragalus
polysaccharide; TPO, thrombopoietin; LY, LY294002. |
The antiapoptotic effect of ASPS were assessed using
the JC-1 assay, which is based on the calculation of the apoptotic
cell populations according to the distribution of the JC-1 compound
in aggregate and monomer form. In healthy cells, JC-1 aggregates
were detected in mitochondria (Fig.
7B; red fluorescence; detected in FL2) by emitting red
fluorescence, while the monomeric form was detected the cytoplasm.
However, in apoptotic cells, JC-1 aggregates emitted green
fluorescence due to the breakdown of the mitochondrial
transmembrane potential (Fig. 7B;
green fluorescence; detected in FL1). In Fig. 7B, R1 represents cells with both JC-1
aggregates and monomers, R2 cells with only JC-1 monomers and R1 +
R2 represents the total number of apoptotic cells. The percentage
of JC-1 monomer positive cells was significantly increased in the
control, LY294002- and ASPS + LY294002-treated cells, compared with
normal M-07e cells (Fig. 7B),
indicating that the induction of apoptosis in these cells.
Interestingly, no significant differences were observed in the
percentage of apoptotic cells between the ASPS or TPO-treated
samples and normal ones, suggesting that ASPS and TPO mediated
protective effects against apoptosis.
Activation of caspase-3, a downstream effector
protein of apoptosis, is indicative of apoptotic occurrence
(22). In the caspase-3 assay,
M-07e cells were treated as aforementioned, labelled with activated
caspase-3-PE dye and subjected to flow cytometric analysis.
Histograms and statistical analyses between the indicated and
normal samples are presented in Fig.
7C. The percentage of activated caspase-3 positive cells in the
ASPS- and TPO-treated cells was significantly decreased compared
with the control group. In addition, ASPS- and TPO-treated cells
were similar compared with in the normal samples. LY294002-treated,
ASPS+LY294002-treated and control cells exhibited a significantly
higher percentage of activated caspase-3 positive cells compared
with normal cells.
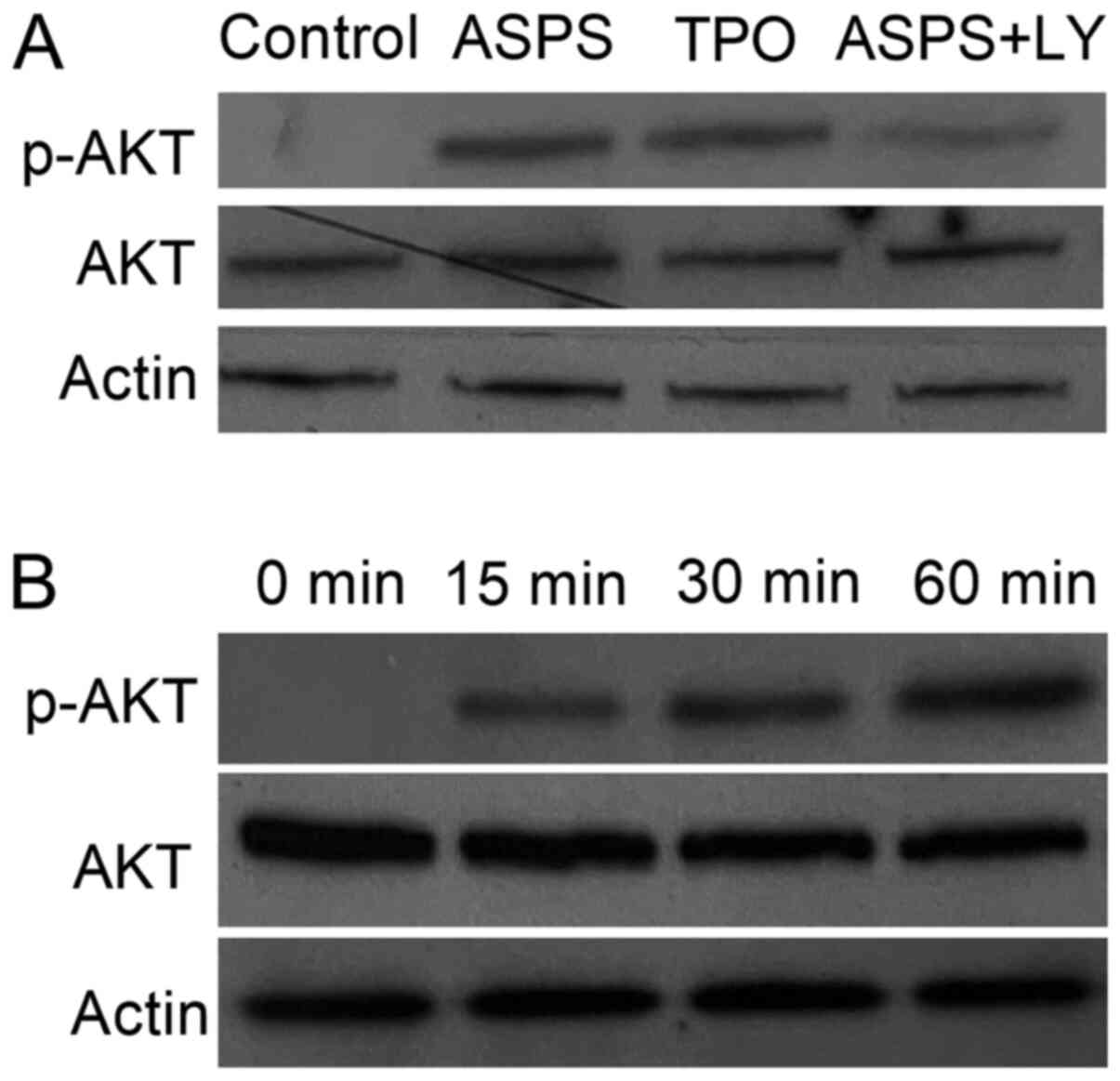
ASPS attenuates apoptosis via
activating the PI3K/AKT pathway
As indicated in Fig.
7, the percentage of apoptotic cells in ASPS + LY294002-treated
cells was significantly higher compared with ASPS-treated cells. To
confirm that ASPS inhibited cell apoptosis via the AKT pathway, the
activation status of p-AKT was detected via western blot analysis
(Fig. 8). p-AKT was detectable 15
min following treatment with ASPS. However, there was no obvious
change in the phosphorylation levels of AKT at 30 and 60 min
(Fig. 8A). Compared with the
control group, ASPS- and TPO-treated groups exhibited markedly
increased AKT activation levels, whereas no visible difference
between ASPS and TPO was observed. Moreover, treatment of M-07e
cells with ASPS + LY294002 notably suppressed the ASPS-induced AKT
activation (Fig. 8B).
Discussion
The present study demonstrated that ASPS enhanced
the in vivo recovery of peripheral blood cells and
platelets, in a murine model of hemocytopenia/thrombocytopenia.
This effect was supported by the CFU analysis results, where
treatment with ASPS stimulated the formation of CFU-MK and
CFU-GEMM. CFU is an indicator for hematopoietic growth function of
bone marrow. In the present experiment, bone marrow cells were
treated with 0, 50, 100 and 200 µg/ml ASPS. It was identified that
100 µg/ml ASPS had maximum effect, and when exceeding this dose,
the effect was gradually declined. In addition, ASPS inhibited
megakaryocyte apoptosis. In summary, the current study supplemented
our previous study on the mechanism of DBT by investigating the
hematopoietic/thrombopoietic effects of ASPS (10,11).
However, in addition to the anti-apoptotic mechanism, other
mechanisms of ASPS on hematopoiesis require further study.
Cell density is a classic indicator for evaluating
the ability of retaining the hematopoietic phenotype in three-line
cells, and has been widely used in previous articles (18,23,24).
The present results suggested that the platelet counts in
ASPS-treated mice were significantly increased at an earlier or
similar time-point compared with TPO-treated mice. In addition,
ASPS exhibited a weak effect on the proliferation of WBCs and RBCs
at day 14 and day 21. No significant difference was observed in WBC
and RBC counts between the ASPS- and TPO-treated groups. These
findings indicated that the radioprotective effect of ASPS in the
recovery of peripheral blood cells was similar to that of TPO,
especially in the recovery of platelets. Therefore, this study
suggested that ASPS could be used as an alternative approach for
the management of thrombocytopenia.
As a potential substitute of TPO, the thrombopoietic
effect of ASPS by promoting the production of TPO remains unknown.
He et al (25) reported that
ASPS could inhibit TNF-α and IL-1β mRNA expression levels.
Furthermore, it has been revealed that ASPS significantly
suppresses NF-κB activation (26),
as well as downregulates the phosphorylation of ERK and JNK
(27), two important signaling
pathways involved in the expression of TNF-α and IL-1β. The current
study suggested that ASPS could increase the number of platelets in
an inflammatory factor-independent manner. Our previous study
demonstrated that IL-1β could upregulate TPO expression, thus
resulting in the proliferation of platelets (28). Taken together, these findings,
combined with the results of the present study, indicated that ASPS
could promote the proliferation of megakaryocytes in a
TPO-independent manner.
In our previous studies, M-07e and HL-60 cells were
treated with 0, 50, 100 and 200 µg/ml ASPS, and it was identified
that 100 µg/ml had a higher stimulatory effect on cell
proliferation (29,30). In the present study, the
antiapoptotic effect of ASPS on M-07e cells was demonstrated by
performing Annexin V and activated caspase-3 assays (18,20).
The Annexin V and caspase-3 assay results demonstrated that the
percentage of the apoptotic M-07e cells in the ASPS-treated group
was significantly decreased compared with that in the control
samples (cytokine- and serum-depleted samples). With respect to the
JC-1 distribution, the number of apoptotic cells in the control
samples was significantly higher compared with that in the
ASPS-treated and normal samples. These finding indicated that ASPS
could enhance peripheral blood cell proliferation via inhibiting
cell apoptosis, but not by directly stimulating bone marrow.
Furthermore, the activation of caspase-3 in the control samples was
elevated compared with that in the ASPS-treated samples, suggesting
that ASPS attenuated megakaryocyte apoptosis via
mitochondrial-mediated pathways.
The PI3K/AKT pathway serves an important role in
cell apoptosis (31) and
proliferation (32) in a large
spectrum of cell types, including megakaryocytes (33). Several studies have reported that
cell apoptosis may be induced by regulating a variety of downstream
effector molecules of the PI3K/AKT pathway (34,35).
As the key molecule of this pathway, p-AKT actively inhibits cell
apoptosis (36). Therefore,
decreased p-AKT levels may increase the apoptotic rate of M-07e
cells. TPO is a growth factor not only for platelets, but also for
hematopoietic stem cells (37). TPO
has been shown to inhibit apoptosis via activating the PI3K pathway
in previous studies (38,39), and it is a common drug for positive
control in the study of hematopoiesis and thrombopoiesis (13,40,41).
The results of the present study indicated that ASPS inhibited
M-07e cell apoptosis via regulating the levels of p-AKT within the
PI3K/AKT pathway.
In conclusion, in the present study, the
hematopoietic/thrombopoietic effects of ASPS were confirmed in
vivo and in vitro, which were attributed to two
different mechanisms. Firstly, ASPS promoted the proliferation of
hematopoietic CFUs, and secondly, inhibited megakaryocyte
apoptosis. These findings may facilitate the development of novel
alternative therapies for patients with myelosuppression, including
thrombocytopenia.
Acknowledgements
The authors would like to thank Mr. Nga Hin Pong
from The Chinese University of Hong Kong for technical assistance
in the animal procedures and other studies.
Funding
This work was supported by the National Natural
Science Foundation of China (grant no. 81770116), the Funding of
Traditional Medicine Project from the Health Department of
Guangming District, the Shenzhen, Science, Technology and
Innovation Commission of Shenzhen, the China's Postdoctoral Science
Foundation (grant no. 2019TQ0383) and the Sanming Project of
Medicine in Shenzhen (grant no. SZSM202011004).
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
MY and LL contributed to the conception and design
of the present study, and provided the final proofs of the
manuscript version to be published. WX and LL conducted the
experiments and collected data. LL, CKL, CY and CC performed the
data and statistical analyses, and drafted the manuscript. MY, HXi,
YC, CC, CKL, XF, LY and HXu analyzed the data and revised the
manuscript repeatedly. All authors read and approved the final
manuscript.
Ethics approval and consent to
participate
The experimental protocol of the present study was
approved by the Animal Research Welfare Committee of Southern
Medical University. The principles of the National Institutes of
Health Guidelines for Laboratory Animals were followed during the
entire course of the experiments. All parts of this report are in
compliance with the ARRIVE Guidelines for reporting animal research
(42).
Patient consent for publication
Not applicable
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Liebman HA: Thrombocytopenia in cancer
patients. Thromb Res. 133 (Suppl 2):S63–S69. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Lieberman L, Bercovitz RS, Sholapur NS,
Heddle NM, Stanworth SJ and Arnold DM: Platelet transfusions for
critically ill patients with thrombocytopenia. Blood.
123:1146–1151; quiz 1280. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Cines DB, Gernsheimer T, Wasser J, Godeau
B, Provan D, Lyons R, Altomare I, Wang X and Lopez A: Integrated
analysis of long-term safety in patients with chronic immune
thrombocytopaenia (ITP) treated with the thrombopoietin (TPO)
receptor agonist romiplostim. Int J Hematol. 102:259–270. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Arnold DM, Vrbensky JR, Karim N, Smith JW,
Liu Y, Ivetic N, Kelton JG and Nazy I: The effect of rituximab on
anti-platelet autoantibody levels in patients with immune
thrombocytopenia. Br J Haematol. 178:302–307. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Aubron C, Flint AW, Bailey M, Pilcher D,
Cheng AC, Hegarty C, Martinelli A, Reade MC, Bellomo R and
McQuilten Z: Is platelet transfusion associated with
hospital-acquired infections in critically ill patients? Crit Care.
21:22017. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Scheinberg P, Singulane CC, Barbosa LS and
Scheinberg M: Successful platelet count recovery in
lupus-associated thrombocytopenia with the thrombopoietin agonist
eltrombopag. Clin Rheumatol. 33:1347–1349. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Shin SK, Pack SP, Oh JG, Kang NK, Chang
MH, Chung YH, Kim SJ, Lee JW and Heo TH: Anti-erythropoietin and
anti-thrombopoietin antibodies induced after administration of
recombinant human erythropoietin. Int Immunopharmacol.
11:2237–2241. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Gong AG, Li N, Lau KM, Lee PS, Yan L, Xu
ML, Lam CT, Kong AY, Lin HQ, Dong TT, et al: Calycosin orchestrates
the functions of Danggui Buxue Tang, a Chinese herbal decoction
composing of Astragali Radix and Angelica Sinensis Radix: An
evaluation by using calycosin-knock out herbal extract. J
Ethnopharmacol. 168:150–157. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Lin HQ, Gong AG, Wang HY, Duan R, Dong TT,
Zhao KJ and Tsim KW: Danggui Buxue Tang (Astragali Radix and
Angelicae Sinensis Radix) for menopausal symptoms: A review. J
Ethnopharmacol. 199:205–210. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Liu C, Li J, Meng FY, Liang SX, Deng R, Li
CK, Pong NH, Lau CP, Cheng SW, Ye JY, et al: Polysaccharides from
the root of Angelica sinensis promotes hematopoiesis and
thrombopoiesis through the PI3K/AKT pathway. BMC Complement Altern
Med. 10:792010. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Yang M, Chan GC, Deng R, Ng MH, Cheng SW,
Lau CP, Ye JY, Wang L and Liu C: An herbal decoction of Radix
astragali and Radix angelicae sinensis promotes hematopoiesis and
thrombopoiesis. J Ethnopharmacol. 124:87–97. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Zhao M, Zhang ZF, Ding Y, Wang JB and Li
Y: Astragalus polysaccharide improves palmitate-induced insulin
resistance by inhibiting PTP1B and NF-κB in
C2C12 myotubes. Molecules. 17:7083–7092.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Xie JH, Jin ML, Morris GA, Zha XQ, Chen
HQ, Yi Y, Li JE, Wang ZJ, Gao J, Nie SP, et al: Advances on
bioactive polysaccharides from medicinal plants. Crit Rev Food Sci
Nutr. 56 (Suppl 1):S60–S84. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
National Research Council, . Guide for the
Care and Use of Laboratory Animals. National Academies Press;
Washington, DC: 1985
|
|
15
|
Yuan S, Piao X, Li D, Kim S, Lee H and Guo
P: Effects of dietary Astragalus polysaccharide on growth
performance and immune function in weaned pigs. Anim Sci.
82:501–507. 2006. View Article : Google Scholar
|
|
16
|
Staub A: Removeal of protein-Sevag method.
Methods Carbohydr Chem. 5:5–6. 1965.
|
|
17
|
Inagaki K, Oda T, Naka Y, Shinkai H,
Komatsu N and Iwamura H: Induction of megakaryocytopoiesis and
thrombocytopoiesis by JTZ-132, a novel small molecule with
thrombopoietin mimetic activities. Blood. 104:58–64. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Ye JY, Chan GC, Qiao L, Lian Q, Meng FY,
Luo XQ, Khachigian LM, Ma M, Deng R, Chen JL, et al:
Platelet-derived growth factor enhances platelet recovery in a
murine model of radiation-induced thrombocytopenia and reduces
apoptosis in megakaryocytes via its receptors and the PI3-k/Akt
pathway. Haematologica. 95:1745–1753. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Yang M, Li K, Chui CM, Yuen PM, Chan PK,
Chuen CK, Li CK and Fok TF: Expression of interleukin (IL) 1 type I
and type II receptors in megakaryocytic cells and enhancing effects
of IL-1beta on megakaryocytopoiesis and NF-E2 expression. Br J
Haematol. 111:371–380. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Yang M, Li K, Ng PC, Chuen CK, Lau TK,
Cheng YS, Liu YS, Li CK, Yuen PM, James AE, et al: Promoting
effects of serotonin on hematopoiesis: Ex vivo expansion of cord
blood CD34+ stem/progenitor cells, proliferation of bone
marrow stromal cells, and antiapoptosis. Stem Cells. 25:1800–1806.
2007. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Li K, Sung RY, Huang WZ, Yang M, Pong NH,
Lee SM, Chan WY, Zhao H, To MY, Fok TF, et al: Thrombopoietin
protects against in vitro and in vivo cardiotoxicity induced by
doxorubicin. Circulation. 113:2211–2220. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Porter AG and Jänicke RU: Emerging roles
of caspase-3 in apoptosis. Cell Death Differ. 6:99–104. 1999.
View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Zhang J, Zhou S, Zhou Y, Feng F, Wang Q,
Zhu X, Zhao J, Fu H, Lv M, Ai H, et al: Adipose-derived mesenchymal
stem cells (ADSCs) with the potential to ameliorate platelet
recovery, enhance megakaryopoiesis, and inhibit apoptosis of bone
marrow cells in a mouse model of radiation-induced
thrombocytopenia. Cell Transplant. 25:261–273. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
MacVittie TJ, Farese AM, Smith WG, Baum
CM, Burton E and McKearn JP: Myelopoietin, an engineered chimeric
IL-3 and G-CSF receptor agonist, stimulates multilineage
hematopoietic recovery in a nonhuman primate model of
radiation-induced myelosuppression. Blood. 95:837–845. 2000.
View Article : Google Scholar : PubMed/NCBI
|
|
25
|
He X, Shu J, Xu L, Lu C and Lu A:
Inhibitory effect of Astragalus polysaccharides on
lipopolysaccharide-induced TNF-α and IL-1β production in THP-1
cells. Molecules. 17:3155–3164. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Xue H, Gan F, Zhang Z, Hu J, Chen X and
Huang K: Astragalus polysaccharides inhibits PCV2 replication by
inhibiting oxidative stress and blocking NF-κB pathway. Int J Biol
Macromol. 81:22–30. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Han L, Wang HX and Lu MI: Effect of
Astragalus polysaccharide on LPS-induced cardiomyocyte apoptosis by
inhibiting NF-κB and JNK signaling pathway. Chinese Pharmacol Bull.
34:243–249. 2018.
|
|
28
|
Chuen CK, Li K, Yang M, Fok TF, Li CK,
Chui CM and Yuen PM: Interleukin-1β up-regulates the expression of
thrombopoietin and transcription factors c-Jun, c-Fos, GATA-1, and
NF-E2 in megakaryocytic cells. J Lab Clin Med. 143:75–88. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Xiao B, Xu Y, He H, Jiang QL, Li SY, Shu
HY, Liang EY, Yi ZS, Ye JY, Huang LF, et al: Anti-apoptotic effect
of Astragalus Polysaccharide on myeloid cells. Zhongguo Shi Yan Xue
Ye Xue Za Zhi. 21:1243–1247. 2013.(In Chinese). PubMed/NCBI
|
|
30
|
Yang M, Jiang Q, Xiao B, Liu C, Li S,
Huang L, Chong B and Meng F: Astragalus polysaccharide has
hematopoietic and thrombopoietic activities in an irradiation mouse
model. Blood. 122:42162013. View Article : Google Scholar
|
|
31
|
Jiang X, Zeng L, Huang J, Zhou H and Liu
Y: Arctigenin, a natural lignan compound, induces apoptotic death
of hepatocellular carcinoma cells via suppression of PI3-K/Akt
signaling. J Biochem Mol Toxicol. 29:458–464. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Liu R, Ding L, Yu MH, Wang HQ, Li WC, Cao
Z, Zhang P, Yao BC, Tang J, Ke Q, et al: Effects of
dihydrotestosterone on adhesion and proliferation via PI3-K/Akt
signaling in endothelial progenitor cells. Endocrine. 46:634–643.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Martini M, De Santis MC, Braccini L,
Gulluni F and Hirsch E: PI3K/AKT signaling pathway and cancer: An
updated review. Ann Med. 46:372–383. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Hu C, Xu M, Qin R, Chen W and Xu X:
Wogonin induces apoptosis and endoplasmic reticulum stress in HL-60
leukemia cells through inhibition of the PI3K-AKT signaling
pathway. Oncol Rep. 33:3146–3154. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Ma W, Wang DD, Li L, Feng YK, Gu HM, Zhu
GM, Piao JH, Yang Y, Gao X and Zhang PX: Caveolin-1 plays a key
role in the oleanolic acid-induced apoptosis of HL-60 cells. Oncol
Rep. 32:293–301. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Liu Y, Bi T, Wang G, Dai W, Wu G, Qian L,
Gao Q and Shen G: Lupeol inhibits proliferation and induces
apoptosis of human pancreatic cancer PCNA-1 cells through AKT/ERK
pathways. Naunyn Schmiedebergs Arch Pharmacol. 388:295–304. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Hitchcock IS and Kaushansky K:
Thrombopoietin from beginning to end. Br J Haematol. 165:259–268.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Pulikkan JA, Madera D, Xue L, Bradley P,
Landrette SF, Kuo YH, Abbas S, Zhu LJ, Valk P and Castilla LH:
Thrombopoietin/MPL participates in initiating and maintaining
RUNX1-ETO acute myeloid leukemia via PI3K/AKT signaling. Blood.
120:868–879. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Chan KY, Xiang P, Zhou L, Li K, Ng PC,
Wang CC, Zhang L, Deng HY, Pong NH, Zhao H, et al: Thrombopoietin
protects against doxorubicin-induced cardiomyopathy, improves
cardiac function, and reversely alters specific signalling
networks. Eur J Heart Fail. 13:366–376. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Su RJ, Zhang XB, Li K, Yang M, Li CK, Fok
TF, James AE, Pong H and Yuen PM: Platelet-derived growth factor
promotes ex vivo expansion of CD34+ cells from human
cord blood and enhances long-term culture-initiating cells,
non-obese diabetic/severe combined immunodeficient repopulating
cells and formation of adherent cells. Br J Haematol. 117:735–746.
2002. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Ye JY, Liang EY, Cheng YS, Chan GC, Ding
Y, Meng F, Ng MH, Chong BH, Lian Q and Yang M: Serotonin enhances
megakaryopoiesis and proplatelet formation via p-Erk1/2 and F-actin
reorganization. Stem Cells. 32:2973–2982. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Kilkenny C, Browne WJ, Cuthill IC, Emerson
M and Altman DG: Improving bioscience research reporting: The
ARRIVE guidelines for reporting animal research. J Pharmacol
Pharmacother. 1:94–99. 2010. View Article : Google Scholar : PubMed/NCBI
|