Introduction
Cervical cancer is the fourth most common type of
malignancy and a prominent cause of cancer-related deaths in women
worldwide (1). Indeed, cervical
cancer was responsible for 570,000 cases and 311,000 deaths in 2018
worldwide, ranking after breast cancer (2.1 million cases),
colorectal cancer (0.8 million) and lung cancer (0.7 million)
(1). Clinical characteristics, such
as advanced tumor stage and lymph node metastasis, have been
associated with poor prognosis and poor therapeutic outcomes
(2). The 5-year overall survival
for women with locally advanced cervical cancers is ~70% following
completion of concurrent chemoradiotherapy (2). Despite a steady decline in cervical
cancer incidence and mortality rates in recent years due to human
papillomavirus (HPV)-based screening programs and HPV vaccination
programs, cervical cancer continues to pose a considerable threat
to middle-aged women (3).
Therefore, there remains an urgent requirement to discover novel
prognostic biomarkers and to determine the important signaling
pathways that are activated or inactivated in the event of cervical
cancer. In addition, more effective therapeutic strategies for
anticancer therapy in cervical cancer need to be developed.
Long non-coding RNAs (lncRNAs) are transcripts of
>200 nucleotides in length that do not encode proteins (4). Previous studies have reported that
lncRNAs have functions in numerous biological activities, including
cell cycle regulation (5), stem
cell differentiation (6), the
immune response (7), cancer
progression (8) and chemotherapy
resistance (9). A few studies have
reported that the lncRNA, long intergenic non-protein coding RNA
861 (LINC00861), was involved in the progression and prognosis of a
wide range of types of cancer. For example, the downregulated
expression levels of LINC00861 were reportedly associated with a
poor prognosis in ovarian cancer (10). LINC00861 expression levels were also
demonstrated to be downregulated in an early recurrence group of
patients with hepatocellular carcinoma compared with a non-early
recurrence group of patients (11).
In addition, in another previous study, LINC00861 was able to bind
to hsa-microRNA (miRNA/miR)-510, and it was associated with
clinical biomarkers, such as the receptor tyrosine protein kinase
ERBB-2, estrogen receptor and progesterone receptor, in breast
cancer (12). Emerging evidence has
revealed that lncRNAs serve as competing endogenous RNAs (ceRNAs)
to regulate the epithelial-mesenchymal transition (EMT) process,
such as the lncRNAs plasmacytoma variant translocation 1 and colon
cancer associated transcript-1, in cervical cancer (13,14).
However, the expression and underlying mechanism of LINC00861 in
cervical cancer progression remain unclear.
In the current study, the expression levels of
LINC00861 were discovered to be significantly downregulated in
cervical cancer tissues, as well as CaSki and ME-180 cell lines.
The downregulated LINC00861 expression levels were associated with
an advanced-stage, lymph node metastasis and the poor survival of
patients with cervical cancer. LINC00861 overexpression was also
demonstrated to inhibit cervical cancer cell proliferation,
migration, invasion and EMT via functioning as a ceRNA for
miR-513b-5p, which subsequently regulated the PTEN/AKT/mTOR
signaling pathway.
Materials and methods
Clinical specimens
A total of 56 cervical cancer tissues from patients
(age, 35–59) with cervical cancer and their matched adjacent normal
tissues were obtained from the Department of Gynecology at Yantai
Hospital of Traditional Chinese Medicine from April 2017 to
December 2018. All samples were collected, snap-frozen in liquid
nitrogen and kept at −80°C until use. Patients were included if
they were pathologically diagnosed with cervical cancer and had not
received anticancer therapy prior to tumor excision. Patients were
excluded if they had other malignant tumors. Written informed
consent was obtained from each patient. The study protocol was
approved by the Research Ethics Committee of Yantai Hospital of
Traditional Chinese Medicine (approval no. YTSZYYY2017-112).
All diagnoses were histologically confirmed
independently by at least two experienced pathologists following
the examination of the sections following hematoxylin and eosin
(H&E) staining. Tissues were fixed in 4% paraformaldehyde for
24 h at 25°C, dehydrated, embedded in paraffin, and cut into 4-µm
slices for histological staining. These sections were stained with
hematoxylin (Beyotime Institute of Biotechnology) for 15 min and
eosin (Beyotime Institute of Biotechnology) for 5 min at 25°C.
Samples were examined under a light microscope (magnification,
×400; Nikon Corporation).
Cell culture and transfection
Human cervical cancer cell lines, CaSki and ME-180,
and the human cervical epithelial cell line, Ect1/E6E7, were
obtained from the American Type Culture Collection. All cells were
maintained in RPMI-1640 medium (HyClone; Cytiva) supplemented with
10% FBS (Gibco; Thermo Fisher Scientific, Inc.), and maintained at
37°C in a humidified atmosphere with 5% CO2.
CaSki and ME-180 cells (1.5×106 cells/ml)
were seeded into 6-well plates and transfected with 50 nM
pcDNA3.1-LINC00861 or empty vector pcDNA3.1 as a negative control
(NC), (both Shanghai GenePharma Co., Ltd.) using
Lipofectamine® 2000 reagent (Invitrogen; Thermo Fisher
Scientific, Inc.) 37°C for 6 h, according to the manufacturer's
protocol. miR-513b-5p expression levels were overexpressed or
knocked down by transfecting CaSki and ME-180 cells
(1.5×106 cells/ml) with 50 nM miR-513b-5p mimic or
miR-513b-5p inhibitor, respectively, or their respective NCs, mimic
NC and inhibitor NC (all Shanghai GenePharma Co., Ltd.). The
sequences are listed in Table I.
The transfections were also performed using
Lipofectamine® 2000 reagent (Invitrogen; Thermo Fisher
Scientific, Inc.) according to the manufacturer's instructions at
37°C for 6 h. CaSki and ME-180 cells were harvested at 48 h after
transfection.
 | Table I.Oligonucleotide sequences. |
Table I.
Oligonucleotide sequences.
| Nucleic acids
name | Sequence
(5′→3′) |
|---|
|
MicroRNA-513b-5p |
UUCACAAGGAGGUGUCAUUUAU |
| mimic |
AAAUGACACCUCCUUGUGAAUU |
| Mimic NC |
UUCUCCGAACGUGUCACGUTTACGUGACACGUUCGGAGAATT |
| MicroRNA-513b-5p
inhibitor |
AUAAAUGACACCUCCUUGUGAA |
| Inhibitor NC |
CAGUACUUUUGUGUAGUACAA |
Cell Counting Kit-8 (CCK-8) assay
CaSki and ME-180 cells were seeded into 96-well
plates at a density of 2×103 cells/100 µl and incubated
at 37°C for 24, 48 and 72 h. Subsequently, 10 µl CCK-8 solution
(Dojindo Molecular Technologies, Inc.) was added to each well and
incubated for 2 h. The absorbance at a wavelength of 450 nm was
measured in each well using a microplate reader (Bio-Rad
Laboratories, Inc.).
Colony formation assay
CaSki and ME-180 cells were plated into six-well
plates at a density of 1×103 cells/100 µl. Following
incubation for 12 days at 37°C, the cells were fixed with 4%
paraformaldehyde at 25°C for 15 min and stained with 0.1% crystal
violet solution at 25°C for 20 min. Colonies were counted under a
light microscope (magnification, ×100; Nikon Corporation), with
>50 cells considered a colony.
Wound healing assay
CaSki and ME-180 cells were seeded in six-well
plates at a density of 4×105 cells/well. The cell
monolayers were scratched using a 200-µl pipette tip once the
cultured cells reached 100% confluence and then cultured in
serum-free RPMI-1640 medium. At 0 and 24 h, the width of the wound
was visualized under a light microscope (magnification, ×100; Nikon
Corporation). The wound healing rate was calculated by using the
formula: The wound healing rate (%) = (width of the wound at 0
h-width of the wound at 24 h)/width of the wound at 0 h ×100%.
Transwell invasion assay
CaSki and ME-180 cells at a density of
4×103 cells/100 µl in serum-free RPMI-1640 medium were
plated into the upper chambers of 24-well Transwell plates
(Corning, Inc.) precoated with Matrigel (EMD Millipore) at 37°C for
60 min. The lower chambers were filled with 500 µl RPMI-1640 medium
supplemented with 10% FBS. Following 24 h of incubation at 37°C,
the invasive cells were fixed with 75% methanol at 25°C for 15 min
and then stained with 0.1% crystal violet (Sigma-Aldrich; Merck
KGaA) for at 25°C 15 min. The invasive cells were visualized under
a light microscope (magnification, ×100; Nikon Corporation).
Dual luciferase reporter assay
The Diana Tools (http://carolina.imis.athena-innovation.gr/diana_tools/web/index.php)
and TargetScan 7.2 (http://www.targetscan.org/vert_72/) bioinformatics
databases were used to identify potential binding sites between
LINC00861, miR-513b-5p and PTEN. The mutations in the miR-513-5p
binding sites were introduced using the GeneArt™ site-directed
mutagenesis system (Thermo Fisher Scientifc, Inc.). The LINC00861
wild-type (WT) or LINC00861 mutant (MUT) and PTEN-WT or PTEN-MUT
binding miR-513-5p were synthesized and cloned into the
Renilla psiCHECK2 vector (Promega Corporation).
Subsequently, CaSki and ME-180 cells at 5×104 cells/well
in 24-well plates were co-transfected with 500 ng the LINC00861-WT,
LINC00861-MUT, PTEN-WT or PTEN-MUT vectors and the miR-513-5p mimic
or mimic NC, using Lipofectamine 2000 reagent. Following 48 h of
transfection at 37°C, the relative luciferase activity was measured
using a Dual Luciferase Reporter assay system (Promega
Corporation). Renilla luciferase activity was used as the
normalization.
Reverse transcription-quantitative PCR
(RT-qPCR)
Total RNA was extracted from frozen tissues and
cultured cells (CaSki, ME-180, and Ect1/E6E7) using
TRIzol® reagent (Invitrogen; Thermo Fisher Scientific,
Inc.). 1 µg RNA was reverse transcribed into cDNA using
PrimeScript™ RT reagent kit with gDNA Eraser (cat. no. RR047A;
Takara Bio, Inc.). qPCR was subsequently performed on an Applied
Biosystems 7500 Real-Time PCR system (Thermo Fisher Scientific,
Inc.) using the Power SYBR™ Green PCR Master mix (Applied
Biosystems; Thermo Fisher Scientific, Inc.). qPCR was amplified
using the following thermocycling conditions: i) Initial
denaturation at 95°C for 30 sec; followed by 35 cycles of
denaturation at 95°C for 5 sec, annealing at 55°C for 20 sec and
extension at 72°C for 20 sec. The primer sequences used for qPCR
are listed in Table II. U6, which
is a small nuclear RNA, and GAPDH were used as the internal
controls for miRNA and mRNA, respectively. Data were analysed using
the 2−ΔΔCq method (15).
| Table II.Primers used for reverse
transcription-quantitative PCR. |
Table II.
Primers used for reverse
transcription-quantitative PCR.
| Gene | Primer sequence
(5′→3′) |
|---|
| Long intergenic
non- | F:
ATGGTTAGGCACATGGGGTG |
| protein coding RNA
861 | R:
CCAGGCTCTGGGCAACATTA |
|
MicroRNA-513b-5p | F:
TGGTACTGATGTGATGGACT |
|
| R:
TCATATCACACAGCACCGAT |
| U6 | F:
CTCGCTTCGGCAGCACA |
|
| R:
AACGCTTCACGAATTTGCGT |
| GAPDH | F:
ATGGGGAAGGTGAAGGT |
|
| R:
AAGCTTCCCGTTCTCAG |
Western blotting
Total protein was extracted from the CaSki and
ME-180 cell lines using RIPA lysis buffer (Thermo Fisher
Scientific, Inc.). Total protein was quantified by using an
Enhanced BCA Protein Assay kit (Beyotime Institute of
Biotechnology) and 50 µg protein/lane was separated via 10%
SDS-PAGE. The separated proteins were subsequently transferred onto
PVDF membranes (EMD Millipore) and blocked by using 5% non-fat milk
for 1 h at room temperature. The membranes were then incubated
overnight at 4°C with the following primary antibodies (all Cell
Signaling Technology, Inc.): Anti-PTEN (1:2,000; cat. no. 9188),
anti-phosphorylated (p)-AKT (1:2,000; cat. no. 4060), anti-AKT
(1:2,000; cat. no. 9272), anti-p-mTOR (1:2,000; cat. no. 5536),
anti-mTOR (1:2,000; cat. no. 2972), anti-E-cadherin (1:2,000; cat.
no. 3195), anti-Snail (1:2,000; cat. no. 3879), anti-vimentin
(1:2,000; cat. no. 5741) and anti-GAPDH (1:2,000; cat. no. 5174).
Following washing with TBST, the membranes were incubated with the
HRP-conjugated anti-rabbit secondary antibodies (1:3,000, A0208;
Beyotime Institute of Biotechnology). Protein bands were visualized
using enhanced chemiluminescence kit (Sigma-Aldrich; Merck KGaA)
and imaged using a gel imaging system. The optical densities of the
protein bands were measured using Image-Pro Plus software (version
6.0; Media Cybernetics, Inc.).
Bioinformatics analysis
Overall survival of cervical cancer patients for
LINC00861 was done using gene expression profiling interactive
analysis (http://gepia2.cancer-pku.cn). The
cervical cancer patients were divided into high LINC00861
expression group (LINC00861 expression > median; n=145) and low
LINC00861 expression group (LINC00861 expression ≤ median; n=146).
Gene Set Enrichment Analysis (GSEA) (16) was performed on The Cancer Genome
Atlas (TCGA) database dataset (CESC; http://cancergenome.nih.gov/) of LINC00861 expression
by using R package clusterProfiler (17). Gene sets (h.all.v7.1.symbols.gmt)
were downloaded from the MSigDB database (https://www.gsea-msigdb.org/gsea/msigdb/genesets.jsp;
Broad Institute, Inc.).
Statistical analysis
All experiments were performed ≥3 times. Statistical
data are presented as the mean ± SD and statistical analysis was
performed using GraphPad Prism 7 software (GraphPad Software,
Inc.). A two-tailed paired Student's t-test was used to determine
the statistical differences in the LINC00861 expression levels
between cervical cancer and matched adjacent normal tissues. The
survival analysis was performed using a Kaplan-Meier test and a
log-rank test was used to determine the statistical significance
between the two groups. A χ2 test was performed to
analyze the association between LINC00861 expression levels and the
clinicopathological features of the patients with cervical cancer.
The remaining two group comparisons were performed using an
unpaired Student's t-test. The comparisons among ≥3 groups were
conducted using a one-way ANOVA followed by a Tukey's multiple
comparisons test. P<0.05 was considered to indicate a
statistically significant difference.
Results
LINC00861 expression levels are
significantly downregulated in cervical cancer tissues and are
associated with prognosis
To investigate the role of LINC00861 in cervical
cancer progression, RT-qPCR analysis was performed to analyze
LINC00861 expression levels in 56 cervical cancer and adjacent
normal tissues. LINC00861 expression levels were significantly
downregulated in the cervical cancer tissues compared with the
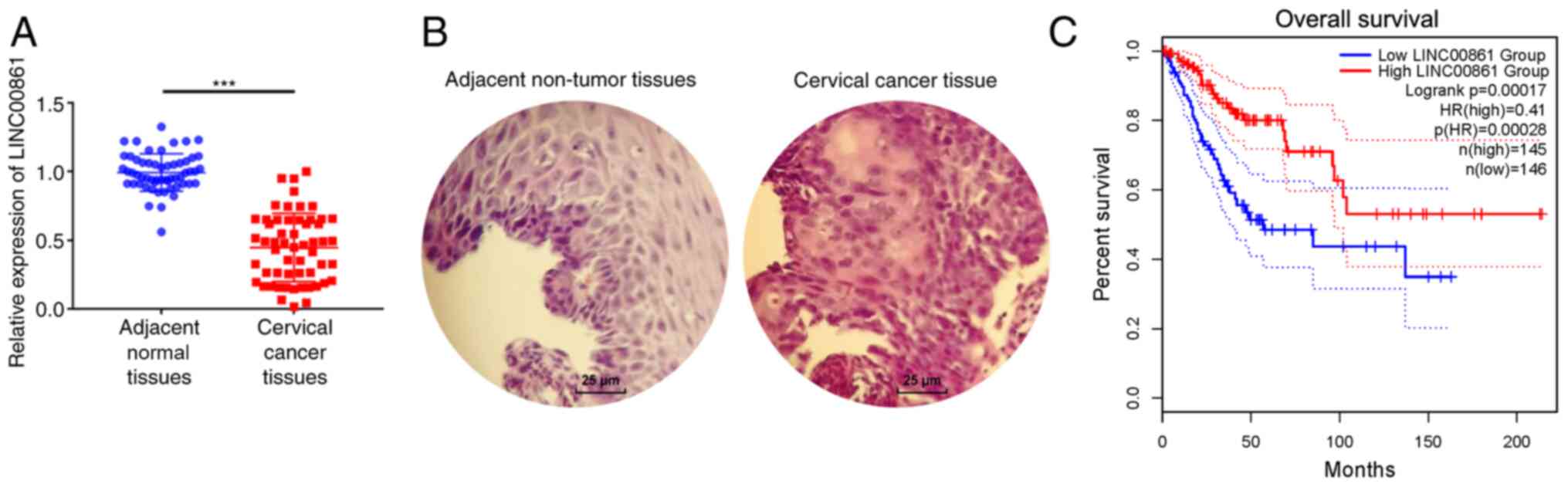
adjacent normal tissues (Fig. 1A).
In addition, the expression levels of LINC00861 were associated
with lymph-node metastasis and advanced tumor stage. No
associations were identified between LINC00861 levels and age,
tumor size, histology, and differentiation (Table III). Representative H&E
staining images are shown in Fig.
1B. Cervical cancer tissues displayed enlarged and
hyperchromatic nuclei, little cytoplasm, and atypical nuclei. TCGA
data indicated that patients with low LINC00861 expression levels
had a shorter survival time compared with patients with high
LINC00861 expression levels (Fig.
1C).
| Table III.Association between LINC00861
expression levels and the clinicopathological features of patients
with cervical cancer. |
Table III.
Association between LINC00861
expression levels and the clinicopathological features of patients
with cervical cancer.
|
|
| LINC00861
expression |
|
|---|
|
|
|
|
|
|---|
| Variable | n | High (n=28) | Low (n=28) | P-value |
|---|
| Age, years |
|
|
| 0.179 |
|
<45 | 25 | 15 | 10 |
|
|
≥45 | 31 | 13 | 18 |
|
| Tumor size, cm |
|
|
| 0.284 |
|
<4 | 26 | 11 | 15 |
|
| ≥4 | 30 | 17 | 13 |
|
| Histology |
|
|
| 0.313 |
|
Squamous cell carcinoma | 45 | 24 | 21 |
|
|
Adenocarcinoma | 11 | 4 | 7 |
|
|
Differentiation |
|
|
| 0.237 |
| Well +
moderate | 40 | 22 | 18 |
|
|
Poor | 16 | 6 | 10 |
|
| Lymph node
metastasis |
|
|
| 0.035a |
| No | 41 | 24 | 17 |
|
|
Yes | 15 | 4 | 11 |
|
| Tumor stage |
|
|
| 0.016a |
|
Ib-IIa | 29 | 19 | 10 |
|
|
IIb-IIIa | 27 | 9 | 18 |
|
Overexpression of LINC00861 attenuates
cervical cancer cell proliferation
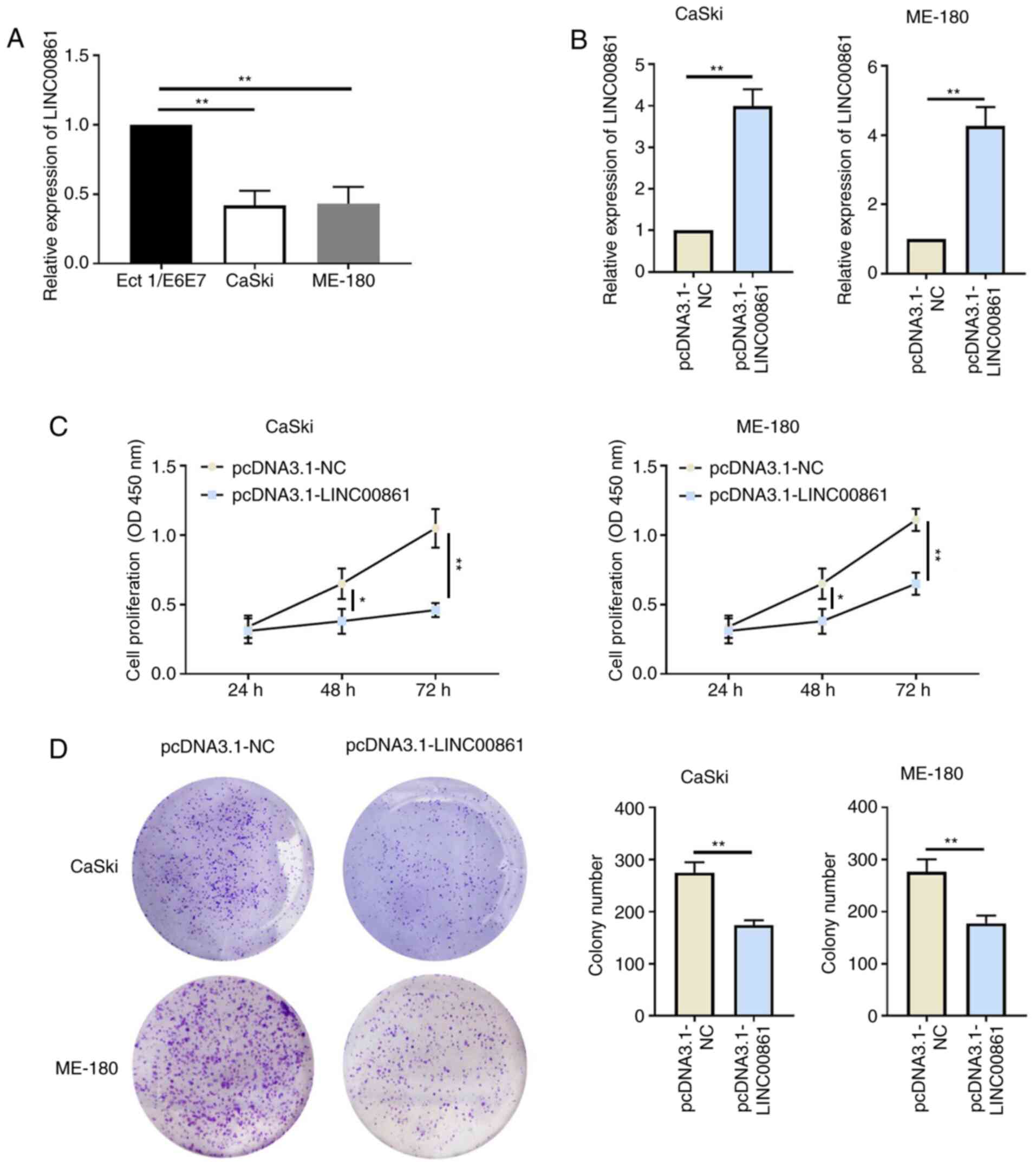
The expression levels of LINC00861 were
significantly downregulated in cervical cancer cells (CaSki and
ME-180) compared with the human cervical epithelial cell line,
Ect1/E6E7 (Fig. 2A). Cervical
cancer cells were subsequently transfected with either pcDNA3.1-NC
or pcDNA3.1-LINC00861 and the transfection efficiency was verified
using RT-qPCR (Fig. 2B). The
proliferative ability of the cervical cancer cells was
significantly attenuated following the overexpression of LINC00861
compared with the cells transfected with pcDNA3.1-NC, as determined
using CCK-8 and colony formation assays (Fig. 2C and D).
Overexpression of LINC00861 attenuates
cervical cancer cell migration and invasion
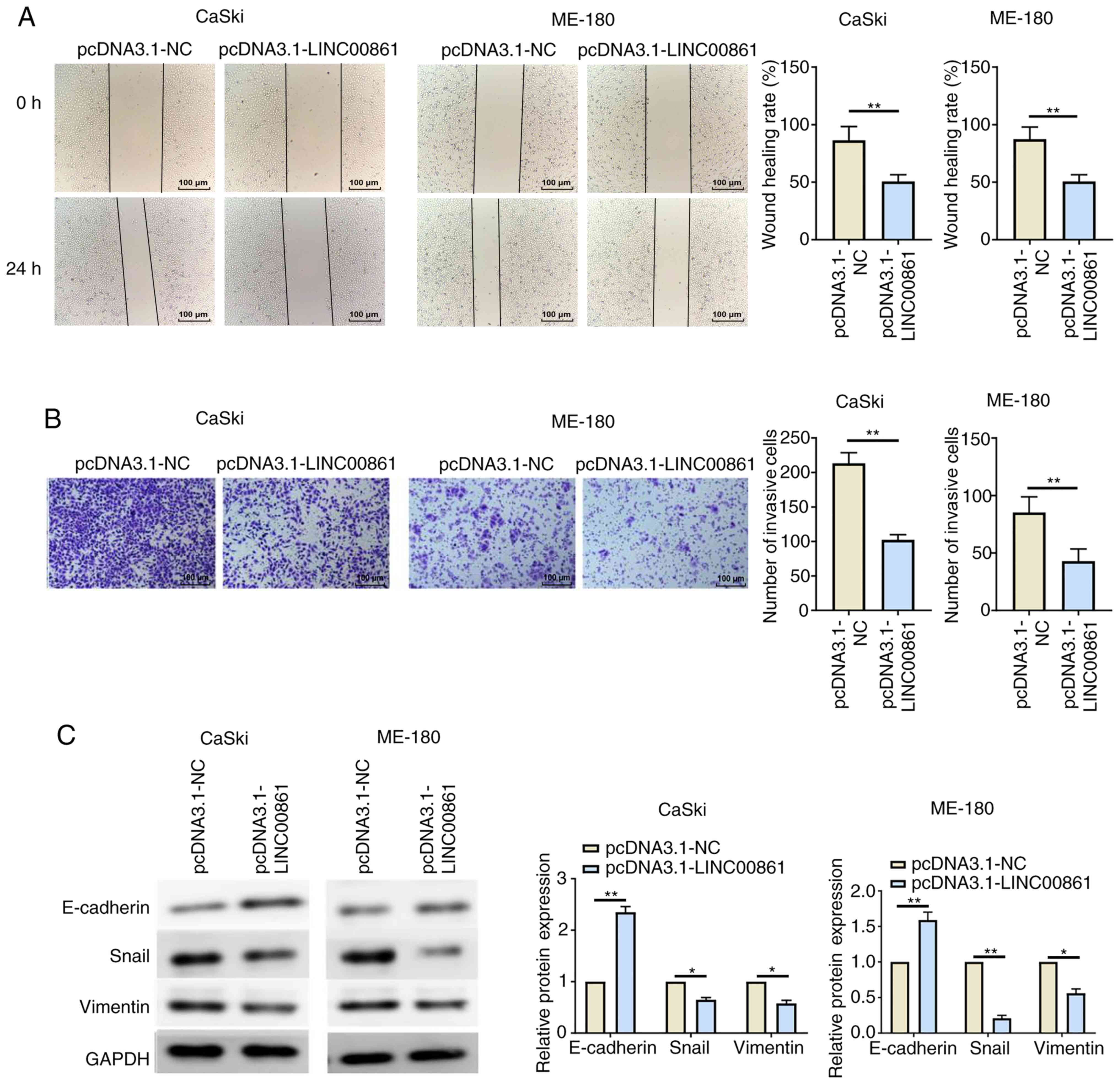
In addition to proliferation, the effects of
LINC00861 on migration and invasion were also investigated. Wound
healing and Transwell invasion assays revealed that the
transfection with pcDNA3.1-LINC00861 resulted in significantly
reduced cell migratory and invasive abilities compared with cells
transfected with pcDNA3.1-NC (Fig. 3A
and B). The expression levels of several EMT-related proteins
were further analyzed using western blotting. The results revealed
that the overexpression of LINC00861 led to the significant
upregulation of E-cadherin expression levels and the significant
downregulation of Snail and vimentin expression levels in both
cervical cancer cell lines compared with the
pcDNA3.1-NC-transfected cells (Fig.
3C).
LINC00861 competitively binds to
miR-51b-5p and regulates PTEN expression levels
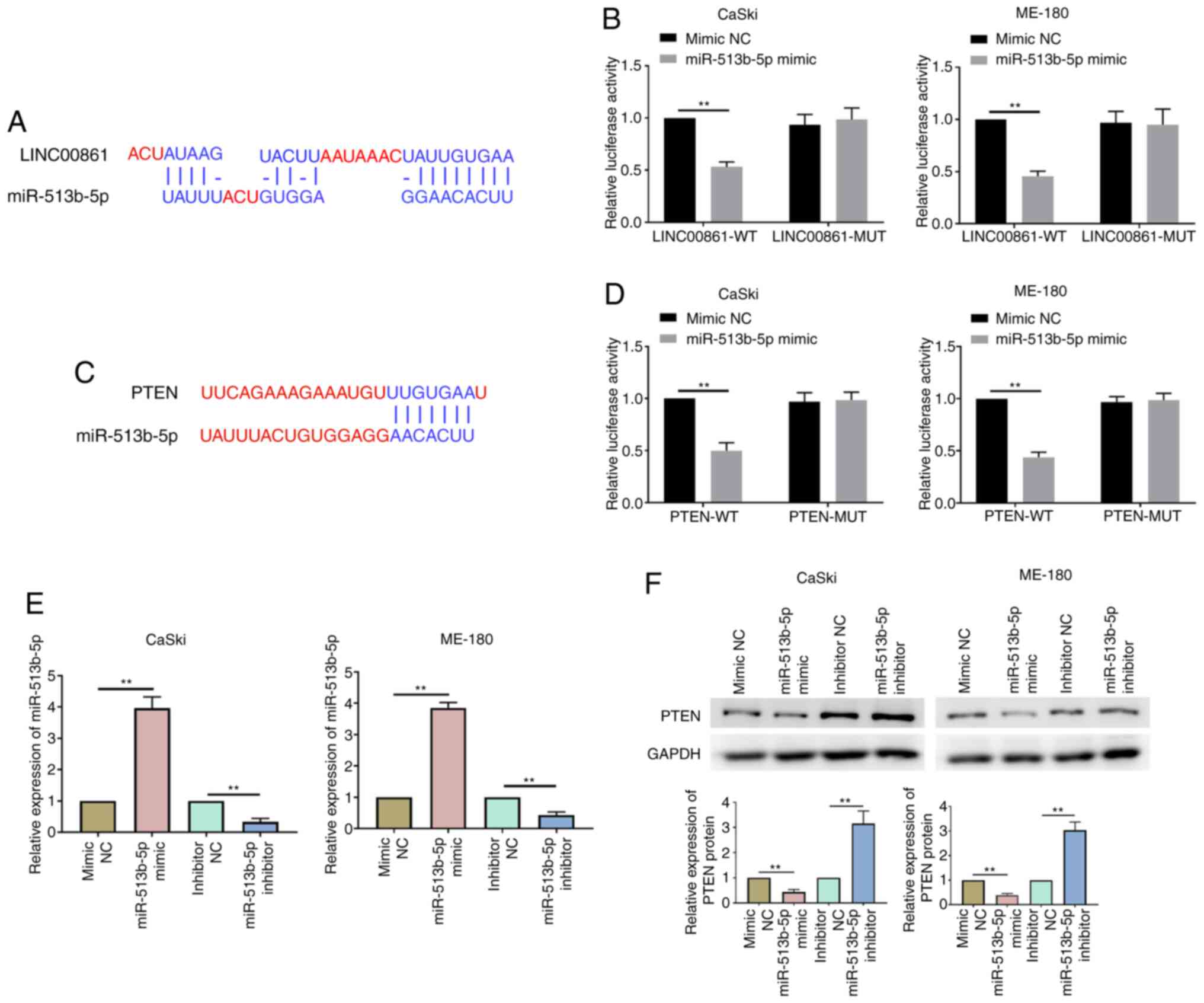
The online bioinformatics software, Diana Tools and
TargetScan, predicted that miR-513b-5p contained putative binding
sites within the 3′-untranslated region (UTR) of LINC00861 and PTEN
(Fig. 4A and C). The dual
luciferase reporter assay identified that the co-transfection with
the miR-513b-5p mimic could significantly decrease the relative
luciferase activity of the LINC0086-WT and PTEN-WT groups compared
with the co-transfection with the mimic NC (Fig. 4B and D). However, miR-513b-5p mimic
transfection had no effect on the luciferase activity of reporter
vector containing MUT 3′UTR of LINC0086 and PTEN. RT-qPCR analysis
demonstrated that the miR-513b-5p mimic significantly upregulated
miR-513b-5p expression levels, whereas the miR-513b-5p inhibitor
significantly downregulated the expression levels of miR-513b-5p in
cervical cancer cells compared with their respective NCs (Fig. 4E). Western blotting further revealed
that PTEN expression levels were significantly downregulated in the
miR-513b-5p mimic group compared with the mimic NC group in both
cell lines (Fig. 4F). Conversely,
the transfection with the miR-513b-5p inhibitor significantly
upregulated the expression levels of PTEN compared with the
inhibitor NC in both cell lines. These data indicated that
LINC00861 may serve as a ceRNA of miR-513b-5p in cervical cancer
progression.
Both LINC00861 and miR-513b-5p
overexpression modulate the PTEN/AKT/mTOR signaling pathway
To understand the underlying mechanism of LINC00861
in cervical cancer progression, GSEA was performed on TCGA cervical
cancer datasets. The results revealed that downregulation of
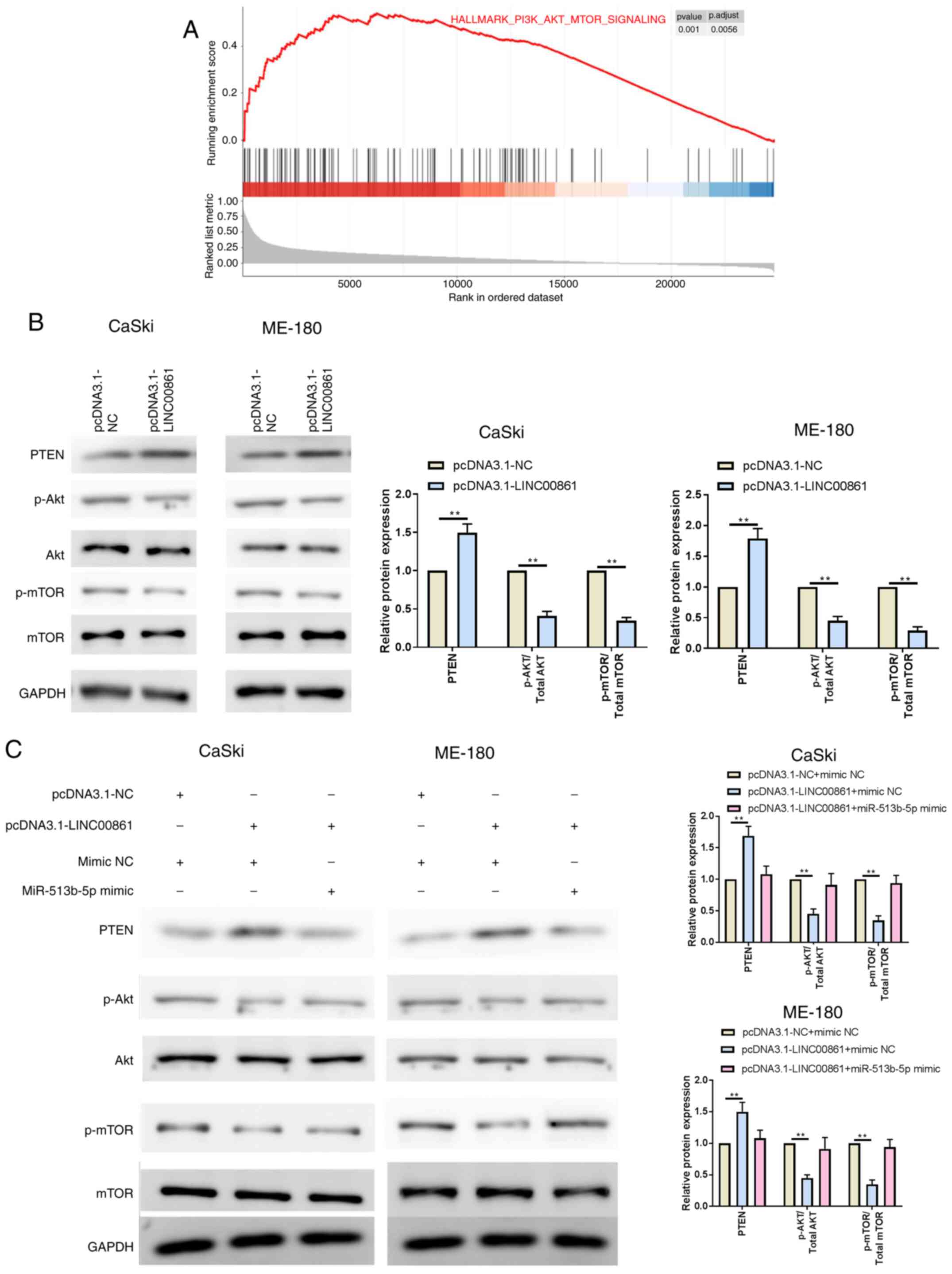
LINC00861 was associated with the PI3K/AKT/mTOR signaling (Fig. 5A), thereby indicating that the
PI3K/AKT/mTOR signaling pathway may be involved in the inhibition
effect of LINC00861 on proliferation, migration and invasion of
cervical cancer cells. The overexpression of LINC00861
significantly upregulated PTEN expression levels, while
downregulating p-AKT and p-mTOR expression levels, compared with
the pcDNA3.1-NC group in both cell lines (Fig. 5B). Cervical cancer cells were
subsequently co-transfected with LINC00861 and miR-513b-5p mimic;
as expected, the expression levels of PTEN, p-AKT and p-mTOR were
recovered following the co-transfection with pcDNA3.1-LINC00861 and
miR-513b-5p compared with the pcDNA3.1-LINC00861 + mimic NC group
in both cell lines (Fig. 5C). Thus,
it was suggested that the miR-513b-5p mimic may block the effect of
the overexpression of LINC00861.
Both LINC00861 and miR-513b-5p
overexpression modulate the EMT phenotype
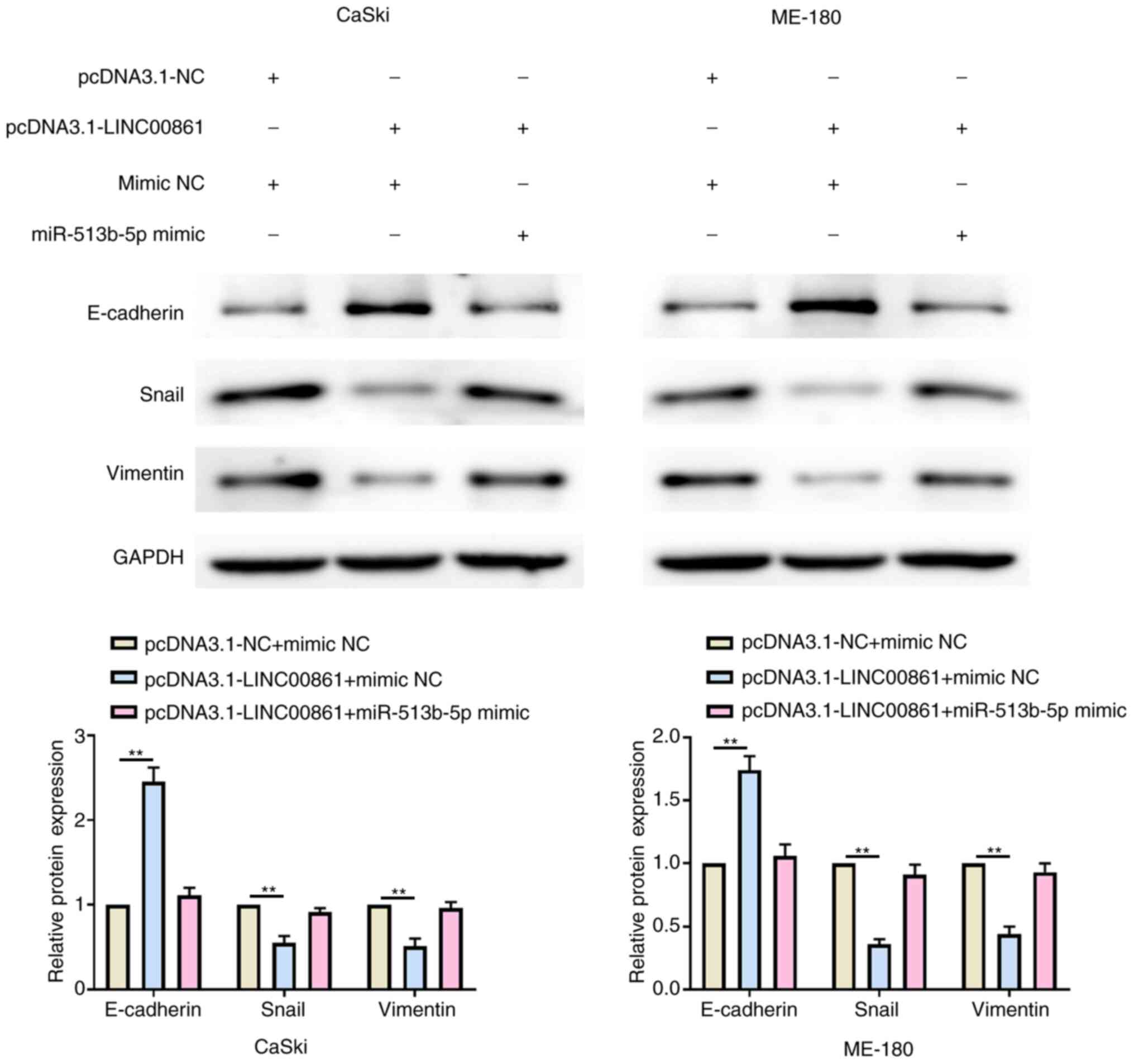
Western blotting revealed that the overexpression of
LINC00861 significantly upregulated E-cadherin expression levels
and downregulated Snail and vimentin expression levels compared
with the pcDNA3.1-NC + mimic NC group in both cell lines (Fig. 6). Conversely, the expression levels
of E-cadherin, Snail and vimentin were recovered following the
co-transfection with the pcDNA3.1-LINC00861 and miR-513b-5p mimic
in both cell lines.
Discussion
The abnormal expression of lncRNAs has been strongly
implicated in a wide range of cancer types, such as prostate
(18), colorectal (19) and cervical cancer (20). Several studies have reported that
lncRNAs could serve as either oncogenes or tumor suppressors
(21). For example, lncRNA nuclear
paraspeckle assembly transcript 1 functions as an oncogene to drive
aggressive endometrial cancer progression (22). LINC00589 and lncRNA maternally
expressed gene 3 are tumor suppressor (23,24).
Previous evidence has suggested that lncRNAs may serve as ceRNAs to
regulate miRNAs, which consequently regulates the expression of
target genes (25). For example,
the expression levels of LINC01234 were discovered to be
significantly upregulated in gastric cancer tissues and associated
with poor clinical outcomes (26).
In addition, the knockdown of LINC01234 inhibited cell
proliferation in vitro by serving as a ceRNA for miR-204-5p
and subsequently regulating core-binding factor subunit β
expression in gastric cancer (26).
Another previous study reported that the overexpression of lncRNA
LINC00673 promoted non-small cell lung cancer (NSCLC)
proliferation, migration, invasion and EMT by sponging miR-150-5p
(27). Furthermore, the lncRNA
proteasome 20S subunit α 3 antisense RNA 1 promoted esophageal
cancer progression by sponging miR-101 to upregulate Histone-lysine
N-methyltransferase EZH2 expression (28). In the present study, the expression
levels of LINC00861 were revealed to be significantly downregulated
in cervical cancer tissues, and were associated with an
advanced-stage, lymph node metastasis and poor survival in patients
with cervical cancer. In addition, the overexpression of LINC00861
was demonstrated to inhibit cervical cancer cell proliferation,
migration, invasion and EMT by functioning as a ceRNA for
miR-513b-5p and upregulating PTEN expression levels. Consistent
with these results, a recent report illustrated that
circ_La-related protein 4 (LARP4) inhibited cell proliferation and
migration in ovarian cancer by sponging miR-513b-5p to regulate the
expression of LARP4 (29). Wang
et al (30) reported that
miR-513b-5p promoted p53 protein expression by targeting interferon
regulatory factor 2, thereby suppressing testicular embryonal
carcinoma cell proliferation and inducing apoptosis in
vitro. It seems contradictory that miR-513b-5p may serve two
completely opposite roles in testicular embryonal carcinoma and
cervical cancer. In the present study, miR-513b-5p reversed the
inhibitory effects of LINC00861 on proliferation of cervical cancer
cells. However, several studies have revealed that the functions of
miRNAs may be specific to individual tissues; for example, miR-182
served an oncogenic role in endometrial carcinoma cells and
promoted cell proliferation, migration and invasion by targeting
forkhead box protein F2 (31).
Meanwhile, Lv et al (32)
demonstrated that miR-182 inhibited the proliferation of laryngeal
squamous cell carcinoma cells by targeting cisplatin
resistance-related protein 9. These two aforementioned studies
supported the findings of the present study, that is, that
miR-513b-5p may serve as an oncogenic miRNA in cervical cancer.
PTEN, an important tumor suppressor gene, is one of
the most commonly deleted or mutated tumor suppressors in a variety
of types of human cancer, including breast (33), prostate (34), and cervical cancer (35). PTEN inactivates the PI3K/AKT/mTOR
signaling pathway, which is involved in an abundance of essential
cellular processes, such as cell growth, differentiation,
proliferation and migration; most of these processes have also been
associated with cancer progression (36). For example, syntaxin binding protein
5-AS1 reduced cervical cancer cell proliferation and invasion by
regulating the miR-96-5p/PTEN axis (37). Wang et al (38) reported that long intergenic
non-protein coding RNA, p53 induced transcript alleviated the
progression of NSCLC by sponging miR-543 and subsequently inducing
PTEN expression. Shi et al (39) have suggested that the knockdown of
miR-17-5p may suppresses the malignant behavior of thyroid cancer
by upregulating PTEN expression levels and by inactivating the
AKT/mTOR signaling pathway. The findings of the present study
supported the results of previous studies.
Tumor metastasis is the cause of cancer mortality in
>90% of cases (40). The EMT
process can be hijacked by epithelial cancer cells to promote
abnormal migration and invasion (41). EMT is characterized by epithelial
cell markers such as E-cadherin are down-regulated, which leads to
loss of cell-cell adhesion, while mesenchymal markers such as
vimentin, Snail and N-cadherin are up-regulated, thereby allowing
the cells to migrate to different organs (42). Currently, numerous lncRNAs,
including lncRNAs pro-transition associated RNA and GATA binding
protein 6-AS1, have been reported to be involved in the EMT process
in ovarian and gastric cancer, respectively (43,44).
In the current study, the lncRNA LINC00861 was indicated to inhibit
the migration and invasion of cervical cancer cells by inhibiting
the EMT process of cervical cancer cells.
In conclusion, the results of the present study
indicated that LINC00861 overexpression may inhibit the
proliferation, migration and invasion of cervical cancer cells by
sponging miR-513b-5p, which subsequently regulated the
PTEN/AKT/mTOR signaling pathway. Therefore, the results suggested
that LINC00861 may serve as a tumor suppressor in the development
and progression of cervical cancer.
Acknowledgements
Not applicable.
Funding
No funding was received.
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
XS conceived and designed the study; HL, LZ and XD
performed the experiments, analyzed the data and wrote the
manuscript. All authors read and approved the final manuscript.
Ethics approval and consent to
participate
The experimental protocol involving patients was
approved by the Research Ethics Committee of Yantai Hospital of
Traditional Chinese Medicine (approval no. YTSZYYY2017-112). All
patients provided written informed consent for participation.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Bray F, Ferlay J, Soerjomataram I, Siegel
RL, Torre LA and Jemal A: Global cancer statistics 2018: GLOBOCAN
estimates of incidence and mortality worldwide for 36 cancers in
185 countries. CA Cancer J Clin. 68:394–424. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Cohen PA, Jhingran A, Oaknin A and Denny
L: Cervical cancer. Lancet. 393:169–182. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Arbyn M, Weiderpass E, Bruni L, de Sanjosé
S, Saraiya M, Ferlay J and Bray F: Estimates of incidence and
mortality of cervical cancer in 2018: A worldwide analysis. Lancet
Glob Health. 8:e191–e203. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Nagano T and Fraser P: No-nonsense
functions for long noncoding RNAs. Cell. 145:178–181. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Hu YW, Kang CM, Zhao JJ, Nie Y, Zheng L,
Li HX, Li X, Wang Q and Qiu YR: LncRNA PLAC2 down-regulates RPL36
expression and blocks cell cycle progression in glioma through a
mechanism involving STAT1. J Cell Mol Med. 22:497–510. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Yang L, Li Y, Gong R, Gao M, Feng C, Liu
T, Sun Y, Jin M, Wang D, Yuan Y, et al: The long non-coding
RNA-ORLNC1 regulates bone mass by directing mesenchymal stem cell
fate. Mol Ther. 27:394–410. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Hu X, Goswami S, Qiu J, Chen Q, Laverdure
S, Sherman BT and Imamichi T: Profiles of long non-coding RNAs and
mRNA expression in human macrophages regulated by interleukin-27.
Int J Mol Sci. 20:62072019. View Article : Google Scholar
|
|
8
|
Wang M, Zhou L, Yu F, Zhang Y, Li P and
Wang K: The functional roles of exosomal long non-coding RNAs in
cancer. Cell Mol Life Sci. 76:2059–2076. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Zhang F, Zhang L and Zhang C: Long
noncoding RNAs and tumorigenesis: Genetic associations, molecular
mechanisms, and therapeutic strategies. Tumour Biol. 37:163–175.
2016. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Zheng M, Hu Y, Gou R, Nie X, Li X, Liu J
and Lin B: Identification three LncRNA prognostic signature of
ovarian cancer based on genome-wide copy number variation. Biomed
Pharmacother. 124:1098102020. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Lv Y, Wei W, Huang Z, Chen Z, Fang Y, Pan
L, Han X and Xu Z: Long non-coding RNA expression profile can
predict early recurrence in hepatocellular carcinoma after curative
resection. Hepatol Res. 48:1140–1148. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Zhang Y, Li Y, Wang Q, Zhang X, Wang D,
Tang HC, Meng X and Ding X: Identification of an lncRNAmiRNAmRNA
interaction mechanism in breast cancer based on bioinformatic
analysis. Mol Med Rep. 16:5113–5120. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Shen CJ, Cheng YM and Wang CL: LncRNA PVT1
epigenetically silences miR-195 and modulates EMT and
chemoresistance in cervical cancer cells. J Drug Target.
25:637–644. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Li R, Liu J and Qi J: Knockdown of long
non-coding RNA CCAT1 suppresses proliferation and EMT of human
cervical cancer cell lines by down-regulating Runx2. Exp Mol
Pathol. 113:1043802020. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Subramanian A, Tamayo P, Mootha VK,
Mukherjee S, Ebert BL, Gillette MA, Paulovich A, Pomeroy SL, Golub
TR, Lander ES and Mesirov JP: Gene set enrichment analysis: A
knowledge-based approach for interpreting genome-wide expression
profiles. Proc Natl Acad Sci USA. 102:15545–15550. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Yu G, Wang LG, Han Y and He QY:
clusterProfiler: An R package for comparing biological themes among
gene clusters. OMICS. 16:284–287. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Hu R and Lu Z: Long non-coding RNA HCP5
promotes prostate cancer cell proliferation by acting as the sponge
of miR-4656 to modulate CEMIP expression. Oncol Rep. 43:328–336.
2020.PubMed/NCBI
|
|
19
|
Bai J, Xu J, Zhao J and Zhang R: LncRNA
NBR2 suppresses migration and invasion of colorectal cancer cells
by downregulating miRNA-21. Hum Cell. 33:98–103. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Liang H, Zhang C, Guan H, Liu J and Cui Y:
LncRNA DANCR promotes cervical cancer progression by upregulating
ROCK1 via sponging miR-335-5p. J Cell Physiol. 234:7266–7278. 2019.
View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Sanchez Calle A, Kawamura Y, Yamamoto Y,
Takeshita F and Ochiya T: Emerging roles of long non-coding RNA in
cancer. Cancer Sci. 109:2093–2100. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Dong P, Xiong Y, Yue J, Xu D, Ihira K,
Konno Y, Kobayashi N, Todo Y and Watari H: Long noncoding RNA NEAT1
drives aggressive endometrial cancer progression via
miR-361-regulated networks involving STAT3 and tumor
microenvironment-related genes. J Exp Clin Cancer Res. 38:2952019.
View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Zhang J, Li Z, Liu L, Wang Q, Li S, Chen
D, Hu Z, Yu T, Ding J, Li J, et al: Long noncoding RNA TSLNC8 is a
tumor suppressor that inactivates the interleukin-6/STAT3 signaling
pathway. Hepatology. 67:171–187. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Ghafouri-Fard S and Taheri M: Maternally
expressed gene 3 (MEG3): A tumor suppressor long non coding RNA.
Biomed Pharmacother. 118:1091292019. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Yao Y, Zhang T, Qi L, Zhou C, Wei J, Feng
F, Liu R and Sun C: Integrated analysis of co-expression and ceRNA
network identifies five lncRNAs as prognostic markers for breast
cancer. J Cell Mol Med. 23:8410–8419. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Chen X, Chen Z, Yu S, Nie F, Yan S, Ma P,
Chen Q, Wei C, Fu H, Xu T, et al: Long noncoding RNA LINC01234
functions as a competing endogenous RNA to regulate CBFB expression
by sponging miR-204-5p in gastric cancer. Clin Cancer Res.
24:2002–2014. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Lu W, Zhang H, Niu Y, Wu Y, Sun W, Li H,
Kong J, Ding K, Shen HM, Wu H, et al: Long non-coding RNA linc00673
regulated non-small cell lung cancer proliferation, migration,
invasion and epithelial mesenchymal transition by sponging
miR-150-5p. Mol Cancer. 16:1182017. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Qiu BQ, Lin XH, Ye XD, Huang W, Pei X,
Xiong D, Long X, Zhu SQ, Lu F, Lin K, et al: Long non-coding RNA
PSMA3-AS1 promotes malignant phenotypes of esophageal cancer by
modulating the miR-101/EZH2 axis as a ceRNA. Aging. 12:1843–1856.
2020. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Lin W, Ye H, You K and Chen L:
Up-regulation of circ_LARP4 suppresses cell proliferation and
migration in ovarian cancer by regulating miR-513b-5p/LARP4 axis.
Cancer Cell Int. 20:52020. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Wang X, Zhang X, Wang G, Wang L, Lin Y and
Sun F: Hsa-miR-513b-5p suppresses cell proliferation and promotes
P53 expression by targeting IRF2 in testicular embryonal carcinoma
cells. Gene. 626:344–353. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Yao H, Kong F and Zhou Y: MiR-182 promotes
cell proliferation, migration and invasion by targeting FoxF2 in
endometrial carcinoma cells. Int J Clin Exp Pathol. 12:1248–1259.
2019.PubMed/NCBI
|
|
32
|
Lv Y, Ye D, Qiu S, Zhang J, Shen Z, Shen Y
and Deng H: MiR-182 regulates cell proliferation and apoptosis in
laryngeal squamous cell carcinoma by targeting the CRR9. Biosci
Rep. 39:BSR201913482019. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Ngeow J, Sesock K and Eng C: Breast cancer
risk and clinical implications for germline PTEN mutation carriers.
Breast Cancer Res Treat. 165:1–8. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Patel R, Brzezinska EA, Repiscak P, Ahmad
I, Mui E, Gao M, Blomme A, Harle V, Tan EH, Malviya G, et al:
Activation of β-catenin cooperates with loss of Pten to drive
AR-independent castration-resistant prostate cancer. Cancer Res.
80:576–590. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Kaur G, Balasubramaniam SD and Lee YJ:
IGFBP-2 in cervical cancer development. Exp Mol Pathol. 113:212020.
View Article : Google Scholar
|
|
36
|
Li A, Qiu M, Zhou H, Wang T and Guo W:
PTEN, insulin resistance and cancer. Curr Pharm Des. 23:3667–3676.
2017. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Shao S, Wang C, Wang S, Zhang H and Zhang
Y: LncRNA STXBP5-AS1 suppressed cervical cancer progression via
targeting miR-96-5p/PTEN axis. Biomed Pharmacother. 117:152019.
View Article : Google Scholar
|
|
38
|
Wang S, Jiang W, Zhang X, Lu Z, Geng Q,
Wang W, Li N and Cai X: LINC-PINT alleviates lung cancer
progression via sponging miR-543 and inducing PTEN. Cancer Med.
9:1999–2009. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Shi YP, Liu GL, Li S and Liu XL: miR-17-5p
knockdown inhibits proliferation, autophagy and promotes apoptosis
in thyroid cancer via targeting PTEN. Neoplasma. 67:249–258. 2020.
View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Fu BM: Tumor metastasis in the
microcirculation. Adv Exp Med Biol. 1097:201–218. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Ombrato L and Malanchi I: The EMT
universe: Space between cancer cell dissemination and metastasis
initiation. Crit Rev Oncog. 19:349–361. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Odero-Marah V, Hawsawi O, Henderson V and
Sweeney J: Epithelial-mesenchymal transition (EMT) and prostate
cancer. Adv Exp Med Biol. 1095:101–110. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Liang H, Yu T, Han Y, Jiang H, Wang C, You
T, Zhao X, Shan H, Yang R, Yang L, et al: LncRNA PTAR promotes EMT
and invasion-metastasis in serous ovarian cancer by competitively
binding miR-101-3p to regulate ZEB1 expression. Mol Cancer.
17:1192018. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Li ZT, Zhang X, Wang DW, Xu J, Kou KJ,
Wang ZW, Yong G, Liang DS and Sun XY: Overexpressed lncRNA
GATA6-AS1 inhibits LNM and EMT via FZD4 through the Wnt/β-catenin
signaling pathway in GC. Mol Ther Nucleic Acids. 19:827–840. 2019.
View Article : Google Scholar : PubMed/NCBI
|