Introduction
Multiple myeloma (MM) is a type of hematological
malignancy characterized by malignant clonal hyperplasia of plasma
cells in the bone marrow, often resulting in susceptibility to
infection, impaired renal function, anemia and hypercalcemia
(1). According to research
statistics, the incidence of MM is the second highest to
non-Hodgkin's lymphoma (2), and the
incidence of MM accounts for ~13% of all hematological tumors
(3). With the development of
molecular biology technology, progress has been made in broadening
the understanding of MM pathogenesis and in the development of new
drugs (4). In particular,
gene-targeted therapy has long been the focus of the study and
treatment of cancer (5). However,
there are no curative therapies for MM. It is therefore urgent to
develop new and more effective treatment methods (6,7).
Therefore, further understanding of the molecular mechanism
underlying MM progression may provide new insight into the
development of novel therapeutic strategies.
MicroRNAs (miRNAs) are non-coding small RNAs ~22
nucleosides in length, which serve important roles in the
regulation of gene expression (8).
miRNAs participate in regulation of gene expression by directly
binding to the 3′-untranslated region (UTR) of specific target
genes, thereby affecting cell differentiation, development,
apoptosis, proliferation and other biological activities (9–11).
Recent evidence has indicated that miRNAs are dysregulated in
several types of tumor and could modulate tumor progression
(12,13). For instance, previous studies
demonstrated that miR-450a inhibited the metastasis of ovarian
cancer cells (14), and that
miR-365-3p could promote metastasis, cancer stem cell-like
properties and drug resistance in oral squamous cells (15). Moreover, miR-140-5p is considered a
tumor suppressor for various types of tumor, including in
retinoblastoma and ovarian cancer (16–18).
Another previous study also indicated that miR-140-5p upregulation
could inhibit autophagy and chemoresistance of myeloma cells
(16). However, whether miR-140-5p
regulates the growth and metastasis of MM remains unknown.
Vascular endothelial growth factor A (VEGFA) is a
key regulator of tumor growth. As a result, VEGFA represents a
potential therapeutic target in cancer treatment (19). Integrated genomic analysis indicated
that VEGFA was commonly overexpressed in adenocarcinoma (20). Notably, VEGFA expression is
correlated with the survival of patients with MM; for instance
patients whose MM cells express low levels of VEGFA have the
longest survival (21). VEGFA is
the target of multiple miRNAs, such as miR-29c in lung
adenocarcinoma (22), and
miR-199a-5p in endometrial mesenchymal stem cells (23). A previous study demonstrated that
miR-16 targeted a large number of downstream target genes,
including VEGFA in MM (24).
However, whether miR-140-5p can also target VEGFA in MM is
unclear.
Therefore, the aim of the present study was to
explore the function served by miR-140-5p in MM and to determine
whether the potential tumor suppressive effects of miR-140-5p in MM
are mediated through VEGFA.
Materials and methods
Cell culture
Human normal plasma cells were separated from the
peripheral blood of three men and three women with a median age of
47 years using flow cytometry (25). All donors were healthy volunteers
who had not previously received any drugs associated with
immunological diseases. The blood (2 ml/person) was collected at
the Jingzhou Central Hospital between 2016.03 and 2017.03. Informed
consent was provided by all volunteers, and the present study was
approved by the Ethics Committee of Jingzhou Central Hospital
(approval no. JCH20150608EZ023). Briefly, peripheral blood
mononuclear cells were separated by Ficoll®-Hypaque
centrifugation from peripheral blood. Plasma cells were isolated
from mononuclear blood cells using CD138 microbeads (Miltenyi
Biotec, Inc.) according to the manufacturer's instructions
(26). The isolated plasma cells
were then cultured. The RPMI 8226, OPM2, U266 (TIB-196), H929,
MM1.S and KMS11 MM cell lines were purchased from the American Type
Culture Collection. RPMI-1640 medium (Gibco; Thermo Fisher
Scientific, Inc.) containing 10% fetal bovine serum (Gibco; Thermo
Fisher Scientific, Inc.) and 1% dual antibiotics (penicillin 100
U/ml; streptomycin 0.1 mg/m1; Sigma-Aldrich; Merck KGaA) was used
to culture all cells at 37°C with 5% CO2.
Transfection
The RPMI 8226 and U266 cell lines were used for
transfection experiments, since these two cell lines displayed low
miR-140-5p expression. Cells were seeded on a 6-well plate at a
density of 1×106 cells/ml and cultured overnight at
37°C. The next day, cells were transfected with miR-140-5p mimic or
a VEGFA overexpression plasmid. A total of 100 pmol of the
miR-140-5p mimic (sense: 5′-ACCAUAGGGUAAAACCACUGUU-3′), mimic
control (5′-UUGUACUACACAAAAGUACUG-3′), VEGFA overexpression plasmid
(VEGFA-pcDNA3.1) and VEGFA negative control (NC) were synthesized
by Sangon Biotech Co., Ltd.. The miR-140-5p mimic, VEGFA-pcDNA3.1
plasmid or their respective controls (100 pmol), were then
dissolved in 50 µl Opti-MEM (HyClone; GE Healthcare Life Sciences).
In addition, 1 µl Lipofectamine® 2000 (Invitrogen;
Thermo Fisher Scientific, Inc.) transfection reagent was then
separately diluted in 50 µl DMEM, then incubated for 5 min at room
temperature. After incubation, the nucleic acid was mixed with the
diluted Lipofectamine® and the mixture was added to the
cells. Cells were cultured at 37°C with 5% CO2 and the
medium was replaced after 24 h. Then, 72 h after changing the
medium, the cells were harvested for subsequent experiments.
Luciferase activity assay
The VEGFA 3′-UTR containing putative miR-140-5p
binding sites was inserted into a pmirGLO dual luciferase vector
(Promega Corporation) to generate wild-type (WT) pmirGLO-VEGFA. The
mutant (MUT) 3′-UTR of VEGFA was inserted into the pmirGLO
luciferase reporter vector at the XhoI/XbaI sites.
RPMI 8226 and U266 cells were co-transfected with pmirGLO vector
containing the WT or MUT VEGFA 3′-UTR and miR-140-5p mimic using
Lipofectamine® 2000 (Invitrogen; Thermo Fisher
Scientific, Inc.). After incubation for 48 h (27), relative luciferase activity in the
cells was assessed using the Dual-Luciferase Reporter Assay kit
(Promega Corporation), according to the manufacturer's protocol.
The Renilla luciferase activity was used for reference.
Reverse transcription-quantitative PCR
(RT-qPCR)
Total RNA was extracted from cells using
TRIzol® reagent (Invitrogen; Thermo Fisher Scientific,
Inc.). The concentration of RNA was measured using a NanoDrop™
spectrophotometer (Thermo Fisher Scientific, Inc.), then diluted to
500 ng/µl. For reverse transcription, 1 µg total RNA was converted
into cDNA using the Superscript II First-Strand cDNA Synthesis kit
(Invitrogen; Thermo Fisher Scientific, Inc.). The mRNA expression
levels were determined with SYBR-Green PCR Master Mix (Thermo
Fisher Scientific, Inc) using the 7500 Real-Time PCR system (Thermo
Fisher Scientific, USA). The thermocycling conditions were as
follows: i) Initial denaturation at 95°C for 1 min; ii) 40 cycles
at 95°C for 30 sec, 58°C for 20 sec, and 70°C for 20 sec; and iii)
a final extension at 72°C for 7 min. The samples were then
maintained at 4°C. The expression levels in the RT-qPCR products
were determined using the 2−ΔΔCq method (28). All primer sequences are listed in
Table I.
 | Table I.Primers used for reverse
transcription-quantitative PCR. |
Table I.
Primers used for reverse
transcription-quantitative PCR.
| Target | Primer
sequence | Species |
|---|
| miR-140-5p | Forward,
5′-GAGTGTCAGTGGTTTTACCCT-3′ |
|
|
| Reverse,
5′-GCAGGGTCCGAGGTATTC-3′ | Human |
| VEGFA | Forward,
5′-GGCCTCCGAAACCATGAACT-3′ |
|
|
| Reverse,
5′-TCGTGATGATTCTGCCCTCC-3′ | Human |
| U6 | Forward,
5′-TGACTTCCAAGTACCATCGCCA-3′ |
|
|
| Reverse,
5′-TTGTAGAGGTAGGTGTGCAGCAT-3′ | Human |
| GAPDH | Forward,
5′-GGTGAAGGTCGGAGTCAACG-3′ |
|
|
| Reverse,
5′-CAAAGTTGTCATGGATGTACC-3′ | Human |
Cell viability detection
Cells were seeded into a 96-well plate
(4×104), and 10 µl Cell Counting Kit-8 (CCK-8; cat. no.
96992, Sigma-Aldrich, Merck KGaA) solution was added into each
well. Cells were then incubated for 4 h at 37°C. Absorbance was
read at 450 nm using a Multiskan GO multiplate reader (Thermo
Fisher Scientific, Inc.) Data were recorded as the average of
triplicate wells for each condition.
Cell apoptosis
Cells were harvested 24 h after transduction and
re-suspended at 1×106 cells/ml in 1X Annexin binding
buffer (cat. no. 422201; BioLegend, Inc.). Next, 5 µl of
fluorescein isothiocyanate (FITC)-conjugated Annexin V (cat. no.
C1062S, Beyotime Institute of Biotechnology) and 1 µl of 100 µg/ml
propidium iodide (cat. no. P1304MP; Thermo Fisher Scientific, Inc.)
solution in 300 µl 1X Annexin binding buffer were added to the cell
suspension for 15 min at room temperature. Stained samples were
acquired using a FACSCalibur flow cytometer (BD Biosciences) and
analyzed using FlowJo version 10.0 (FlowJo LLC).
Wound healing assay
Transfected cells were plated at a density of
5×105 cells/well on a 6-well plate. Cells were allowed
to adhere to the surface of the well for 24 h, then, a
uniform-width scratch was quickly made on the cell layer. The
suspended cells were removed, and the remainder was cultured at low
serum concentration (1%). Cell migration across the scratch was
recorded at 0 and 48 h (light microscope; magnification, ×100), and
the migration distance was measured using ImageJ software version
1.8.0 (National Institutes of Health). To eliminate the differences
in initial width at 0 h among the different experimental groups, a
relative migration rate was calculated as (0 h scratch width-48 h
scratch width)/0 h scratch width × 100%. In order to ensure that
the same location was observed at 0 and 48 h, a cross symbol was
drawn at within the scratches and images obtained at both
time-points were aligned with these signs.
Transwell Matrigel™ assay
Cells were harvested 24 h after transfection and
re-suspended in FBS-free medium at a concentration of
1×106 cells/ml. The cell suspension was then added into
the upper chamber of Transwell plates (Corning Inc.) pre-coated
with Matrigel™ (Thermo Fisher Scientific, Inc.). Medium containing
10% FBS serum was added into the lower chamber. After a 24-h
incubation at 37°C, the migratory cells were stained using 0.5%
crystal violet at room temperature for 15 min, while those
remaining in the upper chamber were removed using a cotton swab.
Invading cells were counted under a light microscope, magnification
×200.
Western blotting
Total protein was extracted from cells using RIPA
buffer (cat. no. 9806; Cell Signaling Technology, Inc.) then boiled
for 5 min at 100°C for denaturation. Protein concentration was
examined using bicinchoninic acid Assay kit (Thermo Fisher
Scientific, Inc.). Proteins (20 µg/lane) were then separated using
12% SDS-PAGE, and then transferred to polyvinylidene fluoride
membranes (EMD Millipore). The membranes were blocked with 5% milk
at room temperature for 1 h. The blots were then probed with
primary antibodies against Ki-67 (rabbit; 1:1,000; product code
ab16667), cyclin D1 (rabbit; 1:1,000; product code ab16663),
vimentin (rabbit; 1:1,000; product code ab193555), Snail (rabbit;
1:1,000; product code ab229701), matrix metalloproteinase (MMP)-2
(rabbit; 1:1,000; product code ab215986), MMP3 (rabbit; 1:1,000;
product code ab52915) and GAPDH (rabbit; 1:2,000; product code
ab181602) at 4°C overnight. All primary antibodies were obtained
from Abcam. The membranes were then washed with PBS three times,
then incubated with a horseradish peroxidase-conjugated Goat
Anti-Rabbit IgG (H+L) secondary antibody (cat. no. SA00001-2;
1:2,000; ProteinTech Group, Inc.) at 37°C for 2 h. The protein
bands were detected using the ECL western blotting kit (cat. no.
93-K820-500, Multi Sciences; http://www.liankebio.com/index.html) and analyzed
using ImageJ version 4.7 (National Institutes of Health).
Statistical analysis
Statistical analysis was carried out using GraphPad
Prism version 6.01 (GraphPad Software, Inc.). Data are presented as
the mean ± SD of at least three independent experiments. Student's
t-test was used for pairwise comparisons between the means of
continuous variables. Multi-group comparisons were carried out
using ANOVA, followed by Bonferroni correction. P<0.05 was
considered to indicate a statistically significant difference.
Results
miR-140-5p expression in MM cell lines
and its effect on the viability and apoptosis of MM cells
The expression of miR-140-5p in MM cells was
significantly lower than in normal plasma cells (P<0.001;
Fig. 1A). The U266 and RPMI 8226
cells were transfected with a miR-140-5p mimic, resulting in a
significant increase in miR-140-5p expression, compared with the
mimic control and blank control cells (P<0.001; Fig. 1B). Moreover, overexpression of
miR-140-5p significantly decreased the viability of U266 and RPMI
8226 cells over time (P<0.001; Fig.
1C and D). Transfection with the miR-140-5p mimic also resulted
in a significant increase in cell apoptosis (P<0.001; Fig. 1E and F).
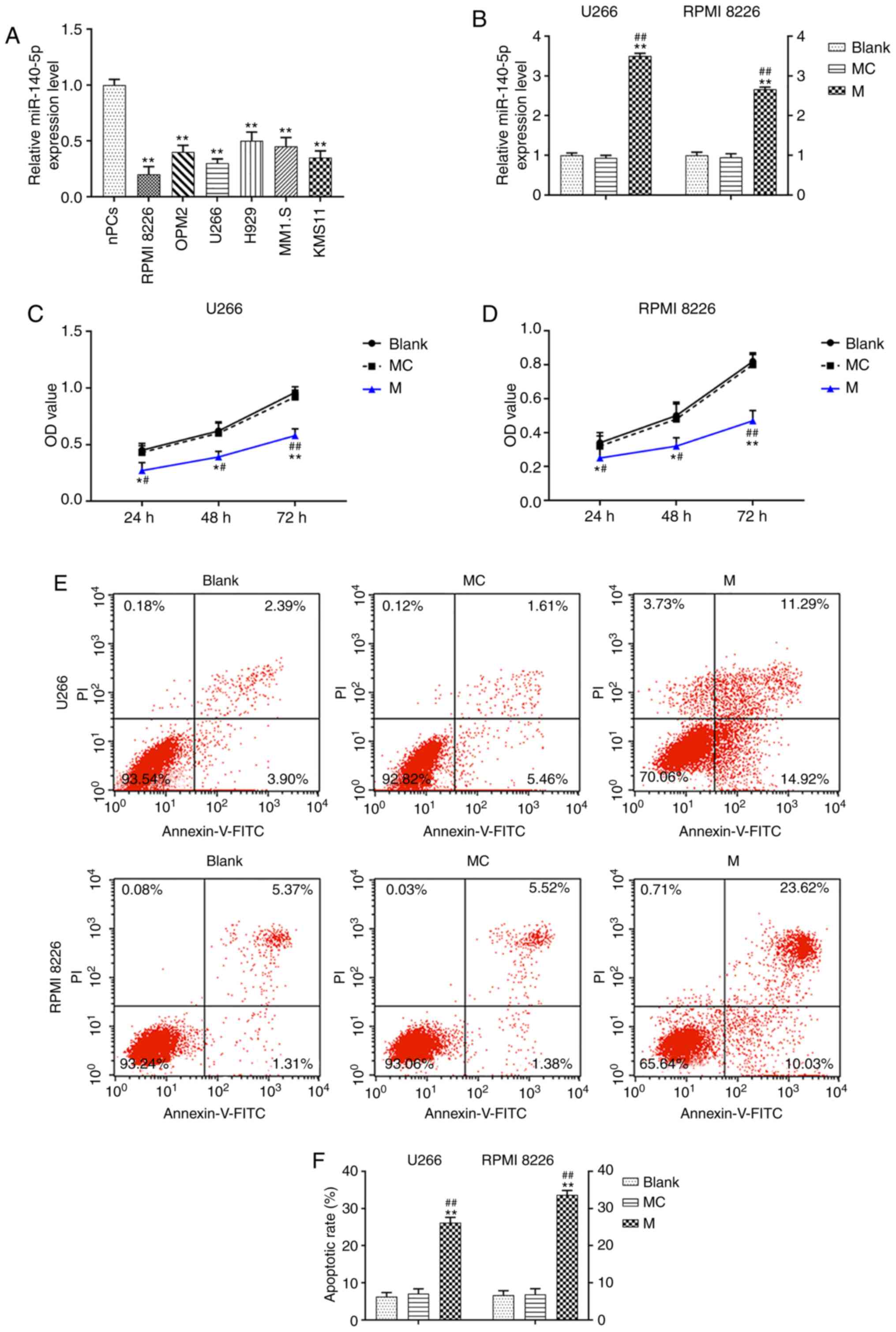 | Figure 1.miR-140-5p overexpression affects the
viability and apoptosis of MM cells. (A) RT-qPCR was used to detect
the expression of miR-140-5p in MM cell lines and nPCs. n=3.
**P<0.001, vs. nPCs. (B) Transfection efficiency of miR-140-5p
in U266 and RPMI 8226 cell lines was detected by RT-qPCR. U6 was
the internal reference. (C and D) Cell Counting Kit-8 was used to
evaluate the effect of miR-140-5p transfection on cell viability.
(E and F) Cell apoptosis was detected by flow cytometry. n=3.
*P<0.05, **P<0.001, vs. Blank; #P<0.05,
##P<0.001, vs. MC. MM, multiple myeloma; miR,
microRNA; MC, mimic control; M, mimic; nPCs, normal plasma cells;
RT-qPCR, reverse transcription-quantitative PCR; OD, optical
density; PI, propidium iodide; FITC, fluorescein
isothiocyanate. |
Effects of miR-140-5p overexpression
on the migration and invasion of MM cells
In order to evaluate cell migration in U266 and RPMI
8226 cells following transfection with miR-140-5p, a wound healing
assay was carried out. Overexpression of miR-140-5p significantly
inhibited cell migration, compared with the mimic control and blank
cells (P<0.001; Fig. 2A and B).
A Transwell Matrigel™ assay was also performed to assess cell
invasion. Transfection with miR-140-5p resulted in significant
inhibition of cell invasion, compared with the mimic control and
blank cells (P<0.001; Fig. 2C and
D).
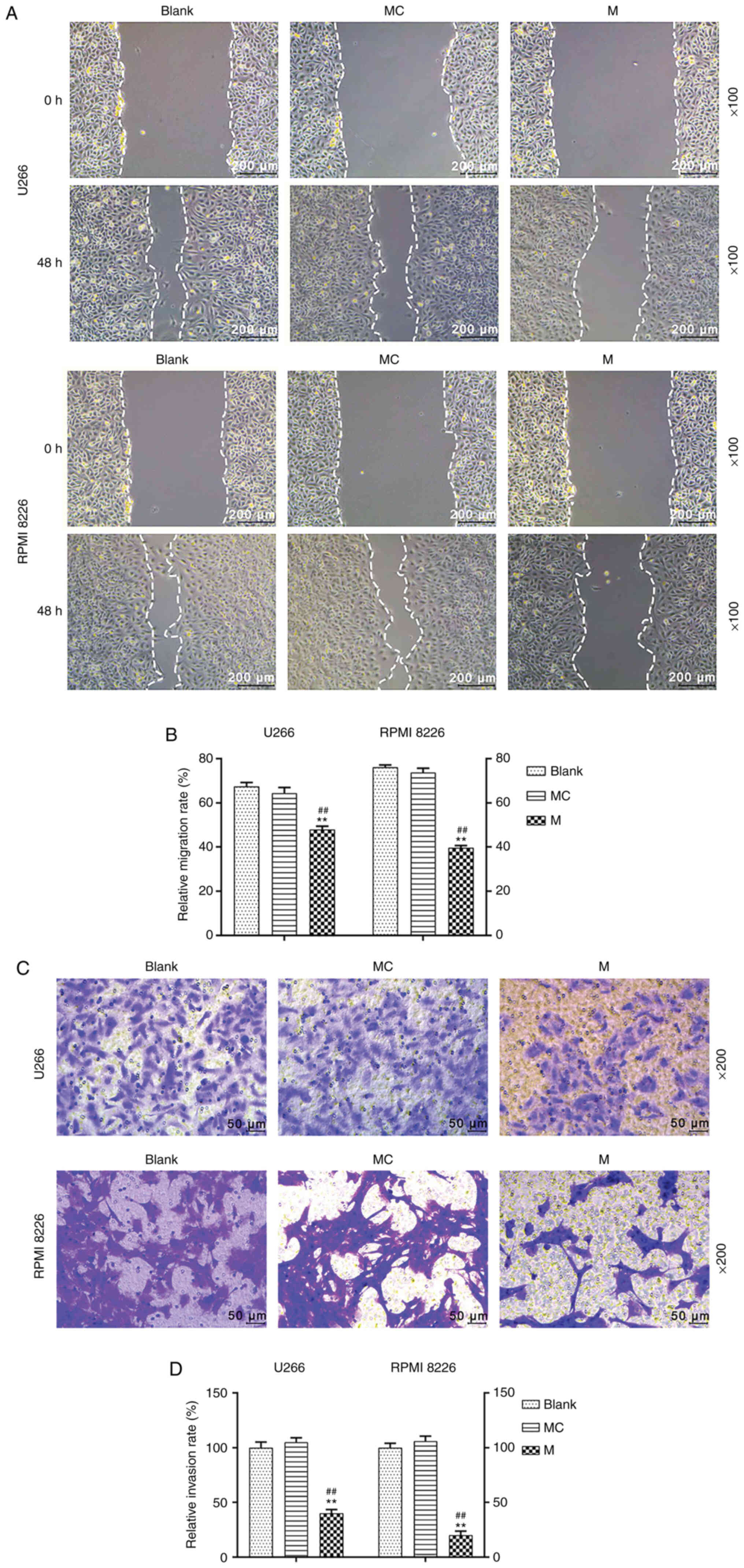 | Figure 2.Effects of miR-140-5p overexpression
on the migration and invasion of MM cells. (A and B) Cell migration
was evaluate using a wound healing assay. Scale bar, 200 µm.
Magnification, ×100. (C and D) Cell invasion was assessed using a
Transwell Matrigel™ assay. Scale bar, 50 µm. Magnification, ×200.
n=3. **P<0.001, vs. Blank; ##P<0.001, vs. MC. miR,
microRNA; MM, multiple myeloma; MC, mimic control; M, mimic. |
Effects of miR-140-5p overexpression
on the expression levels of genes related to proliferation,
migration and invasion
To determine the mechanism of miR-140-5p in MM
cells, western blot assays were performed to determine the
expression levels of proteins related to the proliferation,
migration and invasion of U266 and RPMI 8226 cells. After
miR-140-5p mimic had been transfected into the U266 and RPMI 8226
cells, the results revealed that overexpressed miR-140-5p inhibited
the expression levels of Ki-67, cyclin D1, vimentin, Snail, MMP2
and MMP3 in U266 cells (P<0.001, Fig. 3A-D).
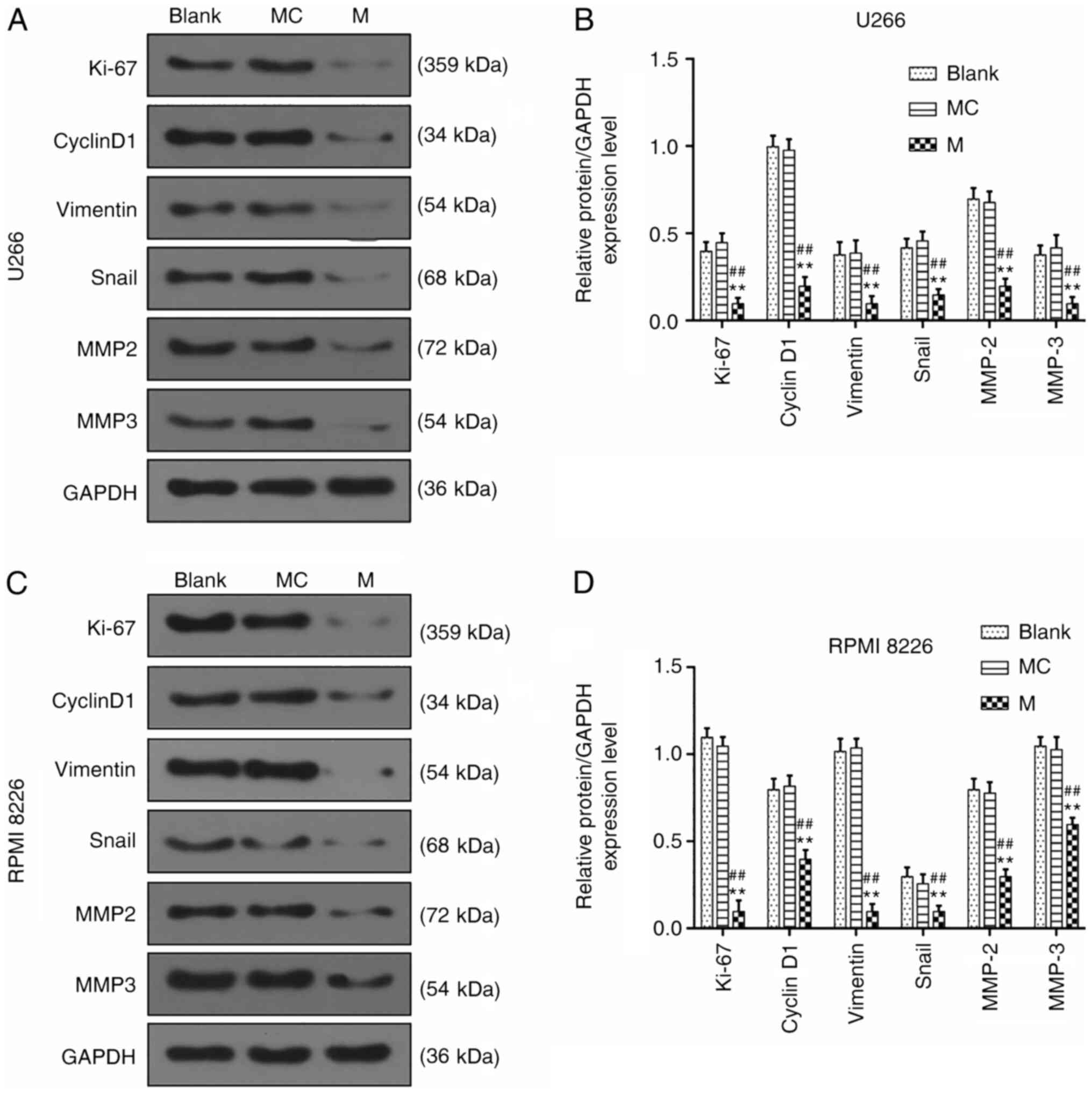 | Figure 3.Effects of miR-140-5p overexpression
on the expression of genes related to proliferation, migration and
invasion. (A and B) Western blotting was performed to detect the
expression levels of Ki-67, cyclin D1, vimentin, Snail, MMP2 and
MMP3 in U266 cells. (C and D) Western blotting was used to detect
the expression levels of Ki-67, cyclin D1, vimentin, Snail, MMP2
and MMP3 in RPMI 8226 cells. n=3. **P<0.001, vs. Blank;
##P<0.001, vs. MC. miR, microRNA; MC, mimic control;
M, mimic; MMP, matrix metalloproteinase. |
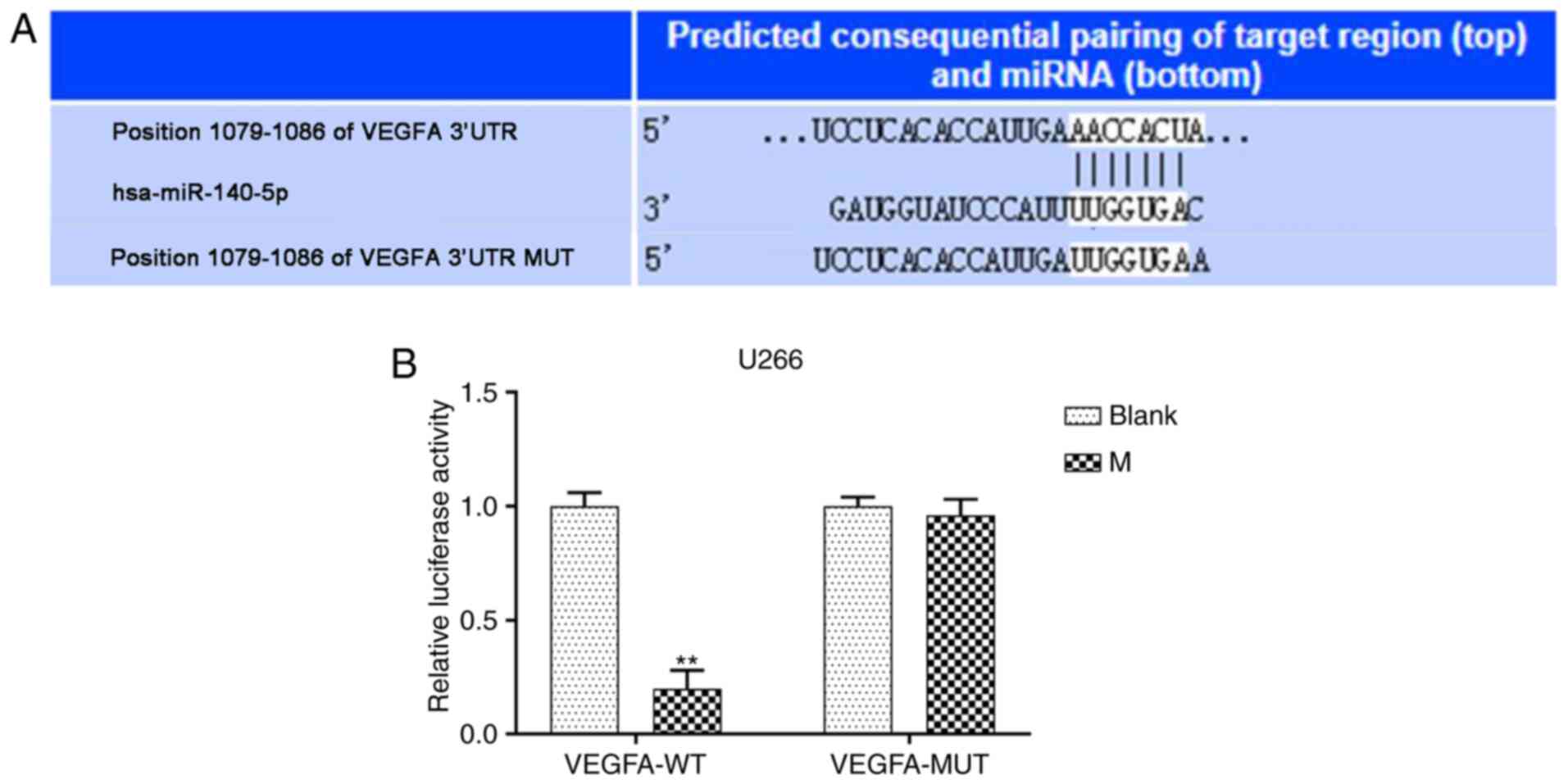
VEGFA directly targets miR-140-5p in
MM cell lines
TargetScan software predicted that VEGFA was the
potential target gene of miR-140-5p (Fig. 4A). Furthermore, pmirGLO dual
luciferase reporter vectors containing VEGFA-WT or VEGFA-MUT 3-UTR
sequences were constructed, and co-transfected with miR-140-5p
mimic into MM cell lines. Relative luciferase activity following
co-transfection with VEGFA-WT and mimic significantly decreased,
compared with the blank, while VEGFA-MUT had no effect (P<0.001,
Fig. 4B). Moreover, transfection
VEGFA-pcDNA3.1 increased VEGFA expression following U266 and RPMI
8226 cell transfection. However, co-transfection of VEGFA-pcDNA3.1
with the miR-140-5p mimic significantly reduced VEGFA expression,
compared with transfection with VEGFA-pcDNA3.1 alone (P<0.001;
Fig. 5A and B).
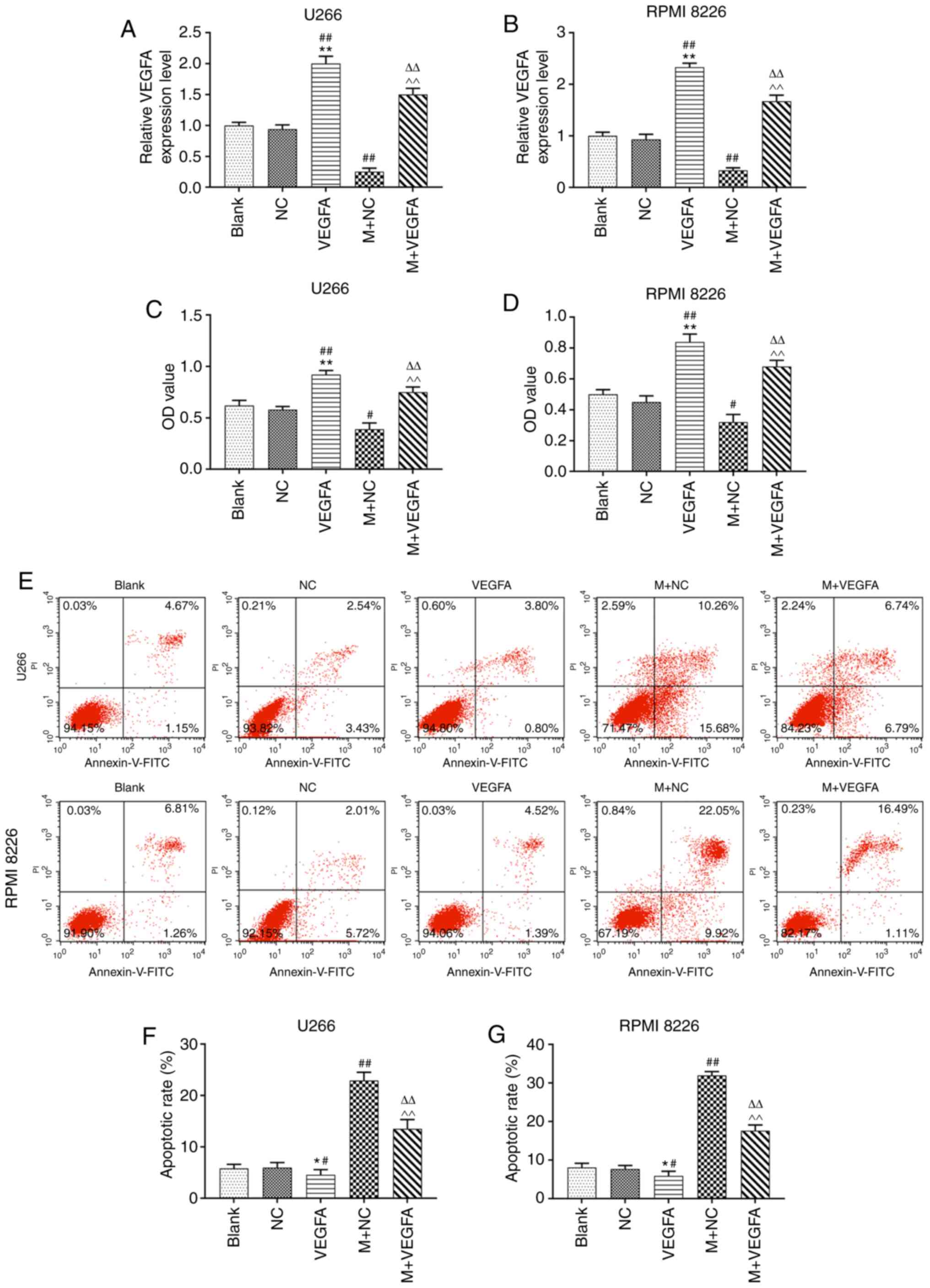 | Figure 5.Effects of miR-140-5p and VEGFA the
on viability and apoptosis of MM cell lines. (A and B) Reverse
transcription-quantitative PCR was used to detect the expression of
VEGFA in U266 and RPMI 8226 cells. GAPDH was the internal
reference. (C and D) Cell Counting Kit-8 was used to assess the
effect of VEGFA and miR-140-5p on cell viability. (E-G) Cell
apoptosis was evaluated using flow cytometry. n=3. *P<0.05,
**P<0.001, vs. Blank; #P<0.05,
##P<0.001, vs. NC; ^^P<0.001, vs.
VEGFA, ΔΔP<0.001, vs. mimic + NC. VEGFA, vascular
endothelial growth factor A; miR, microRNA; MM, multiple myeloma;
M, mimic; NC, negative control; OD, optical density; PI, propidium
iodide; FITC, fluorescein isothiocyanate. |
Effects of miR-140-5p on viability and
apoptosis of MM cell lines by targeting VEGFA
VEGFA overexpression led to a significant increase
in cell viability. However, co-transfection of VEGFA-pcDNA3.1 with
the miR-140-5p mimic significantly reduced cell viability, compared
with transfection with VEGFA-pcDNA3.1 alone (P<0.001; Fig. 5C and D). Moreover, apoptosis was
significantly reduced following VEGFA overexpression, but increased
following co-transfection with by VEGFA-pcDNA3.1 and the miR-140-5p
mimic (P<0.001, Fig. 5E-G).
Effects of miR-140-5p and VEGFA on the
migration and invasion of MM cell lines
MM cell lines were transfected with VEGFA-pcDNA3.1,
alone or with miR-140-5p mimic into the cells, and cell migration
and invasion were assessed. VEGFA overexpression promoted cell
migration, compared with blank cells. However, co-transfection with
VEGFA-pcDNA3.1 and miR-140-5p mimic reduced the migration rate,
compared with VEGFA overexpression alone (P<0.001; Fig. 6A-D). Moreover, overexpression of
VEGFA was also promoted cell invasion, but this effect was reversed
following co-transfection with the miR-140-5p mimic (P<0.001,
Fig. 6E-G).
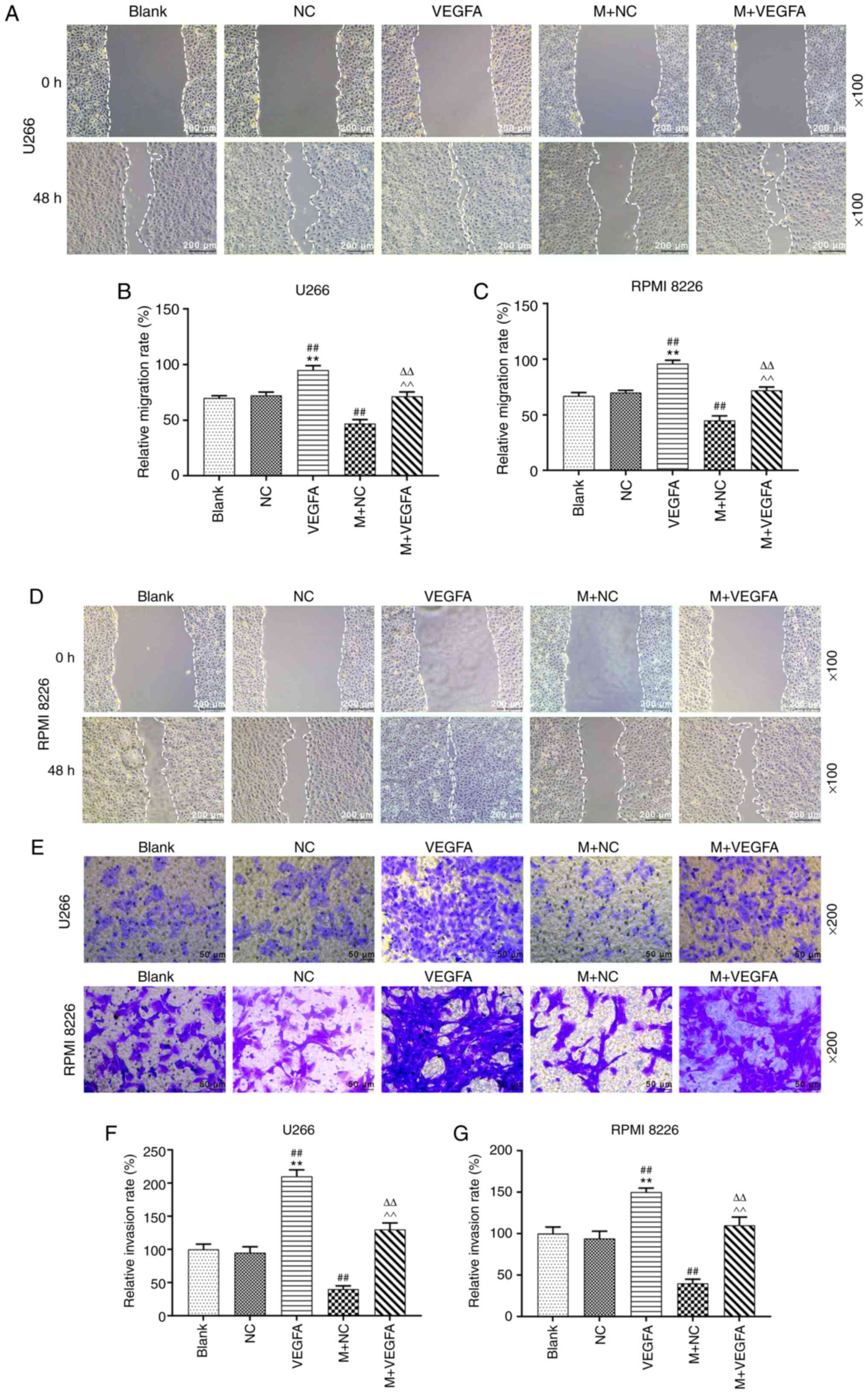 | Figure 6.Effects of miR-140-5p and VEGFA on
migration and invasion of MM cell lines. (A and B) U266 cell
migration was measured using a wound healing scratch assay. (C and
D) RPMI 8226 cell migration was measured using a wound healing
assay. Scale bar, 200 µm. Magnification, ×100. (E-G) Cell invasion
was detected using a Transwell Matrigel™ assay. Scale bar, 50 µm.
Magnification, ×200. n=3. **P<0.001, vs. Blank;
##P<0.001, vs. NC; ^^P<0.001, vs.
VEGFA, ΔΔP<0.001, vs. mimic + NC. MiR, microRNA;
VEGFA, vascular endothelial growth factor A; MM, multiple myeloma;
M, mimic; NC, negative control. |
Discussion
MM often results in susceptibility to infection,
impaired renal function, anemia and hypercalcemia, which severely
impacts on patient quality of life (29,30).
To date, the pathogenesis of MM remains unclear, and specific
treatment avenues for MM are still the focus of research (31). Previous studies have indicated that
miRNAs are aberrantly expressed in different types of tumor, and
that these molecules could act as tumor suppressor genes through
genetic regulation of cell proliferation, migration, cell cycle and
differentiation during tumorigenesis (12,32).
For instance, previous studies demonstrated that miR-196-b-5p and
miR-99-a-5p served important roles in the regulation of autophagy
and apoptosis of MM cells (33),
while miR-125-b and miR-34a affected their viability (34). Moreover, miR-720, miR-1246, miR-16
and miR-25 could serve as biological markers in MM prognostic
evaluation (35,36). Therefore, miRNA function represents
an important research area in the study of MM pathogenesis.
miR-140-5p is a newly discovered small-molecule
coding RNA. Previous studies indicated that miR-140-5p played an
important regulatory role in human gastric cancer (37), glioma (38) and non-small cell lung cancer
(39). Another study demonstrated
that miR-140-5p could regulate vitamin D levels by targeting MAPK,
which provided new insight into the treatment of bone diseases
(40). In addition, miR-140-5p
could be involved in autophagy and drug resistance mechanisms in MM
(41). However, the specific role
of miR-140-5p in the occurrence and development of MM still remains
unclear. Thus, the present study aimed to explore the role of
miR-140-5p in MM.
miR-140-5p is significantly downregulated in gastric
cancer (37) and breast cancer
(42), compared with normal
tissues. Similarly, in the present study, the expression of
miR-140-5p in MM cell lines was significantly lower than in normal
plasma cells. In a previous study, high miR-140-5p expression
promoted cell viability of human osteosarcoma cells decreased by
tumor drugs (43), and
overexpressing miR-140-5p could promote apoptosis in chronic
myeloid leukemia (44).
Furthermore, the migration and invasion of lung cancer cells
significantly decreased following miR-140-5p overexpression
(39). In the present study, MM
cell lines were transfected with a miR-140-5p in order to further
investigate the biological effects of miR-140-5p overexpression on
MM. miR-140-5p overexpression significantly reduced the viability
and migration of MM cells, while increasing apoptosis. These
results indicated that miR-140-5p could regulate MM progression,
and that overexpression of miR-140-5p inhibited the development of
MM.
miRNAs are known to bind to the 3′-UTR of specific
target genes to jointly regulate tumorigenesis and tumor
development (10). miR-140-5p binds
several target genes and regulates the progression of numerous
diseases. For example, miR-140-5p inhibited the growth of ovarian
cancer by targeting platelet-derived growth factor receptor α
(17). In addition, miR-140-5p
could also promote proliferation and inhibit apoptosis of human
pulmonary artery smooth muscle cells during hypoxia by targeting
DNA methyltransferase 1 (45).
Based on these previous findings, it was determined that miR-140-5p
bound to a gene to jointly regulate the disease progression of MM.
Targetscan7.2 software predicted that VEGFA was a possible target
gene for miR-140-5p, and a dual luciferase assay also indicated
that VEGFA interacted with miR-140-5p directly. Moreover,
transfection with a VEGFA overexpression plasmid along with a
miR-140-5p mimic confirmed that miR-140-5p overexpression could
inhibit the expression of VEGFA in MM cells.
VEGFA can increase microvascular permeability, and
promote the proliferation, vascular construction and the migration
of endothelial cells from different tissues (46). The VEGFA signaling pathway is an
important target for the inhibition of tumor angiogenesis (47). Studies demonstrated that miR-140-5p
could inhibit cell proliferation, migration and invasion of lung
cancer and colorectal cancer cells by targeting VEGFA (48,49).
In the present study, overexpression of VEGFA could promote cell
viability, invasion and migration, and inhibit cell apoptosis.
Moreover, miR-140-5p overexpression partially reversed this effect
and promoted cell viability, migration and invasion, while
suppressing apoptosis. Consistent with previous studies, these
results indicated that miR-140-5p could regulate MM progression
through VEGFA (48).
In conclusion, the present study demonstrated that
miR-140-5p suppressed MM progression by targeting VEGFA, and
suggests that miR-140-5p could be a potential therapeutic target
for the treatment of MM. However, a limitation of the present study
was the lack of animal experiments and thus these results require
further validation in vivo.
Acknowledgements
Not applicable.
Funding
No funding was received.
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
ML and HL made substantial contributions to the
conception and design of the study. HL, JZ and ZY acquired,
analyzed and interpreted the data. ML and HL drafted the article
and critically revised it for important intellectual content. All
authors agree to be accountable for all aspects of the work in
ensuring that questions related to the accuracy or integrity of the
work are appropriately investigated and resolved. All authors
approved the final version of the manuscript.
Ethics approval and consent to
participate
The present study was approved by the Ethics
Committee of Jingzhou Central Hospital (approval no.
JCH20150608EZ023). All procedures involving human participants were
in accordance with the ethical standards of the institutional
and/or national research committee and with the 1964 Helsinki
declaration and its later amendments or comparable ethical
standards. All volunteers provided informed consent.
Patient consent for participation
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
Glossary
Abbreviations
Abbreviations:
|
MM
|
multiple myeloma
|
|
RT-qPCR
|
reverse transcription-quantitative
PCR
|
|
miR
|
MicroRNA
|
|
VEGFA
|
vascular endothelial growth factor
A
|
|
CCK-8
|
cell counting kit-8
|
|
RPMI
|
Roswell Park Memorial Institute
|
|
NC
|
negative control
|
|
WT
|
wildtype
|
|
MUT
|
mutant
|
|
UTR
|
untranslated region
|
|
FITC
|
fluorescein isothiocyanate
|
References
|
1
|
Röllig C, Knop S and Bornhäuser M:
Multiple myeloma. Lancet. 385:2197–2208. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Siegel RL, Miller KD and Jemal A: Cancer
statistics, 2017. CA Cancer J Clin. 67:7–30. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Ghobrial IM, Siegel DS, Vij R, Berdeja JG,
Richardson PG, Neuwirth R, Patel CG, Zohren F and Wolf JL: TAK-228
(formerly MLN0128), an investigational oral dual TORC1/2 inhibitor:
A phase I dose escalation study in patients with relapsed or
refractory multiple myeloma, non-Hodgkin lymphoma, or Waldenstrom's
macroglobulinemia. Am J Hematol. 91:400–405. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Agarwal A and Mahadevan D: Novel targeted
therapies and combinations for the treatment of multiple myeloma.
Cardiovasc Hematol Disord Drug Targets. 13:2–15. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Hao S, Du X, Song Y, Ren M, Yang Q, Wang
A, Wang Q, Zhao H, Du Z and Zhang G: Targeted gene therapy of the
HSV-TK/hIL-12 fusion gene controlled by the hSLPI gene promoter of
human non-small cell lung cancer in vitro. Oncol Lett.
15:6503–6512. 2018.PubMed/NCBI
|
|
6
|
Morris LG and Chan TA: Therapeutic
targeting of tumor suppressor genes. Cancer. 121:1357–1368. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Ling H, Fabbri M and Calin GA: MicroRNAs
and other non-coding RNAs as targets for anticancer drug
development. Nat Rev Drug Discov. 12:847–865. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Mohr AM and Mott JL: Overview of microRNA
biology. Semin Liver Dis. 35:3–11. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Liang S, Zhang N, Deng Y, Chen L, Zhang Y,
Zheng Z, Luo W, Lv Z, Li S and Xu T: miR-663 promotes NPC cell
proliferation by directly targeting CDKN2A. Mol Med Rep.
16:4863–4870. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Shi J: Regulatory networks between
neurotrophins and miRNAs in brain diseases and cancers. Acta
Pharmacol Sin. 36:149–157. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Petri R and Jakobsson J: Identifying miRNA
targets using AGO-RIPseq. Methods Mol Biol. 1720:131–140. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Trionfini P and Benigni A: MicroRNAs as
master regulators of glomerular function in health and disease. J
Am Soc Nephrol. 28:1686–1696. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Orso F, Quirico L, Dettori D, Coppo R,
Virga F, Ferreira LC, Paoletti C, Baruffaldi D, Penna E and Taverna
D: Role of miRNAs in tumor and endothelial cell interactions during
tumor progression. Semin Cancer Biol. 60:214–224. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Muys BR, Sousa JF, Plaça JR, de Araújo LF,
Sarshad AA, Anastasakis DG, Wang X, Li XL, de Molfetta GA, Ramão A,
et al: miR-450a acts as a tumor suppressor in ovarian cancer by
regulating energy metabolism. Cancer Res. 79:3294–3305. 2019.
View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Huang WC, Jang TH, Tung SL, Yen TC, Chan
SH and Wang LH: A novel miR-365-3p/EHF/keratin 16 axis promotes
oral squamous cell carcinoma metastasis, cancer stemness and drug
resistance via enhancing β5-integrin/c-met signaling pathway. J Exp
Clin Cancer Res. 38:892019. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Miao X, Wang Z, Chen B, Chen Y, Wang X,
Jiang L, Jiang S, Hao K and Zhang W: miR-140-5p suppresses
retinoblastoma cell proliferation, migration, and invasion by
targeting CEMIP and CADM3. Cell Mol Biol (Noisy-le-grand).
64:42–47. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Lan H, Chen W, He G and Yang S: miR-140-5p
inhibits ovarian cancer growth partially by repression of PDGFRA.
Biomed Pharmacother. 75:117–122. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Zhang D, Yang Y, Wu M, Zhao X, Sun Y, Xie
H, Li H, Li Y, Wang K, Zhang J, et al: The moderating effect of
social support on the relationship between physical health and
suicidal thoughts among Chinese rural elderly: A nursing home
sample. Int J Ment Health Nurs. 27:1371–1382. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Ferrara N and Adamis AP: Ten years of
anti-vascular endothelial growth factor therapy. Nat Rev Drug
Discov. 15:385–403. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Cancer Genome Atlas Research Network;
Analysis Working Group; Asan University; BC Cancer Agency; Brigham
and Women's Hospital; Broad Institute; Brown University; Case
Western Reserve University; Dana-Farber Cancer Institute; Duke
University, et al, . Integrated genomic characterization of
oesophageal carcinoma. Nature. 541:169–175. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Botta C, Di Martino MT, Ciliberto D, Cucè
M, Correale P, Rossi M, Tagliaferri P and Tassone P: A gene
expression inflammatory signature specifically predicts multiple
myeloma evolution and patients survival. Blood Cancer J.
6:e5112016. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Liu L, Bi N, Wu L, Ding X, Men Y, Zhou W,
Li L, Zhang W, Shi S, Song Y and Wang L: MicroRNA-29c functions as
a tumor suppressor by targeting VEGFA in lung adenocarcinoma. Mol
Cancer. 16:502017. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Hsu CY, Hsieh TH, Tsai CF, Tsai HP, Chen
HS, Chang Y, Chuang HY, Lee JN, Hsu YL and Tsai EM: miRNA-199a-5p
regulates VEGFA in endometrial mesenchymal stem cells and
contributes to the pathogenesis of endometriosis. J Pathol.
232:330–343. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Sun CY, She XM, Qin Y, Chu ZB, Chen L, Ai
LS, Zhang L and Hu Y: miR-15a and miR-16 affect the angiogenesis of
multiple myeloma by targeting VEGF. Carcinogenesis. 34:426–435.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Liu D, Lin P, Hu Y, Zhou Y, Tang G, Powers
L, Medeiros LJ, Jorgensen JL and Wang SA: Immunophenotypic
heterogeneity of normal plasma cells: Comparison with minimal
residual plasma cell myeloma. J Clin Pathol. 65:823–829. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Horst A, Hunzelmann N, Arce S, Herber M,
Manz RA, Radbruch A, Nischt R, Schmitz J and Assenmacher M:
Detection and characterization of plasma cells in peripheral blood:
Correlation of IgE+ plasma cell frequency with IgE serum titre.
Clin Exp Immunol. 130:370–378. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Liang B, Yin JJ and Zhan XR: MiR-301a
promotes cell proliferation by directly targeting TIMP2 in multiple
myeloma. Int J Clin Exp Pathol. 8:9168–9174. 2015.PubMed/NCBI
|
|
28
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Fakhri B and Vij R: Clonal evolution in
multiple myeloma. Clin Lymphoma Myeloma Leuk. 16 (Suppl):S130–S134.
2016. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Kiely F, Cran A, Finnerty D and O'Brien T:
Self-reported quality of life and symptom burden in ambulatory
patients with multiple myeloma on disease-modifying treatment. Am J
Hosp Palliat Care. 34:671–676. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Kuroda J and Chinen Y: Multiple myeloma:
Pathophysiology and progress in management. Rinsho Ketsueki.
58:487–497. 2017.PubMed/NCBI
|
|
32
|
Hayes J, Peruzzi PP and Lawler S:
MicroRNAs in cancer: Biomarkers, functions and therapy. Trends Mol
Med. 20:460–469. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Shang J, Chen ZZ, Wang ZH, Wei TN, Wu WB
and Chen WM: Association of miRNA-196b-5p and miRNA-99a-5p with
autophagy and apoptosis in multiple myeloma cells. Zhonghua Xue Ye
Xue Za Zhi. 39:766–772. 2018.(In Chinese). PubMed/NCBI
|
|
34
|
Abdi J, Rastgoo N, Li L, Chen W and Chang
H: Role of tumor suppressor p53 and micro-RNA interplay in multiple
myeloma pathogenesis. J Hematol Oncol. 10:1692017. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Ren Y, Li X, Wang W, He W, Wang J and Wang
Y: Expression of peripheral blood miRNA-720 and miRNA-1246 can be
used as a predictor for outcome in multiple myeloma patients. Clin
Lymphoma Myeloma Leuk. 17:415–423. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Rocci A, Hofmeister CC and Pichiorri F:
The potential of miRNAs as biomarkers for multiple myeloma. Expert
Rev Mol Diagn. 14:947–959. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Fang Z, Yin S, Sun R, Zhang S, Fu M, Wu Y,
Zhang T, Khaliq J and Li Y: miR-140-5p suppresses the
proliferation, migration and invasion of gastric cancer by
regulating YES1. Mol Cancer. 16:1392017. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Yang HL, Gao YM and Zhao JA: miR-140-5p
inhibits human glioma cell growth and invasion by targeting JAG1.
Mol Med Rep. 16:3634–3640. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Flamini V, Jiang WG and Cui Y: Therapeutic
role of MiR-140-5p for the treatment of non-small cell lung cancer.
Anticancer Res. 37:4319–4327. 2017.PubMed/NCBI
|
|
40
|
Luo W, Liu L, Yang L, Dong Y, Liu T, Wei
X, Liu D, Gu H, Kong J, Yuan Z and Zhao Q: The vitamin D receptor
regulates miR-140-5p and targets the MAPK pathway in bone
development. Metabolism. 85:139–150. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Lu D, Yang C, Zhang Z, Cong Y and Xiao M:
Knockdown of Linc00515 inhibits multiple myeloma autophagy and
chemoresistance by upregulating miR-140-5p and downregulating
ATG14. Cell Physiol Biochem. 48:2517–2527. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Lu Y, Qin T, Li J, Wang L, Zhang Q, Jiang
Z and Mao J: MicroRNA-140-5p inhibits invasion and angiogenesis
through targeting VEGF-A in breast cancer. Cancer Gene Ther.
24:386–392. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Wei R, Cao G, Deng Z, Su J and Cai L:
miR-140-5p attenuates chemotherapeutic drug-induced cell death by
regulating autophagy through inositol 1,4,5-trisphosphate kinase 2
(IP3k2) in human osteosarcoma cells. Biosci Rep. 36:e003922016.
View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Nie ZY, Liu XJ, Zhan Y, Liu MH, Zhang XY,
Li ZY, Lu YQ, Luo JM and Yang L: miR-140-5p induces cell apoptosis
and decreases Warburg effect in chronic myeloid leukemia by
targeting SIX1. Biosci Rep. 39:BSR201901502019. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Zhang Y and Xu J: MiR-140-5p regulates
hypoxia-mediated human pulmonary artery smooth muscle cell
proliferation, apoptosis and differentiation by targeting Dnmt1 and
promoting SOD2 expression. Biochem Biophys Res Commun. 473:342–348.
2016. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Barquet LA: Role of VEGF in diseases of
the retina. Arch Soc Esp Oftalmol. 90 (Suppl 1):S3–S5. 2015.(In
Spanish). View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Claesson-Welsh L and Welsh M: VEGFA and
tumour angiogenesis. J Intern Med. 273:114–127. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Yang P, Xiong J, Zuo L, Liu K and Zhang H:
miR-140-5p regulates cell migration and invasion of non-small cell
lung cancer cells through targeting VEGFA. Mol Med Rep.
18:2866–2872. 2018.PubMed/NCBI
|
|
49
|
Zhang W, Zou C, Pan L, Xu Y, Qi W, Ma G,
Hou Y and Jiang P: MicroRNA-140-5p inhibits the progression of
colorectal cancer by targeting VEGFA. Cell Physiol Biochem.
37:1123–1133. 2015. View Article : Google Scholar : PubMed/NCBI
|