Introduction
Keloids are a skin fibroproliferative condition
usually accompanied by the loss of the sense of touch, pain and
itching, amongst other physical and psychosocial symptoms
(including restricted joint movement, anxiety, depression, low
self-esteem) (1). Keloids usually
occur secondary to the abnormal healing of skin wounds, and grow in
an invasive manner, extending beyond the original injured tissue
area (2). Histopathological
examinations have revealed that compared with normal skin, keloids
are characterized by the hyperproliferation of fibroblasts, an
increased proportion of myofibroblasts and the excessive deposition
of extracellular matrix (ECM) components, such as collagen I and
collagen III (3). The increased
synthesis of ECM collagen is considered to be related to the
overactivation of keloid fibroblasts. The proliferation and
migration of fibroblasts have been identified as essential for
keloid formation. To some extent, keloids exhibit invasive and
immortal growth, similar to tumor tissues (4). Based on the RNA-Seq analysis of keloid
and normal skin samples, previous studies indicated that changes in
the expression levels of pro-fibrotic genes are found to serve a
significant role in keloid formation (5,6).
Although various therapeutic modalities are used to treat keloids,
such as local surgical resection combined with radiotherapy, laser
treatments, cryotherapy and chemotherapy, the clinical effects
remain unsatisfactory (7). This is
because these methods have the potential to cause relapse, leading
to contour deformity caused by keloids repeatedly pulling on the
skin and dysfunction caused by keloids around the joint, seriously
affecting the physical and mental health, and social life of
patients (8). These issues remain a
major challenge for keloid treatment in the field of plastic
surgery. Therefore, determining the molecular mechanisms involved
in the genesis of keloids is essential for developing novel and
effective therapeutic strategies.
The emerging development of high-throughput
sequencing has enabled the identification of hub genes and
dysfunctional biological pathways involved in the pathological
process of numerous types of disease. Gene Ontology (GO) functional
term and Kyoto Encyclopedia of Genes and Genomes (KEGG) signaling
pathway enrichment analyses are common methods used in
bioinformatics analysis for annotating genes and identifying
underlying biological pathways (9).
For example, Liu et al (10)
applied weighted gene co-expression network analysis to identify
the main functions of hub genes and keloid-related pathways. The
results of this study revealed that the expression change of some
hub genes (such as DKK3 and MMP3) have been verified by
immunohistochemical staining. In addition, Fagone et al
(11) used functional term and
signaling pathway enrichment analyses to verify that macrophage
migratory inhibitory factor (MIF) receptors were variably expressed
in CD4+ T cells and central nervous lesions. Similarly,
Presti et al (12) also used
The Cancer Genome Atlas data to determine the accurate and
underlying biological mechanism in glioblastoma multiforme, and
further confirmed that MIF and its receptors were associated with
glioblastoma multiforme progression and maintenance. Runt-related
transcription factors (Runx) are a family of three genes, which
have been discovered to serve an important role in cell migration,
proliferation and apoptosis (13).
Runx2 is a member of the Runx family of transcription factors
containing the runt DNA-binding domain (14). Runx2 has been revealed to be
essential for osteoblastic differentiation and skeletal
morphogenesis, as well as tumor formation and progression (15). In addition, Runx2 contributed to the
profibrotic function and promoted the progression of lung fibrosis,
suggesting that targeting Runx2 may be of therapeutic value for
curing fibrosis-related diseases (14). Numerous specific molecular pathways
have been suggested to serve a role in liver fibrosis, mainly
related to chronic viral hepatitis B or C (HBV or HCV) (16). For example, Fagone et al
(17) reported that Runx2 was
identified as a hub gene that participated in the progression of
hepatic fibrosis, while the knockdown of Runx2 expression levels
could ameliorate liver cirrhosis. Similarly, Chen et al
(18) revealed that the protein
expression levels of Runx2 were upregulated in aortic valve
fibrosis, indicating that the AMPK-activated protein kinase α/Runx2
pathway may participate in high-fat diet-induced fibrosis in aortic
valves. A previous study also demonstrated that Runx2 was
differentially expressed in alveolar epithelial cells and
fibroblasts in pulmonary fibrosis. In fact, the cell-specific
targeting of Runx2 signaling pathways was suggested as a
therapeutic approach for idiopathic pulmonary fibrosis (14). Runx2 expression levels were also
found to be crucial in autoimmune diseases. For instance, Tchetina
et al (19) discovered that
Runx2 expression levels were upregulated in the peripheral blood
and may predict an improved response of patients with rheumatoid
arthritis to methotrexate treatment, indicating the potential of
Runx2 as a prognostic marker. However, to the best of our
knowledge, the role of Runx2 in keloid pathogenesis remains
unknown. Therefore, the present study aimed to investigate the
expression levels of Runx2 in human keloid fibroblasts (HKFs) and
to determine its role in keloid formation.
Materials and methods
Patient samples
Keloid tissues and normal skin tissues were obtained
from 9 patients (age range, 13–36, mean, 25.4 years; 5 women and 4
men) with keloids between September 2018 and May 2020 at Tongji
Hospital of Huazhong University of Science and Technology (Wuhan,
China). The patients recruited in the present study had not been
pretreated for keloid for ≥3 months and the patients keloids were
in the active stage. The diagnosis of keloid was based on clinical
appearance, symptoms, persistence for >1 year and extension
beyond the original margins. The nature of the samples was
confirmed by pathological examination. The present study was
approved by the ethics committee of Tongji Hospital of Huazhong
University of Science and Technology. Written informed consent was
obtained from all participants.
Bioinformatics analysis
Genome-wide microarray data for HKFs (from nine
different patients with keloids) and human dermal fibroblasts
(HDFs, from four different control patients) (dataset no. GSE44270)
(20) were downloaded from the Gene
Expression Omnibus (GEO) database (https://www.ncbi.nlm.nih.gov/geo). GEO2R (https://www.ncbi.nlm.nih.gov/geo/geo2r)
was used to identify the differentially expressed genes (DEGs,
|fold change| >1.5 and P<0.05) between HKFs and HDFs in the
GSE44270 dataset (21). DEGs were
then subjected to functional term enrichment analysis using Gene
Ontology (GO) analysis and signaling pathway enrichment analysis
using Kyoto Encyclopedia for Genes and Genomes (KEGG) analysis on
the Database of Annotation, Visualization and Integrated Discovery
(DAVID; http://david.abcc.ncifcrf.gov). In addition, a
protein-protein interaction (PPI) network of the DEGs was
constructed using the Search Tool for the Retrieval of Interacting
Genes/Proteins (STRING) database (http://string-db.org) and a threshold combined score
of ≥0.4 was set as the cut-off. Cytoscape software (https://cytoscape.org/index.html; version 3.6.1)
was used to visualize the PPI network. The plug-in MCODE
(https://cytoscape.org/index.html;
version 3.6.1) in Cytoscape was used to analyze the genes in the
PPI network and to identify hub genes based on the node degree
value (degree cut-off ≥2, node score cut-off ≥0.2, K-core ≥2, and
max depth = 100).
Primary cell culture and small
interfering RNA (siRNA/si) transfection
HKFs were isolated from keloid tissues (n=3) with
high Runx2 expression level and HDFs were isolated from normal skin
tissues (n=3) with low Runx2 expression level, respectively, using
the collagenase digestion method. Briefly, the adipose tissues and
epidermis were removed from the samples. Subsequently, specimens
were cut into 1×1 mm sections and digested with collagenase (0.5
mg/ml) and trypsin (0.2 mg/ml) for 6 h at 37°C. Following
centrifugation at 300 × g at 37°C. for 5 min, the supernatant was
discarded and the precipitate retained. Fibroblasts were cultured
in DMEM/F12 medium (Gibco; Thermo Fisher Scientific, Inc.)
supplemented with 10% FBS (Gibco; Thermo Fisher Scientific, Inc.),
100 IU/ml penicillin and 100 IU/ml streptomycin, and maintained at
37°C with 5% CO2. HDFs and HKFs were used in the present
study following 3–5 cell passages (22).
siRNAs targeting Runx2 (si-Runx2;
5′-CAGAAGAATGGTACAAATCCAAG-3′) and a negative control (NC) siRNA
(si-NC; 5′-TTCTCCGAACGTGTCACGTdTdT-3′) were constructed by
Guangzhou RiboBio Co., Ltd. HKFs at a density of
~2×106/well were cultured to 50% confluence in 6-well
plates and transfected with 20 µmol/ml of si-Runx2 or si-NC using
10 µl Lipofectamine® 3000 reagent (Invitrogen; Thermo
Fisher Scientific, Inc.) in Opti-MEM medium (Gibco; Thermo Fisher
Scientific, Inc.) at 37°C for 24 h, according to the manufacturer's
protocol. Following 24 h of transfection, HKFs were harvested for
further experiments.
Fluorescence and light microscopy
The expression and distribution of Runx2, collagen
I, collagen III, and fibronectin from human keloid tissues and
normal skin tissues were analyzed using fluorescence microscopy
(magnification, ×200). Meanwhile, the expression and distribution
of vimentin in HKFs and HDFs was also analyzed using fluorescence
microscopy (magnification, ×400). The morphology of fibroblasts
(included HKFs and HDFs) was observed under the light microscope
(magnification, ×40). Briefly, fresh tissues collected in the
operating rooms were immediately fixed with 4% paraformaldehyde at
37°C for 24 h and embedded in paraffin after dehydration with an
ascending alcohol series (23).
Sections were cut at 5 µm. Following paraffinization, the tissues
were rehydrated using a descending alcohol series. Sections were
washed with xylene at 37°C, deparaffinized, blocked with 5% bovine
serum albumin (Sangon Biotech Co., Ltd.) for 30 min at 37°C and
incubated overnight at 4°C with the specific primary antibodies
(anti-Runx2 1:1,000, cat. no. 55725-1-AP; anti-collagen I 1:1,000,
cat. no. 14695-1-AP; anti-collagen III 1:1,000, cat. no.
14737-1-AP; anti-fibronectin 1:1,000, cat. no. 49225-1-AP;
anti-vimentin, 1:1,000, cat. no. 82132-1-AP, all from ProteinTech
Group, Inc.). Following the primary antibody incubation, the slides
were incubated for 1 h at 37°C with the corresponding specific
secondary antibody (HRP-conjugated Affinipure Goat Anti-Rabbit
IgG(H+L), 1:2,500, cat. no. SA00001-2; HRP-conjugated Affinipure
Goat Anti-Mouse IgG(H+L), 1:2,500, cat. no. SA00001-1; both from
ProteinTech Group, Inc.), and staining with the DAB solution (Dako;
Agilent Technologies, Inc.). Nuclei were stained with DAPI for 45
min at 37°C.
Western blotting
HKFs at a density of approximately
2×106/well were cultured to 50% confluence in 6-well
plates and transfected with si-Runx2 or si-NC at 37°C for 24 h.
Total protein was extracted from cells using RIPA lysis buffer
(Boster Biological Technology). Total protein was quantified using
a BCA protein assay kit and a total of 50 µg proteins loaded per
lane were separated via 12% SDS-PAGE. The separated proteins were
subsequently transferred onto polyvinylidene fluoride membranes
(24) and blocked with 5% blocking
buffer (Invitrogen; Thermo Fisher Scientific, Inc.) in TBS-0.1%
Tween-20 (TBST) for 2 h at 37°C. The membranes were then incubated
with the following primary antibodies overnight at 4°C: Anti-Runx2
(1:1,000, cat. no. 55725-1-AP), anti-α-smooth muscle actin (SMA,
1:1,000, cat. no. 55135-1-AP), anti-collagen I (1:1,000, cat. no.
14695-1-AP), anti-collagen III (1:1,000, cat. no. 14737-1-AP),
anti-fibronectin (1:1,000, cat. no. 49225-1-AP),
anti-phosphorylated (p)-AKT (1:500, cat. no. 66444-1-Ig), anti-AKT
(1:500, cat. no. 60203-2-Ig), anti-p-PI3K (1:500, cat. no.
56321-1-Ig), anti-PI3K (1:1,000, cat. no. 48631-2-Ig) and
anti-GAPDH (1:2,500, cat. no. 63714-1-AP; all from ProteinTech
Group, Inc.). Following the primary antibody incubation, the
membranes were washed three times with TBST and incubated with the
secondary antibody [HRP-conjugated Affinipure Goat Anti-Rabbit
IgG(H+L), 1:2,500, cat. no. SA00001-2; HRP-conjugated Affinipure
Goat Anti-Mouse IgG(H+L), 1:2,500, cat. no. SA00001-1; both from
ProteinTech Group, Inc.], and staining with the DAB solution (Dako;
Agilent Technologies, Inc.) for 2 h at 37°C. Protein bands were
visualized using an ECL detection kit (Beyotime Institute of
Biotechnology) and then the signal intensity of proteins were
quantified using ImageJ software (version 1.8.0; National
Institutes of Health). All experiments were performed in
triplicate.
Reverse transcription-quantitative PCR
(RT-qPCR)
Total RNA was extracted from HKFs using
TRIzol® reagent (Invitrogen; Thermo Fisher Scientific,
Inc.). Total RNA was spectrophotometrically quantified at an
absorbance of 260 nm by a spectrophotometer (Genova; Jenway) and
reverse transcribed into cDNA using a PrimeScript RT reagent kit
(Takara Bio, Inc.) at 85°C for 5 sec, 37°C for 10 min and 4°C for
15 min. qPCR was subsequently performed (one initial cycle at 95°C
for 30 sec, followed by 40 cycles at 95°C for 5 sec and at 60°C for
30 sec.) using a Power SYBR Green PCR Master mix (Takara Bio, Inc.)
on a Real-Time Thermal cycler (Bio-Rad Laboratories, Inc.). The
following primers were used for the qPCR: Fibronectin forward,
5′-CGGTGGCTGTCAGTCAAAG-3′ and reverse, 5′-AAACCTCGGCTTCCTCCATAA-3′;
Runx2 forward, 5′-ACGAGGCAAGAGTTTCACCT-3′ and reverse,
5′-TGTCTGTGCCTTCTTGGTTC-3′; GAPDH forward,
5′-ACCACAGTCATGCCATCAC-3′ and reverse, 5′-TCCACCACCCTGTTGCTGTA-3′;
collagen I forward, 5′-GGGCAAGACAGTGATTGAATA-3′ and reverse,
5′-ACGTCFAAGCCGAATTCCT-3′; collagen III forward,
5′-AGGTCCTGCGGGTAACACT-3′ and reverse, 5′-ACTTTCACCCTTGACACCCTG-3′;
and α-SMA forward, 5′-CTGTTCCAGCCATCCTTCAT-3′ and reverse,
5′-CCGTGATCTCCTTCTGCATT-3′. The expression levels were quantified
using the 2−∆∆Cq method and normalized to GAPDH
expression levels (25).
Transwell migration assay
The migratory ability of HKFs from different groups
was evaluated using a Transwell assay. Briefly, 1×104
HKFs/well in 200 µl serum-free DMEM/F12 Glucose medium were seeded
into the upper compartments of the Transwell chambers (Corning,
Inc.). The lower compartments were filled with basic medium
supplemented with 20% FBS as a chemoattractant. The cells were
incubated at 37°C with 5% CO2 to allow the migration
through the porous membrane. Following 24 h of incubation, the
cells on the upper surface of the membrane were gently removed with
a cotton swab. The migratory cells in the lower chamber were fixed
with 4% paraformaldehyde for 30 min, stained with 0.1% crystal
violet for 10 min at 37°C and gently rinsed 3 times with PBS. The
stained cells were counted in 3 randomly selected fields under a
light microscope (magnification, ×200) (26).
Wound healing assay
A wound healing assay was used to analyze the
migratory ability of HKFs. Briefly, the fibroblasts were seeded
into 6-well culture plates at a density of 1×106
cells/well and transfected with si-Runx2 or si-NC for 24 h at 37°C.
A scratch was created using a sterile 200-µl pipette tip in the
cell monolayer upon the fibroblasts reaching 95–100% confluence.
The wells were rinsed with PBS and incubated in serum-free DMEM/F12
Glucose medium at 37°C with 5% CO2. The migration of the
cells in the different groups into the wound area was observed
under a light microscope (magnification, ×100) at 0, 12 and 24 h.
ImageJ software (version 1.8.0; National Institutes of Health) was
used for analysis.
Proliferation assay
The proliferative ability of cells was determined
using a Cell Counting Kit-8 (CCK-8) assay (Dojindo Molecular
Technologies, Inc.). Following 24 h of transfection at 37°C, HKFs
were seeded into 96-well culture plates at a density of
5×104 cells/well in 100 µl DMEM/F12 Glucose medium
supplemented with 10% FBS and cultured for 24 h at 37°C. Following
the incubation, 10 µl CCK-8 reagent was added to each well and
incubated for 1 h at 37°C. The absorbance at 450 nm was measured
using a microplate reader.
Flow cytometric analysis of early and
late apoptosis
The apoptotic rate of HKFs was analyzed using an
Annexin V-FITC/PI apoptosis detection kit (BD Biosciences).
Briefly, HKFs were seeded into 6-well plates at a density of
1×106 cells/well and incubated for 12 h at 37°C. Cells
were transfected with si-Runx2 or si-NC for 24 h at 37°C,
subsequently harvested (300 × g; 5 min at 37°C), washed with PBS
and resuspended with 200 µl the Annexin V-PI binding buffer at a
final density of 2×106 cells/flow tube. The cell
suspension was mixed with 5 µl Annexin V-FITC and 5 µl propidium
iodide (PI) solution and incubated in the dark for 15 min at 4°C.
The apoptotic cells from each sample were detected using a
fluorescence-activated cell sorting flow cytometer (BD Biosciences)
and analyzed using FlowJo software (version 10.2; FlowJo LLC). The
total number of cells was assessed using quadrant statistical
method (upper right quadrant-advanced stage apoptosis cell
percentage; lower right quadrant-prophase apoptosis cell
percentage) (27).
Statistical analysis
Statistical analysis was performed using GraphPad
Prism version 8.0.1 software (GraphPad Software, Inc.) and all data
are presented as the mean ± SD. Statistical differences between two
groups were analyzed using an unpaired two-tailed Student's t-test,
whereas differences between ≥3 groups were analyzed using a one-way
ANOVA followed by a Tukey's post hoc test for multiple comparisons.
The primary fibroblasts (including HKFs and HDFs) used in the
experiment were derived from 3 different individuals. P<0.05 was
considered to indicate a statistically significant difference.
Results
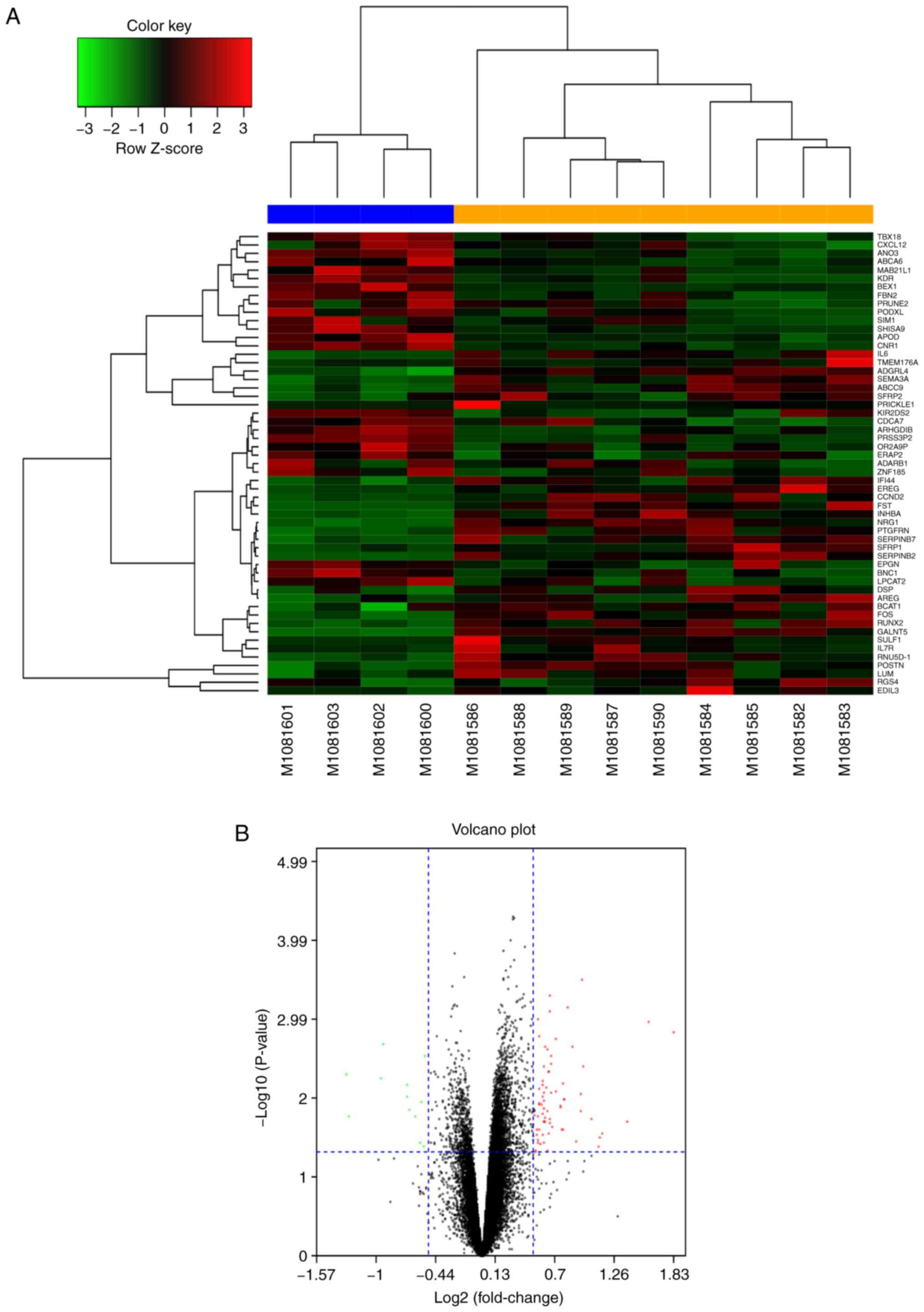
Identification of differentially
expressed mRNAs in keloid fibroblasts
The microarray dataset GSE44270 was downloaded from
the GEO database. This dataset comprised nine samples of HKFs
(GSM1081582-GSM1081590) and four samples of HDFs
(GSM1081600-GSM1081603). GEO2R was used to identify the DEGs in the
GSE44270 dataset. The 60 most DEGs (30 upregulated, including
Runx2, FOS, AREG and BCAT1 and 30 downregulated, including TBX18,
ANO3, SIM1 and APOD) were represented in hierarchical clustering
maps (Fig. 1A) and volcano maps
(Fig. 1B). A PPI network was
generated using the STRING online tool and the result revealed that
MMP3, VEGF, Runx2 and CTGF were the most prominent proteins and had
the most prominent interactions in the PPI network (Fig. 2A). The hub genes (network-centric
genes, including Runx2) from the PPI network were identified and
visualized using the plug-in MCODE in Cytoscape (Fig. 2B). These results indicated that
Runx2 may serve an important regulatory role in the pathological
process of keloids.
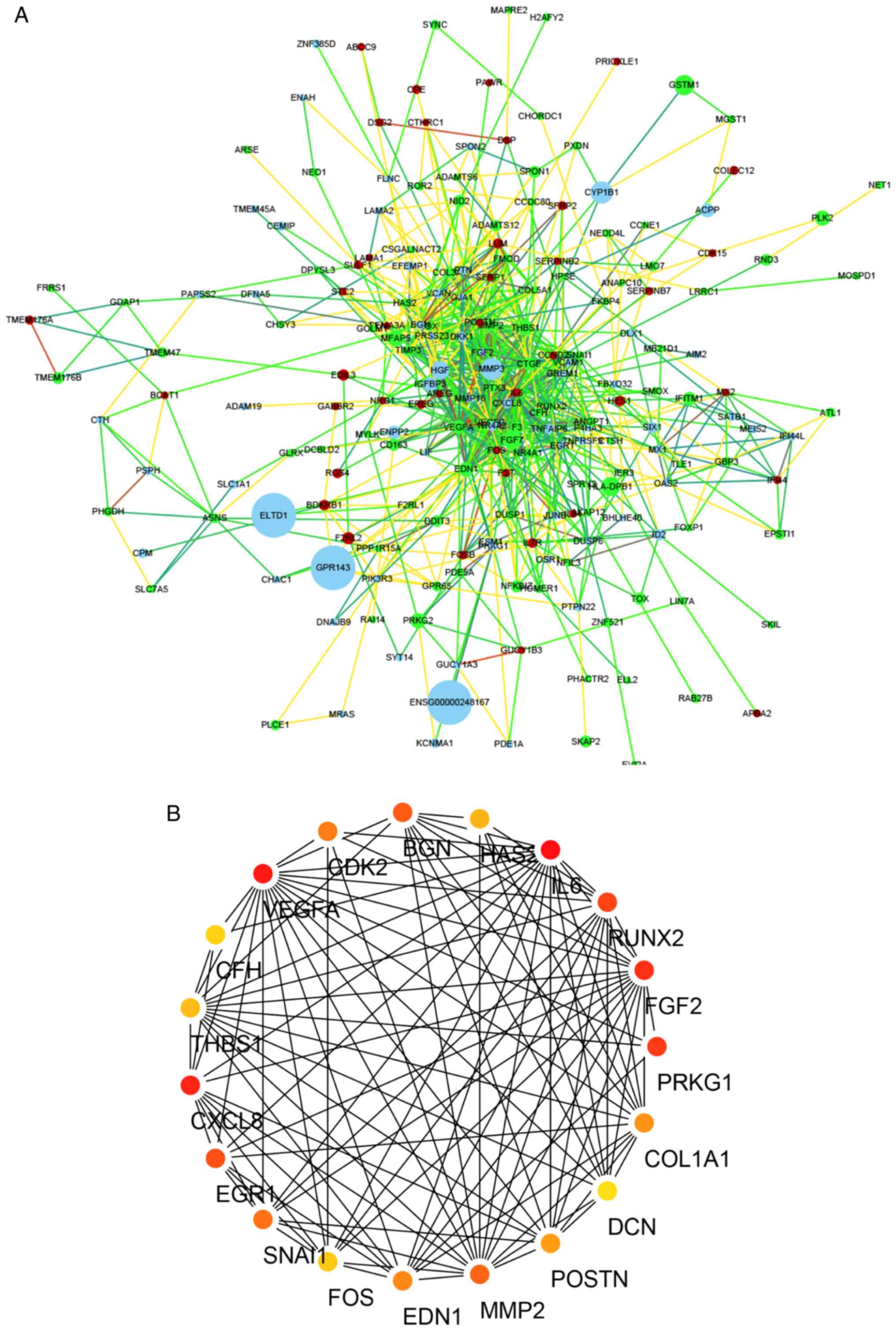 | Figure 2.DEGs PPI network complex. (A) PPI
network of DEGs was generated using the Search Tool for the
Retrieval of Interacting Genes/Proteins database. The gradual node
color represented the fold change (HKFs vs. HDFs) of DEGs in
GSE44270 dataset. The redder the color of the node, the greater the
fold change of the gene. Conversely, the greener the color of the
node, the smaller the fold change of the gene. The gradient color
line represented the degree of connection between each node (the
green line represented the closest connection, the red line
represented the most distant connection) The gradual node volume
represented the P-value of each node, and the larger the P-value,
the larger the volume of the node. (B) The hub genes
(network-centric genes; including PRKG1, FGF2, Runx2, IL6, HAS2,
BGN, CDK2, VEGFA, CFH, THBS1, CXCL8, EGR1, SNAI1, FOS, EDN1, MMP2,
POSTN, DCN, and COL1A1) from the PPI network were identified and
visualized using the plug-in MCODE in Cytoscape. The different
colors represented different degree value and the redder the color,
the greater the degree value of the node. DEGs, differentially
expressed genes; PPI, protein-protein interaction; HKFs, primary
human keloid fibroblasts; HDFs, human dermal fibroblasts; PRKG1,
protein kinase cGMP-dependent 1; FGF2, fibroblast growth factor 2;
Runx2, RUNX family transcription factor 2; IL6, interleukin 6;
HAS2, hyaluronan synthase 2; BGN, biglycan; CDK2, cyclin dependent
kinase 2; VEGFA, vascular endothelial growth factor A; CFH,
complement factor H; THBS1, thrombospondin 1; CXCL8, C-X-C motif
chemokine ligand 8; EGR1, early growth response 1; SNAI1, snail
family transcriptional repressor 1; FOS, Fos proto-oncogene, AP-1
transcription factor subuni; EDN1, endothelin 1; MMP2, matrix
metallopeptidase 2; POSTN, periostin; DCN, decorin; COL1A1,
collagen type I α 1 chain. |
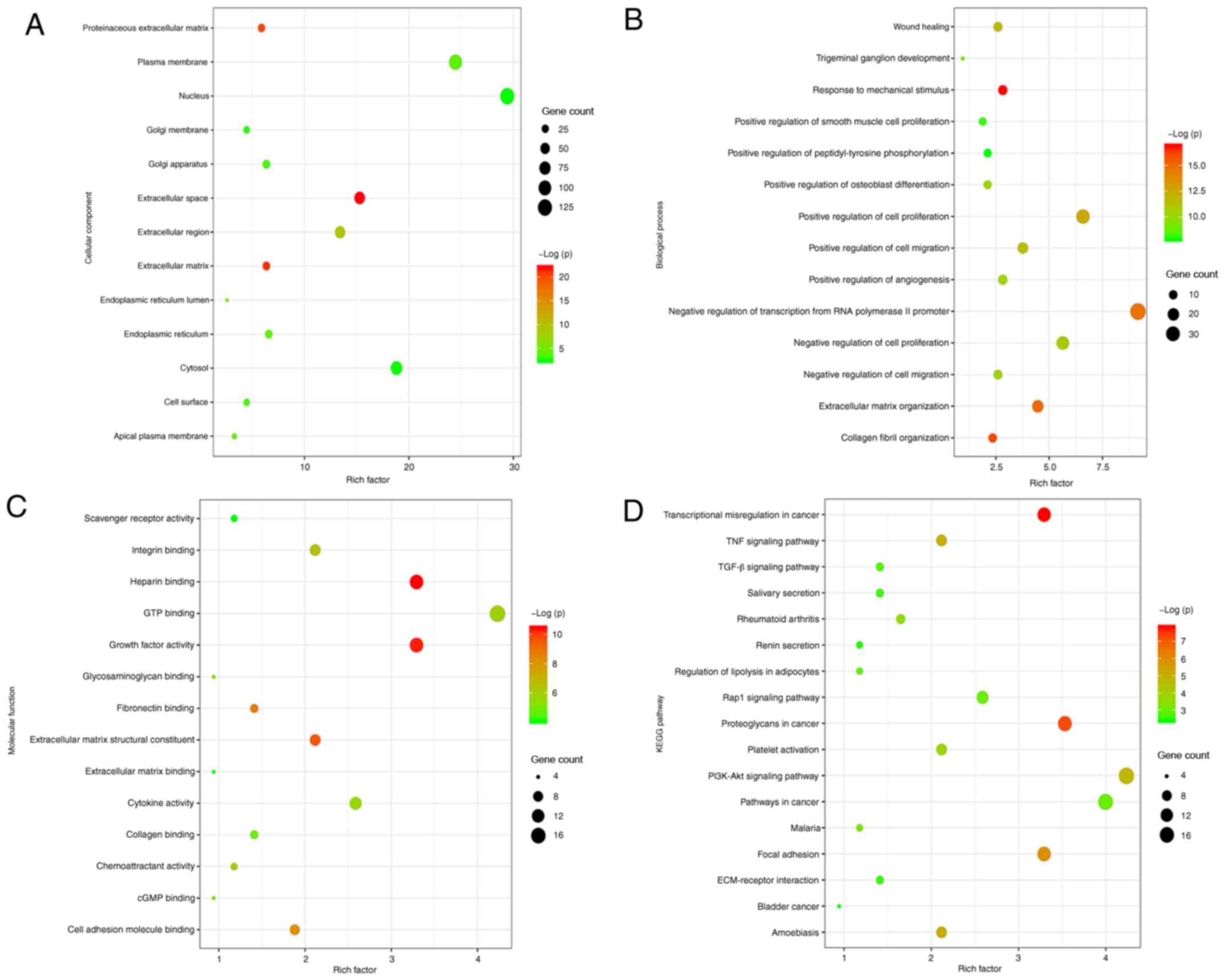
Functional term and signaling pathway
enrichment analyses
GO consists of three structured ontologies that
describe gene function terms based on their associated biological
processes (BP), cellular components (CC) and molecular functions
(MF) (28). Meanwhile, the KEGG
database is a database applied to search genomes, promising drug
candidates and biological pathways (29). GO functional term and KEGG signaling
pathway enrichment analyses are widely applied to investigate
pathogenetic pathways and novel therapeutic approaches for the
treatment of several pathological conditions, including autoimmune,
neoplastic, psychiatric and neurodegenerative disorders (11,30).
DAVID was used to analyze the BPs, CCs and MFs, and the associated
signaling pathways of the DEGs (Fig.
3). A total of 427 upregulated genes associated with keloids
were screened for further functional term and signaling pathway
enrichment analyses. CC analyses indicated that the DEGs closely
associated with the pathological progression of keloids were
strongly enriched in the ‘Extracellular matrix’, ‘Extracellular
space’ and ‘Extracellular region’ (Fig.
3A). BP analyses identified that the genes closely associated
with the pathological progression of keloids were mainly enriched
in ‘Extracellular matrix organization’, ‘Positive regulation of
cell proliferation’ and ‘Positive regulation of cell migration’
(Fig. 3B). MF analyses revealed
that the DEGs closely associated with the pathological progression
of keloids were enriched in ‘Extracellular matrix structural
constituents’, ‘Fibronectin binding’, ‘Growth factor activity’ and
‘Collagen binding’ (Fig. 3C). KEGG
analyses revealed that the genes closely associated with the
pathological progression of keloids were mainly enriched in the
‘PI3K-Akt signaling pathway’, ‘TGF-β signaling pathway’,
‘ECM-receptor interaction’ and ‘Focal adhesion’ (Fig. 3D). Overall, GO and KEGG analysis
revealed that most of the DEGs closely associated with the
pathological progression of keloids (including Runx2) were
significantly enriched in the BP ‘Positive regulation of cell
proliferation’, in the CC ‘Extracellular matrix’, in the MF
‘Extracellular matrix structural constituents’ and in the KEGG
‘PI3K-Akt signaling pathway’.
Expression levels of Runx2 in keloid
tissues and HKFs
The expression levels of Runx2 in keloid tissues and
HKFs compared with normal skin tissues and HDFs were analyzed. The
protein expression levels of Runx2 were determined by western
blotting and the results indicated that Runx2 expression levels
were significantly upregulated in human keloid tissues and HFKs
compared with normal skin tissues and HDFs, respectively (Figs. 4A and B and S1). Consistent results were obtained in
the mRNA expression levels using RT-qPCR (Fig. 4C). Immunofluorescence subsequently
revealed that the fluorescence intensities of Runx2, collagen I,
collagen III and fibronectin in keloid tissues were markedly higher
compared with the normal skin tissues (Fig. 4D). Primary fibroblasts (including
HKFs and HDFs) appeared as long fusiform cells under the light
microscope (magnification, ×40) and immunofluorescence imaging of
vimentin expression was performed using fluorescence microscope
(magnification, ×400; Fig. 4E). The
results of the immunofluorescence experiments revealed that
mesenchymal-derived skin fibroblasts (including HKFs and HDFs)
could express vimentin. To investigate the biological function of
Runx2 in HKFs, Runx2 was knocked down by transfection with
si-Runx2. The result of western blotting demonstrated that Runx2
expression was markedly decreased after si-Runx2 transfection
compared to the si-NC transfected HKFs (Fig. 4F). Compared with si-1 and si-3, si-2
had the most significant silencing efficiency and then was selected
for subsequent experiments.
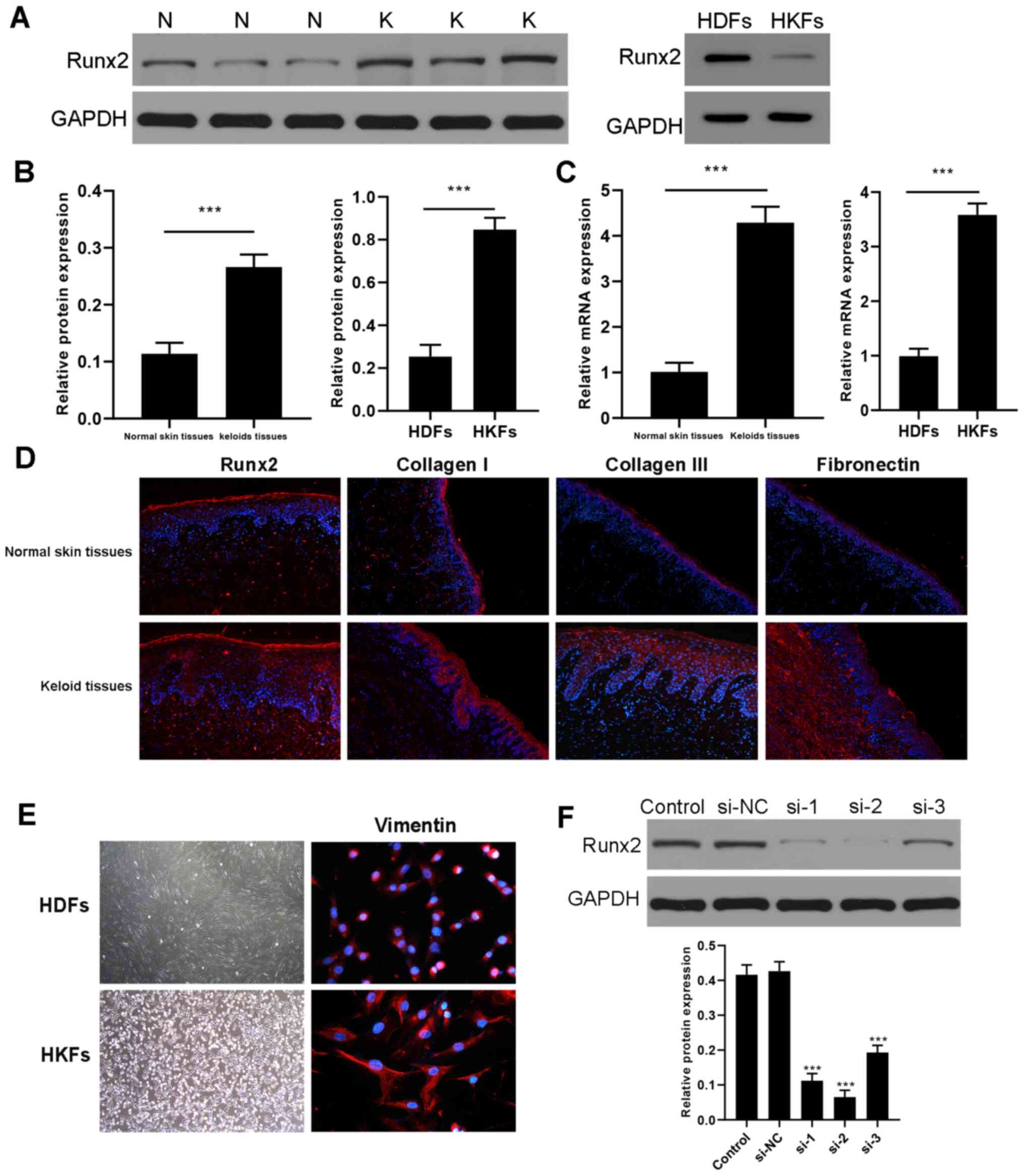 | Figure 4.Upregulation of Runx2 expression
levels in keloid tissues and HKFs. (A) Expression levels of Runx2
were analyzed in keloid and normal tissue and HDFs and HKFs using
western blotting. (B) Semi-quantification of the expression levels
from part (A). ***P<0.001 keloid tissues vs. normal skin tissues
(n=3); ***P<0.001 HKFs vs. HDFs (n=3). (C) mRNA expression
levels of Runx2 were analyzed using reverse
transcription-quantitative PCR. ***P<0.001 keloid tissues vs.
normal skin tissues (n=3); ***P<0.001 HKFs vs. HDFs (n=3). (D)
Immunofluorescence was used to determine that the expression of
Runx2, collagen I, collagen III and fibronectin in the keloid
tissues was upregulated compared with normal skin tissues
(magnification, ×200). (E) Left panel: Morphology of fibroblasts
under the light microscope (magnification, ×40). Primary
fibroblasts (including HKFs and HDFs) appeared as long fusiform
cells. Right panel: immunofluorescence imaging of vimentin
expression (red field, magnification, ×400). (F) Western blotting
revealed a significantly downregulation in the expression levels of
Runx2 in HKFs transfected with si-Runx2 (si-1, si-2 and si-3)
compared with si-NC. The silencing effect of si-2 was much stronger
than those of si-1 and si-3 in HKFs. ***P<0.001 vs. si-NC (N=3).
Data are presented as the mean ± SD of 3 independent experiments.
N, normal skin tissue; K, keloid tissue; HKFs, human keloid
fibroblasts; HDFs, human dermal fibroblasts; si, small interfering
RNA; NC, negative control; Runx2, Runt-related transcription factor
2. |
Regulation of proliferation, migration
and apoptosis of HKFs following Runx2 knockdown
The effect of Runx2 on HKF proliferation was
analyzed using CCK-8 assays. As a consequence of Runx2 knockdown by
the transfection with si-Runx2, HKF proliferation was significantly
suppressed compared with the si-NC group (Fig. 5A). Using Annexin V//PI staining, the
percentage of apoptotic cells was markedly increased after Runx2
expression was knocked down (Fig.
5B). The population of migratory HKFs was significantly
decreased following the knockdown of Runx2 compared with the si-NC
group (Fig. 5C). A wound healing
assay demonstrated that the cell number in the wound of the
si-Runx2 group was significantly decreased compared with the si-NC
group at 12 and 24 h (Fig. 5D).
Taken together, these results suggested that the knockdown of Runx2
in HKFs may inhibit cell proliferation and migration.
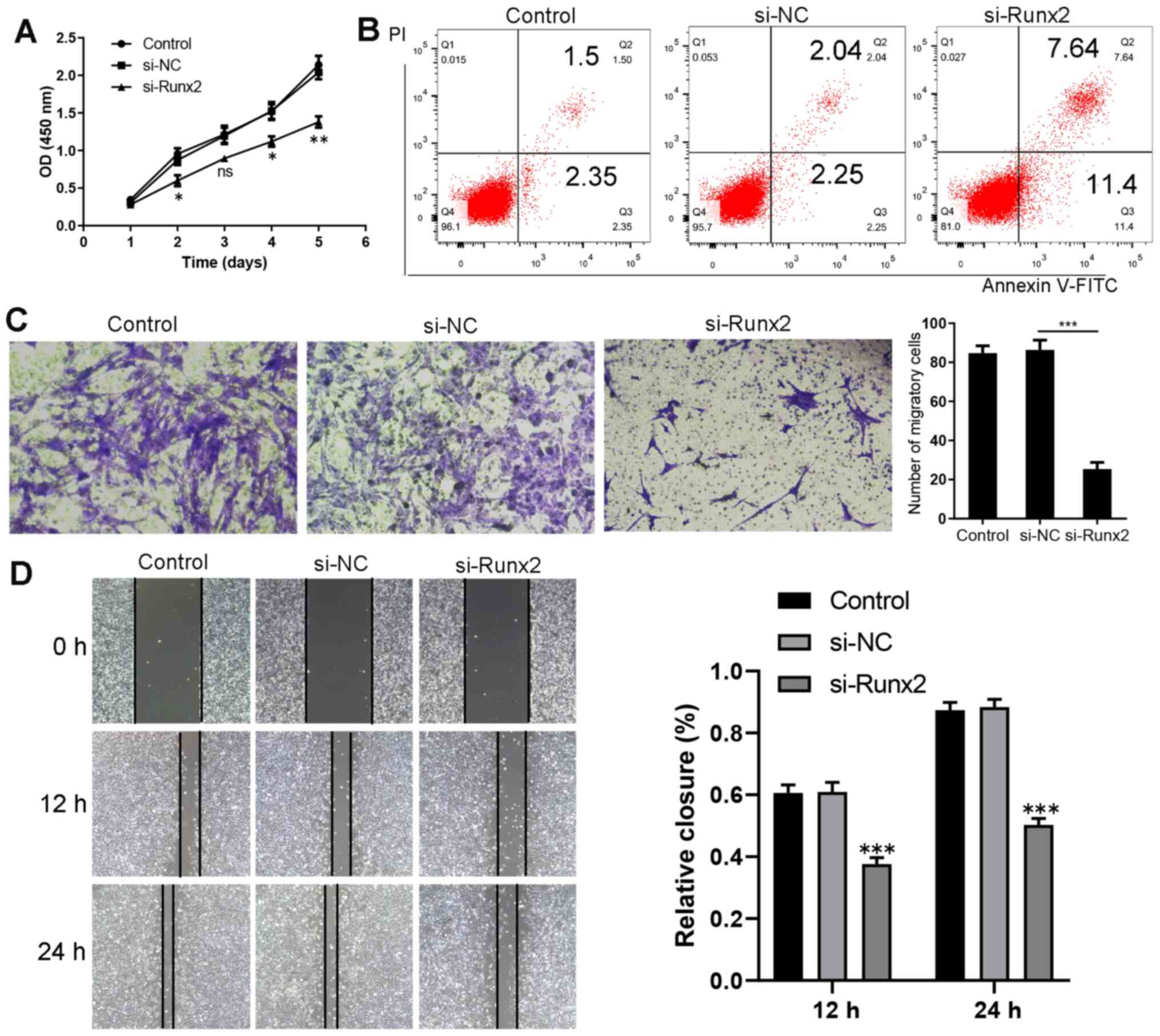 | Figure 5.Runx2 knockdown inhibits the
migration and proliferation, and promotes the apoptosis of HKFs.
(A) Cell Counting Kit-8 assay revealed that the proliferation of
HKFs was inhibited following Runx2 knockdown. *P<0.05,
**P<0.01 vs. si-NC (n=3). (B) Promotion of apoptosis following
the transfection of HKFs with si-Runx2, as shown by flow cytometry
using an Annexin V-FITC/PI assay. The lower left quadrant (Annexin
V-/PI-) represents live cells, the lower right quadrant (Annexin
V+/PI-) represents early apoptotic stage cells, the upper right
quadrant (Annexin V+/PI+) represents late apoptotic stage or
necrotic cells. (C) ***P<0.001 si-Runx2 vs. si-NC (n=3). (D)
Evaluation of cell migration using a wound healing assay
(magnification, ×100). 12 h ***P<0.001 si-Runx2 vs. si-NC (n=3);
24 h ***P<0.001 si-Runx2 vs. si-NC (n=3). Data are presented as
the mean ± SD of three independent experiments. HKFs, human keloid
fibroblasts; si, small interfering RNA; NC, negative control;
Runx2, Runt-related transcription factor 2; PI, propidium iodide;
OD, optical density. |
Downregulation of ECM protein
expression levels due to Runx2 knockdown in HKFs
Keloids are characterized by the presence of large
numbers of myofibroblasts and the excessive deposition of ECM
components, such as collagen I and collagen III (31). The transfection of HKFs with
si-Runx2 resulted in the significant downregulation of α-SMA,
collagen I, collagen III and fibronectin expression levels at both
the mRNA and protein level compared with the si-NC group (Fig. 6A and B).
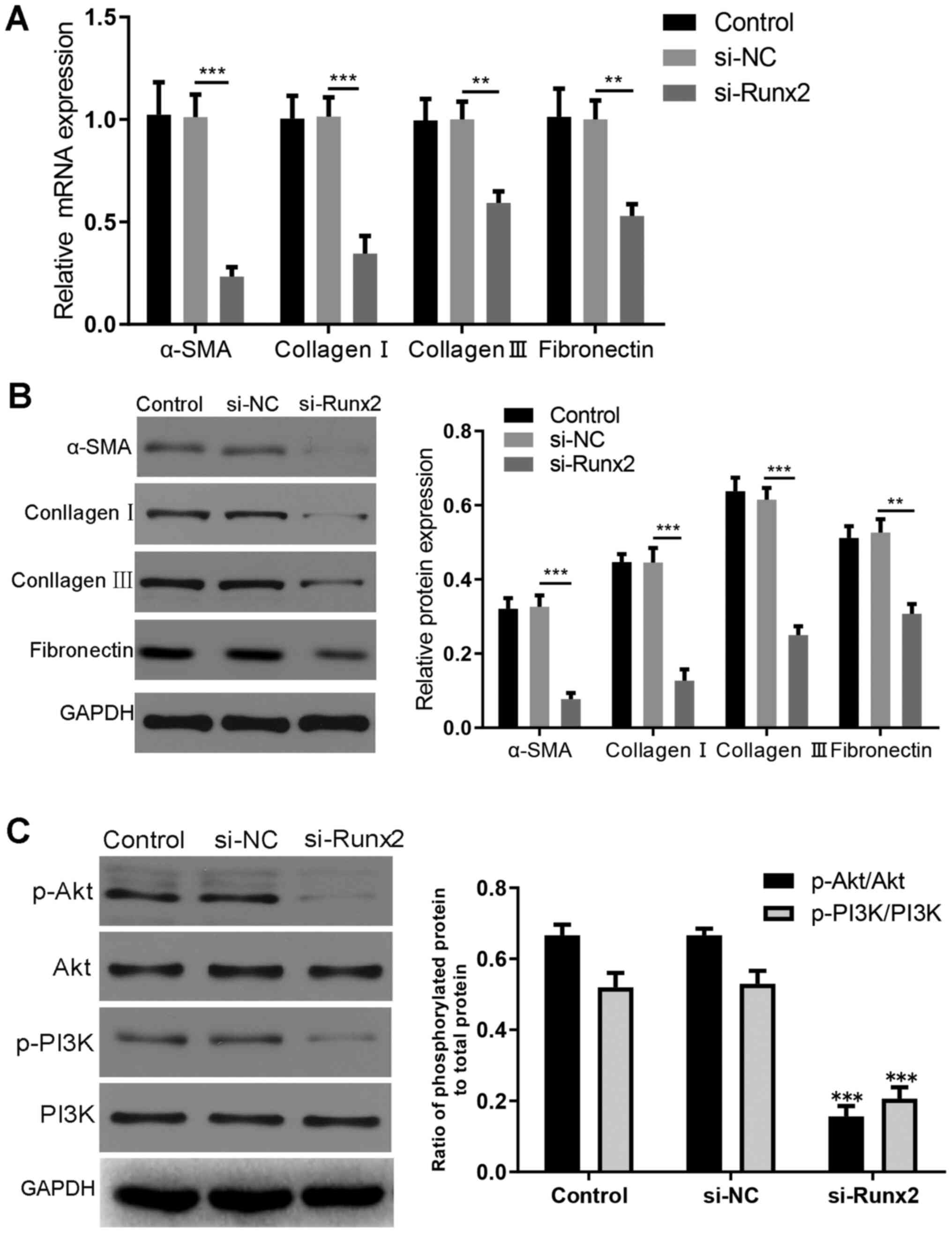 | Figure 6.Regulation of ECM deposition through
the PI3K/AKT signaling pathway. (A) Reverse
transcription-quantitative PCR was used to demonstrate that the
expression levels of α-SMA, collagen I, collagen III and
fibronectin were downregulated in HKFs following transfection.
α-SMA, ***P<0.001 si-Runx2 vs. si-NC (n=3); collagen I,
***P<0.001si-Runx2 vs. si-NC (n=3); collagen III, **P<0.01
si-Runx2 vs. si-NC (n=3); fibronectin, **P<0.01 si-Runx2 vs.
si-NC (n=3). (B) Western blotting was used to demonstrate that the
expression levels of α-SMA, collagen I, collagen III and
fibronectin were downregulated in HKFs following transfection.
α-SMA, ***P<0.001 si-Runx2 vs. si-NC (n=3); collagen I,
***P<0.001 si-Runx2 vs. si-NC (n=3); collagen III, ***P<0.001
si-Runx2 vs. si-NC (n=3); fibronectin, **P<0.01 vs. si-NC (n=3).
(C) Expression levels of AKT, p-AKT, PI3K and p-PI3K were analyzed
using western blotting. Statistical analysis was based on the ratio
between the levels of phosphorylated protein and total protein.
Following normalization to GAPDH, no difference was observed in the
PI3K and AKT total protein expression levels in the si-Runx2 group
compared with the si-NC group. ***P<0.001, p-AKT vs. AKT
(n=3);***P<0.001, p-PI3K vs. PI3K (n=3). The relative protein
expression levels were semi-quantified using ImageJ software. Data
are presented as the mean ± SD of 3 independent experiments. α-SMA,
α-smooth muscle actin; si, small interfering RNA; NC, negative
control; Runx2, Runt-related transcription factor 2; p-,
phosphorylated. |
Knockdown of Runx2 suppresses the
phosphorylation of AKT and PI3K in HKFs
KEGG signaling pathway enrichment analyses revealed
that the genes that served an important role in the pathology of
keloids were mainly enriched in the PI3K/AKT signaling pathway
(Fig. 3D). Therefore, to
investigate the effect of Runx2 knockdown on the PI3K/AKT signaling
pathway, the expression levels of AKT, p-AKT, PI3K and p-PI3K were
analyzed using western blotting. The results revealed that the
expression levels of p-AKT and p-PI3K were significantly
downregulated in HKFs following Runx2 knockdown compared with the
si-NC group (Fig. 6C).
Discussion
Keloids are a type of benign mass that occur in
different shapes, which are characterized by a reddish protuberance
of the skin and a tough texture (32). Patients with keloids often
experience pain and itching as the main symptoms and often suffer
from infections due to scratching or physical friction (33). Although keloids are benign masses
and do not metastasize, they sometimes seriously impact the
confidence of an individual with their appearance, which
subsequently affects the life quality (8). However, there is currently no
effective treatment for improving keloids prognosis and reducing
recurrence (34). At present, it is
known that genetics, gene mutation, inflammatory cytokines and
autoimmunity are associated with keloid formation, but the specific
pathogenesis remains unclear (35).
Therefore, further investigations to determine the process of
keloid formation using HKFs and to identify effective treatments
have become necessary for plastic surgery clinicians and
researchers. HKFs are the main effector cells involved in the
development of keloids. The proliferation and migration of HKFs was
discovered to be closely related to the formation of keloids
(36). Compared with normal skin
tissues, HKFs were reported to secrete more ECM components, such as
collagen I, collagen III and mucin. Simultaneously, HKFs can also
secrete related cytokines, including TGF-β and platelet-derived
growth factor to promote keloid hyperplasia (37). Therefore, an in-depth understanding
of the molecular mechanisms that regulate HKF function may provide
novel therapeutic strategies for keloids.
In the present study, GO and KEGG pathway analysis
were applied to investigate the possible biological functions and
potential mechanisms of DEGs in keloid formation. GO analysis
revealed that most of the DEGs (including Runx2) were significantly
enriched in the BP ‘Positive regulation of cell proliferation’, in
the CC ‘Extracellular matrix’, in the MF ‘Extracellular matrix
structural constituents’. Previous studies have confirmed that the
proliferation of fibroblasts and the excessive deposition of ECM
served an indispensable role in the pathological process of keloids
(3,31,36),
which is consistent with the GO enrichment analysis. KEGG analysis
of these DEGs (including Runx2) proposed that the ‘PI3K-Akt
signaling pathway’, ‘TGF-β signaling pathway’, ‘ECM-receptor
interaction’ and ‘Focal adhesion’ might serve a pathological role
in keloid formation. The abovementioned findings revealed that the
identified hub genes and pathways using GO and KEGG analysis could
significantly enrich our understanding of the development of
keloids and provide the potential mechanisms in keloid
formation.
Runx2 has been discovered to be closely associated
with the differentiation of mesenchymal cells to osteoblast lineage
cells and is essential for cell proliferation and ECM formation
(38). Mümmler et al
(14) reported that Runx2
expression levels were upregulated in ATII cells isolated from
fibrotic mouse lungs, and Runx2 knockdown using siRNA decreased the
migratory ability of A549 cells. Herreño et al (15) demonstrated that Runx2 promoted
epithelial-mesenchymal transition and increased the migratory
capacity in lung adenocarcinoma cells. In the present study, the
results revealed that the expression levels of Runx2 in HKFs and
keloid tissues were significantly upregulated compared with HDFs
and normal skin tissues. Runx2 knockdown with siRNA inhibited HKF
proliferation, migration, and the deposition of the ECM (including
collagen I, collagen III, and fibronectin). Furthermore, the
apoptotic cell population in HKFs was markedly increased following
Runx2 knockdown. Consequently, this may cause HKFs to lose their
highly proliferative and migratory nature, and the excessive
deposition of the ECM, which may thereby result in the delayed
progression of keloids, indicating the possibility of Runx2 as a
potential target for the treatment of keloids.
Previously, it was demonstrated that the PI3K/AKT
signaling pathway was involved in the regulation of cell
proliferation, differentiation, apoptosis and glucose transport
(39). In the current study, KEGG
signaling pathway enrichment analysis revealed that the DEGs in
keloids were mainly enriched in the PI3K/AKT signaling pathway. It
was also discovered that the phosphorylation levels of PI3K and AKT
significantly decreased following the transfection with si-Runx2,
which indicated that the PI3K/AKT signaling pathway was involved in
HFK functions.
The protein mTOR consists of the serine/threonine
kinase TOR and at ≥5 other proteins. mTOR participates in several
signaling transduction networks, especially the PI3K/AKT signaling
pathway, to regulate cell cycle progression, cell survival,
proliferation, invasion and apoptosis (40). Research has revealed that mTOR is a
key component of the PI3K/AKT/mTOR signaling pathway, and it is
considered as an attractive therapeutic target in breast cancer
(41). In addition, mTOR was
reportedly involved in regulating the pathogenesis of autoimmune
diseases and viral infections, such as human immunodeficiency virus
(HIV) and SARS-CoV-2 (42). By
analyzing the clinical data and measuring the plasma levels of
p-PI3K, p-AKT and p-mTOR using ELISAs, Ge et al (43) found that the PI3K/AKT/mTOR signaling
pathway was closely associated with the pathogenesis of lupus
nephritis. In addition, to determine possible novel therapeutic
targets, Fagone et al (44)
applied the anti-signature perturbation analysis to predict mTOR as
a potential target to delay the progression of Covid-19 through
analyzing transcriptomic profiles of primary human lung epithelium
following SARS-CoV-2 infection. Nicoletti et al (45) also identified a relationship between
dysregulated mTOR activation and the pathogenesis of HIV, which
suggested that mTOR might be a multifunctional therapeutic target
in HIV infection. Interestingly, Tu et al (46) discovered that CUDC-907, which is a
PI3K/Akt/mTOR pathway inhibitor, could significantly inhibit the
proliferation, migration, invasion and ECM deposition of HKFs.
Therefore, based on these aforementioned findings, it was
hypothesized that inhibiting the PI3K/AKT/mTOR signaling pathway
may reverse the pathological phenotype of HKFs.
Gene therapy is a recently developed and novel
approach for treatment, which has demonstrated tremendous potential
(47). For example, the feasibility
of gene therapy applications of a retroviral vector expressing
LAMB3 cDNA has been definitely confirmed by the successful results
obtained in a clinical trial for the LAMB3-deficient form of
junctional epidermolysis bullosa (48). RNA interference is a universal
mechanism of post-transcriptional gene silencing induced by siRNAs,
which are double-stranded non-coding RNAs of 21–22 nucleotides in
length generated from longer double strand RNA by the action of
Dicer (49). siRNA is able to
prevent target gene expression by inducing the degradation of
specific mRNA (50). In the present
study, Runx2 siRNA was designed to knockdown Runx2 mRNA and
transfected into HKFs. The results revealed that the biological
functions (including proliferation, migration and ECM deposition)
of HKFs were significantly inhibited by si-Runx2 transfection,
suggesting that siRNA treatment could be used for the treatment of
keloids.
Nonetheless, there are several limitations to the
current study. Firstly, mTOR is a key component of the
PI3K/AKT/mTOR signaling pathway and is considered as an attractive
therapeutic target in the pathological process of keloids (46). However, the present study only
investigated the PI3K and AKT phosphorylation levels and did not
further confirm the potential mechanism of mTOR in regulating the
progression of keloids. In addition, relatively few patients with
keloids need to undergo surgical resection. Therefore, it is
difficult to collect a large number of samples, which is a general
limitation of keloid research. For this reason, the sample number
(9 pairs) in the present study is small, and further large-scale
analysis will be required in the future.
In conclusion, the results of the current study
indicated that Runx2 expression levels may be upregulated in keloid
tissues. Silencing Runx2 in HKFs inhibited the cell proliferation,
migration, expression levels of ECM-related proteins through the
suppression of the PI3K/AKT signaling pathway. Meanwhile, we found
that silencing Runx2 inhibited the expression of α-SMA in HKFs and
α-SMA was a marker of myofibroblasts, indicating that silencing
Runx2 inhibited the differentiation into myofibroblasts. Therefore,
Runx2 may be a potential therapeutic target for keloids and could
be applied in the clinic as a novel therapeutic strategy.
Supplementary Material
Supporting Data
Acknowledgements
Not applicable.
Funding
No funding was received.
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
WL, MW and YR designed and performed the
experiments, and contributed to the writing of the manuscript. QZ
and YW made substantial contributions to the conception and design
of the present study, analysis and interpretation of data. XL and
WH were involved in revising the manuscript critically for
important intellectual content and the acquisition of data. All
authors read and approved the final manuscript.
Ethics approval and consent to
participate
The present study was approved by the ethics
committee of Tongji Hospital of Huazhong University of Science and
Technology. Written, informed consent was obtained from all
participants. The collection and use of samples were in accordance
with the Declaration of Helsinki principles.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Ud-Din S and Bayat A: New insights on
keloids, hypertrophic scars, and striae. Dermatol Clin. 32:193–209.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
de Oliveira GV and Gold MH: Hydrocolloid
dressings can be used to treat hypertrophic scars: An outpatient
dermatology service devoted to treat keloids and challenging scars.
J Cosmet Dermatol. Oct 26–2020.(Epub ahead of print). doi:
10.1111/jocd.13792 2020.
|
|
3
|
Li Y, Liu H, Liang Y, Peng P, Ma X and
Zhang X: DKK3 regulates cell proliferation, apoptosis and collagen
synthesis in keloid fibroblasts via TGF-β1/Smad signaling pathway.
Biomed Pharmacother. 91:174–180. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Lee JY, Yang CC, Chao SC and Wong TW:
Histopathological differential diagnosis of keloid and hypertrophic
scar. Am J Dermatopathol. 26:379–384. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Lee YS, Liang YC, Wu P, Kulber DA, Tanabe
K, Chuong CM, Widelitz R and Tuan TL: STAT3 signalling pathway is
implicated in keloid pathogenesis by preliminary transcriptome and
open chromatin analyses. Exp Dermatol. 28:480–484. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Zhang Q, Cai L, Wang M, Ke X, Zhao X and
Huang Y: Identification of a novel mutation in the
mechanoreceptor-encoding gene CXCR1 in patients with keloid. Arch
Dermatol Res. 310:561–566. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Liu J, Zhu H, Wang H, Li J, Han F, Liu Y,
Zhang W, He T, Li N, Zheng Z, et al: Methylation of secreted
frizzled-related protein 1 (SFRP1) promoter downregulates
Wnt/β-catenin activity in keloids. J Mol Histol. 49:185–193. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Bijlard E, Kouwenberg CA, Timman R, Hovius
SE, Busschbach JJ and Mureau MA: Burden of Keloid Disease: A
Cross-sectional Health-related Quality of Life Assessment. Acta
Derm Venereol. 97:225–229. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Chen L, Zhang YH, Wang S, Zhang Y, Huang T
and Cai YD: Prediction and analysis of essential genes using the
enrichments of gene ontology and KEGG pathways. PLoS One.
12:e01841292017. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Liu W, Huang X, Liang X, Zhou Y, Li H, Yu
Q and Li Q: Identification of Key Modules and Hub Genes of Keloids
with Weighted Gene Coexpression Network Analysis. Plast Reconstr
Surg. 139:376–390. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Fagone P, Mazzon E, Cavalli E, Bramanti A,
Petralia MC, Mangano K, Al-Abed Y, Bramati P and Nicoletti F:
Contribution of the macrophage migration inhibitory factor
superfamily of cytokines in the pathogenesis of preclinical and
human multiple sclerosis: In silico and in vivo evidences. J
Neuroimmunol. 322:46–56. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Presti M, Mazzon E, Basile MS, Petralia
MC, Bramanti A, Colletti G, Bramanti P, Nicoletti F and Fagone P:
Overexpression of macrophage migration inhibitory factor and
functionally-related genes, D-DT, CD74, CD44, CXCR2 and CXCR4, in
glioblastoma. Oncol Lett. 16:2881–2886. 2018.PubMed/NCBI
|
|
13
|
Mevel R, Draper JE, Lie ALM, Kouskoff V
and Lacaud G: RUNX transcription factors: orchestrators of
development. Development. 146:dev1482962019. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Mümmler C, Burgy O, Hermann S, Mutze K,
Günther A and Königshoff M: Cell-specific expression of
runt-related transcription factor 2 contributes to pulmonary
fibrosis. FASEB J. 32:703–716. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Herreño AM, Ramírez AC, Chaparro VP,
Fernandez MJ, Cañas A, Morantes CF, Moreno OM, Brugés RE, Mejía JA,
Bustos FJ, et al: Role of RUNX2 transcription factor in epithelial
mesenchymal transition in non-small cell lung cancer lung cancer:
Epigenetic control of the RUNX2 P1 promoter. Tumour Biol.
41:1010428319851014. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Fagone P, Mangano K, Pesce A, Portale TR,
Puleo S and Nicoletti F: Emerging therapeutic targets for the
treatment of hepatic fibrosis. Drug Discov Today. 21:369–375. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Fagone P, Mangano K, Mammana S, Pesce A,
Pesce A, Caltabiano R, Giorlandino A, Portale TR, Cavalli E,
Lombardo GA, et al: Identification of novel targets for the
diagnosis and treatment of liver fibrosis. Int J Mol Med.
36:747–752. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Chen J, Lin Y and Sun Z: Deficiency in the
anti-aging gene Klotho promotes aortic valve fibrosis through
AMPKα-mediated activation of RUNX2. Aging Cell. 15:853–860. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Tchetina EV, Demidova NV, Markova GA,
Taskina EA, Glukhova SI and Karateev DE: Increased baseline RUNX2,
caspase 3 and p21 gene expressions in the peripheral blood of
disease-modifying anti-rheumatic drug-naïve rheumatoid arthritis
patients are associated with improved clinical response to
methotrexate therapy. Int J Rheum Dis. 20:1468–1480. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Hahn JM, Glaser K, McFarland KL, Aronow
BJ, Boyce ST and Supp DM: Keloid-derived keratinocytes exhibit an
abnormal gene expression profile consistent with a distinct causal
role in keloid pathology. Wound Repair Regen. 21:530–544. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Chen YT, Yao JN, Qin YT, Hu K, Wu F and
Fang YY: Biological role and clinical value of miR-99a-5p in head
and neck squamous cell carcinoma (HNSCC): A bioinformatics-based
study. FEBS Open Bio. 8:1280–1298. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Kang SU, Kim YS, Kim YE, Park JK, Lee YS,
Kang HY, Jang JW, Ryeo JB, Lee Y, Shin YS, et al: Opposite effects
of non-thermal plasma on cell migration and collagen production in
keloid and normal fibroblasts. PLoS One. 12:e01879782017.
View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Garcia-Morales V, Friedrich J, Jorna LM,
Campos-Toimil M, Hammes HP, Schmidt M and Krenning G: The
microRNA-7-mediated reduction in EPAC-1 contributes to vascular
endothelial permeability and eNOS uncoupling in murine experimental
retinopathy. Acta Diabetol. 54:581–591. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Fang B, Liu Y, Zheng D, Shan S, Wang C,
Gao Y, Wang J, Xie Y, Zhang Y and Li Q: The effects of mechanical
stretch on the biological characteristics of human adipose-derived
stem cells. J Cell Mol Med. 23:4244–4255. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) Method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Zhang M, Zeng J, Zhao Z and Liu Z: Loss of
MiR-424-3p, not miR-424-5p, confers chemoresistance through
targeting YAP1 in non-small cell lung cancer. Mol Carcinog.
56:821–832. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Rotgers E, Nurmio M, Pietilä E,
Cisneros-Montalvo S and Toppari J: E2F1 controls germ cell
apoptosis during the first wave of spermatogenesis. Andrology.
3:1000–1014. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Gene Ontology C; Gene Ontology Consortium,
: Gene Ontology Consortium: Going forward. Nucleic Acids Res.
43D:D1049–D1056. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Kanehisa M, Furumichi M, Tanabe M, Sato Y
and Morishima K: KEGG: New perspectives on genomes, pathways,
diseases and drugs. Nucleic Acids Res. 45D1:D353–D361. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Cavalli E, Battaglia G, Basile MS, Bruno
V, Petralia MC, Lombardo SD, Pennisi M, Kalfin R, Tancheva L,
Fagone P, et al: Exploratory Analysis of iPSCS-Derived Neuronal
Cells as Predictors of Diagnosis and Treatment of Alzheimer
Disease. Brain Sci. 10:1662020. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Zhang Y, Cheng C, Wang S, Xu M, Zhang D
and Zeng W: Knockdown of FOXM1 inhibits activation of keloid
fibroblasts and extracellular matrix production via inhibition of
TGF-β1/Smad pathway. Life Sci. 232:1166372019. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Berman B, Maderal A and Raphael B: Keloids
and Hypertrophic Scars: Pathophysiology, Classification, and
Treatment. Dermatol Surg. 43 (Suppl 1):S3–S18. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Bock O, Schmid-Ott G, Malewski P and
Mrowietz U: Quality of life of patients with keloid and
hypertrophic scarring. Arch Dermatol Res. 297:433–438. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Liu Q, Wang X and Jia Y: Heat Shock
Protein 90 Inhibitor Decreases Collagen Synthesis of Keloid
Fibroblasts and Attenuates the Extracellular Matrix on the Keloid
Spheroid Model. Plast Reconstr Surg. 137:759e–760e. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Philandrianos C, Kerfant N, Jaloux C Jr,
Martinet L, Bertrand B and Casanova D: Keloid scars (part I):
Clinical presentation, epidemiology, histology and pathogenesis.
Ann Chir Plast Esthet. 61:128–135. 2016.(In French). View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Chen J, Liu K, Liu Y, Wang X and Zhang Z:
Targeting mTORC1/2 with OSI-027 inhibits proliferation and
migration of keloid keratinocytes. Exp Dermatol. 28:270–275. 2019.
View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Ogawa R: Keloid and Hypertrophic Scars Are
the Result of Chronic Inflammation in the Reticular Dermis. Int J
Mol Sci. 18:6062017. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Komori T: Regulation of Proliferation,
Differentiation and Functions of Osteoblasts by Runx2. Int J Mol
Sci. 20:16942019. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Li G, Li YY, Sun JE, Lin WH and Zhou RX:
ILK-PI3K/AKT pathway participates in cutaneous wound contraction by
regulating fibroblast migration and differentiation to
myofibroblast. Lab Invest. 96:741–751. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Evangelisti C, Evangelisti C, Chiarini F,
Lonetti A, Buontempo F, Bressanin D, Cappellini A, Orsini E,
McCubrey JA and Martelli AM: Therapeutic potential of targeting
mTOR in T-cell acute lymphoblastic leukemia (review). Int J Oncol.
45:909–918. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Sharma VR, Gupta GK and Sharma AK, Batra
N, Sharma DK, Joshi A and Sharma AK: PI3K/Akt/mTOR Intracellular
Pathway and Breast Cancer: Factors, Mechanism and Regulation. Curr
Pharm Des. 23:1633–1638. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Steelman LS, Martelli AM, Cocco L, Libra
M, Nicoletti F, Abrams SL and McCubrey JA: The therapeutic
potential of mTOR inhibitors in breast cancer. Br J Clin Pharmacol.
82:1189–1212. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Ge F, Wang F, Yan X, Li Z and Wang X:
Association of BAFF with PI3K/Akt/mTOR signaling in lupus
nephritis. Mol Med Rep. 16:5793–5798. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Fagone P, Ciurleo R, Lombardo SD,
Iacobello C, Palermo CI, Shoenfeld Y, Bendtzen K, Bramanti P and
Nicoletti F: Transcriptional landscape of SARS-CoV-2 infection
dismantles pathogenic pathways activated by the virus, proposes
unique sex-specific differences and predicts tailored therapeutic
strategies. Autoimmun Rev. 19:1025712020. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Nicoletti F, Fagone P, Meroni P, McCubrey
J and Bendtzen K: mTOR as a multifunctional therapeutic target in
HIV infection. Drug Discov Today. 16:715–721. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Tu T, Huang J, Lin M, Gao Z, Wu X, Zhang
W, Zhou G, Wang W and Liu W: CUDC 907 reverses pathological
phenotype of keloid fibroblasts in vitro and in vivo via
dual inhibition of PI3K/Akt/mTOR signaling and HDAC2. Int J Mol
Med. 44:1789–1800. 2019.PubMed/NCBI
|
|
47
|
Aragona M and Blanpain C: Gene therapy:
Transgenic stem cells replace skin. Nature. 551:306–307. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Mavilio F, Pellegrini G, Ferrari S, Di
Nunzio F, Di Iorio E, Recchia A, Maruggi G, Ferrari G, Provasi E,
Bonini C, et al: Correction of junctional epidermolysis bullosa by
transplantation of genetically modified epidermal stem cells. Nat
Med. 12:1397–1402. 2006. View
Article : Google Scholar : PubMed/NCBI
|
|
49
|
Wang W, Qu M, Xu L, Wu X, Gao Z, Gu T,
Zhang W, Ding X, Liu W and Chen YL: Sorafenib exerts an anti-keloid
activity by antagonizing TGF-β/Smad and MAPK/ERK signaling
pathways. J Mol Med (Berl). 94:1181–1194. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Tian Y, Jin L, Zhang W, Ya Z, Cheng Y and
Zhao H: AMF siRNA treatment of keloid through inhibition signaling
pathway of RhoA/ROCK1. Genes Dis. 6:185–192. 2018. View Article : Google Scholar : PubMed/NCBI
|