Introduction
Diabetes mellitus is a common group of clinical
syndromes that are mainly manifested in disorders of glucose
metabolism and fluid imbalance (1,2). More
than 400 million patients are diagnosed with the disease worldwide,
and it has high mortality and disability rates (3). Notably, type-2 diabetes mellitus
(T2DM) accounts for up to 90% of these patients. T2DM is
characterized by hyperglycemia induced by multiple factors,
including genetic inheritance, various environmental factors (such
as exercise habits and work pressure) and lifestyle (4). T2DM is caused by impaired islet β-cell
function and insulin resistance (5).
In recent years, researchers have carried out
exploratory work on T2DM in the field of epigenetics, including DNA
methylation (6) and histone
acetylation (7), in an effort to
explain the epigenetic mechanism of T2DM (8). Changes in dietary habits cause
epigenetic changes (9). Epigenetic
research related to the environment is becoming increasingly
important in the prevention and treatment of diseases (10). DNA methylation is one of the most
well-studied epigenetic processes in transcriptional regulation and
gene silencing. PDX-1 is a gene that plays an important role in
pancreatic development and function (11,12).
Previous studies have suggested that the methylation of PDX-1 plays
a role in the development of T2DM (13,14).
Acarbose is an α-glucosidase inhibitor that
functions by competing with oligosaccharides to inhibit
α-glucosidase activity of intestinal wall cells, thereby delaying
the process of carbohydrate degradation and effectively delaying
the absorption of glucose by pancreatic tissues (15). However, the effect of acarbose on
PDX-1 methylation in islet β cells remains unclear. The purpose of
the study was to evaluate the effect of acarbose on PDX-1
methylation in islet β-cells in spontaneous type 2 diabetic db/db
mice.
Materials and methods
Animal studies and experimental
design
Male, 8-week-old db/db mice and male -+/db mice
(n=28; weight, ~18 g) were purchased from Shanghai Xipu-bikai
Experimental Animal Co., Ltd. The mice were housed at 22–25°C with
60±5%, 12-h light/dark cycles and sufficient water and were fed a
commercial diet over one week of adaptation. The mice were then
randomly divided into four groups. In the normal group (n=8), male
-+/db mice were fed a commercial diet. Mice with fasting blood
glucose >11.1 mmol/l were considered diabetic and selected in
the T2DM groups. In the T2DM group (n=8), mice were fed a high-fat
diet (T2DM model group). The mice in the acarbose-treated diabetic
group (T2DM+acarbose group; n=8) were given 9 g/kg/day acarbose
extract (cat. no. 1906208B; Zhongmei Huadong Pharmaceutical Co.,
Ltd.) by gavage for four weeks. The mice in the alkylresorcinol
(AR; cat. no. 3158-56-3; Sigma-Aldrich; Merck KGaA)-treated T2DM
group (T2DM+AR group; n=8) received 60 mg/kg/day AR extract by
gavage for 4 weeks as a positive control (16). This study was performed in strict
accordance with the NIH guidelines for the Care and Use of
Laboratory Animals (NIH Publication No. 85-23 Rev. 1985) and was
approved by the Animal Care and Use Committee of Zhejiang Chinese
Medical University (approval no. ZSLL-2019-102). Food intake, body
weight, serum total cholesterol (TC) and triglyceride (TG) levels,
glucagon, and fasting blood glucose (FBG) were measured. All mice
were euthanized with 5% isoflurane. Mice were monitored until
complete cessation of the heartbeat, sustained involuntary
respiration for 2–3 min and absence of blinking or toe contraction
reflex.
Routine measurements
During the experimental period, food intake, body
weight, and FBG were recorded weekly. Before measuring FBG, the
mice were fasted for 24 h (with sufficient water), blood (1 ml;
total 5 ml) was collected from the tail vein, and FBG was measured
with a blood glucose meter (Jiangsu Yuyue Medical Equipment &
Supply Co., Ltd.) weekly until the mice were sacrificed. Four weeks
after treatment, glycosylated hemoglobin (HbA1c) was measured using
a DCA2000 analyzer (Bayer AG). The levels of glucagon were measured
by ELISA (cat. no. MOES01072; USCN Kit, Inc.).
Lipid profiles in serum
The serum total cholesterol (TC) and triglyceride
(TG) levels were measured by direct enzymatic methods using a CX-7
Biochemical Autoanalyser (Beckman Coulter, Inc.).
Glucose and insulin tolerance
tests
Intraperitoneal glucose tolerance tests (IPGTTs) and
insulin tolerance tests (ITTs) were performed on different days.
Four weeks after treatment, all mice were fasted overnight before
analysis. Glucose was administered at a dose of 2 g/kg by
intraperitoneal injection, and tail vein blood glucose levels were
measured using a glucose meter (Yuyue301) at 0, 30, 60, and 120
min. The mice were injected with 1 U/kg humulin R (Eli Lilly) after
fasting for 4 h for the IPGTT. The glucose levels were measured at
0, 30, 60, 90 and 120 min after injection.
Methylation-specific PCR (MSP)
The MSP was performed on a 7300 thermocycler
(Applied Biosystems; Thermo Fisher Scientific, Inc.). The primers
for Methylation PDX-1 and unmethylated PDX-1 were based on the
genomic sequence (accession nos. NT_024524100 and NT_024524) of the
PDX-1 gene. Primer sequences were as follows: i) Unmethylated
PDX-1-forward, 5′-GCCTCGAGCGCAGACTGAGTCCAGAGTG-3′ and reverse,
5′-CGAAGCTTCGGTGATTTCTTCAGGGAAA-3′; ii) Methylation PDX-1-forward,
5′-GTTTTTTTCGGAGGGAGTC-3′ and reverse, 5′-TTTCCACGCGTAAACTTTAA-3′.
Cellular DNA was extracted using the QIamp DNA mini kit (Qiagen
GmbH) from mouse pancreatic samples according to the manufacturers
instructions. Bisulphate modification of the DNA was performed
using the EZ Methylation-Gold kit (Zymo Research, Corp.) according
to the manufacturers protocol. DNA elements with methylated
cytosins remained unmodified after bisulfite treatment, whereas
unmethylated cytosins were completely converted into uracil
nucleosides and further to thymine after PCR, thus giving different
primer recognition sequences for MSP. PCR-mix (2X; 12.5 µl;
Fermentas; Thermo Fisher Scientific, Inc.) was added to 2 µl
bisulfonated/unmodified DNA sample followed by 0.8 µl forward and
reverse primers (400 nM each) and 8.5 µl water.
The PCR reaction was run using the Taq GC Buffer
amplification system (Takara Biotechnology Co., Ltd.) and the
following thermocycling conditions: 94°C for 2 min; followed by 35
cycles at 94°C for 10 sec, 60–70°C for 20 sec and 72°C for 20 sec;
and terminated at 72°C for 4 min followed by immediate cooling to
4°C.
After staining with 6X loading dye solution
(Fermantas; Thermo Fisher Scientific, Inc.), the PCR products were
analyzed on 2% agarose gels and visualized under UV illumination
using a LabWorks gel imaging system (version 4.6; UVP, LLC).
Extraction of nuclear extracts and
pancreatic tissue homogenate
Nuclear extracts were isolated from pancreatic
tissues using NE-PER™ Extraction Reagent (Thermo Fisher Scientific,
Inc.), according to the manufacturers protocol. Frozen pancreatic
tissues were placed on ice and pulverized in a pre-cooled mortar
with liquid nitrogen. Tissues were lysed in 500 µl RIPA buffer
(Beyotime Institute of Biotechnology) supplemented with phosphatase
and protease inhibitor cocktail (Thermo Fisher Scientific, Inc.)
and then centrifuged at 10,000 × g for 20 min at 4°C. The soluble
extracts were collected for western blotting. The determination of
protein concentrations was performed using a bicinchoninic acid
assay (Thermo Fisher Scientific, Inc.).
Western blot analysis
Soluble lysates (40 µg) and nuclear extracts (20 µg)
were separated by SDS-PAGE on 8% gels and transferred to PVDF
membranes (EMD Millipore). The membranes were blocked with 5%
nonfat milk in TBS-T (10 mM Tris, pH 7.6, 150 mM NaCl, and 0.05%
Tween-20) for 2 h at room temperature and probed with primary
antibodies against PDX-1 (42 kDa; rabbit; 1:1,000; cat. no. 5679;
Cell Signaling Technology), β-actin (45 kDa; rabbit; 1:1,000; cat.
no. ab8227; Abcam) and PCNA (36 kDa; rabbit; 1:1,000; cat. no.
13110; Cell Signaling Technology) overnight at 4°C. A
peroxidase-conjugated goat anti-mouse IgG secondary antibody
(1:2,000; cat. no. sc-516102; Santa Cruz Biotechnology, Inc.) was
then added for 1.5 h at room temperature. The bands were visualized
using an enhanced chemiluminescence kit (Beyotime Institute of
Biotechnology) and quantified using ImageJ software (version 6.0;
National Institutes of Health).
β-cell proliferation assessment
After fixation by perfusion with 4% paraformaldehyde
for 5 min at room temperature, the mouse pancreas was removed.
After sectioning into 5-µm, dewaxing and microwave antigen
recovery, the slides were briefly incubated with 0.2% Triton X-100
and 10% BSA (cat. no. A3858; Sigma-Aldrich; Merck KGaA), then kept
overnight at 4°C with guinea pig anti-insulin (cat. no. ab7842;
1:1,000; Abcam) and mouse anti-BrdU (cat. no. ab8152; 1:1,000;
Abcam) antibodies followed by Alexa Fluor 568 goat anti-guinea pig
(cat. no. AP108B; 1:1,000; Sigma-Aldrich; Merck KGaA) and FITC goat
anti-mouse (cat. no. ab6785; 1:1,000; Abcam) secondary antibodies.
Nuclei were counterstained with DAPI (cat. no. ab104139; Abcam).
All BrdU+ cells and insulin+ cells (representing total β-cells)
from 50 pancreatic islets per mouse were counted using a confocal
microscope (Zeiss AG). The percentage of β-cell proliferation was
determined by dividing the number of BrdU+ β-cells by the total
number of β-cells.
Immunohistochemistry
Pancreatic tissues were fixed in 10%
neutral-buffered formalin for 5 min at room temperature and
embedded in paraffin. After a xylene wash to remove the paraffin
and a rehydration step with serial dilutions of alcohol, the 5-µm
thick sections were incubated in 0.3% H2O2
for 15 min to block endogenous peroxidases and heated at 98°C for
15 min for antigen retrieval. Samples were blocked with 5% BSA for
1 h at room temperature. The sections were then incubated overnight
at 4°C with primary antibodies against inducible Neurogenin 3
(NGN3; cat. no. sc-374442; mouse; 1:60; Santa Cruz Biotechnology,
Inc.), Insulin (mouse; cat. no. BM4310; 1:50; Wuhan Boster
Biological Technology, Ltd.), PDX-1 (cat. no. 5679; rabbit; 1:50;
Cell Signaling Technology, Inc.) and Forkhead box O1 (FoXO1; cat.
no. 2880; rabbit, 1:100; Cell Signaling Technology). After a series
of washes, peroxidase-conjugated goat anti-mouse IgG/goat
anti-rabbit IgG secondary antibodies (cat. nos. sc-516102 and
sc-2357; 1:100; Santa Cruz Biotechnology, Inc.) were applied. The
sections were stained with diaminobenzidine solution (OriGene
Technologies, Inc.) washed, dehydrated, permeabilized, mounted and
examined under bright-field microscopy. Images were examined using
an LSM 710 laser scanning confocal microscope (Carl Zeiss AG).
Statistical analysis
Data are presented as the mean ± SEM of at least
three independent experiments. Statistical analysis was performed
using SPSS version 13.0 (SPSS, Inc.). One-way ANOVA followed by
Bonferroni correction was used for multi-group comparisons. A
t-test was performed to identify significant differences between
two groups. P<0.05 were considered to indicate a statistically
significant difference.
Results
Effect of acarbose on glucose and
lipid metabolism in diabetic mice
T2DM mice exhibited a significant increase and body
weight, compared with normal mice (Fig.
1A and B). Furthermore, T2DM mice demonstrated a significant
increase in FBG compared with normal mice, and the increased FBG
levels were maintained throughout the entire experiment (Fig. 1C). These results are considered
common signs of diabetes. Importantly, in all diabetic mice mice,
glucagon, HbA1c, TC, TG and LDL-C levels were all significantly
increased, compared with the normal group (Fig. 1D-H), while HDL-C levels remained
unchanged (Fig. 1I), indicating
that hyperlipidemia symptoms were also successfully established in
the diabetes group. Moreover, acarbose significantly decreased
glucagon, HbA1c, TC, TG and LDL-C levels in serum, compared with
untreated T2DM mice.
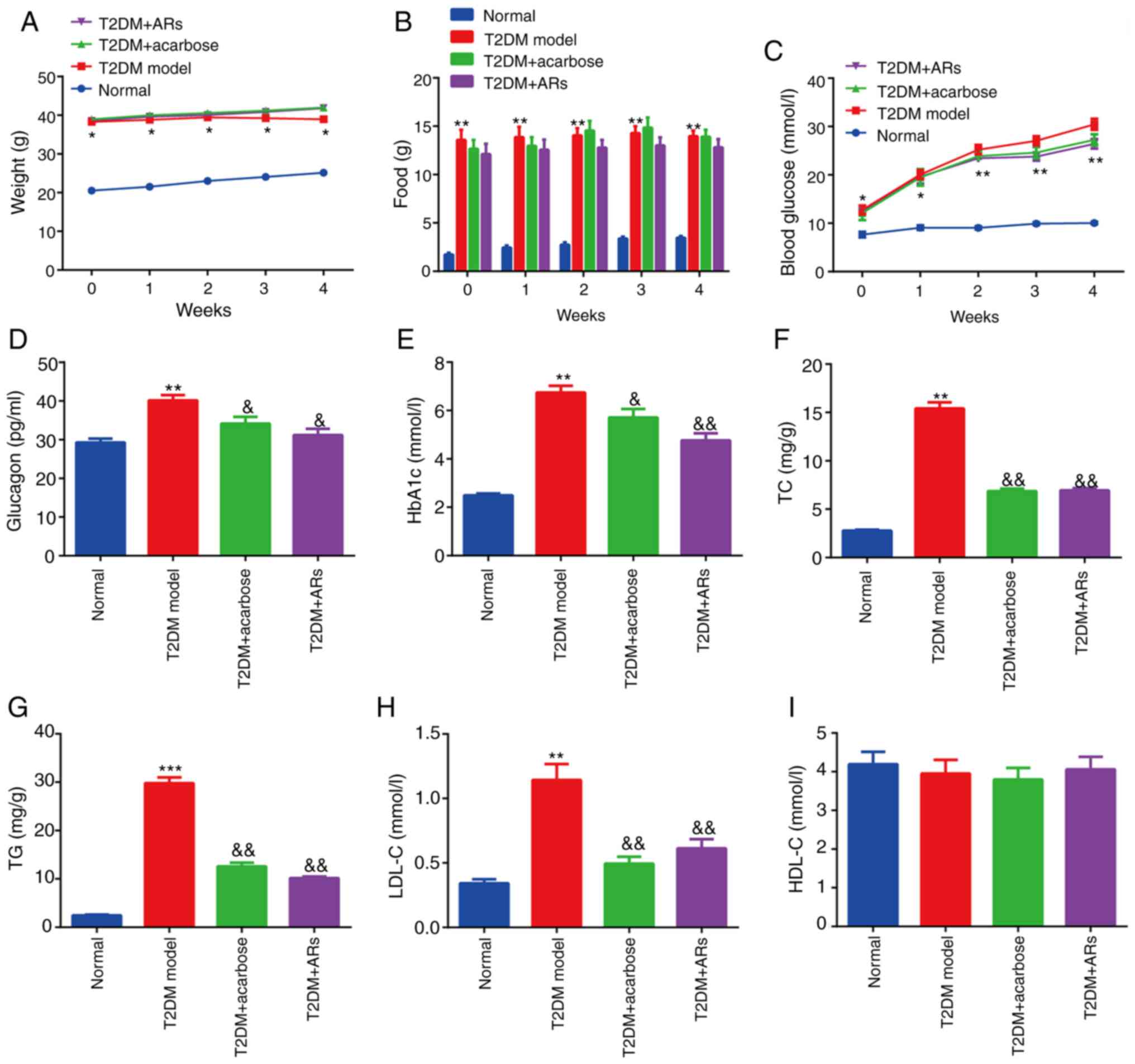 | Figure 1.Effect of acarbose on glucose and
lipid metabolism in diabetic mice. (A) Weight, (B) food, (C)
fasting blood glucose, (D) glucagon, (E) HbA1c, (F) TC, (G) TG (H)
LDL-C and (I) HDL-C levels were determined. n=8. *P<0.05,
**P<0.01, ***P<0.001, vs. normal group;
&P<0.05, &&P<0.01, vs. T2DM
group. AR, alkylresorcinol; T2DM, type-2 diabetes mellitus; HbA1c,
glycated hemoglobin; TC, total cholesterol; TG, triglycerides;
LDL-C, low-density lipoprotein-cholesterol; HDL-C, high-density
lipoprotein-cholesterol. |
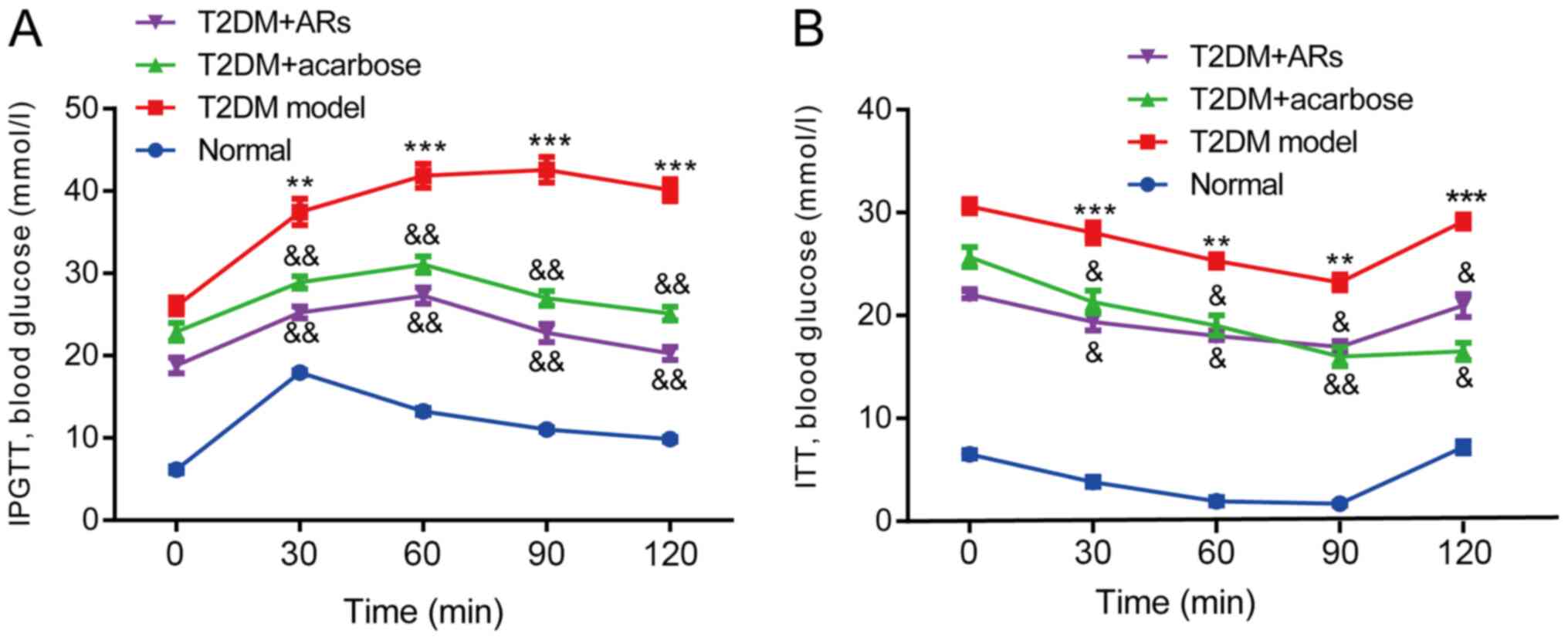
Effect of acarbose on the IPGTT and
ITT
Blood glucose levels were measured at 30, 60, 90 and
120 min. In the normal group, the initial and final blood glucose
levels were similar. By contrast, blood glucose levels remained
high in T2DM mice at the 120-min timepoint, compared with the
initial timepoint (Fig. 2A).
Interestingly, blood glucose significantly decreased in the
acarbose- or AR-treated groups, compared, with T2DM mice,
suggesting that acarbose could reduce glucose rapidly and stably.
Similarly, in the ITT, the acarbose-treated mice displayed reduced
blood glucose levels at 30, 60, 90 and 120 min compared to the T2DM
model mice (Fig. 2B).
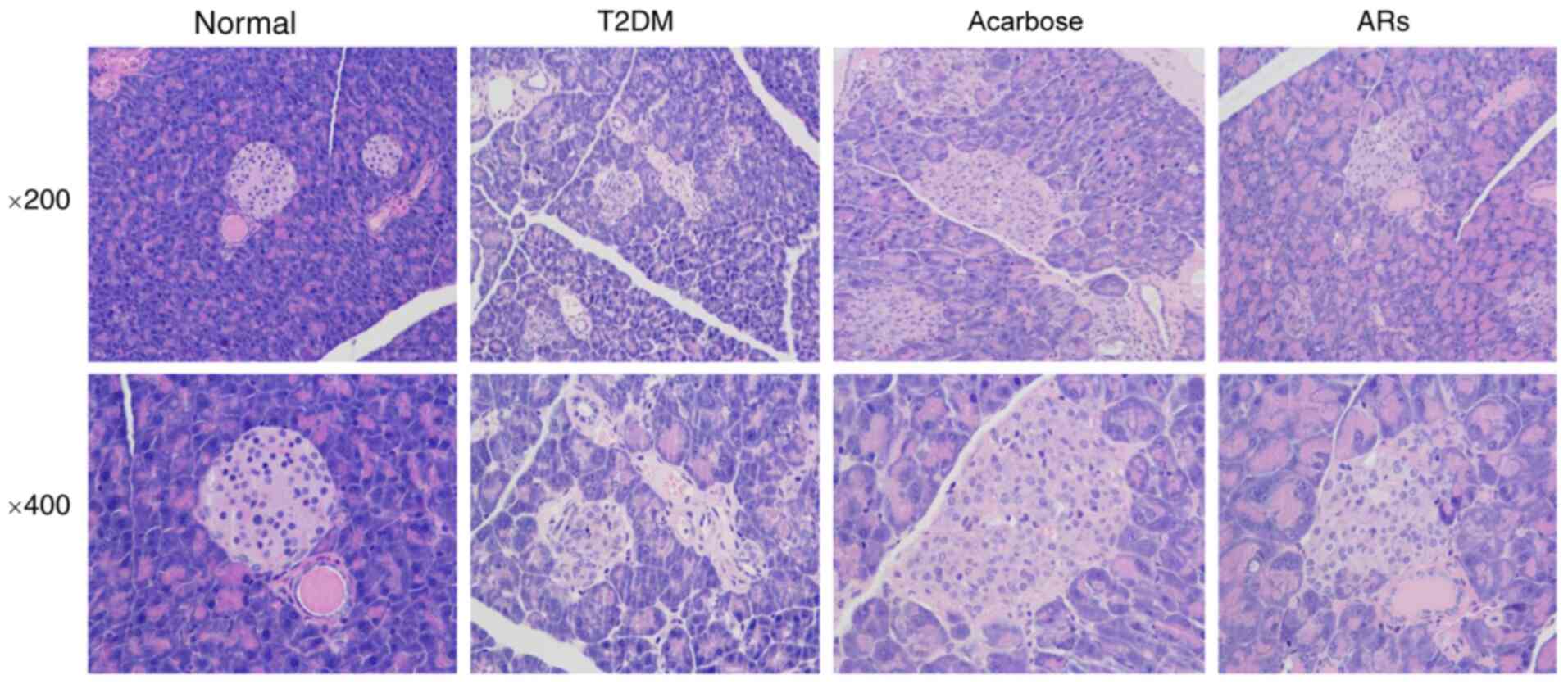
Effects of acarbose on pancreatic
islet number and structure in diabetic mice
HE staining of pancreatic tissue demonstrated that
high-fat diet administration elicited severe injury to the
pancreas, including decreased islet cells and diminished pancreatic
islet diameter. Additionally, the structure of the pancreatic
islets was disordered, vacuoles appeared, and nuclei were swollen
in T2DM mice. These effects were reversed by acarbose (Fig. 3).
Effects of acarbose on insulin, PDX-1,
FoxO1 and Ngn3 expression in islets
Active PDX-1 induces the efficient production of
insulin. The strong nuclear expression of PDX-1 was shown in the
islet β-cells of NC pancreatic tissues, while PDX-1 staining was
weak in the DM group and was restored by acarbose (Fig. 4A). Conversely, strong nuclear
expression of FoxO1 was observed in the pancreatic tissues of T2DM
model mice, which was reversed by acarbose (Fig. 4A). The Ngn3 level in the pancreas
was obviously increased in the T2DM model mice and was obviously
decreased by acarbose (Fig. 4A).
Moreover, strong insulin staining was observed in normal mice but
weak staining was observed in T2DM model mice. However, acarbose
increased insulin levels (Fig. 4A).
the level of GLP-1 was determined, and the results showed that
GLP-1 level was decreased in T2DM group compared to normal group,
which partly reversed by acarbose and alkylresorcinol (Fig. 4B). Similarly, the number of
insulin-positive/BrdU-positive β-cells increased dramatically in
the T2DM + acarbose group compared with the T2DM group (Fig. 4C).
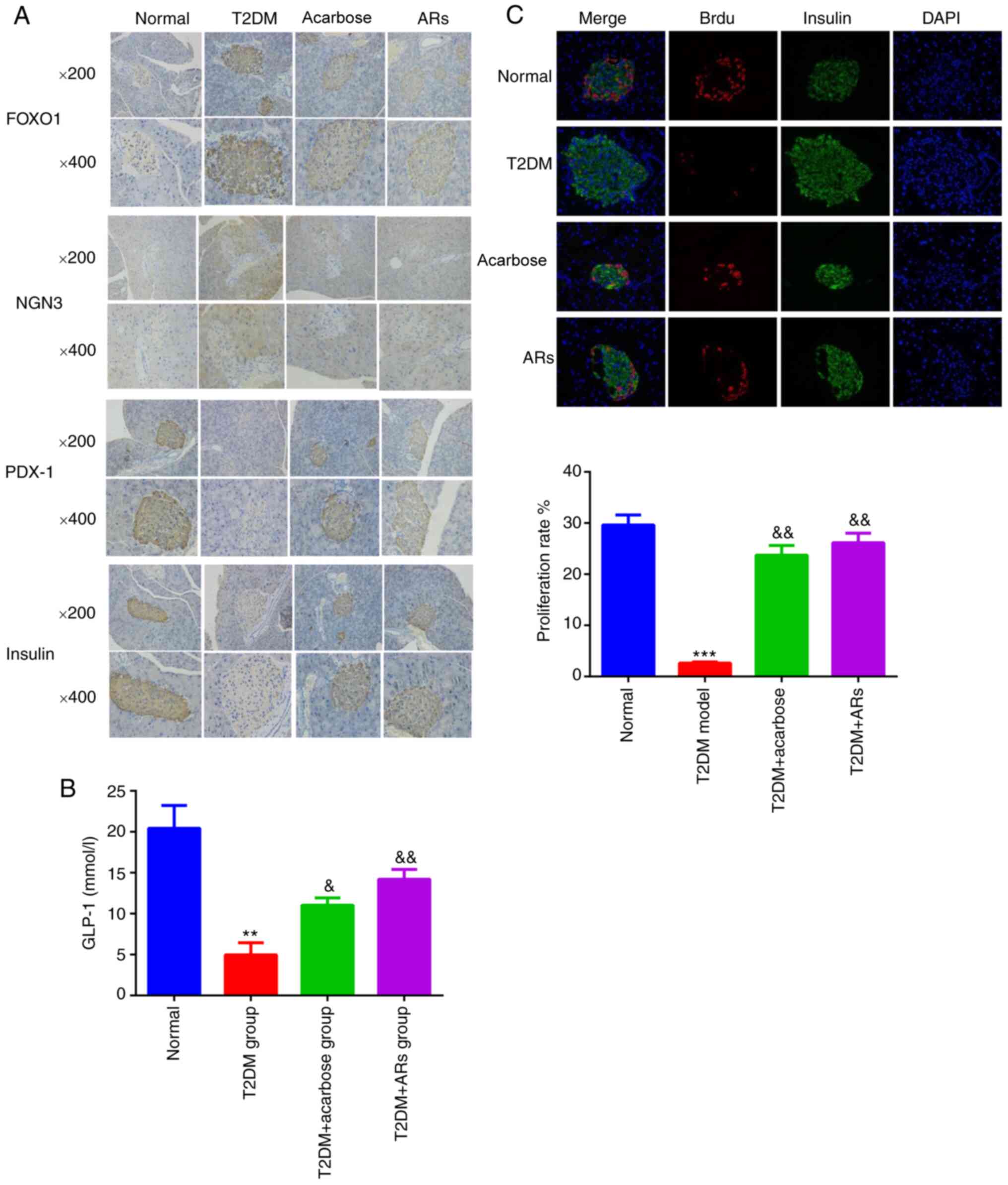 | Figure 4.Effects of acarbose on insulin, PDX-1,
FoxO1 and NGN 3 expression in islets. (A) Insulin, PDX-1, FoxO1 and
Ngn3 expression in the islet β-cells of pancreatic tissue sections.
Magnification, ×200 or ×400. (B) GLP-1 levels. (C) Numbers of
insulin-positive and BrdU-positive β-cells. n=8. **P<0.01,
***P<0.001, vs. the normal group; &P<0.05,
&&P<0.01, vs. the T2DM group. AR,
alkylresorcinol; T2DM, type-2 diabetes mellitus; FOXO1, forkhead
box O1; NGN3, neurogenin 3; PDX-1, pancreatic and duodenal
homeobox-1; GLP-1, zinc finger GATA-like protein 1. |
Effects of acarbose on PDX-1 promoter
hypermethylation in diabetic mice
Hypermethylation of the PDX-1 promoter was not
observed in normal mice but was significantly increased in T2DM
model mice. Additionally, PDX-1 hypermethylation was inhibited by
acarbose and alkylresorcinol (Fig.
5A-C). Moreover, a significant redistribution of PDX-1 from the
nucleus to the cytoplasm was observed in pancreatic tissues of T2DM
model mice compared with that in normal mice. In contrast, acarbose
prevented the nuclear export of PDX-1 in T2DM model mice (Fig. 5D).
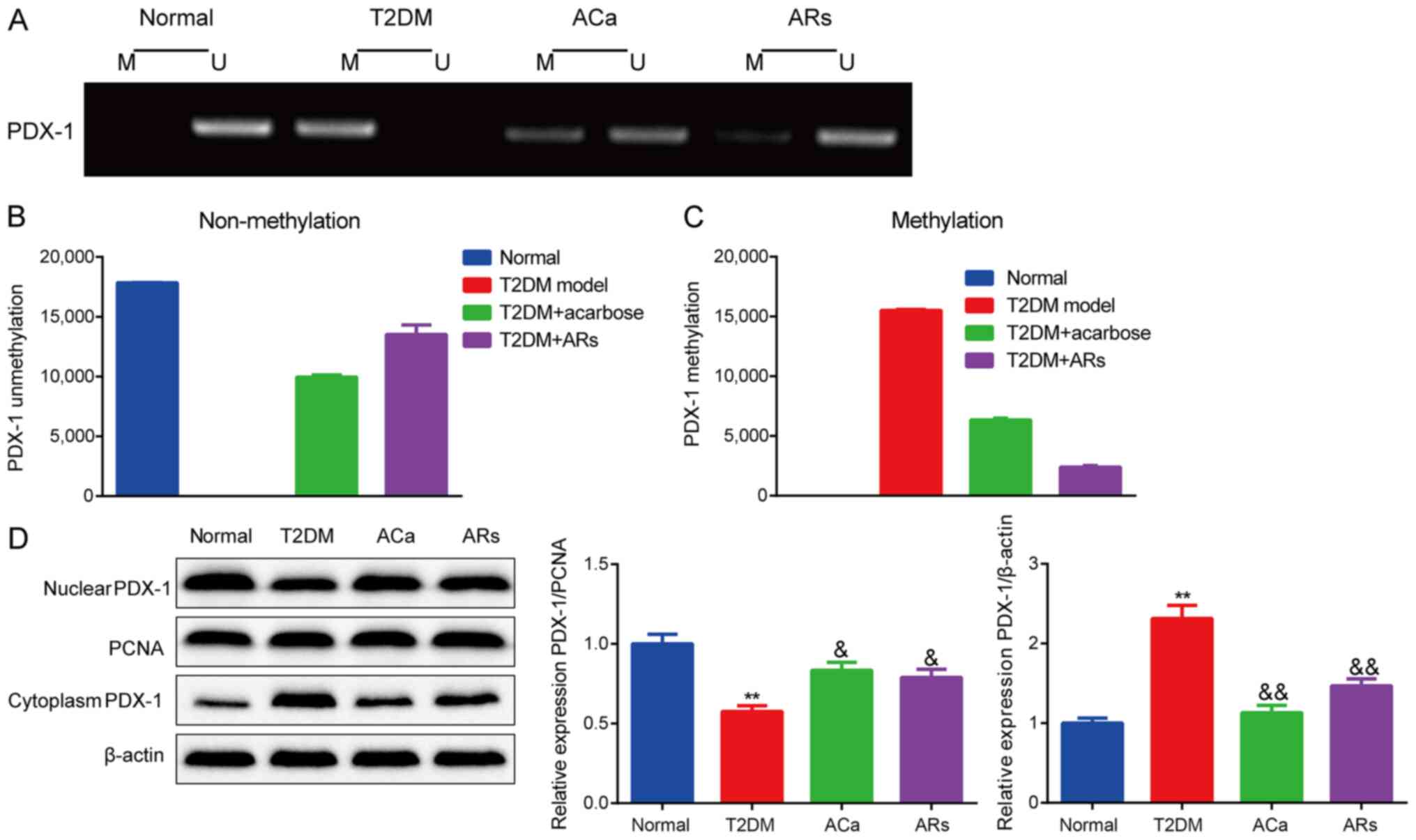 | Figure 5.Effects of acarbose on PDX-1 promoter
hypermethylation in diabetic mice. (A-C) PDX-1 hypermethylation was
inhibited by acarbose and ARs. (D) Acarbose prevented the nuclear
export of PDX-1 in T2DM model mice, PCNA and β-actin were used as
loading controls for nuclear and cytoplasmic fractions,
respectively. n=8. **P<0.01, vs. normal group;
&P<0.05, &&P<0.01, vs. T2DM
group. AR, alkylresorcinol; T2DM, type-2 diabetes mellitus; PDX-1,
pancreatic and duodenal homeobox-1; PCNA, proliferating cell
nuclear antigen; M methylated; U, unmethylated. |
Discussion
The prevalence of T2DM has been increasing each
year, in parallel to the increase in the population with a high
risk for diabetes development (17). Acarbose is one of the safest
antidiabetic agents available and is commonly prescribed for the
treatment or prevention of T2DM (18). The most important function of
pancreatic β-cells is to maintain blood glucose homeostasis by
secreting insulin upon glucose stimulation. Loss of pancreatic
islet mass and β-cell dysfunction accompanied by a decrease in
insulin synthesis are crucial factors for initiating the
development of DM (19). The
present study confirmed that acarbose could alleviate pancreatic
β-cell damage and impaired insulin secretion in T2DM mice by
reducing the methylation of PDX-1.
Insulin resistance is reported to be involved in the
disorder of glucose metabolism and lipid metabolism (20). In the present study, a diabetic
mouse model was established that exhibited hyperglycemia and
insulin resistance. Acarbose significantly reduced blood glucose
levels and improved insulin sensitivity in diabetic mice. It also
reduced the elevated levels of glucagon, TC and TG seen in diabetic
mice and promoted β-cell proliferation. In addition, acarbose
promoted the nuclear expression of PDX-1 with a concomitant
decrease in acetylated FoxO1 levels, both of which led to increased
production of insulin.
PDX-1 is predominately expressed in β-cells and can
induce β-cell differentiation and insulin transcription (21). Conversely, mutation or
downregulation of PDX-1 results in β-cell dysfunction and glucose
intolerance in humans and animals (22), indicating that the activation of
PDX-1-regulated responses is an important therapeutic target for
preventing the development of DM. The present study demonstrated
increased cytoplasmic PDX-1 levels and decreased nuclear PDX-1 in
β-cells from diabetic mice, compared with control mice, and
acarbose blocked the diabetes-mediated nucleocytoplasmic shuttling
of PDX-1, suggesting that acarbose could promote insulin production
by preserving nuclear PDX-1. Interestingly, in PDX-1+
β-cells, FoxO1 is largely localized in the cytoplasm, but in
PDX-1− β-cells, FoxO1 mainly accumulates in the nucleus
(23), suggesting the mutual
exclusion of FoxO1 and PDX-1. Ngn3 is a basic helix-loop-helix
protein and a progenitor cell marker in islets (24). Moreover, the increase in Ngn3
resulted from the dedifferentiation of pancreatic β-cells in the
islets and was probably attributed to the decrease in β-cells
(25). A significant increase in
Ngn3 levels was observed in the model group compared with the
control group. Moreover, the levels of insulin, glucagon and PDX-1
in islets were increased in the T2DM group. These data may support
the hypothesis that alterations to the genes leads to a decrease in
β-cells and that acarbose could prevent β-cell proliferation.
A previous study suggested that DNA methylation may
be involved in controlling cell-specific PDX-1 expression in human
islets (22). In support of this,
increased PDX-1 methylation was observed in T2DM mouse β-cells,
leading to reduced PDX-1 levels. Moreover, acarbose blocked the
increase in methylated PDX-1 in T2DM mouse β-cells.
However, our study had some limitations. First, the
levels of DNA methylases Dnmt1, Dnmt3a or Dnmt3b were not measured
in islets. Second, whether the Notch1/Ngn3 signaling pathway
mediates dedifferentiation needs further examination in
vitro. Third, the effect of acarbose on oxidative stress was
not analyzed.
In summary, the present study demonstrated that
acarbose may improve glucose metabolism in db/db mice. Furthermore,
acarbose maintained β-cell identity by preventing proliferation
through the regulation of Ngn3, FoxO1 and PDX-1. This process
probably involved PDX-1 methylation.
Acknowledgements
Not applicable.
Funding
This work was supported by the National Natural
Science Foundation Youth Fund Project (grant no. 81703996),
National Natural Science Foundation (grant no. 81774217) and Topic
of Zhejiang Traditional Chinese Medicine Administration (grant nos.
2017ZKL016 and 2019ZB096).
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors contributions
DZ and XM designed the study. LC contributed to the
literature search. DZ wrote the initial draft of the manuscript. XM
reviewed and edited the manuscript. All authors read and approved
the final manuscript.
Ethics approval and consent to
participate
All procedures performed in studies involving
animals were approved by The Animal Care and Use Committee of
Zhejiang Chinese Medical University (approval no.
ZSLL-2019-102).
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Sharma AK, Bharti S, Goyal S, Arora S,
Nepal S, Kishore K, Joshi S, Kumari S and Arya DS: Upregulation of
PPARγ by Aegle marmelos ameliorates insulin resistance and β-cell
dysfunction in high fat diet fed-streptozotocin induced type 2
diabetic rats. Phytother Res. 25:1457–1465. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Orchard TJ, Olson JC, Erbey JR, Williams
K, Forrest KY, Smithline Kinder L, Ellis D and Becker DJ: Insulin
resistance-related factors, but not glycemia, predict coronary
artery disease in type 1 diabetes: 10-year follow-up data from the
Pittsburgh epidemiology of diabetes complications study. Diabetes
Care. 26:1374–1379. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Regazzi R: MicroRNAs as therapeutic
targets for the treatment of diabetes mellitus and its
complications. Expert Opin Ther Targets. 22:153–160. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Oza MJ and Kulkarni YA: Biochanin A
improves insulin sensitivity and controls hyperglycemia in type 2
diabetes. Biomed Pharmacother. 107:1119–1127. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Wing RR, Rosen RC, Fava JL, Bahnson J,
Brancati F, Gendrano Iii IN, Kitabchi A, Schneider SH and Wadden
TA: Effects of weight loss intervention on erectile function in
older men with type 2 diabetes in the Look AHEAD trial. J Sex Med.
7:156–165. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Zhou Z, Sun B, Li X and Zhu C: DNA
methylation landscapes in the pathogenesis of type 2 diabetes
mellitus. Nutr Metab (Lond). 15:472018. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Sharma S and Taliyan R: Histone
deacetylase inhibitors: Future therapeutics for insulin resistance
and type 2 diabetes. Pharmacol Res. 113:320–326. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Malipatil N, Lunt M, Narayanan RP, Siddals
K, Cortés Moreno GY, Gibson MJ, Gu HF, Heald AH and Donn RP:
Assessment of global long interspersed nucleotide element-1
(LINE-1) DNA methylation in a longitudinal cohort of type 2
diabetes mellitus (T2DM) individuals. Int J Clin Pract. e13270Oct
21–2018.(Epub ahead of print). PubMed/NCBI
|
|
9
|
Hudec M, Dankova P, Solc R, Bettazova N
and Cerna M: Epigenetic regulation of circadian rhythm and its
possible role in diabetes mellitus. Int J Mol Sci. 21:30052020.
View Article : Google Scholar
|
|
10
|
Paneni F, Costantino S, Volpe M, Lüscher
TF and Cosentino F: Epigenetic signatures and vascular risk in type
2 diabetes: A clinical perspective. Atherosclerosis. 230:191–197.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Pedica F, Beccari S, Pedron S, Montagna L,
Piccoli P, Doglioni C and Chilosi M: PDX-1 (pancreatic/duodenal
homeobox-1 protein 1). Pathologica. 106:315–321. 2014.PubMed/NCBI
|
|
12
|
Zhou G, Yu J, Wang A, Liu SH,
Sinnett-Smith J, Wu J, Sanchez R, Nemunaitis J, Ricordi C,
Rozengurt E and Brunicardi FC: Metformin restrains pancreatic
duodenal homeobox-1 (PDX-1) function by inhibiting ERK signaling in
pancreatic ductal adenocarcinoma. Curr Mol Med. 16:83–90. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Rafati R, Jalal R, Asoodeh A and Matin MM:
Association of rs12255372 (TCF7L2) and D76N (PDX-1) polymorphisms
with type 2 diabetes in a population living in Northeast Iran. Arch
Iran Med. 18:376–378. 2015.PubMed/NCBI
|
|
14
|
Kaneto H, Matsuoka TA, Kimura T, Obata A,
Shimoda M, Kamei S, Mune T and Kaku K: Appropriate therapy for type
2 diabetes mellitus in view of pancreatic β-cell glucose toxicity:
‘The earlier, the better’. J Diabetes. 8:183–189. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
McIver LA and Tripp J: Acarbose.
StatPearls. StatPearls Publishing Copyright © 2020. StatPearls
Publishing LLC.; Treasure Island (FL): 2020, simplehttps://www.ncbi.nlm.nih.gov/books/NBK493214Aughust
10–2020
|
|
16
|
Tu J, Liu G, Cao X, Zhu S, Li Q, Ji G, Han
Y and Xiao H: Hypoglycemic effects of wheat bran alkyresorcinols in
high-fat/high-sucrose diet and low-dose streptozotocin-induced type
2 diabetic male mice and protection of pancreatic β cells. Food
Funct. 10:3282–3290. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Triplitt C, Cersosimo E and DeFronzo RA:
Pioglitazone and alogliptin combination therapy in type 2 diabetes:
A pathophysiologically sound treatment. Vasc Health Risk Manag.
6:671–690. 2010.PubMed/NCBI
|
|
18
|
Li FF, Xu XH, Fu LY, Su XF, Wu JD, Lu CF,
Ye L and Ma JH: Influence of acarbose on plasma glucose
fluctuations in insulin-treated patients with type 2 diabetes: A
pilot study. Int J Endocrinol. 2015:9035242015. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Butler AE, Janson J, Bonner-Weir S, Ritzel
R, Rizza RA and Butler PC: Beta-cell deficit and increased
beta-cell apoptosis in humans with type 2 diabetes. Diabetes.
52:102–110. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Wang X, Zhou L, Li G, Luo T, Gu Y, Qian L,
Fu X, Li F, Li J and Luo M: Palmitate activates AMP-activated
protein kinase and regulates insulin secretion from beta cells.
Biochem Biophys Res Commun. 352:463–468. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Shao S, Fang Z, Yu X and Zhang M:
Transcription factors involved in glucose-stimulated insulin
secretion of pancreatic beta cells. Biochem Biophys Res Commun.
384:401–404. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Brissova M, Shiota M, Nicholson WE, Gannon
M, Knobel SM, Piston DW, Wright CV and Powers AC: Reduction in
pancreatic transcription factor PDX-1 impairs glucose-stimulated
insulin secretion. J Biol Chem. 277:11225–11232. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Kitamura T, Nakae J, Kitamura Y, Kido Y,
Biggs WH III, Wright CV, White MF, Arden KC and Accili D: The
forkhead transcription factor Foxo1 links insulin signaling to Pdx1
regulation of pancreatic beta cell growth. J Clin Invest.
110:1839–1847. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Zhu Y, Liu Q, Zhou Z and Ikeda Y: PDX1,
neurogenin-3, and MAFA: Critical transcription regulators for beta
cell development and regeneration. Stem Cell Res Ther. 8:2402017.
View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Van de Casteele M, Leuckx G, Baeyens L,
Cai Y, Yuchi Y, Coppens V, De Groef S, Eriksson M, Svensson C,
Ahlgren U, et al: Neurogenin 3+ cells contribute to β-cell
neogenesis and proliferation in injured adult mouse pancreas. Cell
Death Dis. 4:e5232013. View Article : Google Scholar : PubMed/NCBI
|