Introduction
Knee osteoarthritis (KOA) is a common degenerative
disease in the orthopedic field, which has a high disability rate.
With the increasing aging of the social population, the incidence
of KOA has increased to 45% in the last 20 years, and the incidence
of KOA has exceeded 80% among people >65 years old (1). KOA is a chronic degenerative joint
disease caused by multifactorial etiologies involving the loss of
articular cartilage, subchondral bone sclerosis, cyst formation,
osteophyte development, chronic pain, stiffness and lower extremity
disability, which significantly affect the independence and quality
of life of patients, primarily among elderly individuals (2).
Pyroptosis is a caspase-1-dependent form of
programmed cell death that substantially differs from apoptosis,
and involves canonical and non-canonical inflammasome activation
pathways (3). Nod-like receptor
protein (NLRP) inflammasomes, mainly the NLRP1 and NLRP3
inflammasomes, belong to a group of seven widely accepted
inflammasome complexes: NLRP1, NLRP3, NLRC4, interferon γ inducible
protein 16, pyrin, caspase-4 and absent in melanoma 2 (4). NLRPs directly interact with
apoptosis-ssociated speck-like protein with a caspase-recruitment
domain (ASC) to activate caspase-1 (cleaved caspase-1p10 and
caspase-1p20), which in turn promotes gasdermin D (GSDMD) cleavage
and translocation (5–7). This leads to the formation of a hole
in the cell membrane that facilitates the release of intracellular
inflammatory factors, such as IL-1β and IL-18 and ultimately
results in cell death (8). Previous
studies have reported that fibroblast-like synoviocytes (FLSs)
undergo lipopolysaccharide (LPS) + ATP-induced pyroptosis in a
caspase-1-dependent manner, mediated by activation of the NLRP1 and
NLRP3 inflammasomes (9,10).
High mobility group box 1 (HMGB1) was first
identified as a nuclear protein that regulates transcription,
replication and DNA repair (11).
Extracellular HMGB1 has been identified as a crucial cytokine
involved in the intestinal inflammatory response (12) and multiple inflammatory diseases,
including osteoarthritis (13),
rheumatoid arthritis (14),
endotoxemia (15), epilepticus
(16), stroke (17), diabetes (18), innate immunity, collagen disease,
atherosclerosis, cancer and acute lung injury (19,20).
HMGB1 can be secreted by FLSs after stimulation with inflammatory
factors in KOA (21–25), and HMGB1 levels in synovial tissues
increase during KOA progression and correlate positively with KOA
synovitis (26). HMGB1 can also
stimulate chondrocytes to secrete MMPs, as well as promote
cartilage autophagy and degeneration in the pathogenesis of KOA
(21). Moreover, the HMGB1-LPS
complex is reportedly transported into macrophages via RNA for
advanced-glycation end products (RAGE)-dependent endocytosis, and
intracellular LPS activates caspase-1 and leads to pyroptosis
(27), indicating HMGB1 as an
important factor in the inflammatory response in KOA. However, the
specific mechanism via which FLSs promote HMGB1 secretion in KOA is
yet to be elucidated. Notably, a previous study revealed that
ASC-mediated caspase-1 activation can promote abnormal HMGB1
secretion during alveolar macrophage pyroptosis in acute lung
injury (28). Therefore, the
relationship between HMGB1 secretion and FLS pyroptosis in KOA
requires further investigation.
The present study aimed to investigate the
relationship between HMGB1 secretion and FLS pyroptosis in
vivo and in vitro. Small interfering RNAs (siRNAs) and
inhibitors were used to further elucidate the specific effect of
FLS pyroptosis on HMGB1 secretion.
Materials and methods
Animals and samples
A total of 15 male Sprague-Dawley (SD) rats (Animal
House Grant Certificate no. 201810A001; Beijing Vital River
Laboratory Animal Technology Co., Ltd.; age, 2 months; weight,
200–300 g) were maintained in a specific pathogen-free laminar-flow
housing apparatus, under controlled temperature (25±2°C) and
humidity (60±5%) conditions with a 12-h light/dark cycle, all rats
had free access to food and water. All animal protocols were
approved by the Animal Care and Use Committee of the Nanjing
University of Chinese Medicine. All experiments were conducted in
accordance with the National Institutes of Health Guidelines for
the Care and Use of Laboratory Animals (29).
The rats were randomly assigned to three groups: the
normal group (n=5), the KOA group (n=5) and the caspase-1 inhibitor
group (n=5). KOA was induced in the KOA and caspase-1 inhibitor
groups via anterior cruciate ligament transection (ACLT) as
previously described (30). Rats
were anesthetized with an intraperitoneal injection of 30 mg/kg
pentobarbital sodium (3%). The joint capsule was cut along its
medial side (normal group), or the anterior cruciate ligament was
transected (KOA, caspase-1 inhibitor and HMGB1 inhibitor groups).
The surgical results were examined via the front drawer test
(31). Amikacin (Mutian Animal
Pharmaceutical Co., Ltd.) was injected into the thigh of each rat
at a dose of 10 mg/kg per day for 4 days after the operation. The
rats were placed in cages allowing free movement after the
operation, and the joints were not fixed. From day 14, the rats in
the caspase-1 inhibitor group were given intra-articular injection
of caspase-1 inhibitor VX765 (MedChemExpress, Inc.) at a dose of 50
mg/kg in 50 µl sterilized physiological saline once a day for 2
weeks. Rats in the normal and KOA groups were given intra-articular
injection of 50 µl sterilized physiological saline. At day 28, all
rats were sacrificed, and synovial tissues and cartilage were
harvested. Synovial tissues were collected to detect protein and
gene expression levels, as well as caspase-1 activity. Cartilage
was subjected to hematoxylin and eosin (H&E) staining to
evaluate damage. Serum proteins were detected using specific ELISA
kits.
FLS culture and treatment
Synovial tissues removed from five 2-month-old male
SD rats were snipped into pieces of ~3 mm3, homogenized
in DMEM (Gibco; Thermo Fisher Scientific, Inc.) and incubated for 1
h at 37°C with 1 mg/ml type I collagenase (Sigma-Aldrich; Merck
KGaA). The samples were filtered through a 100-µm cell strainer.
After dissociation, the FLSs were pelleted via centrifugation at
300 × g at ~25°C for 5 min and plated in DMEM supplemented with 10%
FBS (Gibco; Thermo Fisher Scientific, Inc.) and 1% antibiotics (100
U/ml penicillin and 100 µg/ml streptomycin; Invitrogen; Thermo
Fisher Scientific, Inc.). Cells were cultured at 37°C in a
humidified atmosphere with 95% air and 5% CO2, and were
identified as described in our previous studies (9,10).
Primary FLSs from passages 3–5 were used for subsequent
experiments.
To induce pyroptosis, FLSs were stimulated with LPS
(3 µg/ml; Sigma-Aldrich; Merck KGaA) in DMEM for 12 h and then
treated with ATP (Beijing Solarbio Science & Technology Co.,
Ltd.) at 37°C (3 mM) for 4 h. For the control group, FLSs were
treated with DMEM for 12 h and then treated with DMEM in
combination with 0.9% saline of the same volume as ATP for 4 h.
Compounds in the supernatant were detected using specific ELISA
kits. FLSs were collected to detect protein and gene expression
levels, and caspase-1 activity, as well as to perform
immunofluorescence.
siRNA transfection
To inhibit NLRP1, NLRP3, ASC and caspase-1 mRNA
expression levels in FLSs, commercially available specific siRNAs
and control siRNAs (Guangzhou RiboBio Co., Ltd.) were used. The
core nucleotide of siRNAs were as follows: NLRP1 siRNA,
5′-GGUGGAGCUGCAUCACAUATT-dTdT-3′ and Vehicle,
5′-UUCUCCGAACGUGUCACGU-dTdT-3′; NLRP3 siRNA,
5′-GGAGAGACCUUUAUGAGAATT-dTdT-3′ and Vehicle,
5′-UUCUCCGAACGUGUCACGU-dTdT-3′; ASC siRNA,
5′-CTGATAAACTCGTCAGCTA-dTdT-3′ and Vehicle,
5′-UUCUCCGAACGUGUCACGU-dTdT-3′; and caspase-1 siRNA,
5′-GGGCAAGCCAGAUGUUUAU-dTdT-3′ and Vehicle,
5′-UUCUCCGAACGUGUCACGU-dTdT-3′. The vehicles are the negative
control siRNAs.
FLSs were plated in 6-well plates with the density
of 1×104/ml and transfected with siRNAs using
Lipofectamine® 2,000 (Invitrogen; Thermo Fisher
Scientific, Inc.) according to the manufacturer's instructions.
siRNAs were diluted in the transfection reagent and culture medium,
and the FLSs in each plate were incubated with 20 pmol siRNA at
37°C for 6 h before the addition of LPS + ATP to induce pyroptosis.
To assess whether siRNA transfection was successful, the
verification experiments were conducted independently before the
formal transfection experiments. FLSs plated in 6-well plates with
the density of 1×104/ml were divided into control, siRNA
and vehicle groups.
Western blot analysis
Synovial tissues from individual rats were
homogenized and lysed in RIPA lysis buffer (Beyotime Institute of
Biotechnology). FLSs in different groups were also lysed in RIPA
lysis buffer. Lysates were collected and centrifuged at 4°C at
12,000 × g for 10 min, and protein concentrations were quantified
with a BCA protein assay kit (Beyotime Institute of Biotechnology).
Individual samples (20 µg/lane) were separated via 10% SDS-PAGE and
were transferred onto PVDF membranes (Beyotime Institute of
Biotechnology). The membranes were blocked with 5% BSA (Beyotime
Institute of Biotechnology) at 25°C for 2 h and then incubated
overnight at 4°C with primary antibodies against the following
proteins: Caspase-1p10 (1:1,000; cat. no. sc-56036; Santa Cruz
Biotechnology, Inc.), NLRP3 (1:500; cat. no. ab214185; Abcam),
NLRP1 (1:1,000; cat. no. sc-166368; Santa Cruz Biotechnology,
Inc.), pro-IL-1β (1:2,000; cat. no. ab9722; Abcam), IL-1β (1:1,000;
cat. no. ab9722; Abcam), GSDMD-C (1:500; cat. no. sc-393581; Santa
Cruz Biotechnology, Inc.), HMGB1 (1:5,000; cat. no. ab79823; Abcam)
and β-actin (1:2,000; cat. no. 60008-1-Ig; ProteinTech Group,
Inc.). The membranes were then incubated with horseradish
peroxidase-conjugated Affinipure goat anti-rabbit IgG (H+L)
(1:20,000; cat. no. SA00001-2; ProteinTech Group, Inc.) for 2 h at
room temperature. The bands were visualized using an enhanced
chemiluminescent (Thermo Fisher Scientific, Inc.) method, and the
grayscale values of the protein bands (average gray value) were
quantified with Photoshop CS5 (Adobe Systems, Inc.) using β-actin
as an internal reference: Target protein gray value/internal
reference gray value.
Reverse transcription-quantitative PCR
(RT-qPCR)
Total RNA was extracted from synovial tissues and
FLSs using an RNA-Quick Purification kit (ES Science Biotech Co.,
Ltd.; www.esunbio.com). RNA concentration and
purity were measured with a spectrophotometer, and the target 260
nm/280 nm ratio was between 1.8 and 2.0. RT was performed using a
HiScriptQ RT SuperMix kit (cat. no. R222-01; Vazyme Biotech Co.,
Ltd.) according to the manufacturer's instructions (stage 1, 1 rep,
50°C, 15 min; stage 2, 1 rep, −80°C, 5 sec). RNA expression was
measured with a ChamQ SYBR qPCR Master Mix kit (cat. no. Q331;
Vazyme Biotech Co., Ltd.) and an ABI 7500 real-time PCR system
(Applied Biosystems; Thermo Fisher Scientific, Inc.) according to
the manufacturer's instructions (stage 1, 1 rep, 95°C, 30 sec;
stage 2, 40 reps, 95°C, 10 sec, 60°C, 30 sec; and stage 3, 1 rep,
95°C, 15 sec, 60°C, 60 sec, 95°C, 15 sec). GAPDH was used as an
internal reference. All the primer sequences are provided in
Table I. All reactions were
performed in triplicate, and the results were analyzed using the
2−ΔΔCq method (32). The
experiment was repeated ≥3 times.
 | Table I.Upstream primers and downstream
primers for HMGB1 and GAPDH. |
Table I.
Upstream primers and downstream
primers for HMGB1 and GAPDH.
| Gene | Primer | Sequences
(5′→3′) |
|---|
| HMGB1 | Forward |
CTGCCTTCTCTTGTGACAAAGTGGAC |
|
| Reverse |
ACATACTCAGCACCAGCATCACC |
| GAPDH | Forward |
CCTCCTTCGGCCTTCTTCTTGTTC |
|
| Reverse |
TCATCCGCAGCAGTGTTGTTCC |
Caspase-1 activity assay
To investigate pyroptosis, caspase-1 activity in
individual samples was determined based on p-nitroaniline (pNA)
production using a colorimetric assay kit (cat. no. C1101; Beyotime
Institute of Biotechnology). Briefly, after detecting the caspase-1
protein concentration in individual cellular and synovial tissue
samples, the samples were diluted to 1–3 mg/ml. Triplicate aliquots
of each sample (10 µl) were incubated with 10 µl Ac-YVAD-pNA (2 mM)
in 96-well microplates at 37°C for 100 min, and caspase-1 activity
in individual wells was measured based on the absorbance at 405 nm
using a Multimode plate reader (PerkinElmer, Inc.).
Hoechst 33342/PI double staining
Hoechst 33342/PI double staining was used for
morphological analysis. FLSs were plated in replicates at
1×106 cells per well in 6-well plates. The cells were
collected via centrifugation at 300 × g for 5 min ~25°C, suspended
in cell staining buffer, incubated with 5 µl Hoechst 33342 and 5 µl
PI solution (cat. no. CA1120; Beijing Solarbio Science &
Technology Co., Ltd.) at 4°C for 30 min and observed under a
fluorescence microscope (magnification, ×200; Leica Microsystems
GmbH).
ELISA
HMGB1 and IL-1β levels in serum and culture
supernatants were assessed using specific ELISA kits (HMGB1. cat.
no. F15640; IL-1β. cat. no. F15810; Westang Biotechnology Co.,
Ltd.) according to the manufacturer's instructions. The blood (~6
ml per rat) was collected from the abdominal aorta of rats and the
serum sample was collected via centrifugation at 1,000 × g for 15
min at 4°C. Experimental and control samples were tested
simultaneously in triplicate.
Histopathological analysis
Tissue was collected from rats and fixed with 4%
paraformaldehyde at 25°C for 72 h, soaked in 15% EDTA at 25°C for
12 weeks for decalcification, then tissue was dehydrated in alcohol
for 2 h. The dehydrated tissue was treated with xylene solution for
dehydration and then was paraffin-embedded. Paraffin-embedded
tissue was cut into sections (thickness, 5 µm). Then,
paraffin-embedded sections were treated with xylene solution for 30
min for dewaxing. Tissue sections were treated with gradient
concentration of ethanol (100, 90, 80, 70 and 65%; 2 min per
concentration) for rehydration. The sections were then stained in
hematoxylin solution at 25°C for 3–5 min and rinsed with running
water for 30 sec, then stained sections were divided with
hydrochloric acid for 30 sec and rinsed again with running water
for 15 min. Stained sections were stained in eosin solution at 25°C
for 2–3 min after dehydration in alcohol with the concentration of
70 and 90% for 10 min. Then, the stained sections were placed into
xylene solution for 20 min. Finally, an appropriate amount of
neutral gum was added, the glass was covered and sealed. All the
stained sections were observed under optical electron microscope
(magnification, ×200). The degree of KOA in individual rats was
assessed by observing cartilage destruction using the Mankin's
scoring system (0, normal; 1, mild; 2, moderate; 3, severe).
Statistical analysis
Each experiment was repeated three times.
Statistical analysis was performed using GraphPad Prism 6.0
Software (GraphPad Software, Inc.). Data are presented as the mean
± SD. Group comparisons were assessed with the one-way ANOVA with
Bonferroni's post hoc test or Student's t-test, or two-way ANOVA
with Bonferroni's post hoc test for comparison of multiple columns.
P<0.05 was considered to indicate a statistically significant
difference.
Results
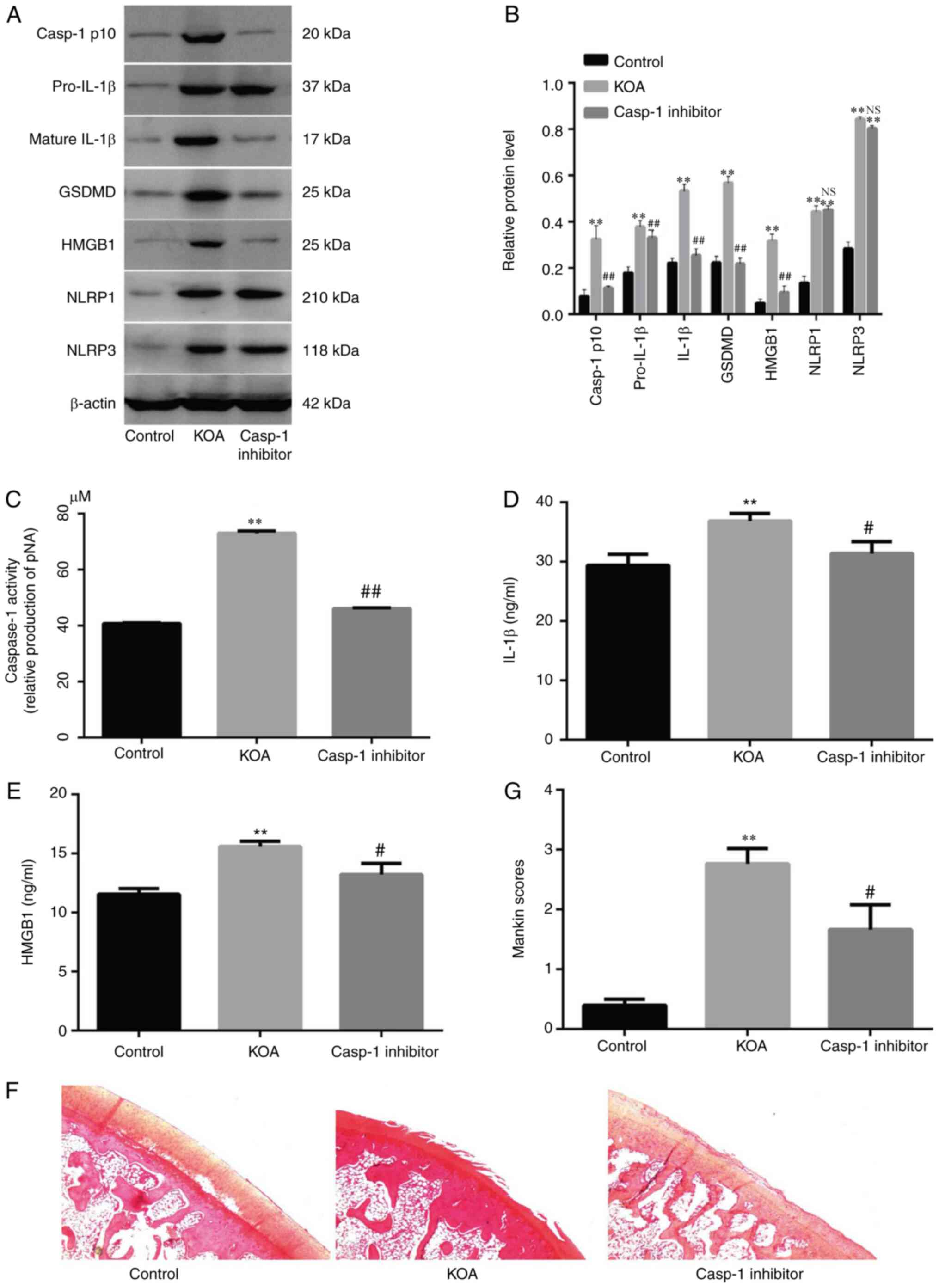
ACLT-induced pyroptosis increases
HMGB1 secretion in vivo
ACLT significantly increased the expression levels
of pyroptosis-associated proteins (NLRP1, NLRP3, GSDMD, IL-1β,
pro-IL-1β and caspase-1 p10; Fig. 1A, B
and D) and HMGB1 (Fig. 1A and
E) in synovial tissue and serum; however, these effects were
attenuated by a caspase-1 inhibitor, which also decreased caspase-1
activity in synovial tissue (Fig.
1C). It was identified that cartilage degeneration occurred in
ACLT-induced inflammatory arthritis and was markedly inhibited by
the caspase-1 inhibitor (Fig.
1F).
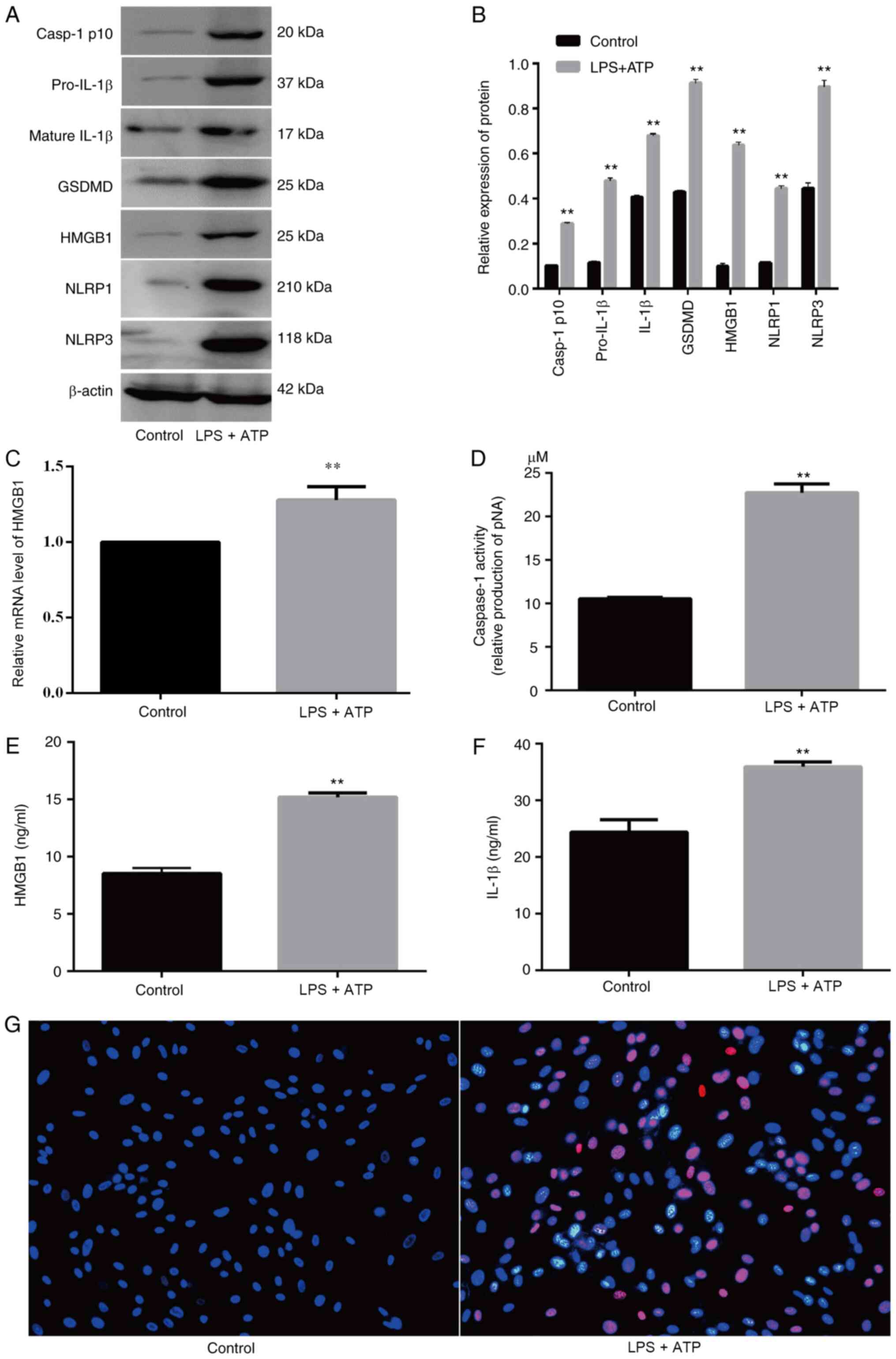
LPS + ATP-induced FLS pyroptosis
increases HMGB1 secretion in vitro
LPS + ATP significantly induced FLS pyroptosis,
which was accompanied by increased caspase-1 activity (Fig. 2D) and upregulated expression levels
of pyroptosis-related proteins (NLRP1, NLRP3, GSDMD, IL-1β,
pro-IL-1β and caspase-1 p10; Fig. 2A, B
and F). The FLSs with pyroptosis were stained pink (Fig. 2G). Moreover, LPS + ATP significantly
increased HMGB1 levels (Fig. 2A-C and
E).
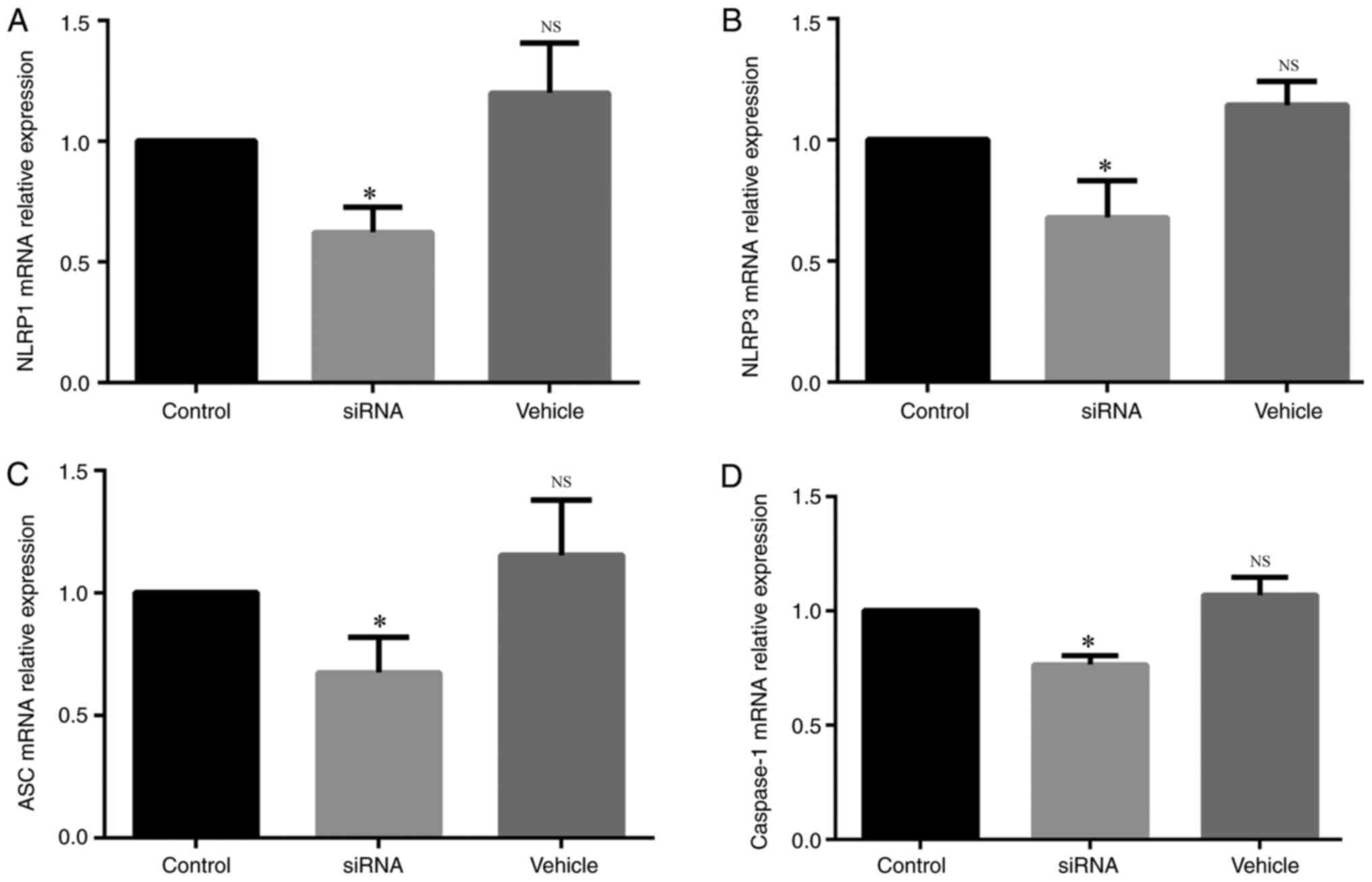
siRNA transfection decreases mRNA
expression in vitro
To observe whether the siRNA transfection was
successful, the different siRNAs were used to knockdown the mRNA
expression levels of NLRP1, NLRP3, ASC and caspase-1. The mRNA
expression of NLRP1 was significantly lower in the siRNA group
compared with the control group (Fig.
3A), and the same results were identified for the mRNA
expression levels of NLRP3 (Fig.
3B), ASC (Fig. 3C) and
caspase-1 (Fig. 3D).
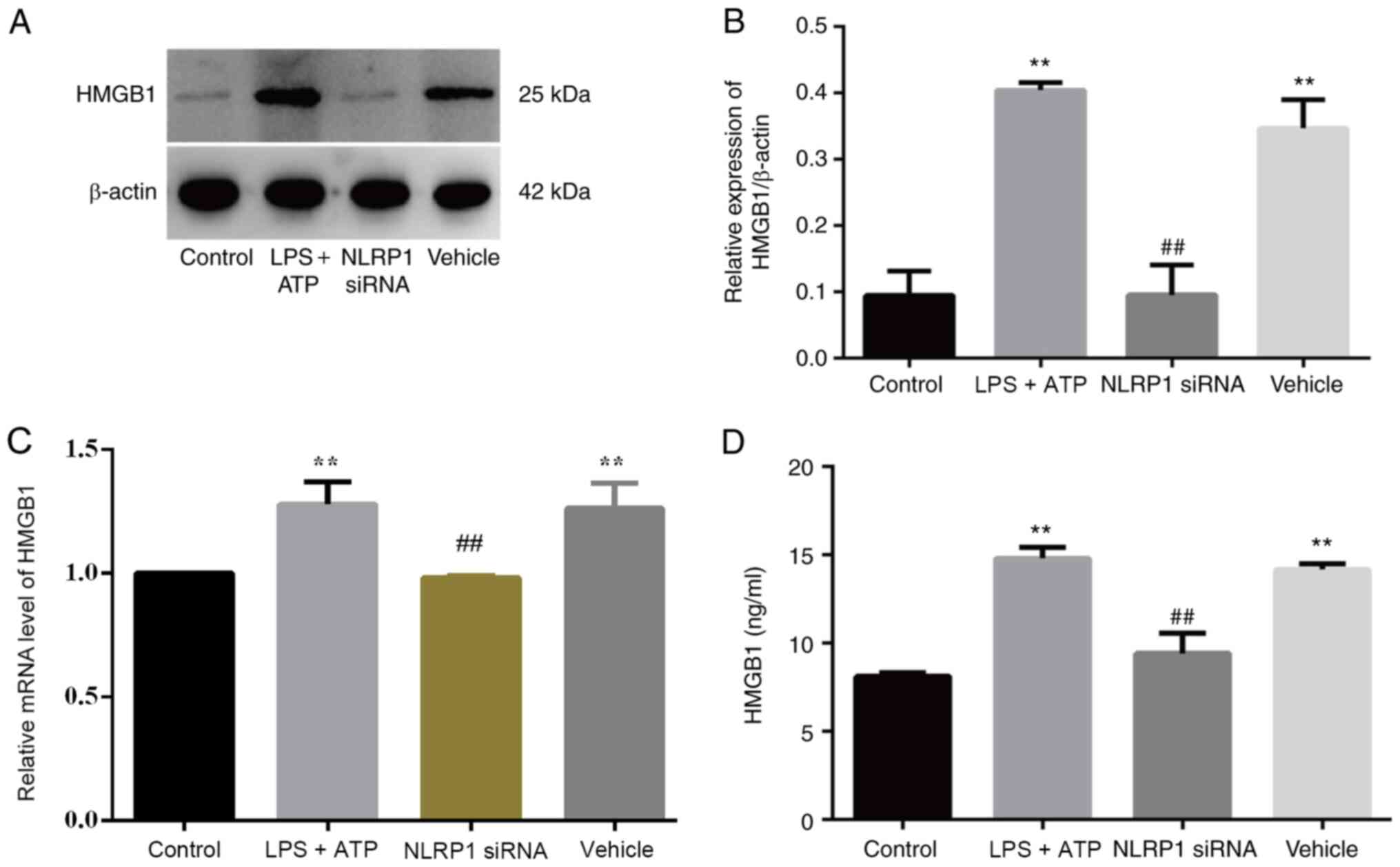
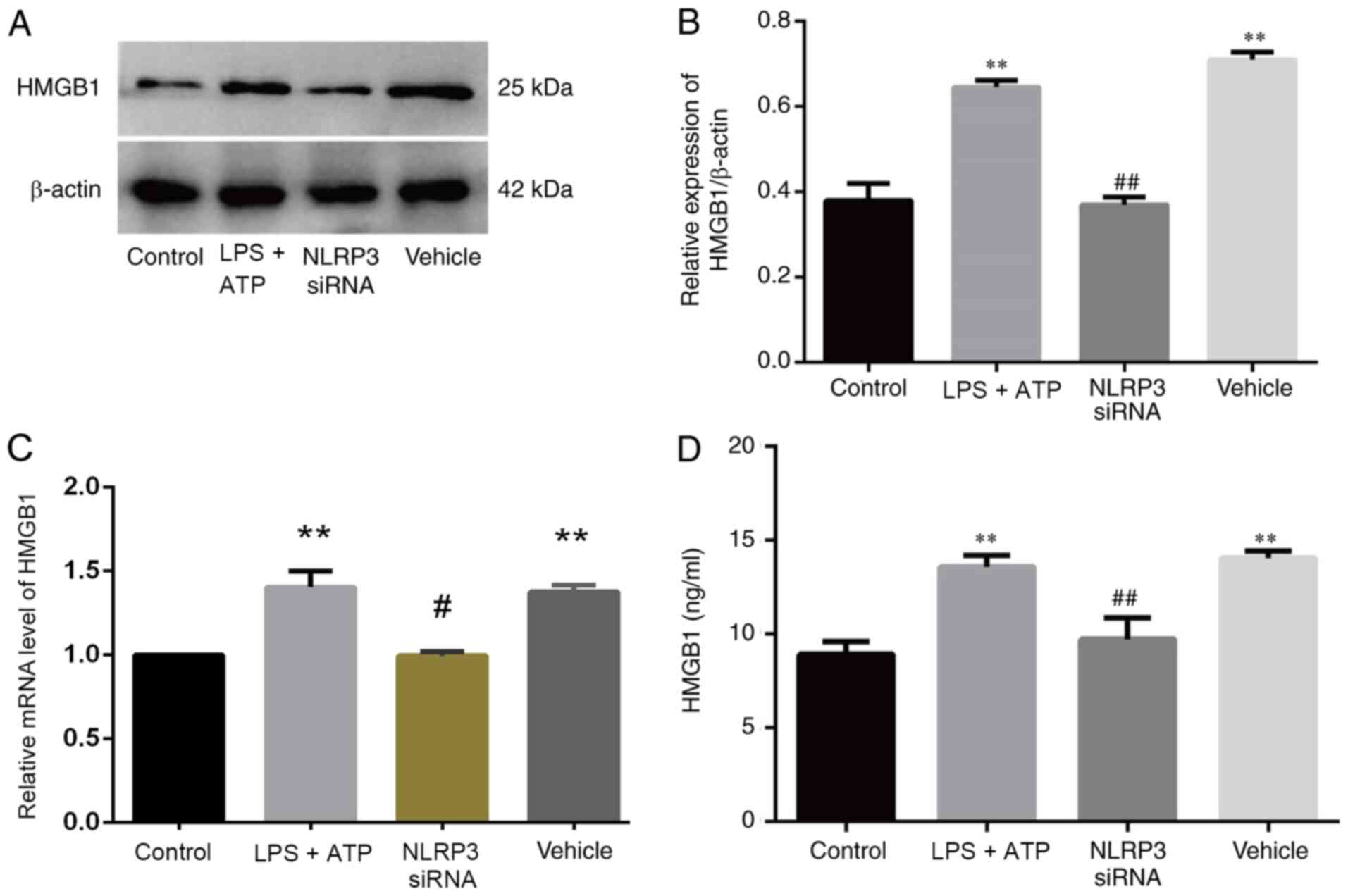
NLRP1 knockdown attenuates HMGB1
secretion in vitro
Our previous study revealed that NLRP1 inflammasomes
are important in the pathogenesis of KOA as they induce FLS
pyroptosis (9). To observe the
specific effect of NLRP1 on HMGB1 secretion, FLSs were exposed to a
NLRP1-specific siRNA or the control. HMGB1 protein expression was
significantly higher in the LPS + ATP and vehicle groups compared
with the NLRP1 siRNA and control groups (Fig. 4A and B), and similar results were
obtained for relative HMGB1 mRNA expression (Fig. 4C) and HMGB1 levels in the
supernatant (Fig. 4D).
NLRP3 knockdown attenuates HMGB1
secretion in vitro
It was previously proved that NLRP3 inflammasomes
are involved in the pathogenesis of KOA by inducing FLS pyroptosis
(9). Thus, NLRP3-specific siRNA was
used to assess the effect of NLRP3 on HMGB1 secretion. Healthy FLSs
served as the control group. HMGB1 protein expression (Fig. 5A and B) was significantly higher in
the LPS + ATP and vehicle groups compared with in the NLRP3 siRNA
and control groups. Similar results were obtained for relative
HMGB1 RNA expression (Fig. 5C) and
HMGB1 levels in the supernatant (Fig.
5D).
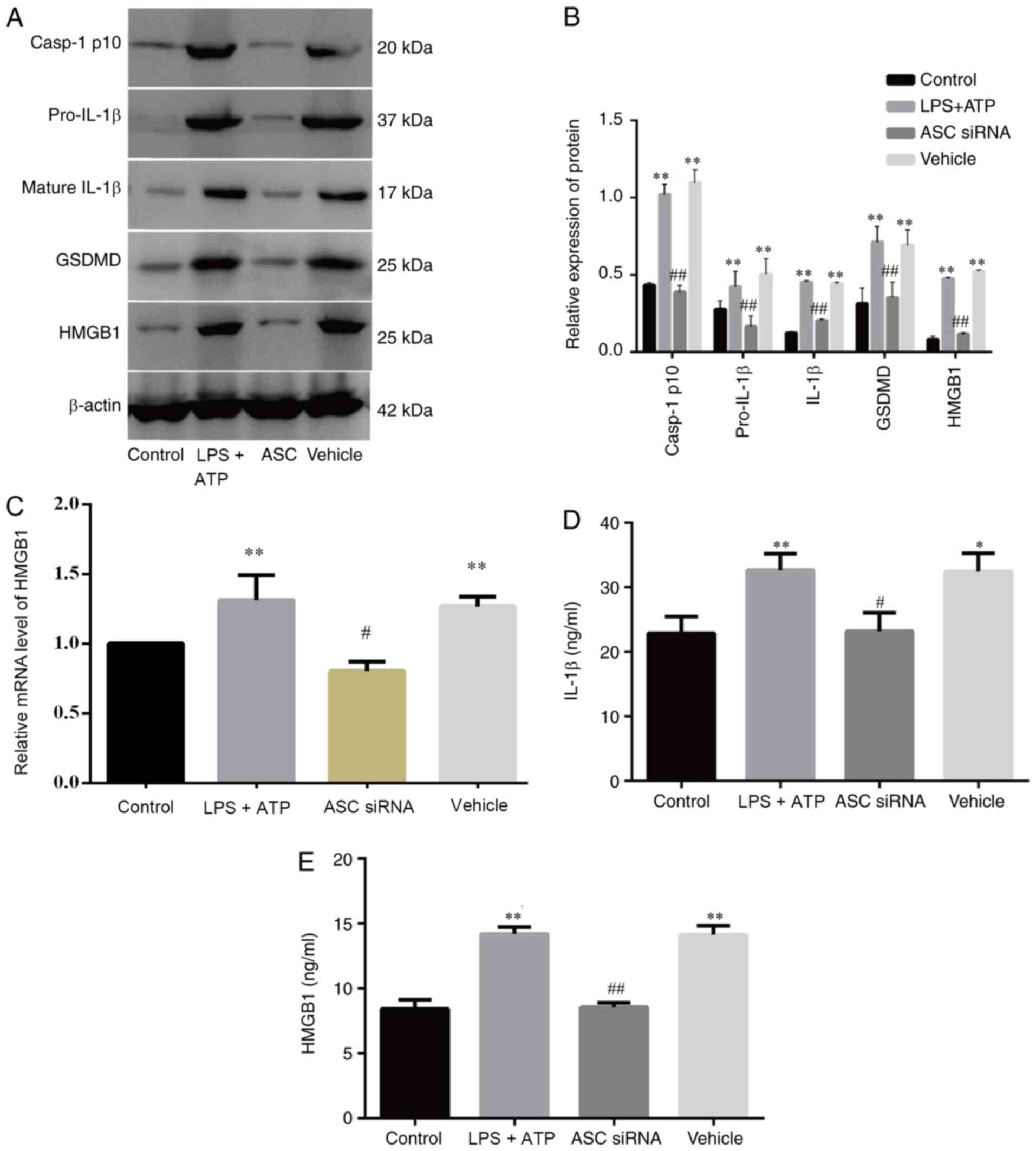
ASC knockdown mitigates FLS pyroptosis
and reduces HMGB1 secretion in vitro
ASC is an important component of inflammasomes
(33). To examine whether ASC
knockdown affects LPS + ATP-induced FLS pyroptosis and HMGB1
secretion, FLSs were exposed to an ASC-specific siRNA or a control.
The expression levels of pyroptosis-related proteins and HMGB1
(Fig. 6A and B) were significantly
higher in the LPS + ATP and vehicle groups compared with the ASC
siRNA and control groups, and the relative RNA expression of HMGB1
demonstrated the same pattern (Fig.
6C). Similar results were obtained for HMGB1 and IL-1β levels
in the supernatant of the FLSs in different groups (Fig. 6D and E). Therefore, ASC may be
important for FLS pyroptosis and HMGB1 secretion.
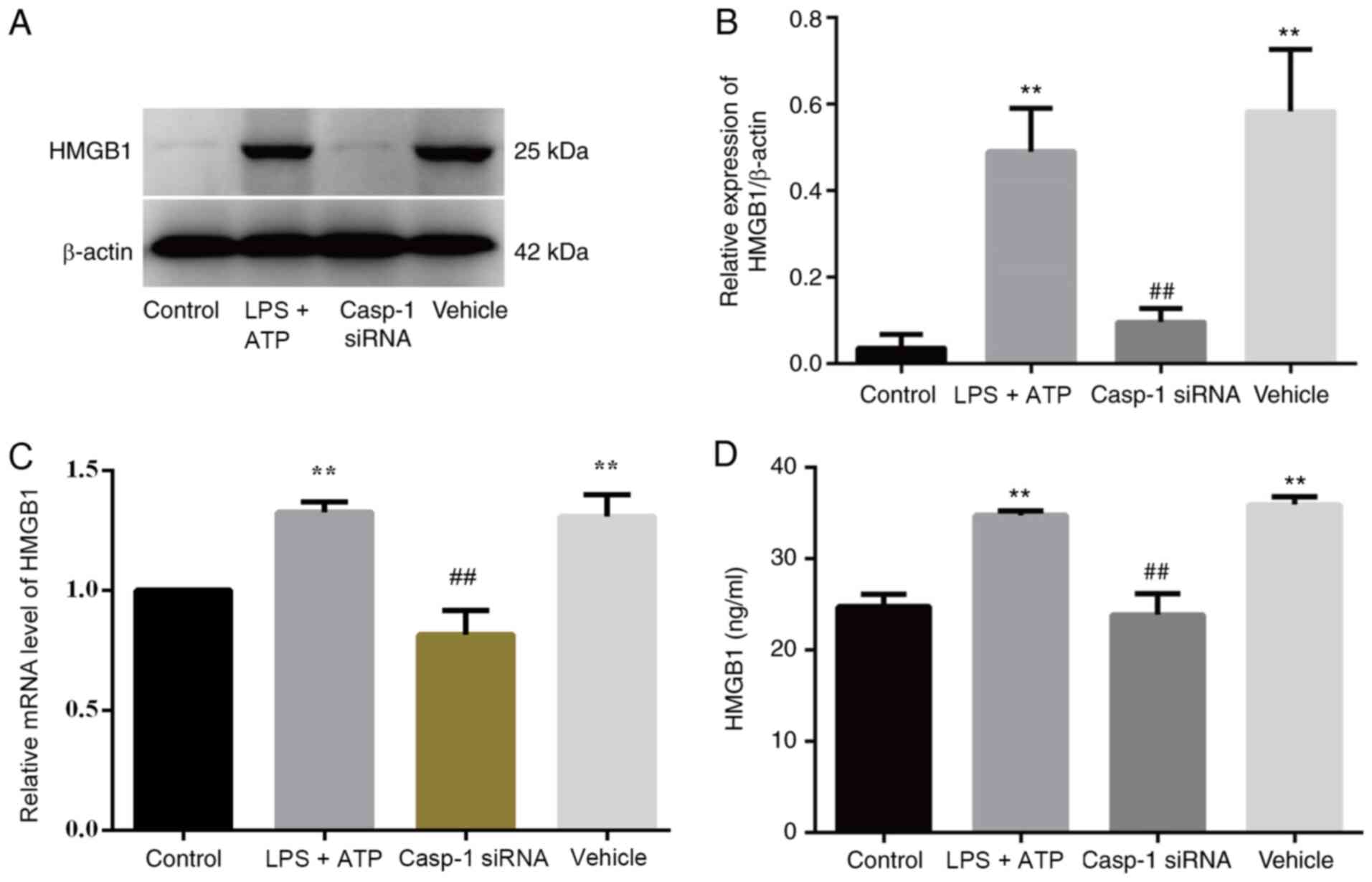
Caspase-1 knockdown decreases HMGB1
secretion in vivo
To further verify the correlation between FLS
pyroptosis and HMGB1 secretion, caspase-1-specific siRNA was used
to inhibit FLS pyroptosis, and untreated FLSs served as the control
group. HMGB1 protein expression was significantly increased in the
LPS + ATP and vehicle groups compared with the caspase-1 siRNA and
control groups (Fig. 7A and B).
Moreover, similar results were obtained for relative HMGB1 RNA
expression (Fig. 7C) and HMGB1
levels in the supernatant (Fig.
7D).
Discussion
Synovitis is common in KOA, and most patients with
KOA with obvious clinical symptoms have synovitis (34,35).
Inflammatory reactions in synovial tissues are accompanied by
congestion, edema, the accumulation of inflammatory mediators and
the secretion of various types of proteases, ultimately resulting
in abnormal cartilage metabolism (36,37).
FLSs are important factors in KOA inflammation and joint
destruction during the pathological progression of KOA, primarily
by secreting a wide range of proinflammatory mediators, such as
IL-1β, IL-18, TNF-α and MMPs (38).
HMGB1 has been reported to serve a proinflammatory
role in KOA (23,26), and HMGB1 levels in synovial fluid
are closely associated with the severity of KOA (39). On the one hand, as an important
proinflammatory factor in KOA, HMGB1 is responsible for promoting
cartilage degradation and inducing synovitis by binding to RAGE,
Toll-like receptor 4 and other receptor (40). On the other hand, HMGB1 in complex
with LPS or IL-1 enhances the production of proinflammatory
cytokines in OA FLSs (41). These
findings indicate that HMGB1 is an important factor in KOA
pathogenesis and that HMGB1 levels are highly associated with
synovitis in KOA. While the specific mechanism of HMGB1 production
in KOA is unknown, HMGB1 is found to be secreted by FLSs stimulated
with inflammatory factors in KOA (42,43).
Moreover, whether HMGB1 release is involved in FLS pyroptosis in
KOA is yet to be fully elucidated. Therefore, the present study
investigated the specific association between HMGB1 secretion and
FLS pyroptosis.
NLRP1 and NLRP3 expression levels are significantly
higher in patients with KOA (44).
The NLRP1 and NLRP3 inflammasomes are key regulators in the innate
immune system that promote the release of downstream
caspase-1-dependent proinflammatory cytokines, such as IL-1β and
IL-18 (45), suggesting that these
inflammasomes are important mediators of FLS pyroptosis.
Furthermore, inflammasomes mediate pyroptosis by activating
caspase-1 (46). In the present
study, in vivo experiments in a rat ACLT-induced KOA model
were conducted, and increased levels of HMGB1 and
pyroptosis-related proteins were found in synovial tissues, which
were significantly decreased by treatment with a caspase-1
inhibitor. Therefore, it was suggested that there was an
association between HMGB1 secretion and pyroptosis in synovial
tissue. To further demonstrate the association between HMGB1
secretion and pyroptosis, LPS + ATP was used to induce FLS
pyroptosis in vitro. Increased caspase-1 activity and
pyroptosis-related protein expression levels indicated FLS
pyroptosis. It was also identified that HMGB1 secretion by FLSs was
mediated via NLRP1 and NLRP3 inflammasomes and caspase-1 activity.
Knockdown of NLRP1, NLRP3 or caspase-1 attenuated HMGB1 secretion
by FLSs, suggesting that FLS pyroptosis may be responsible for
HMGB1 secretion by FLSs in KOA.
ASC is important for the formation of the NLRP3 and
NLRP1 inflammasomes in inflammatory diseases (47), suggesting that ASC is an important
component of pyroptosis. A previous study reported that
transfection of alveolar macrophages with ASC-specific siRNA
decreased ASC expression and mitigated pyroptosis (28). In the present study, it was
demonstrated that ASC was an important factor in FLS pyroptosis, as
transfection with ASC-specific siRNA not only inhibited
caspase-1-dependent FLS pyroptosis, but also decreased HMGB1
secretion. The aforementioned experimental results indicated that
HMGB1 production was closely associated with caspase-1-dependent
FLS pyroptosis induced by NLRP1 and NLRP3 inflammasomes and ASC
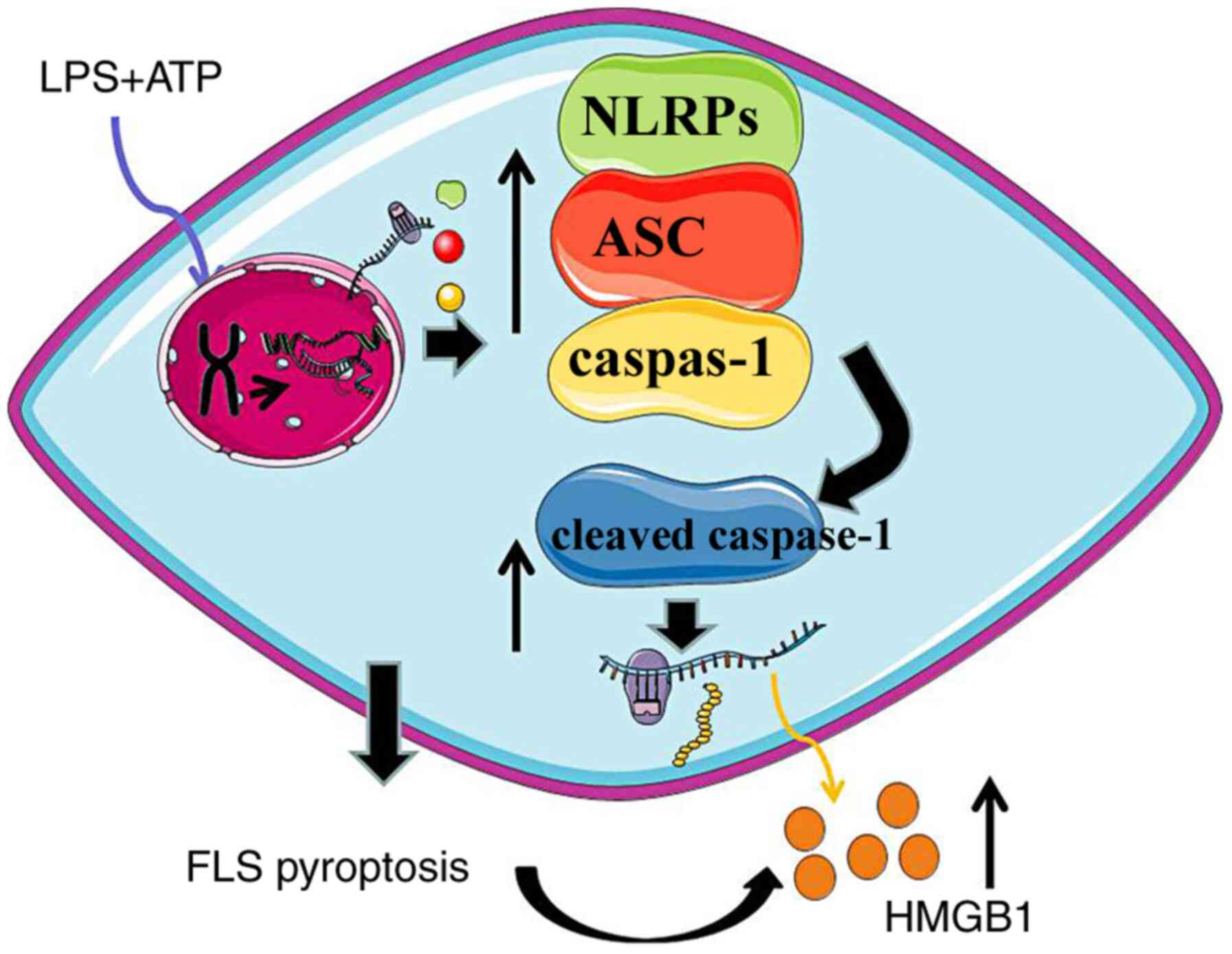
expression. Hence, it was considered that HMGB1 secretion may be
closely associated with the function of activated caspase-1
(Fig. 8). HMGB1 has been reported
to increase IL-1β production via the NLRP3 inflammasome in vascular
and liver injury (48,49), suggesting that HMGB1 may be a
pyroptosis induction factor, but it remains unknown whether HMGB1
has the same function in KOA. The present results indicated that
pyroptosis was an important factor in HMGB1 secretion, but the
specific effect of HMGB1 on pyroptosis in synovial cells requires
further investigation.
In conclusion, the present findings provide a novel
insight into the role of HMGB1 secretion in KOA pathogenesis. The
current in vivo and in vitro experiments demonstrated
that NLRP1 and NLRP3 inflammasome-mediated FLS pyroptosis enhanced
HMGB1 secretion. Thus, decreasing FLS pyroptosis by inhibiting the
activation of the NLRP1 and NLRP3 inflammasomes and caspase-1 may
be an effective strategy for treating KOA.
Acknowledgements
Not applicable.
Funding
The present study was supported by The National
Natural Science Foundation of China (grant no. 8187151765) and the
Jiangsu provincial Leading Talents Program of Traditional Chinese
Medicine of China (grant no. SLJ0207).
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
YX mainly participated in the conceptualization,
experiment operation and writing-original draft of the study. LD
mainly participated in the conceptualization and experiment
operation. SY mainly participated in experiment operation. ZH was
involved in the data analysis and methodolody. LZ participated in
purchasing the experiment materials, the resource administration
and the data analysis. WM wrote the original draft, contributed to
the validation of the study and data analysis. PWu was involved in
formal analysis of the study, data analysis and image processing.
PWa mainly participated in editing the manuscript, modifying the
draft, analysis and interpretation of data, as well as purchasing
and feeding of rats. KP participated in the conceptualization,
supervision of the study and modification of draft, and analysis
and interpretation of data. All authors read and approved the final
manuscript.
Ethics approval and consent to
participate
All animal protocols were approved by the Animal
Care and Use Committee of the Nanjing University of Chinese
Medicine.
Patient consent for publication
Not applicable.
Competing interests
All the authors declare that they have no competing
interests.
Glossary
Abbreviations
Abbreviations:
|
KOA
|
knee osteoarthritis
|
|
FLS
|
fibroblast-like synoviocytes
|
|
HMGB1
|
high mobility group box 1
|
|
NLRP
|
Nod-like receptor protein
|
|
LPS
|
lipopolysaccharide
|
|
ASC
|
apoptosis-associated speck-like
protein with a caspase-recruitment domain
|
|
GSDMD
|
gasdermin D
|
|
ACLT
|
anterior cruciate ligament
resection
|
References
|
1
|
Johnson VL and Hunter DJ: The epidemiology
of osteoarthritis. Best Pract Res Clin Rheumatol. 28:5–15. 2014.
View Article : Google Scholar
|
|
2
|
Gong Z, Liu R, Yu W, Wong TK, Guo Y and
Sun Y: Acutherapy for knee osteoarthritis relief in the elderly: A
systematic review and meta-analysis. Evid Based Complement Alternat
Med. 2019:18681072019. View Article : Google Scholar
|
|
3
|
Xia X, Wang X, Zheng Y, Jiang J and Hu J:
What role does pyroptosis play in microbial infection? J Cell
Physiol. 234:7885–7892. 2019. View Article : Google Scholar
|
|
4
|
Platnich JM and Muruve DA: NOD-like
receptors and inflammasomes: A review of their canonical and
non-canonical signaling pathways. Arch Biochem Biophys. 670:4–14.
2019. View Article : Google Scholar
|
|
5
|
Bergsbaken T, Fink SL and Cookson BT:
Pyroptosis: Host cell death and inflammation. Nat Rev Microbiol.
7:99–109. 2009. View Article : Google Scholar
|
|
6
|
Schroder K, Muruve DA and Tschopp J:
Innate immunity: Cytoplasmic DNA sensing by the AIM2 inflammasome.
Curr Biol. 19:R262–R265. 2009. View Article : Google Scholar
|
|
7
|
Duprez L, Wirawan E, Vanden Berghe T and
Vandenabeele P: Major cell death pathways at a glance. Microbes
Infect. 11:1050–1062. 2009. View Article : Google Scholar
|
|
8
|
Dong W, Zhu Q, Yang B, Qin Q, Wang Y, Xia
X, Zhu X, Liu Z, Song E and Song Y: Polychlorinated biphenyl
quinone induces caspase 1-mediated pyroptosis through induction of
pro-inflammatory HMGB1-TLR4-NLRP3-GSDMD signal axis. Chem Res
Toxicol. 32:1051–1057. 2019. View Article : Google Scholar
|
|
9
|
Zhao LR, Xing RL, Wang PM, Zhang NS, Yin
SJ, Li XC and Zhang L: NLRP1 and NLRP3 inflammasomes mediate
LPS/ATP-induced pyroptosis in knee osteoarthritis. Mol Med Rep.
17:5463–5469. 2018.
|
|
10
|
Zhang L, Zhang L, Huang Z, Xing R, Li X,
Yin S, Mao J, Zhang N, Mei W, Ding L and Wang P: Increased HIF-1α
in knee osteoarthritis aggravate synovial fibrosis via
fibroblast-like synoviocyte pyroptosis. Oxid Med Cell Longev.
2019:63265172019.
|
|
11
|
Venereau E, De Leo F, Mezzapelle R,
Careccia G, Musco G and Bianchi ME: HMGB1 as biomarker and drug
target. Pharmacol Res. 111:534–544. 2016. View Article : Google Scholar
|
|
12
|
Burzynski LC and Clarke M: Death is coming
and the clot thickens, as pyroptosis feeds the fire. Immunity.
50:1339–1341. 2019. View Article : Google Scholar
|
|
13
|
Biscetti F, Flex A, Pecorini G, Angelini
F, Arena V, Stigliano E, Gremese E, Tolusso B and Ferraccioli G:
The role of high-mobility group box protein 1 in collagen
antibody-induced arthritis is dependent on vascular endothelial
growth factor. Clin Exp Immunol. 184:62–72. 2016. View Article : Google Scholar
|
|
14
|
Chung H, Hong SJ, Choi SW, Koo JY, Kim M,
Kim HJ, Park SB and Park CG: High mobility group box 1 secretion
blockade results in the reduction of early pancreatic islet graft
loss. Biochem Biophys Res Commun. 514:1081–1086. 2019. View Article : Google Scholar
|
|
15
|
Magna M and Pisetsky DS: The role of HMGB1
in the pathogenesis of inflammatory and autoimmune diseases. Mol
Med. 20:138–146. 2014. View Article : Google Scholar
|
|
16
|
Wang FC, Pei JX, Zhu J, Zhou NJ, Liu DS,
Xiong HF, Liu XQ, Lin DJ and Xie Y: Overexpression of HMGB1 A-box
reduced lipopolysaccharide-induced intestinal inflammation via
HMGB1/TLR4 signaling in vitro. World J Gastroenterol. 21:7764–7776.
2015. View Article : Google Scholar
|
|
17
|
Yu S, Zhang H, Hei Y, Yi X, Baskys A, Liu
W and Long Q: High mobility group box-1 (HMGB1) antagonist BoxA
suppresses status epilepticus-induced neuroinflammatory responses
associated with Toll-like receptor 2/4 down-regulation in rats.
Brain Res. 1717:44–51. 2019. View Article : Google Scholar
|
|
18
|
Tian X, Liu C, Shu Z and Chen G: Review:
Therapeutic targeting of HMGB1 in stroke. Curr Drug Deliv.
14:785–790. 2017. View Article : Google Scholar
|
|
19
|
Paudel YN, Angelopoulou E, Piperi C,
Balasubramaniam V, Othman I and Shaikh MF: Enlightening the role of
high mobility group box 1 (HMGB1) in inflammation: Updates on
receptor signalling. Eur J Pharmacol. 858:1724872019. View Article : Google Scholar
|
|
20
|
Vijayakumar EC, Bhatt LK and Prabhavalkar
KS: High mobility group box-1 (HMGB1): A potential target in
therapeutics. Curr Drug Targets. 20:1474–1485. 2019. View Article : Google Scholar
|
|
21
|
Lin SS, Yuan LJ, Niu CC, Tu YK, Yang CY
and Ueng SWN: Hyperbaric oxygen inhibits the HMGB1/RAGE signaling
pathway by upregulating Mir-107 expression in human osteoarthritic
chondrocytes. Osteoarthritis Cartilage. 27:1372–1381. 2019.
View Article : Google Scholar
|
|
22
|
Wagner G, Lehmann C, Bode C, Miosge N and
Schubert A: High mobility group box 1 protein in osteoarthritic
knee tissue and chondrogenic progenitor cells: An ex vivo and in
vitro study. Cartilage. Mar 26–2019.(Online ahead of print).
View Article : Google Scholar
|
|
23
|
Taniguchi N, Kawakami Y, Maruyama I and
Lotz M: HMGB proteins and arthritis. Hum Cell. 31:1–9. 2018.
View Article : Google Scholar
|
|
24
|
Alsousi AA and Igwe OJ: Redox-active trace
metal-induced release of high mobility group box 1(HMGB1) and
inflammatory cytokines in fibroblast-like synovial cells is
Toll-like receptor 4 (TLR4) dependent. Biochim Biophys Acta Mol
Basis Dis. 1864:3847–3858. 2018. View Article : Google Scholar
|
|
25
|
Nefla M, Holzinger D, Berenbaum F and
Jacques C: The danger from within: Alarmins in arthritis. Nat Rev
Rheumatol. 12:669–683. 2016. View Article : Google Scholar
|
|
26
|
Ke X, Jin G, Yang Y, Cao X, Fang R, Feng X
and Lei B: Synovial fluid HMGB-1 levels are associated with
osteoarthritis severity. Clin Lab. 61:809–818. 2015. View Article : Google Scholar
|
|
27
|
Kim HM and Kim YM: HMGB1: LPS delivery
vehicle for caspase-11-mediated pyroptosis. Immunity. 49:582–584.
2018. View Article : Google Scholar
|
|
28
|
Hou L, Yang Z, Wang Z, Zhang X, Zhao Y,
Yang H, Zheng B, Tian W, Wang S, He Z and Wang X:
NLRP3/ASC-mediated alveolar macrophage pyroptosis enhances HMGB1
secretion in acute lung injury induced by cardiopulmonary bypass.
Lab Invest. 98:1052–1064. 2018. View Article : Google Scholar
|
|
29
|
National Research Council (US) Institute
for Laboratory Animal Research, . Guide for the Care and Use of
Laboratory Animals. Washington (DC): National Academies Press (US);
1996, simplehttps://www.ncbi.nlm.nih.gov/books/NBK232589/doi:
10.17226/5140.
|
|
30
|
Xing R, Wang P, Zhao L, Xu B, Zhang N and
Li X: Mechanism of TRPA1 and TRPV4 participating in mechanical
hyperalgesia of rat experimental knee osteoarthritis. Arch
Rheumatol. 32:96–104. 2017. View Article : Google Scholar
|
|
31
|
Miao EA, Leaf IA, Treuting PM, Mao DP,
Dors M, Sarkar A, Warren SE, Wewers MD and Aderem A:
Caspase-1-induced pyroptosis is an innate immune effector mechanism
against intracellular bacteria. Nat Immunol. 11:1136–1142. 2010.
View Article : Google Scholar
|
|
32
|
Schmittgen TD and Livak KJ: Analyzing
real-time PCR data by the comparative CT method. Nat Protoc.
3:1101–1108. 2008. View Article : Google Scholar
|
|
33
|
Martel-Pelletier J, Raynauld JP, Mineau F,
Abram F, Paiement P, Delorme P and Pelletier JP: Levels of serum
biomarkers from a two-year multicentre trial are associated with
treatment response on knee osteoarthritis cartilage loss as
assessed by magnetic resonance imaging: An exploratory study.
Arthritis Res Ther. 19:1692017. View Article : Google Scholar
|
|
34
|
Samuelsson K, Magnussen RA, Alentorn-Geli
E, Krupic F, Spindler KP, Johansson C, Forssblad M and Karlsson J:
Equivalent knee injury and osteoarthritis outcome scores 12 and 24
months after anterior cruciate ligament reconstruction: Results
from the swedish national knee ligament register. Am J Sports Med.
45:2085–2091. 2017. View Article : Google Scholar
|
|
35
|
Steinhaus ME, Christ AB and Cross MB:
Total knee arthroplasty for knee osteoarthritis: Support for a
foregone conclusion? HSS J. 13:207–210. 2017. View Article : Google Scholar
|
|
36
|
Kong R, Gao J, Si Y and Zhao D:
Combination of circulating miR-19b-3p, miR-122-5p and miR-486-5p
expressions correlates with risk and disease severity of knee
osteoarthritis. Am J Transl Res. 9:2852–2864. 2017.
|
|
37
|
Zhang L, Xing R, Huang Z, Zhang N, Zhang
L, Li X and Wang P: Inhibition of synovial macrophage pyroptosis
alleviates synovitis and fibrosis in knee osteoarthritis. Mediators
Inflamm. 2019:21659182019. View Article : Google Scholar
|
|
38
|
Zhang C, Yu W, Huang C, Ding Q, Liang C,
Wang L, Hou Z and Zhang Z: Chrysin protects human osteoarthritis
chondrocytes by inhibiting inflammatory mediator expression via
HMGB1 suppression. Mol Med Rep. 19:1222–1229. 2019.
|
|
39
|
Shu Z, Miao X, Tang T, Zhan P, Zeng L and
Jiang Y: The GSK-3β/β-catenin signaling pathway is involved in
HMGB1-induced chondrocyte apoptosis and cartilage matrix
degradation. Int J Mol Med. 45:769–778. 2020.
|
|
40
|
Wähämaa H, Schierbeck H, Hreggvidsdottir
HS, Palmblad K, Aveberger AC, Andersson U and Harris HE: High
mobility group box protein 1 in complex with lipopolysaccharide or
IL-1 promotes an increased inflammatory phenotype in synovial
fibroblasts. Arthritis Res Ther. 13:R1362011. View Article : Google Scholar
|
|
41
|
Garcia-Arnandis I, Guillen MI, Gomar F,
Pelletier JP, Martel-Pelletier J and Alcaraz MJ: High mobility
group box 1 potentiates the pro-inflammatory effects of
interleukin-1β in osteoarthritic synoviocytes. Arthritis Res Ther.
12:R1652010. View
Article : Google Scholar
|
|
42
|
Loeser RF, Goldring SR, Scanzello CR and
Goldring MB: Osteoarthritis: A disease of the joint as an organ.
Arthritis Rheum. 64:1697–1707. 2012. View Article : Google Scholar
|
|
43
|
Terada C, Yoshida A, Nasu Y, Mori S,
Tomono Y, Tanaka M, Takahashi HK, Nishibori M, Ozaki T and Nishida
K: Gene expression and localization of high-mobility group box
chromosomal protein-1 (HMGB-1)in human osteoarthritic cartilage.
Acta Med Okayama. 65:369–377. 2011.
|
|
44
|
Fan C, Zhao X, Guo X, Cao X and Cai J:
P2X4 promotes interleukin-1β production in osteoarthritis via
NLRP1. Mol Med Rep. 9:340–344. 2014. View Article : Google Scholar
|
|
45
|
Harapas CR, Steiner A, Davidson S and
Masters SL: An update on autoinflammatory diseases:
Inflammasomopathies. Curr Rheumatol Rep. 20:402018. View Article : Google Scholar
|
|
46
|
Kesavardhana S and Kanneganti TD:
Mechanisms governing inflammasome activation, assembly and
pyroptosis induction. Int Immunol. 29:201–210. 2017. View Article : Google Scholar
|
|
47
|
Zhang B, Zhang Y, Xu T, Yin Y, Huang R,
Wang Y, Zhang J, Huang D and Li W: Chronic dexamethasone treatment
results in hippocampal neurons injury due to activate NLRP1
inflammasome in vitro. Int Immunopharmacol. 49:222–230. 2017.
View Article : Google Scholar
|
|
48
|
Kim EJ, Park SY, Baek SE, Jang MA, Lee WS,
Bae SS, Kim K and Kim CD: HMGB1 Increases IL-1β production in
vascular smooth muscle cells via NLRP3 inflammasome. Front Physiol.
9:3132018. View Article : Google Scholar
|
|
49
|
Geng Y, Ma Q, Liu YN, Peng N, Yuan FF, Li
XG, Li M, Wu YS, Li BL, Song WB, et al: Heatstroke induces liver
injury via IL-1β and HMGB1-induced pyroptosis. J Hepatol.
63:622–633. 2015. View Article : Google Scholar
|