Introduction
Intermedin (IMD), also termed adrenomedullin-2, is a
member of the calcitonin/calcitonin gene-related peptide family and
is widely expressed in the heart, blood vessels, brain,
hypothalamus, kidney, lung, spleen, thymus, ovary and adipose
tissue (1). The plasma levels of
IMD are relatively low, between 100 and 200 pg/ml (2). Following proteolytic cleavage,
pre-proIMD generates IMD1-53, IMD1-40 and
IMD1-47, the major active fragments that act through the
calcitonin-like receptor/receptor activity-modifying protein
complexes to induce multiple biological effects, such as increased
prolactin release, antidiuretic and natriuretic effects, and
reduced food intake (3).
Accumulating evidence has indicated extensive functions of IMD in
maintaining cardiovascular homeostasis, such as accelerating
angiogenesis, anti-apoptotic and fibroblast activity, and
increasing cardiac contractility and perfusion (4–6). The
plasma levels of IMD and the endogenous IMD in cardiomyocytes have
been reported to be significantly increased in response to heart
failure and acute cardiac infarction stimuli, and IMD
supplementation inhibits cardiomyocyte injury induced by heart
failure and cardiac infarction (4,7,8). These
results also suggested that IMD may be a potential endogenous
protector of the heart. Although these studies have confirmed the
cardioprotective effects of IMD, the underlying protective
mechanisms are still unclear. Teng et al (3) and Zhang et al (9) have reported that IMD attenuates
tunicamycin and dithiothreitol-induced myocardial injury in rats by
inhibiting endoplasmic reticulum (ER) stress. Tunicamycin and
dithiothreitol induce ER stress via inhibition of protein
glycosylation and disulfide bond formation, respectively. Similar
to tunicamycin or dithiothreitol, thapsigargin is also an inducer
of ER stress. Thapsigargin, originally isolated from the plant
Thapsia garganica, is a specific and potent inhibitor for
sarco/endoplasmic reticulum calcium ATPase (SERCA) (10). By inhibiting SERCAs, thapsigargin
interferes with the ER lumen Ca2+ flux that subsequently
leads to ER stress (10,11). SERCA has been reported to exert
important effects in the heart, and a number of heart diseases are
associated with SERCA dysfunction, such as cardiac hypertrophy and
heart failure (12,13). Thapsigargin induces ER
stress-related apoptosis by inhibiting SERCA activity to partially
simulate the pathophysiological processes of various cardiovascular
diseases (14).
The SERCA that is present in all organisms is a 110
kDa transmembrane protein encoded by three homologous genes
(SERCA1, SERCA2 and SERCA3), with a dominant expression of SERCA2a
in cardiomyocytes (15). SERCA2a is
crucial for regulating Ca2+ homeostasis by transporting
cytosolic Ca2+ into the sarcoplasmic reticulum (16,17).
Phospholamban (PLB) located in the cardiac sarcoplasmic reticulum
and, as an endogenous SERCA inhibitor, compromises SERCA affinity
for Ca2+ (18). PLB
phosphorylation is the primary regulator of SERCA activity in
cardiomyocytes (19,20).
To date, there have been no published reports on the
effects of IMD on cardiomyocyte injury induced by thapsigargin to
the best of our knowledge. Therefore, the preset study focused on
the role of IMD in thapsigargin-induced cardiomyocyte apoptosis and
examined SERCA activity during thapsigargin treatment to explore
whether IMD restored ER stress in cardiomyocytes via a
SERCA-dependent mechanism. Protein kinase A (PKA) has also been
reported to increase SERCA activity (21); to further explore the underlying
mechanisms through which IMD regulates SERCA activity and ER
stress, the present study examined the involvement of the PKA
pathway in the IMD-mediated protective effects in
cardiomyocytes.
Materials and methods
Cell culture and treatment
The experimental protocols were approved by the
Ethical Committee of Shanxi Medical University (Taiyuan, China) and
complied with internationally accepted principles of laboratory
animal care and use (22). Animals
were housed at 23°C with 50% humidity, 12-h light/dark cycles, and
free access to food and water. Briefly, 16 neonatal SD rats (male
and female; age, 1–3 days; Laboratory Animal Center of Shanxi
Medical University) were anesthetized by pentobarbital sodium (100
mg/kg) and decapitated for cardiac tissue harvesting. Left
ventricular tissues were digested with collagenase II
(Sigma-Aldrich; Merck KGaA) in Hank's balanced salt solution
(Ca2+- and Mg2+-free) for 1 h at 37°C, as
described previously (23). After
centrifugation at 500 × g for 10 min at 4°C, the supernatants were
discarded, and the cells were resuspended; ~90 min later,
non-myocytes were attached to the dishes. The viable non-attached
cells were collected and plated on 60-mm culture dishes at a
density of 4×106 cells per dish and cultured in DMEM
supplemented with 1% penicillin-streptomycin and 20% FBS (all
purchased from Sigma-Aldrich; Merck KGaA) supplemented with 0.1 mM
bromodeoxyuridine (Sigma-Aldrich; Merck KGaA). Using this strategy,
a myocyte population was obtained, and all experiments were
performed within 3 days after incubation. To evaluate the effects
of IMD on thapsigargin-induced ER stress, cardiomyocytes were
divided into 6 groups: i) Control; ii) IMD, cells incubated with
100 nM IMD for 24 h at 37°C (100 nM; Phoenix Pharmaceuticals, Inc.)
under normoxic conditions (5% CO2; iii) thapsigargin,
cells incubated with 3 µM thapsigargin (Sigma-Aldrich; Merck KGaA),
a classic ER stress inducer, for 24 h; iv-vi) thapsigargin +1, 10
and 100 nM IMD, cells incubated with the indicated dose of IMD
(thapsigargin was administered 30 min after the addition of IMD).
To determine the role of PKA in IMD-mediated cardioprotection,
cardiomyocytes were pretreated with 10 µM protein kinase A
antagonist H-89 (Sigma-Aldrich; Merck KGaA) for 24 h at 37°C.
Western blot analysis
Cardiomyocytes were lysed in the presence of 10 mM
phenylmethylsulfonyl fluoride (Sigma-Aldrich; Merck KGaA), and the
protein concentration of the lysates was determined using a BCA
Protein Assay kit (Beyotime Institute of Biotechnology). Proteins
from each sample (~30 µg) were separated by 12% sodium dodecyl
sulfate-polyacrylamide gel electrophoresis, and then transferred to
polyvinylidene fluoride membranes. Following incubation with 5%
bovine serum albumin for 2 h at room temperature, the membranes
were incubated overnight at 4°C with primary antibodies against 78
kDa glucose-regulated protein (GRP78; 1:200; cat. no. sc-13539),
C/EBP-homologous protein (CHOP; 1:200; cat. no. sc-7351),
caspase-12 (1:200; cat. no. sc-21747), GAPDH (1:500; cat. no.
sc-32233; Santa Cruz Biotechnology, Inc.), SERCA2 (1:500; cat. no.
S1439), phosphorylated (p-)PLB (1:200; cat. no. SAB1305590), total
(t-)PLB (1:200; cat. no. HPA026900; Sigma-Aldrich; Merck KGaA).
Subsequently, the blots were incubated with the appropriate
secondary antibodies (cat. no. sc-2004; Santa Cruz Biotechnology,
Inc.) for 3 h at 37°C. The membranes were developed with an
enhanced chemiluminescence detection system (Amersham; Cytiva).
Signal intensities were analyzed using a Gel Imaging System
(Bio-Rad Laboratories, Inc.). All assays were performed in
triplicate. The relative protein expression levels were determined
by normalization to GAPDH.
Measurement of intracellular
Ca2+ concentration
Intracellular Ca2+ concentration
([Ca2+]i) was measured using the
Ca2+-specific fluorescent probe fluo-3/AM (cat. no.
50013; Biotium, Inc.). Cardiomyocytes were loaded with 5 µM
fluo-3/AM for 30 min at 37°C. The mean fluorescence intensity of
the cells was monitored using an Olympus FV1000 laser scanning
confocal microscope (Olympus Corporation). Fluo-3/AM was excited at
488 nm, and the emission intensity was measured at 528 nm. All
experiments were repeated at least four times.
Measurement of SERCA activity
The sarcoplasmic reticulum of cardiomyocytes was
obtained using an ER isolation kit (cat. no. ER0100; Sigma-Aldrich;
Merck KGaA) according to the manufacturer's instructions. SERCA
activity was measured using a Ca2+-ATPase Assay kit
(Nanjing Jiancheng Bioengineering Institute) according to the
manufacturer's instructions, normalized to protein concentration
and expressed as nanomoles of phosphorus ions per milligram of
protein per minute (nmol Pi/mg prot/min).
Flow cytometry analysis
Apoptotic cells were detected using an
Annexin-V-FITC kit (BD Biosciences) with propidium iodide (PI)
staining and flow cytometric analysis according to the
manufacturer's instructions. The double-negative (Annexin
V-negative/PI-negative) cells were defined as viable, the
single-positive populations were considered to be early apoptotic
(Annexin V-positive/PI-negative) or necrotic (Annexin
V-negative/PI-positive) cells, and double-positive (Annexin
V-positive/PI-positive) cells were considered to be in a late stage
of apoptosis. Apoptotic cells were analyzed using a FACScan flow
cytometer (BD Biosciences) and CellQuest Pro software (version 5.1;
BD Biosciences). Apoptosis was calculated as the total of early and
late apoptotic cells.
Statistical analysis
Data are expressed as the mean ± standard deviation.
Statistical analysis was performed using GraphPad Prism 6.0
(GraphPad Software, Inc.). All data on dose response and time
series were analyzed with a one-way ANOVA followed by a Tukey's
post hoc test. Additional data were analyzed with student's
t-test for two group comparisons. P<0.05 was considered
to indicate a statistically significant difference.
Results
Thapsigargin induces cardiomyocyte
apoptosis in a dose- and time-dependent manner
Neonatal rat cardiomyocytes were incubated with 1.5,
3 or 6 µM thapsigargin for 24 h to determine the working
concentration of thapsigargin. Cell viability was determined using
an MTT assay. As demonstrated in Fig.
1A, treatment with increasing concentrations of thapsigargin
led to a dose-dependent loss of cardiomyocyte viability compared
with the control group. Following exposure to 3 µM thapsigargin,
69.33±4.63% of the cardiomyocytes were viable relative to the
control group (P<0.01). However, with 6 µM thapsigargin, the
viability of cells was significantly reduced to 44.52±6.88%
(Fig. 1A). Apoptosis was analyzed
by flow cytometry using the Annexin V/PI double staining assay. The
percentages of apoptotic cells were 9.8±7.55, 26.47.0±10.31 and
50.87±13.51% in the 1.5, 3 and 6 µM thapsigargin groups,
respectively (Fig. 1B and C).
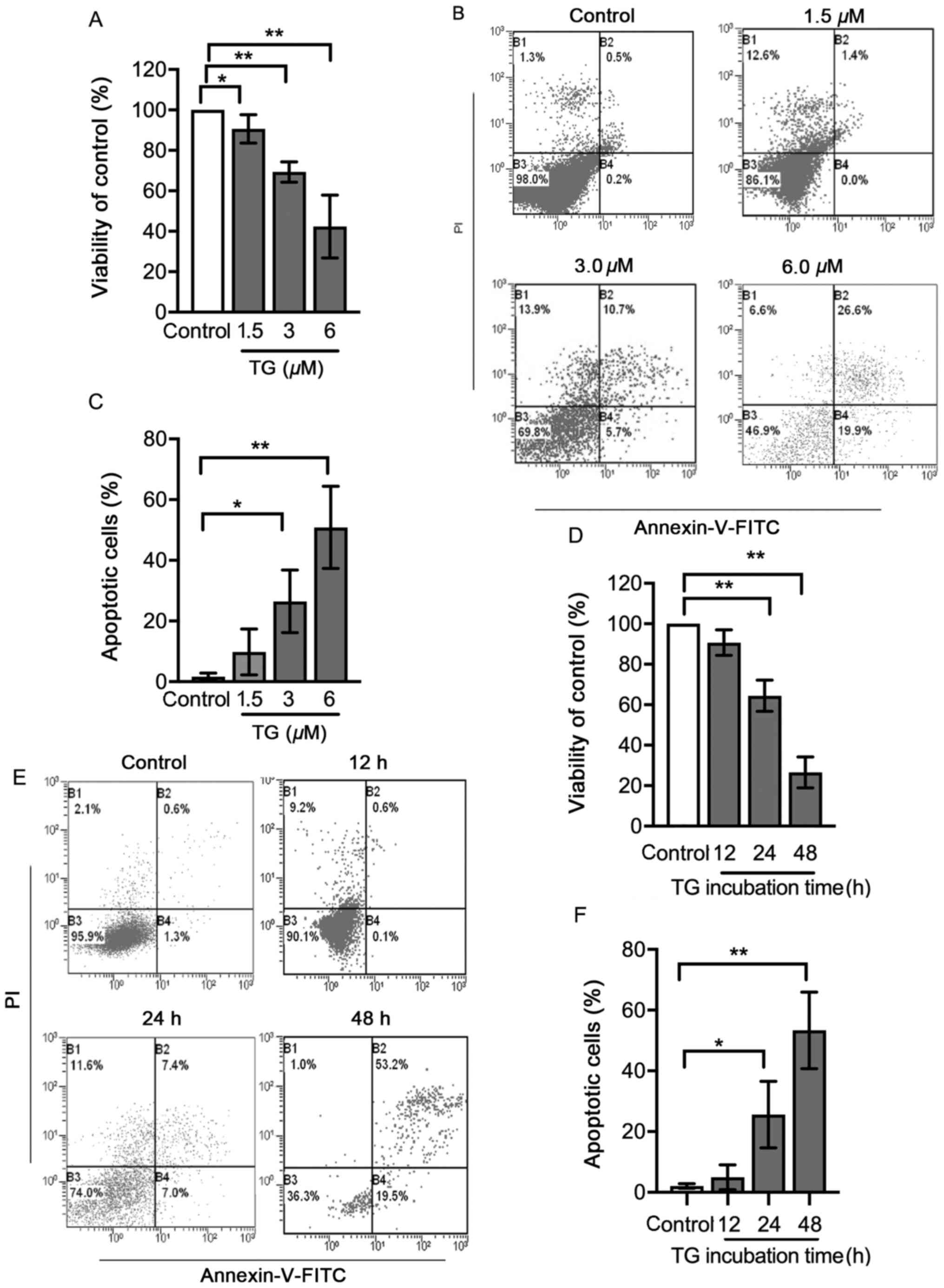 | Figure 1.Viability and apoptosis of cells
treated with TG. Cultured neonatal rat cardiomyocytes were left
untreated or treated with 1.5, 3 or 6 µM TG for 24 h. Following
this, (A) cell viability was determined using an MTT assay, (B)
apoptosis was determined using flow cytometry and (C) the rates of
apoptotic cells are presented in bar graphs. Cultured neonatal rat
cardiomyocytes were left untreated or treated with 3 µM TG for 12,
24 or 48 h. Following this, (D) cell viability was determined using
an MTT assay. (E) Apoptosis was assessed using flow cytometry
analysis and (F) the rates of apoptotic cells are presented in bar
graphs. In the flow cytometry plots, the horizontal axis represents
the Annexin V intensity, and the vertical axis represents PI
staining. Lower left quadrant, living cells; upper left quadrant,
necrotic cells; right quadrants, apoptotic cells. All data on dose
response and time series were analyzed with a one-way ANOVA
followed by a Tukey's test. Data are presented as the mean ± SD.
n=3. *P<0.05 and **P<0.01. TG, thapsigargin; PI, propidium
iodide. |
In order to further study the effects of
thapsigargin, cardiomyocytes were treated with 3 µM thapsigargin
for 12, 24 or 48 h. After exposure to 3 µM thapsigargin for the
indicated times, cell viability was significantly decreased in the
24 and 48 h groups compared with that of the control group
(Fig. 1D). The apoptotic rates in
the 24 and 48 h groups, but not the 12 h group, were significantly
higher compared with those in the control group (Fig. 1E and F). Thapsigargin induced
apoptosis in cardiomyocytes in a dose- and time-dependent manner;
thus, 3 µM thapsigargin treatment for 24 h was used in subsequent
experiments.
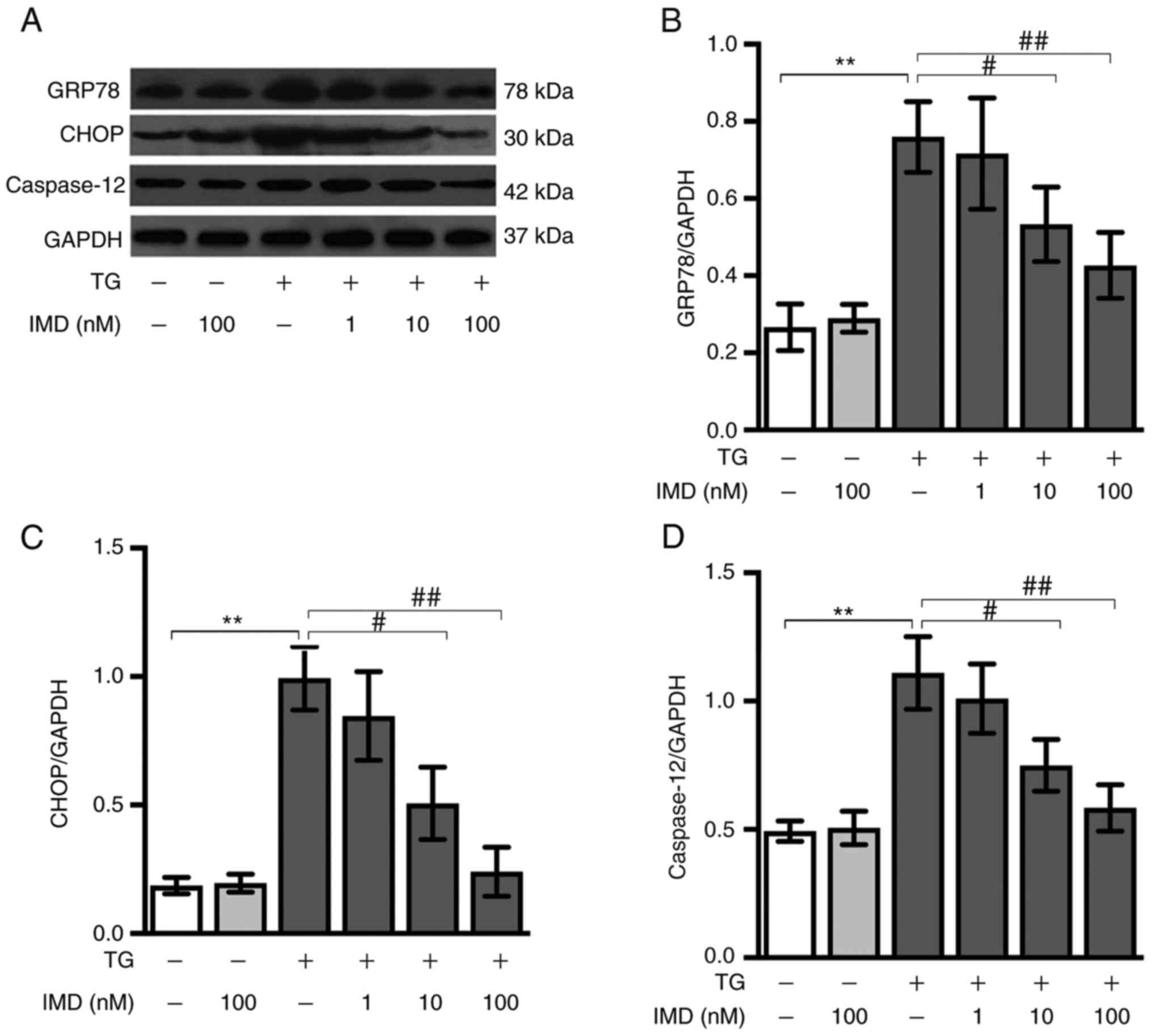
IMD inhibits thapsigargin-induced ER
stress in cardiomyocytes
Cardiomyocytes were incubated with 3 µM thapsigargin
in the presence or absence of IMD for 24 h. CHOP, GRP78 and
caspase-12 are molecular markers specific for ER stress (24). Compared with the control group, the
thapsigargin group exhibited significantly upregulated protein
expression levels of GRP78, CHOP and caspase-12, indicating that
thapsigargin treatment induced ER stress. IMD (10 and 100 nM)
reduced the thapsigargin-mediated upregulation of ER stress in a
dose-dependent manner. Pretreatment with 100 nM IMD resulted in a
strong protective effect against thapsigargin-induced injury
(Fig. 2A-D). Collectively, these
results demonstrated that IMD pretreatment inhibited the
thapsigargin-induced ER stress in a dose-dependent manner.
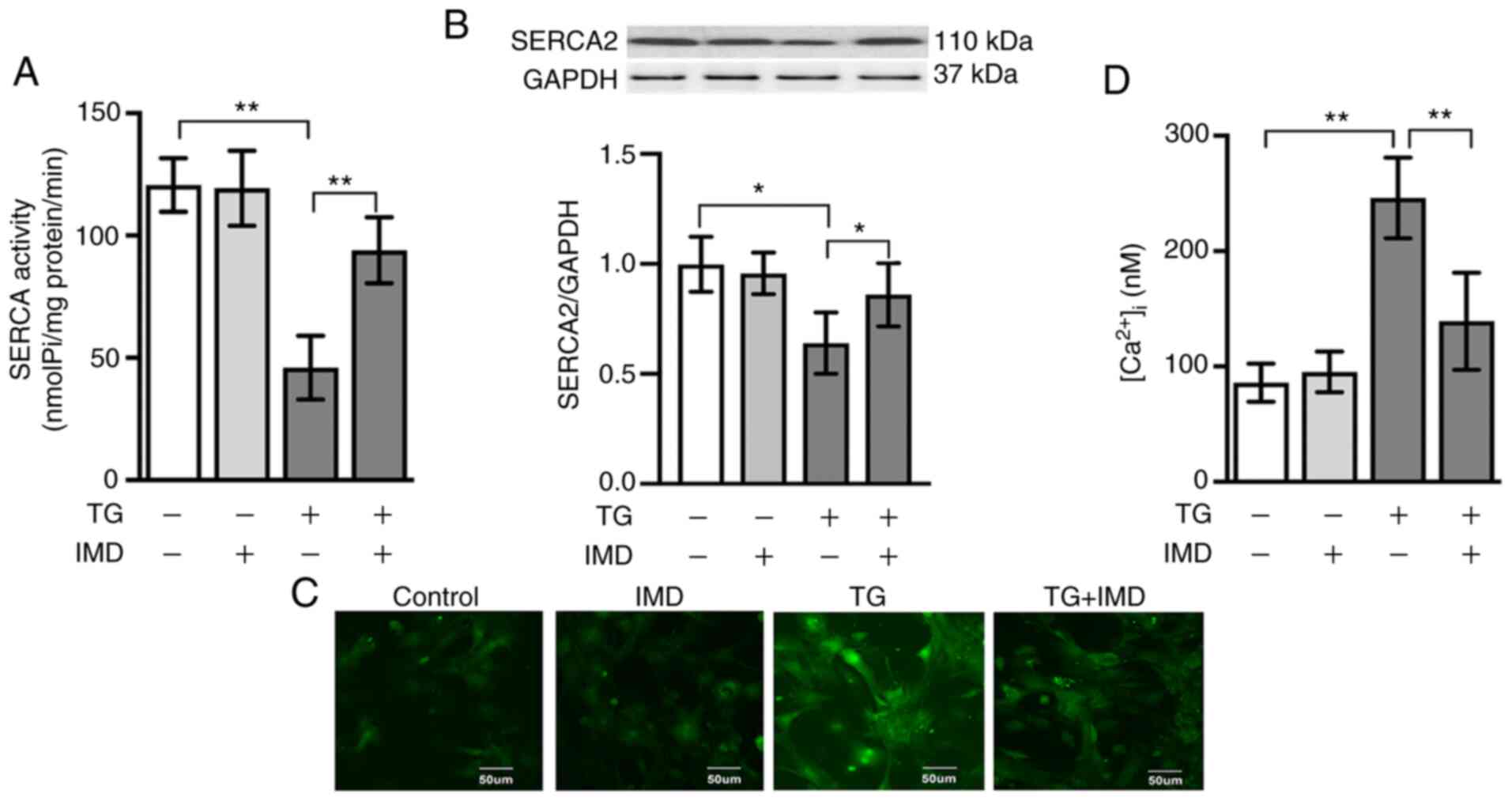
IMD attenuates SERCA suppression and
[Ca2+]i overload induced by thapsigargin in
cardiomyocytes
Thapsigargin is a highly selective inhibitor of
SERCA. Therefore, the present study examined SERCA activity in
cardiomyocytes treated with thapsigargin in the presence or absence
of IMD (100 nM) to further investigate the possible relationship
between IMD and SERCA. As demonstrated in Fig. 3A and B, thapsigargin treatment
significantly reduced the activity and protein expression levels of
SERCA2, which were reversed by IMD treatment.
Since SERCA serves a crucial role in Ca2+
homeostasis, subsequent experiments were performed to determine
whether IMD decreased cytosolic Ca2+ overload in
cardiomyocytes using the fluorescent indicator fluo-3AM. As
presented in Fig. 3C and D, the
intensity of [Ca2+]i fluorescence was
significantly elevated in the thapsigargin group compared with that
in the vehicle group. This increase in
[Ca2+]i fluorescence was significantly
reduced by IMD pretreatment (Fig. 3C
and D).
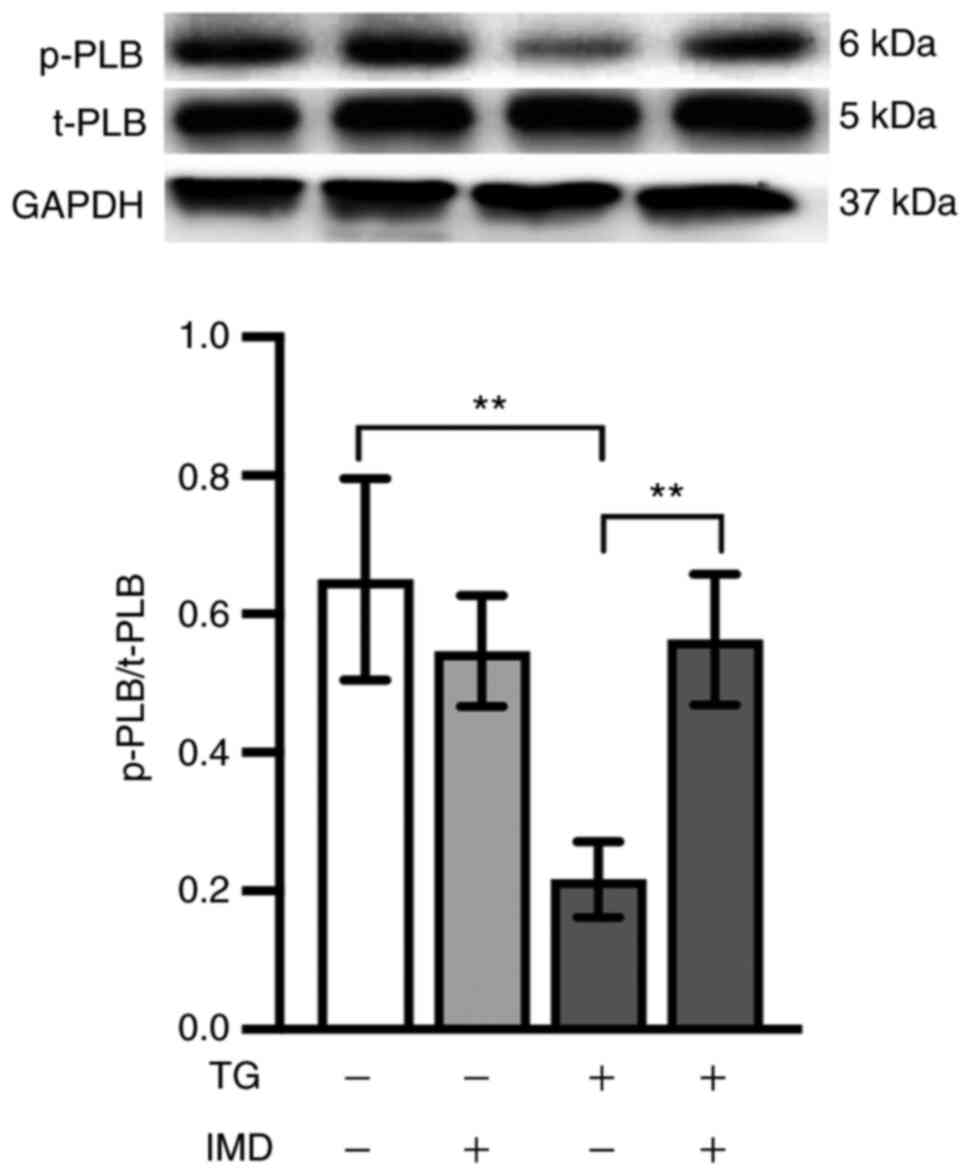
IMD improves SERCA function by
regulating PLB phosphorylation
PLB is a key regulatory protein of SERCA (25). Unphosphorylated PLB binds SERCA and
inhibits its activity, which is abolished upon PLB phosphorylation
(25). Therefore, elucidating the
changes in the SERCA regulatory protein PLB may help understand the
protective mechanisms of IMD. Thus, t-PLB and p-PLB protein levels
were analyzed in the present study by western blotting. As
demonstrated in Fig. 4, the levels
of p-PLB, which is the active form of the SERCA regulatory protein,
were reduced in the thapsigargin group compared with those in the
vehicle group. This reduction was reversed by IMD pretreatment
(Fig. 4).
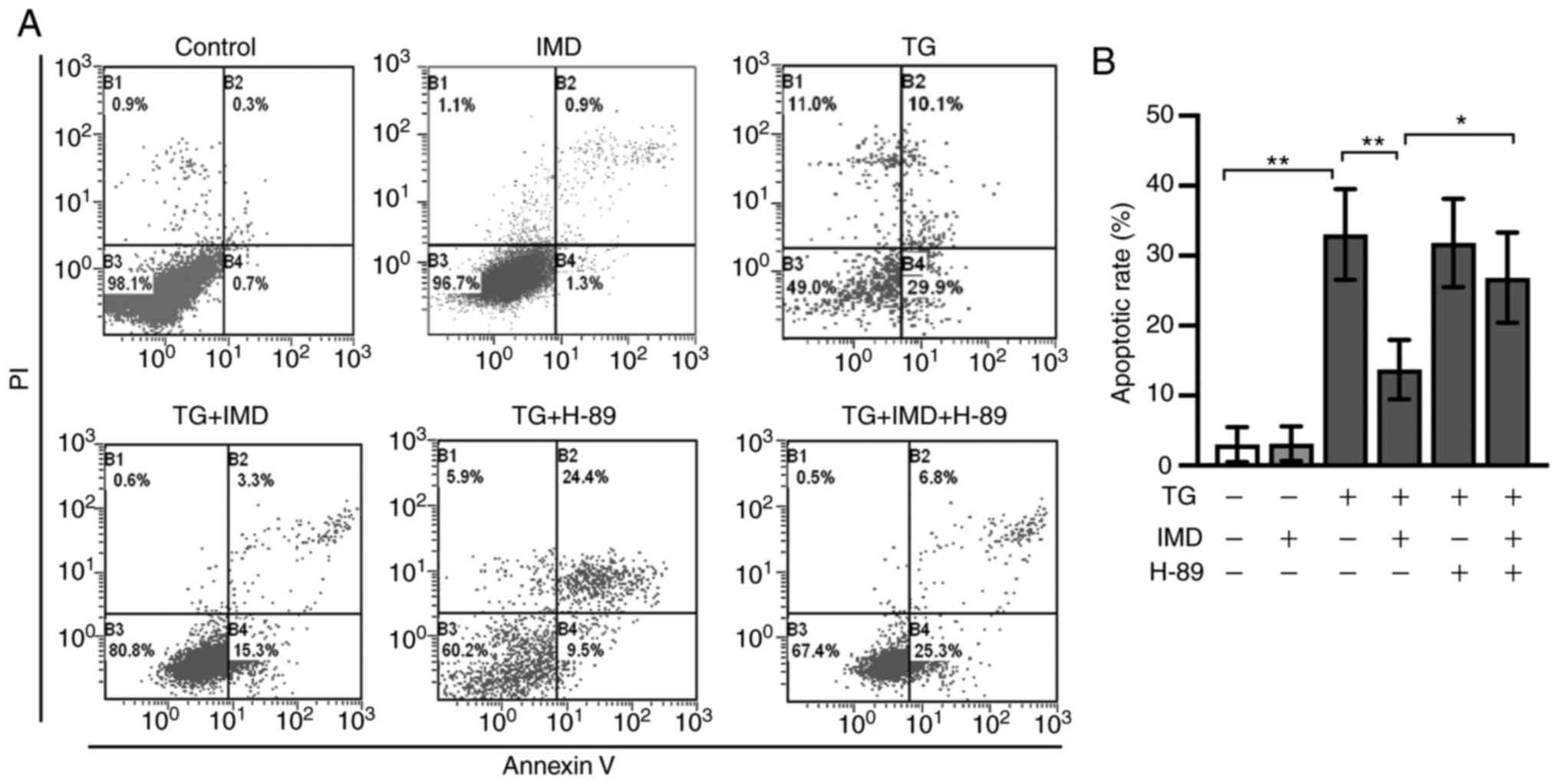
PKA contributes to SERCA function and
ER stress regulated by IMD
As PKA regulates PLB phosphorylation and SERCA
activity (21,26), the present study further examined
whether the PKA signaling pathway was involved in IMD-mediated
cardioprotection, PLB phosphorylation and SERCA function. As
presented in Fig. 5A-D,
co-treatment with the PKA inhibitor H-89 (10 µM) inhibited the
effects of IMD on PLB phosphorylation and SERCA activity in
cardiomyocytes treated with thapsigargin. These results suggested
that IMD-induced PLB phosphorylation and restoration of SERCA
activity were mediated in part by the PKA pathway.
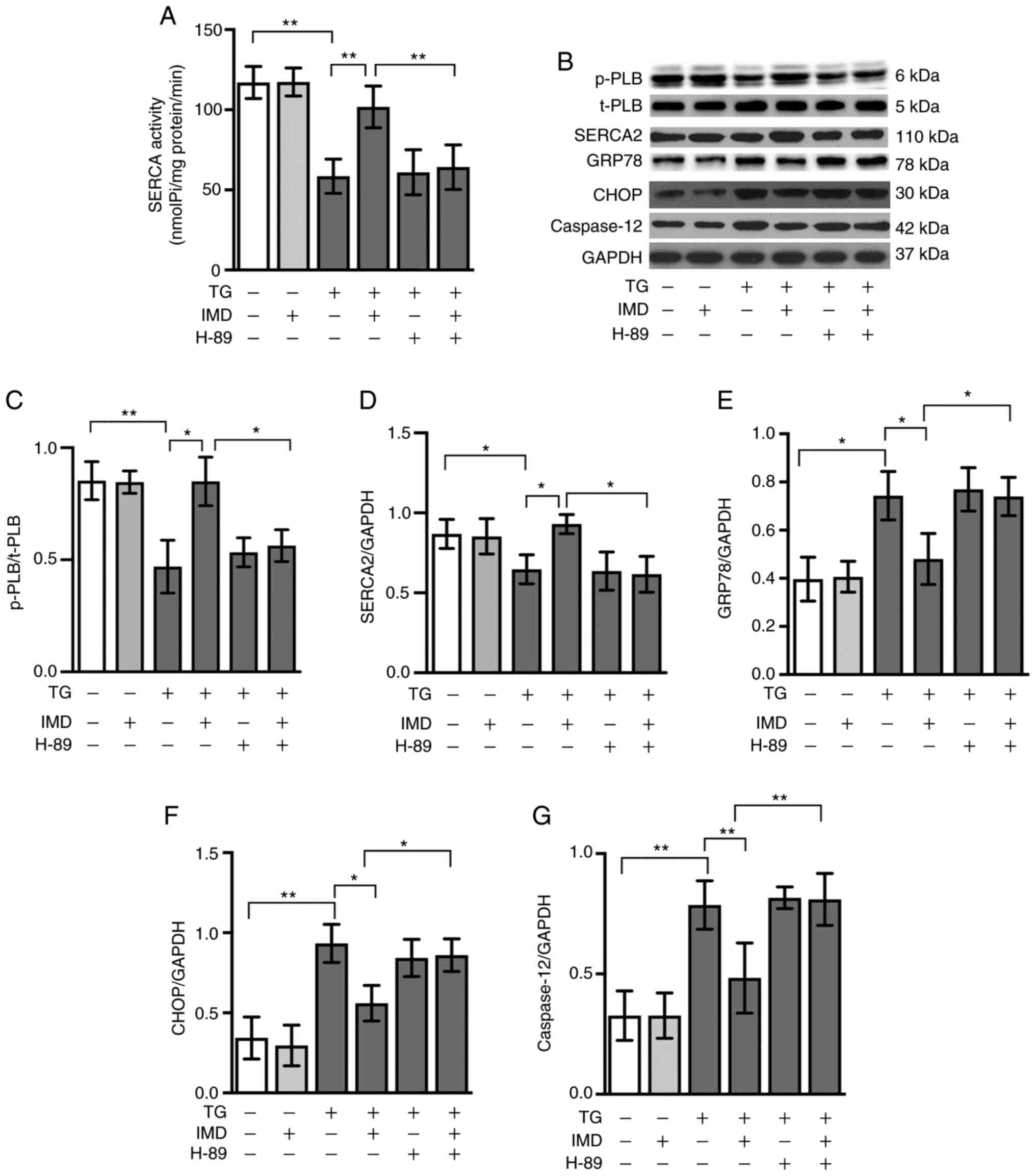 | Figure 5.PKA signaling pathway is involved in
the effects of IMD on SERCA activity and endoplasmic reticulum
stress. Cardiomyocytes were treated with TG in the presence or
absence of 100 nM IMD or 10 µM PKA inhibitor H-89 for 24 h. (A)
SERCA activity. (B) Representative western blot images of p-PLB,
t-PLB, SERCA2, GRP78, CHOP and caspase-12. Semi-quantification of
(C) p-PLB/t-PLB, (D) SERCA2, (E) GRP78, (F) CHOP and (G) caspase-12
protein levels. Data were analyzed with a student's t-test for two
group comparison. Data are presented as the mean ± SD. n=3.
*P<0.05 and **P<0.01. IMD, intermedin; SERCA,
sarco/endoplasmic reticulum calcium ATPase; PLB, phospholamban; p-,
phosphorylated; t-, total; GRP78, 78 kDa glucose-regulated protein;
CHOP, C/EBP-homologous protein; PKA, protein kinase A; TG,
thapsigargin; nmol Pi/mg prot/min, nanomoles of phosphorus ions per
milligram of protein per minute. |
PKA inhibitor H-89 treatment also abolished the
protective effects of IMD on ER stress and ER stress-related
cardiomyocyte apoptosis induced by thapsigargin, as demonstrated by
western blotting and Annexin V/PI double staining assay (Figs. 5B, 5E-G, 6A
and 6B). These results suggested
that IMD may ameliorate ER stress and ER stress-related
cardiomyocyte apoptosis induced by thapsigargin at least in part
via the PKA/SERCA pathway.
Discussion
The results of the present study demonstrated that
IMD decreased the thapsigargin-induced upregulation of GRP78, CHOP
and caspase-12, the specific markers of ER stress. IMD alleviated
thapsigargin-induced ER stress by restoring SERCA activity and
Ca2+ overload in cultured cardiomyocytes. PLB
phosphorylation may also contribute to IMD-enhanced SERCA activity.
Co-treatment with the PAK inhibitor H89 appeared to counteract the
protective effects of IMD on PLB phosphorylation and restoration of
SERCA activity, suggesting the involvement of the PKA signaling
pathway in the effects of IMD.
IMD belongs to a multifunctional
calcitonin/calcitonin gene-related peptide superfamily and shares
common receptors with calcitonin gene-related peptide,
adrenomedullin and amylin, with unique and important
cardioprotective functions, including improving cardiac function,
pro-angiogenesis, anti-oxidation and anti-ER stress (5). ER stress is increasingly recognized as
an important contributor to myocardial injury (27). ER, as the primary site of proper
folding and sorting of proteins, is vulnerable to ischemia, hypoxia
and oxidative stress. Various pathophysiological conditions disturb
the ER function and initiate the unfolded protein response to
promote cell survival (24).
However, persistent and excessive ER stress triggers apoptosis and
aggravates cardiovascular diseases, including heart failure,
cardiac hypertrophy and ischemic heart disease. ER stress and its
induced apoptosis have been highlighted as important mechanisms
underlying myocardial injury (28,29).
Alleviating ER stress is accepted as a promising therapeutic
approach for the treatment of cardiovascular diseases (30).
In previous studies, IMD has been demonstrated to
exert a cardioprotective effect against ER stress induced by
tunicamycin and dithiothreitol (6,7). The
pathways by which IMD inhibits ER stress and protects myocardial
injury are not well understood. Thapsigargin is also a classic ER
stress inducer. Tunicamycin and dithiothreitol induce ER stress via
inhibition of protein glycosylation or disulfide bond formation,
respectively (31), whereas
thapsigargin possesses a unique mechanism to induce ER stress. To
date, there have been no published reports on the effects of IMD on
thapsigargin-induced cardiomyocyte injury. The present study
assessed the protein expression levels of ER stress-related markers
GRP78, CHOP and caspase-12 in cardiomyocytes by western blotting;
thapsigargin treatment significantly upregulated the expression
levels of GRP78, CHOP and caspase-12, whereas IMD pretreatment
significantly ameliorated these changes in a dose-dependent manner,
suggesting that IMD inhibited thapsigargin-induced ER stress. To
the best of our knowledge, the present study demonstrated for the
first time that IMD may rescue cardiomyocytes from
thapsigargin-triggered ER stress.
Experimental and clinical studies have indicated
that IMD suppresses cardiac hypertrophy and heart failure, a major
health issue that is a leading cause of death worldwide (32–34).
During this process, cytosolic Ca2+ overload serves a
crucial role in the development of pathological cardiac hypertrophy
and heart failure. Cytosolic Ca2+ homeostasis is tightly
controlled by Ca2+-handling enzymes, proteins, channels
and transporters in the plasma membrane and Ca2+ storage
organelles (35). The ER is the
major Ca2+ storage organelle that releases
Ca2+ predominately via the Inositol 1,4,5-thriphosphate
and the ryanodine receptor, and uptakes Ca2+ via SERCA,
which is the only active Ca2+ transporter from the
cytosol to the ER in the heart (36). Suppression of SERCA activity and
subsequent alteration of cytosolic Ca2+ signaling can
severely impair the systolic and diastolic function of the heart,
which are major etiologies of cardiac hypertrophy and heart failure
(37–39). As the primary function of SERCA is
to replenish the sarcoplasmic reticulum Ca2+ load during
the contraction-relaxation cycle of the heart, resulting in
cytosolic Ca2+ overload, which is associated with heart
failure and cardiac hypertrophy, it was hypothesized in the present
study that restoration of SERCA activity may mediate the
cardioprotective effects of IMD on ER stress and ER stress-related
apoptosis. Thapsigargin is a highly selective inhibitor of SERCAs
(40). By inhibiting SERCAs,
thapsigargin disrupts Ca2+ transport into the ER lumen,
leading to an increase in cytoplasmic Ca2+
concentration, and subsequently activates ER stress (41). To further investigate this signaling
pathway, thapsigargin was used in the present study, as it is a
well-established model to study ER stress and SERCA activity
(42). The present results
demonstrated that thapsigargin attenuated the protein expression
and activity of SERCA2a in cardiomyocytes, and IMD pretreatment
reversed this change. Furthermore, [Ca2+]i in
thapsigargin-treated cardiomyocytes was significantly increased,
whereas IMD pretreatment inhibited this increase, suggesting that
modulating SERCA function and Ca2+ homeostasis may
contribute to IMD-mediated cardioprotection. Thus, the results of
the present study indicated that the regulation of SERCA activity
may serve an important role in the IMD-mediated protection of
cardiomyocytes against ER stress, which in part explains the
underlying mechanism of IMD improving cardiac hypertrophy and heart
failure.
PLB inhibits SERCA activity by reducing its affinity
for Ca2+ (42).
Unphosphorylated PLB binds SERCA to inhibit SERCA activity; this
inhibition is abolished upon PLB phosphorylation (25). PLB is a key regulator of SERCA
(42). Considering the relationship
between PLB and SERCA, the present study assessed the protein
levels of p-PLB and t-PLB in cardiomyocytes treated with
thapsigargin in the presence or absence of IMD to further
investigate the underlying mechanisms of IMD. The results
demonstrated that the expression of p-PLB was decreased in the
thapsigargin group compared with those in the vehicle group,
whereas IMD reversed this change, suggesting that PLB
phosphorylation may be associated with the protective effects of
IMD on SERCA activity and cytosolic Ca2+ influx.
IMD stimulates cardiomyocyte PKA activity (43), and SERCA activity can be increased
by PKA (20), which provides a
potential mechanism by which IMD stimulates SERCA function and
subsequently attenuates ER stress-related apoptosis in
thapsigargin-treated cardiomyocytes. To investigate this in the
present study, cardiomyocytes were co-pretreated with the PKA
inhibitor H-89 and/or IMD, and incubated with thapsigargin. The
results demonstrated that the PKA inhibitor H-89 inhibited the
protective effect of IMD on ER stress and SERCA function, as
indicated by the changes in the expression levels of GRP78, CHOP
and caspase-12, and SERCA activity. H-89 pretreatment also
abrogated the antiapoptotic effects of IMD. These results suggested
that IMD may exert antiapoptotic effects at least partly via the
regulation of the PKA signaling pathway in cardiomyocytes treated
with thapsigargin.
H-89 is a highly selective, but not exclusive
inhibitor of PKA, and reportedly also binds to other protein
kinases, including ribosomal protein S6 kinase β1, ribosomal
protein S6 kinase α5, rho-associated protein kinase 2, RAC-α
serine/threonine-protein kinase and ribosomal protein S6 kinase
alpha-1, with a low binding affinity (44,45).
Thus, the possibility that these protein kinases may in part
contribute to the thapsigargin-induced cardiomyocyte apoptosis
cannot be eliminated. In addition, the present study did not
investigate whether other Ca2+-handling enzymes,
proteins, channels and transporters, such as inositol
1,4,5-trisphosphate receptor type, ryanodine receptor 1 and stromal
interaction molecule 1, are involved in the protection of IMD on
thapsigargin-induced cardiomyocyte apoptosis. Accumulating evidence
has suggested that thapsigargin-induced apoptosis is mediated by
autophagy in cardiomyocytes (46,47).
The present results indicated that IMD attenuated
thapsigargin-induced cardiomyocyte apoptosis by inhibiting ER
stress with the possible involvement of the PKA/SERCA signaling
pathway. The present study focused on the mechanisms upstream of ER
stress, but did not explore the involvement of autophagy, which is
downstream of ER stress. This is a limitation of this study, and
future studies should evaluate whether autophagy contributes to the
effects of IMD on ER stress-related apoptosis. In addition, PLB has
been demonstrated to be a major regulator of SERCA activity. It is
the only SERCA-associated protein directly involved in the
development of cardiac disease, including heart failure (18). Thus, the present study focused on
the PLB/SERCA signaling pathway. Accumulating evidence has
indicated that sarcolipin also inhibits the affinity of SERCA for
Ca2+ (48,49). In future studies, we will explore
whether the effect of IMD on cardiomyocyte apoptosis is associated
with sarcolipin and other SERCA regulators.
In conclusion, the results of the present study
identified an additional signaling pathway through which IMD
responds to ER stress. The protective role of IMD in attenuating
thapsigargin-induced cardiomyocyte apoptosis may be mediated by
inhibiting ER stress with the possible involvement of the PKA/SERCA
signaling pathway. These findings provided a novel insight into the
mechanisms underlying the cardioprotective effects of IMD.
Acknowledgements
Not applicable.
Funding
The present work was supported by the National
Natural Science Foundation of China (grant no. 81600256), the
Natural Science Foundation of Shanxi Province (grant no.
2014021038-2) and the Postdoctoral Research Startup Fund of the
First Hospital of Shanxi Medical University.
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
MZ and ZL designed the experiments. ZL and JG
performed the experiments. YB and MZ analyzed the data. ZL and JG
wrote the manuscript. All authors read and approved the final
manuscript.
Ethics approval and consent to
participate
The experimental protocols were approved by the
Ethical Committee of Shanxi Medical University (approval no.
SYXk:2015-0507) and complied with the internationally accepted
principles of laboratory animal care and use.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
Glossary
Abbreviations
Abbreviations:
|
IMD
|
intermedin
|
|
ER
|
endoplasmic reticulum
|
|
GRP78
|
glucose-regulated protein 78
|
|
CHOP
|
C/EBP-homologous protein
|
|
GAPDH
|
glyceraldehyde 3-phosphate
dehydrogenase
|
|
SERCA
|
sarco/endoplasmic reticulum calcium
ATPase
|
|
PKA
|
cAMP-dependent protein kinase A
|
|
PLB
|
phospholamban
|
References
|
1
|
Takei Y, Inoue K, Ogoshi M, Kawahara T,
Bannai H and Miyano S: Identification of novel adrenomedullin in
mammals: A potent cardiovascular and renal regulator. FEBS Lett.
556:53–58. 2004. View Article : Google Scholar
|
|
2
|
Taylor MM and Samson WK: Stress hormone
secretion is altered by central administration of
intermedin/adrenomedullin-2. Brain Res. 1045:199–205. 2005.
View Article : Google Scholar
|
|
3
|
Teng X, Song J, Zhang G, Cai Y, Yuan F, Du
J, Tang C and Qi YF: Inhibition of endoplasmic reticulum stress by
intermedin(1–53) protects against myocardial injury through a PI3
kinase-Akt signaling pathway. J Mol Med (Berl). 89:1195–1205. 2011.
View Article : Google Scholar
|
|
4
|
Tang B, Zhong Z, Shen HW, Wu HP, Xiang P
and Hu B: Intermedin as a prognostic factor for major adverse
cardiovascular events in patients with ST-segment elevation acute
myocardial infarction. Peptides. 58:98–102. 2014. View Article : Google Scholar
|
|
5
|
Yang SM, Liu J and Li CX: Intermedin
protects against myocardial ischemia-reperfusion injury in
hyperlipidemia rats. Genet Mol Res. 13:8309–8319. 2014. View Article : Google Scholar
|
|
6
|
Ni X, Zhang J, Tang CX and Qi YF:
Intermedin/adrenomedullin2: An autocrine/paracrine factor in
vascular homeostasis and disease. Sci China Life Sci. 57:781–789.
2014. View Article : Google Scholar
|
|
7
|
Bell D, Gordon BJ, Lavery A, Megaw K,
Kinney MO and Harbinson MT: Plasma levels of intermedin
(adrenomedullin-2) in healthy human volunteers and patients with
heart failure. Peptides. 76:19–29. 2016. View Article : Google Scholar
|
|
8
|
Lv Z, Wu K, Chen X, Zhang X and Hong B:
Plasma intermedin levels in patients with acute myocardial
infarction. Peptides. 43:121–125. 2013. View Article : Google Scholar
|
|
9
|
Zhang JS, Hou YL, Lu WW, Ni XQ, Lin F, Yu
YR, Tang CS and Qi YF: Intermedin1-53 protects against
myocardial fibrosis by inhibiting endoplasmic reticulum stress and
inflammation induced by homocysteine in apolipoprotein E-deficient
mice. J Atheroscler Thromb. 23:1294–1306. 2016. View Article : Google Scholar
|
|
10
|
Canova NK, Kmonickova E, Martinek J, Zidek
Z and Farghali H: Thapsigargin, a selective inhibitor of
sarco-endoplasmic reticulum Ca2+ -ATPases, modulates nitric oxide
production and cell death of primary rat hepatocytes in culture.
Cell Biol Toxicol. 23:337–354. 2007. View Article : Google Scholar
|
|
11
|
Chen G, Shen Y, Li X, Jiang Q, Cheng S, Gu
Y, Liu L and Cao Y: The endoplasmic reticulum stress inducer
thapsigargin enhances the toxicity of ZnO nanoparticles to
macrophages and macrophage-endothelial co-culture. Environ Toxicol
Pharmacol. 50:103–110. 2017. View Article : Google Scholar
|
|
12
|
Chen X, Zhang X, Gross S, Houser SR and
Soboloff J: Acetylation of SERCA2a, another target for heart
failure treatment? Circ Res. 124:1285–1287. 2019. View Article : Google Scholar
|
|
13
|
Prasad AM, Ma H, Sumbilla C, Lee DI, Klein
MG and Inesi G: Phenylephrine hypertrophy, Ca2+-ATPase
(SERCA2), and Ca2+ signaling in neonatal rat cardiac
myocytes. Am J Physiol Cell Physiol. 292:C2269–C2275. 2007.
View Article : Google Scholar
|
|
14
|
Liu M, Xue M, Wang XR, Tao TQ, Xu FF, Liu
XH and Shi DZ: Panax quinquefolium saponin attenuates cardiomyocyte
apoptosis induced by thapsigargin through inhibition of endoplasmic
reticulum stress. J Geriatr Cardiol. 12:540–546. 2015.
|
|
15
|
Adachi T: Modulation of vascular
sarco/endoplasmic reticulum calcium ATPase in cardiovascular
pathophysiology. Adv Pharmacol. 59:165–195. 2010. View Article : Google Scholar
|
|
16
|
Cook NL, Viola HM, Sharov VS, Hool LC,
Schoneich C and Davies MJ: Myeloperoxidase-derived oxidants inhibit
sarco/endoplasmic reticulum Ca2+-ATPase activity and perturb Ca2+
homeostasis in human coronary artery endothelial cells. Free Radic
Biol Med. 52:951–961. 2012. View Article : Google Scholar
|
|
17
|
Zhang C, Bose DD and Thomas DW:
Paradoxical effects of sarco/endoplasmic reticulum Ca(2+)-ATPase
(SERCA) activator gingerol on NG115-401L neuronal cells: Failure to
augment ER Ca(2+) uptake and protect against ER stress-induced cell
death. Eur J Pharmacol. 762:165–173. 2015. View Article : Google Scholar
|
|
18
|
Kranias EG and Hajjar RJ: Modulation of
cardiac contractility by the phospholamban/SERCA2a regulatome. Circ
Res. 110:1646–1660. 2012. View Article : Google Scholar
|
|
19
|
Gorski PA, Ceholski DK and Young HS:
Structure-function relationship of the SERCA pump and its
regulation by phospholamban and sarcolipin. Adv Exp Med Biol.
981:77–119. 2017. View Article : Google Scholar
|
|
20
|
Cerra MC and Imbrogno S: Phospholamban and
cardiac function: A comparative perspective in vertebrates. Acta
Physiol (Oxf). 205:9–25. 2012. View Article : Google Scholar
|
|
21
|
Xu J, Han Q, Shi H, Liu W, Chu T and Li H:
Role of PKA in the process of neonatal cardiomyocyte hypertrophy
induced by urotensin II. Int J Mol Med. 40:499–504. 2017.
View Article : Google Scholar
|
|
22
|
Ogden BE, Pang William W, Agui T and Lee
BH: Laboratory Animal Laws, Regulations, Guidelines and Standards
in China Mainland, Japan, and Korea. ILAR J. 57:301–311. 2016.
View Article : Google Scholar
|
|
23
|
Bian YF, Hao XY, Gao F, Yang HY, Zhang N
and Xiao CS: Adiponectin attenuates hypoxia/reoxygenation-induced
cardiomyocyte injury through inhibition of endoplasmic reticulum
stress. J Investiq Med. 59:921–925. 2011. View Article : Google Scholar
|
|
24
|
Sozen E, Karademir B and Ozer NK: Basic
mechanisms in endoplasmic reticulum stress andrelation to
cardiovascular diseases. Free Radic Biol Med. 78:30–41. 2015.
View Article : Google Scholar
|
|
25
|
Gustavsson M, Traaseth NJ, Karim CB,
Lockamy EL, Thomas DD and Veglia G: Lipid-mediated
folding/unfolding of phospholamban as a regulatory mechanism for
the sarcoplasmic reticulum Ca2+-ATPase. J Mol Biol. 408:755–765.
2011. View Article : Google Scholar
|
|
26
|
Periasamy M, Bhupathy P and Babu GJ:
Regulation of sarcoplasmic reticulum Ca2+ ATPase pump expression
and its relevance to cardiac muscle physiology and pathology.
Cardiovasc Res. 77:265–273. 2008. View Article : Google Scholar
|
|
27
|
Dickhout JG, Carlisle RE and Austin RC:
Interrelationship between cardiac hypertrophy, heart failure, and
chronic kidney disease: Endoplasmic reticulum stress as a mediator
of pathogenesis. Circ Res. 108:629–642. 2011. View Article : Google Scholar
|
|
28
|
Wang J, Hu X and Jiang H: ER
stress-induced apoptosis: A novel therapeutic target in myocardial
ischemia and reperfusion injury. Int J Cardiol. 214:233–234. 2016.
View Article : Google Scholar
|
|
29
|
Wang M, Meng XB, Yu YL, Sun GB, Xu XD,
Zhang XP, Dong X, Ye JX, Xu HB, Sun YF and Sun XB: Elatoside C
protects against hypoxia/reoxygenation-induced apoptosis in H9c2
cardiomyocytes through the reduction of endoplasmic reticulum
stress partially depending on STAT3 activation. Apoptosis.
19:1727–1735. 2014. View Article : Google Scholar
|
|
30
|
Hong J, Kim K, Kim JH and Park Y: The role
of endoplasmic reticulum stress in cardiovascular disease and
exercise. Int J Vasc Med. 2017:20492172017.
|
|
31
|
Li B, Yi P, Zhang B, Xu C, Liu Q, Pi Z, Xu
X, Chevet E and Liu J: Differences in endoplasmic reticulum stress
signalling kinetics determine cell survival outcome through
activation of MKP-1. Cell Signal. 23:35–45. 2011. View Article : Google Scholar
|
|
32
|
Chen H, Wang X, Tong M, Wu D, Wu S, Chen
J, Wang X, Wang X, Kang Y, Tang H, Tang C and Jiang W: Intermedin
suppresses pressure overload cardiac hypertrophy through activation
of autophagy. PLoS One. 8:e647572013. View Article : Google Scholar
|
|
33
|
Liu K, Deng X, Gong L, Chen X, Wang S,
Chen H, Chen X, Amrit B and He S: The effect of intermedin on
angiotensin II and endothelin-1 induced ventricular myocyte
hypertrophy in neonatal rat. Clin Lab. 59:589–596. 2013. View Article : Google Scholar
|
|
34
|
Hirose T, Totsune K, Mori N, Morimoto R,
Hashimoto M, Nakashige Y, Metoki H, Asayama K, Kikuya M, Ohkubo T,
et al: Increased expression of adrenomedullin 2/intermedin in rat
hearts with congestive heart failure. Eur J Heart Fail. 10:840–849.
2008. View Article : Google Scholar
|
|
35
|
Reddish FN, Miller CL, Gorkhali R and Yang
JJ: Calcium dynamics mediated by the endoplasmic/sarcoplasmic
reticulum and related diseases. Int J Mol Sci. 18:10242017.
View Article : Google Scholar
|
|
36
|
Chemaly ER, Troncone L and Lebeche D:
SERCA control of cell death and survival. Cell Calcium. 69:46–61.
2018. View Article : Google Scholar
|
|
37
|
Li L, Louch WE, Niederer SA, Aronsen JM,
Christensen G, Sejersted OM and Smith NP: Sodium accumulation in
SERCA knockout-induced heart failure. Biophys J. 102:2039–2048.
2012. View Article : Google Scholar
|
|
38
|
Roe AT, Ruud M, Espe EK, Manfra O,
Longobardi S, Aronsen JM, Norden ES, Husebye T, Kolstad TRS,
Cataliotti A, et al: Regional diastolic dysfunction in
post-infarction heart failure: Role of local mechanical load and
SERCA expression. Cardiovasc Res. 115:752–764. 2019. View Article : Google Scholar
|
|
39
|
Shi H, Han Q, Xu J, Liu W, Chu T and Zhao
L: Urotensin II induction of neonatal cardiomyocyte hypertrophy
involves the CaMKII/PLN/SERCA 2a signaling pathway. Gene. 583:8–14.
2016. View Article : Google Scholar
|
|
40
|
Kamiya T, Hara H and Adachi T: Effect of
endoplasmic reticulum (ER) stress inducer thapsigargin on the
expression of extracellular-superoxide dismutase in mouse 3T3-L1
adipocytes. J Clin Biochem Nutr. 52:101–105. 2013. View Article : Google Scholar
|
|
41
|
Lytton J, Westlin M and Hanley MR:
Thapsigargin inhibits the sarcoplasmic or endoplasmic reticulum
Ca-ATPase family of calcium pumps. J Biol Chem. 266:17067–17071.
1991.
|
|
42
|
Schmitt JP, Ahmad F, Lorenz K, Hein L,
Schulz S, Asahi M, Maclennan DH, Seidman CE, Seidman JG and Lohse
MJ: Alterations of phospholamban function can exhibit cardiotoxic
effects independent of excessive sarcoplasmic reticulum
Ca2+-ATPase inhibition. Circulation. 119:436–444. 2009.
View Article : Google Scholar
|
|
43
|
Bell D and McDermott BJ: Intermedin
(adrenomedullin-2): A novel counter-regulatory peptide in the
cardiovascular and renal systems. Br J Pharmacol. 153 (Suppl
1):S247–S262. 2008. View Article : Google Scholar
|
|
44
|
Lochner A and Moolman JA: The many faces
of H89: A review. Cardiovasc Drug Rev. 24:261–274. 2006. View Article : Google Scholar
|
|
45
|
Saad NS, Elnakish MT, Ahmed AAE and
Janssen PML: Protein kinase a as a promising target for heart
failure drug development. Arch Med Res. 49:530–537. 2018.
View Article : Google Scholar
|
|
46
|
Zhang X, Yuan Y, Jiang L, Zhang J, Gao J,
Shen Z, Zheng Y, Deng T, Yan H, Li W, et al: Endoplasmic reticulum
stress induced by tunicamycin and thapsigargin protects against
transient ischemic brain injury: Involvement of PARK2-dependent
mitophagy. Autophagy. 10:1801–1013. 2014. View Article : Google Scholar
|
|
47
|
Lindner P, Christensen B, Nissen P, Møller
JV and Engedal N: Cell death induced by the ER stressor
thapsigargin involves death receptor 5, a non-autophagic function
of MAP1LC3B, and distinct contributions from unfolded protein
response components. Cell Commun Signal. 18:122020. View Article : Google Scholar
|
|
48
|
Bhupathy P, Babu GJ and Periasamy M:
Sarcolipin and phospholamban as regulators of cardiac sarcoplasmic
reticulum Ca2+ ATPase. J Mol Cell Cardiol. 42:903–911. 2007.
View Article : Google Scholar
|
|
49
|
Asahi M, Nakayama H, Tada M and Otsu K:
Regulation of sarco(endo)plasmic reticulum Ca2+ adenosine
triphosphatase by phospholamban and sarcolipin: Implication for
cardiac hypertrophy and failure. Trends Cardiovasc Med. 13:152–157.
2003. View Article : Google Scholar
|