Introduction
Cerebral ischemia is considered to be a leading
cause of morbidity and mortality in the developed world (1). Although reperfusion is the recommended
therapy for effective treatment of cerebral ischemia, it may also
exacerbate brain injury and functional damage in a process termed
cerebral ischemia/reperfusion (I/R) injury (2). In recent years, numerous studies have
demonstrated that multiple mechanisms involving a variety of
signaling pathways and biological processes are involved in the
process of cerebral I/R injury (3–5);
however, the mechanism is complex and our knowledge on the subject
is unsatisfactory at present. A number of studies have revealed
that oxidative stress and inflammation are the primary pathological
causes of neuronal loss following cerebral I/R (6,7).
Therefore, a focus on understanding the molecular mechanisms
related to oxidative stress and the inflammatory response behind
cerebral I/R injury may promote the development of more efficient
therapeutic agents.
MicroRNAs (miRs/miRNAs) are a class of small
non-coding RNAs that are 18–23 nucleotides in length that can
regulate a wide range of biological processes in cerebral ischemia,
including oxidative stress and the inflammatory reaction. It has
been proposed that they could be potential diagnostic markers and
promising therapeutic agents in cerebral ischemia (8,9). Since
miR-126 has been reported to regulate vascular integrity and
angiogenesis, numerous studies have demonstrated that miR-126
serves an important role in the pathogenesis and complications of
cerebral ischemic stroke (10,11).
Pan et al (12) revealed
that miR-126 overexpression may promote the functions of
endothelial progenitor cells (EPCs) under hypoxic conditions,
resulting in the enhanced therapeutic efficacy of EPCs in ischemic
stroke. Furthermore, the promotion of the miR-126-related signaling
pathway induced by long non-coding RNAs attenuates ischemic
neuronal death and apoptosis (10).
However, the underlying protective mechanisms of miR-126 in
cerebral I/R injury remain to be elucidated.
NAD-dependent protein deacetylase sirtuin-1(SIRT1),
a nicotinamide adenine dinucleotide-dependent histone deacetylase,
regulates numerous physiological and pathological processes,
including cell survival, apoptosis, oxidative stress, inflammation
and neuroprotection (13–15). Additionally, increasing evidence has
suggested that SIRT1 may have beneficial effects in cerebral I/R
injury (16). Nuclear factor
erythroid 2-related factor 2 (Nrf2) is capable of multiple
biological effects, and the activation of this protein, which
promotes Nrf2 translocation from the cytoplasm to the nucleus and
activates the transcription of antioxidant, cytoprotective and
anti-inflammatory genes, such as heme oxygenase-1 (HO-1), is
crucial for cellular defense mechanisms (17). Furthermore, it has been reported
that SIRT1 is associated with the activation of Nrf2 (18). Several studies have demonstrated
that activation of the SIRT1/Nrf2 signaling pathway increases
resistance to oxidative stress injury and has protective effects
against inflammation (19,20). Recent studies have demonstrated that
the dysregulation of the SIRT1/Nrf2 signaling pathway is involved
in the pathogenesis of a number of neurodegenerative diseases
(20,21). Furthermore, Chan et al
(22) has demonstrated that the
SIRT1/Nrf2 signaling pathway mediates the protective effects
against diabetic hyperglycemia-exacerbated cerebral I/R injury.
Notably, a study by Xu et al (23) revealed that SIRT1 can be regulated
by miR-126. However, at present, the role of the SIRT1/Nrf2
signaling pathway in the development of cerebral I/R injury and the
protective effects of miR-126 on cerebral I/R injury remains
largely unknown.
The present study aimed to investigate the
protective effects of miR-126 overexpression on oxidative stress
and the inflammatory response in human umbilical vein endothelial
cells (HUVECs) subjected to oxygen-glucose
deprivation/reoxygenation (OGD/R), and the role of the SIRT1/Nrf2
signaling pathway in this process. The present study demonstrated
that miR-126 overexpression attenuated OGD/R-induced cell injury of
HUVECs by inhibiting oxidative stress and inflammatory response via
the activation of SIRT1/Nrf2 signaling pathway.
Materials and methods
Chemicals and reagents
Dulbecco's modified Eagle's medium (DMEM; cat. no.
11965092) and fetal bovine serum (FBS; cat. no. 16140071) were
purchased from Gibco (Thermo Fisher Scientific, Inc.). The Cell
Counting Kit-8 (CCK-8) assay (cat. no. C0038) and Reactive Oxygen
Species (ROS) assay kit (cat. no. S0033M) were obtained from
Beyotime Institute of Biotechnology. The Annexin V-FITC/propidium
iodide (PI) apoptosis kit (cat. no. 556547) was purchased from BD
Biosciences. The lactate dehydrogenase (LDH) assay kit (cat. no.
A020-2-2), cell malondialdehyde (MDA) assay kit (cat. no.
A003-4-1), superoxide dismutase (SOD) assay kit (cat. no.
A001-3-2), glutathione peroxidase (GSH-Px) assay kit (cat. no.
A005-1-2), interleukin-6 (IL-6) assay kit (cat. no. H007),
interleukin-1β (IL-1β) assay kit (cat. no. H002), tumor necrosis
factor-α (TNF-α) assay kit (cat. no. H052) and interleukin-10
(IL-10) assay kit (cat. no. H009) were obtained from Nanjing
Jiancheng Bioengineering Institute. Antibodies against SIRT1 (cat.
no. 8469), Nrf2 (cat. no. 12721), HO-1 (cat. no. 82206), histone H3
(cat. no. 4499) and GAPDH (cat. no. 5174) were purchased from Cell
Signaling Technology, Inc. The antibody against GAPDH (cat. no.
ab181602) was obtained from Abcam.
Cell culture and OGD/R treatment
HUVECs were provided by The Cell Bank of Type
Culture Collection of the Chinese Academy of Sciences, and cultured
in DMEM containing 10% FBS, 100 U/ml penicillin and 100 U/ml
streptomycin in a humidified incubator with 5% CO2 at
37°C. Culture media were exchanged every 2 days. To induce OGD/R
injury, HUVECs were incubated with glucose-free DMEM in a
humidified chamber filled with 95% N2 and 5%
CO2 at 37°C for 4 h. Subsequently, cells were
transferred to complete culture medium and maintained in normoxic
conditions for 6, 12 or 24 h to induce reperfusion.
Cell transfection
miR-126 mimics (cat. no. miR10000444-1-5;
5′-UCGUACCGUGAGUAAUAAUGCG-3′) and miR-negative control (miR-NC;
cat. no. miR1N0000002-1-5; 5′-UUCUCCGAACGUGUCACGUTT-3′) were
synthesized by Guangzhou RiboBio Co, Ltd. SIRT1-small interfering
RNA (SIRT1-siRNA; forward, 5′-ACUUUGCUGUAACCCUGUA-3′ and reverse,
5′-UACAGGGUUACAGCAAAGU-3′) or negative control siRNA sequence
(Con-siRNA; 5′-CUAGCUUAUGUGGACCUCG-3′) were synthesized by Sangon
Biotech Co., Ltd. Transfection of miR-126 mimics, miR-NC,
SIRT1-siRNA or Con-siRNA at a final concentration of 50 nM were
performed using Lipofectamine® 3000 (Invitrogen; Thermo
Fisher Scientific, Inc.), according to the manufacturer's
protocols. At 48 h after transfection, transfection efficiency was
determined by reverse transcription-quantitative PCR (RT-qPCR) or
western blotting.
RT-qPCR
Total RNA was extracted from HUVECs using
TRIzol® reagent (cat. no. 15596018; Invitrogen; Thermo
Fisher Scientific, Inc.) according to the manufacturer's protocols.
cDNA was synthesized with the PrimeScript™ One Step RT-PCR kit
(cat. no. RR055B; Takara Biotechnology Co., Ltd.) at a temperature
of 45°C for 60 min and 72°C for 5 min using the extracted total RNA
(30 µg) as the template. miR-126 was amplified using a Power
SYBR-Green PCR Master mix (Applied Biosystems; Thermo Fisher
Scientific, Inc.) with an Applied Biosystems 7500 Real-Time PCR
system (Applied Biosystems; Thermo Fisher Scientific, Inc.). The
primers were as follows: miR-126 forward,
5′-GCAATGCTAGATTTAGTAATT-3′ and reverse,
3′-GCGCAUGGUUUCAUUAUUAC-5′; and U6 forward,
5′-TGACACGCAAATTCGTGAAGCGTTC-3′ and reverse,
5′-CCAGTCTCAGGGTCCGAGGTATTC-3′. U6 was used as the endogenous
control. The RT-qPCR conditions were as follows: 94°C for 4 min,
followed by 94°C for 20 sec, 60°C for 30 sec and 72°C for 30 sec
(35 cycles). Relative gene expression was calculated according to
the 2−ΔΔCq method (24).
CCK-8 assay
The viability of HUVECs was detected using the CCK-8
assay kit according to the manufacturer's protocol. Briefly, cells
in 96-well plates were incubated with CCK-8 solution (10 µl/well)
for 3 h at 37°C, and then the optical density (OD) at an emission
wavelength of 450 nm was measured on a microplate reader (Spectra
Max M5; Molecular Devices, LLC).
Lactate dehydrogenase (LDH) releases
assay
HUVECs injury was determined by the release of LDH
using a LDH assay kit (cat. no. C0016; Beyotime Institute of
Biotechnology) according to manufacturer's protocols. After
treatment as detailed above, the culture supernatant was collected
and then incubated with LDH reagent for 30 min at room temperature
in the dark. The absorbance at 490 nm was assessed by a microplate
reader (Spectra Max M5; Molecular Devices, LLC).
Measurement of intracellular ROS
generation
Intracellular ROS production was determined using
the non-fluorescent probe 2′,7′-dichlorofluorescein diacetate
(DCFH-DA; cat. no. S0033M; Beyotime Institute of Biotechnology)
according to the manufacturer's protocol. In brief, HUVECs cultured
in 6-well plates were incubated with DCFH-DA (10 µM) in FBS-free
DMEM at 37°C for 20 min. After three washes with FBS-free DMEM, the
cells were collected and the fluorescent density of each group was
quantified using a FACSCanto II flow cytometer (BD Biosciences) and
the data were analyzed by the FlowJo software v10.2 (FlowJo
LLC).
MDA content, and SOD and GSH-Px
activity assays
HUVECs were lysed in RIPA buffer (Beyotime Institute
of Biotechnology) on ice for 30 min. Following centrifugation at
13,400 × g at 4°C for 10 min, the MDA content, and SOD and GSH-Px
activity in the supernatant were evaluated using the appropriate
kits (Nanjing Jiancheng Bioengineering Institute), according to the
manufacturer's protocols.
Evaluation of IL-1β, IL-6, TNF-α and
IL-10 levels via ELISA
HUVECs were collected and homogenized in ice-cold
PBS. Following centrifugation at 13,400 × g for 10 min at 4°C, the
supernatant was assayed for IL-1β, IL-6, TNF-α and IL-10 levels in
accordance with the manufacturer's protocols. The OD was measured
at a wavelength of 450 nm on a microplate reader
(Infinite® 200 PRO NanoQuant; Tecan Group, Ltd.).
Caspase-3 activity assay
Caspase-3 activity was measured using the
Apo-ONE® Homogeneous Caspase-3 assay kit (Promega
Corporation), according to the manufacturer's protocols. The OD at
an emission wavelength of 570 nm was measured on a microplate
reader (Spectra Max M5; Molecular Devices, LLC).
Flow cytometry
HUVEC apoptosis was detected using the Annexin
V-FITC/PI Apoptosis Detection kit according to the manufacturer's
protocols. Following the aforementioned treatment, cells were
harvested, washed twice with PBS and then resuspended in 1X buffer
solution. Subsequently, cells were stained successively with
Annexin V-FITC (50 µl) and PI (10 µl) at 37°C for 15 min. The
apoptotic cells were evaluated using a flow cytometer (FACScan; BD
Biosciences) and data were analyzed using CellQuest software
(version 3.1; BD Biosciences). The experiment was performed
independently three times. The percentage of apoptotic cells was
calculated by the ratio of cells with FITC-Annexin
V+PI− (early apoptosis) and FITC-Annexin
V+PI+ (late apoptosis) to the total number of
cells using a FACSCanto II flow cytometer (BD Biosciences) and the
data were analyzed by the FlowJo software v10.2 (FlowJo LLC).
Western blotting
HUVECs were lysed with RIPA buffer containing 1%
(v/v) protease and phosphatase inhibitors (Beyotime Institute of
Biotechnology). Cytoplasmic and nuclear proteins were extracted
using a Nuclear and Cytoplasmic Protein Extraction Kit (cat. no.
P0028; Beyotime Institute of Biotechnology), according to the
manufacturer's instructions. Protein concentration was determined
by a BCA Protein Assay Kit (cat. no. P0012; Beyotime Institute of
Biotechnology). Equal amounts of protein (30 µg) were separated via
SDS-PAGE on a 12% gel, and then transferred to PVDF membranes (EMD
Millipore). Following blocking with 5% non-fat dry milk at room
temperature for 2 h, the membranes were incubated with the
following primary antibodies at 4°C overnight: SIRT1, Nrf2, HO-1,
histone H3 and GAPDH (1:2,000). Histone H3 was used as an internal
reference for nuclear protein and GAPDH was used as an internal
reference for total protein. Subsequently, the membranes were
washed three times. Afterwards, the membranes were incubated with
anti-rabbit HRP-conjugated secondary antibody (cat. no. 7074;
1:5,000; Cell Signaling Technology, Inc.) and anti-rat
HRP-conjugated secondary antibody (cat. no. 7077; 1:5,000; Cell
Signaling Technology, Inc.) at room temperature for 2 h, and then
the bands were visualized using enhanced chemiluminescence
(Amersham; Cytiva). The band densities were quantified using ImageJ
software (version 1.41; National Institutes of Health).
Statistical analysis
All data were analyzed using GraphPad Prism 5.0
(GraphPad Software, Inc.) and presented as the mean ± SD. One-way
ANOVA followed by the Tukey's post hoc test was used to compare
data from different groups. P<0.05 was considered to indicate a
statistically significant difference.
Results
miR-126 mimics reverse OGD/R-induced
HUVEC injury
As shown in Fig. 1,
compared with the control group, cell activity was significantly
decreased with increasing hypoxia duration in the OGD/R group
(Fig. 1A). Furthermore, LDH release
assay results revealed that OGD/R increased LDH release in the
culture supernatant (Fig. 1B).
Additionally, the expression of miR-126 in the OGD/R group was
significantly reduced compared with those in the control group
(Fig. 1C). OGD treatment for 4 h
and R for 24 h were selected for subsequent experiments.
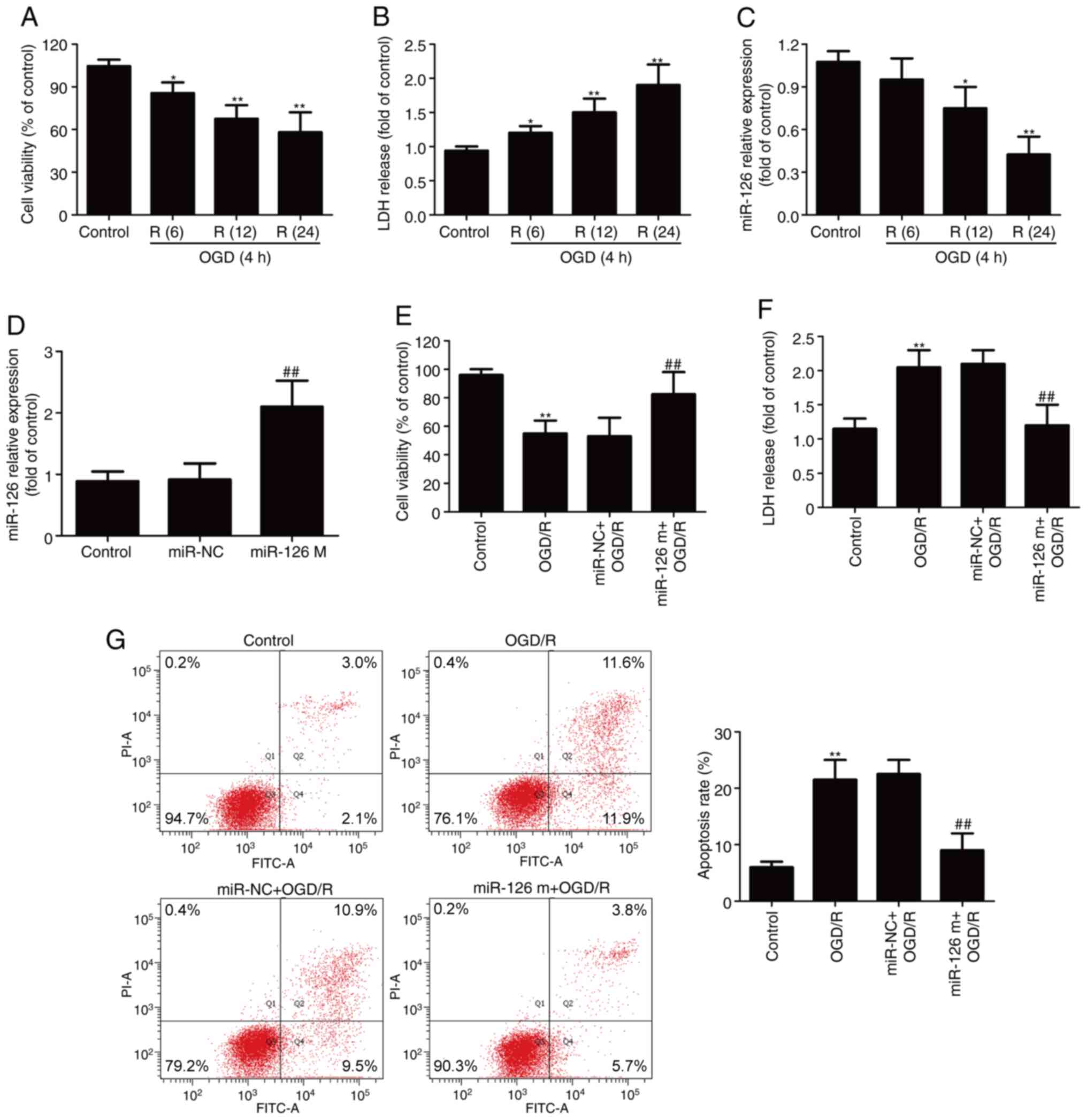 | Figure 1.Effect of OGD/R on cell survival and
apoptosis in the presence or absence of miR-126 mimics in HUVECs.
Cells were exposed to OGD for 4 h, followed by reoxygenation for 6,
12 and 24 h. (A) The cell viability of each group was determined by
CCK-8 assay. (B) LDH release from HUVECs was measured using a LDH
Cytotoxicity assay kit. (C) miR-126 expression was determined via
RT-qPCR. HUVECs were transfected with miR-126 m or miR-NC, and (D)
miR-126 expression was determined via RT-qPCR. HUVECs were
transfected with miR-126 m or miR-NC before OGD (4 h)/R (24 h)
exposure. (E) The cell viability of each group was determined by
CCK-8 assay. (F) LDH release from HUVECs was measured using a LDH
Cytotoxicity assay kit. (G) The rate of apoptosis was detected by
Annexin V/PI double staining followed by flow cytometry. Data are
presented as the mean ± SD of three independent experiments.
*P<0.05 and **P<0.01 vs. control group;
##P<0.01 vs. miR-NC or miR-NC + OGD/R group. miR-126
m, miR-126 mimics; miR-NC, miR-negative control; miR, microRNA;
OGD, oxygen-glucose deprivation; R, reoxygenation; HUVEC, human
umbilical vein endothelial cell; LDH, lactate dehydrogenase;
RT-qPCR, reverse transcription-quantitative PCR. |
Based on the decrease in miR-126 expression and the
protective effects of miR-126 in nervous system diseases (10–12),
it was speculated that the gain-of-function of miR-126 may protect
against neurological deficits induced by OGD/R injury. To verify
this assumption, HUVECs were transfected with miR-126 mimics to
promote the expression of miR-126. The results demonstrated that
miR-126 mimics successfully increased the level of miR-126 compared
with miR-NC transfection (Fig. 1D).
Based on this, it was found that transfection with miR-126 mimics
resulted in an increase in cell viability (Fig. 1E) and a decrease in LDH release
(Fig. 1F) compared with miR-NC
transfection under OGD/R conditions. Additionally, Annexin V/PI
double staining followed by flow cytometry revealed that, compared
with the control group, OGD/R increased the apoptotic rate of
HUVECs. However, this effect of OGD/R was reversed by transfection
with miR-126 mimics (Fig. 1G).
These results indicated that miR-126 overexpression protected
HUVECs against OGD/R injury and apoptosis.
miR-126 mimics attenuate OGD/R-induced
inhibition of the SIRT1/Nrf2 signaling pathway in HUVECs
SIRT1 and Nrf2 are pivotal regulators of I/R injury
in the brain (16,17). To investigate whether the SIRT1/Nrf2
signaling pathway was involved in the neuroprotective effects of
miR-126, the expression levels of proteins related to the
SIRT1/Nrf2 signaling pathway were detected in OGD/R-treated HUVECs
in the presence of miR-126 mimics. Western blotting (Fig. 2A) revealed that transfection with
miR-126 mimics increased the expression level of SIRT1 in
OGD/R-treated HUVECs compared with miR-NC transfection (Fig. 2B). In addition, transfection with
miR-126 mimics significantly decreased Nrf2 expression in the
cytoplasm (Fig. 2C), with a
corresponding increase in the nucleus (Fig. 2D and E), compared with miR-NC
transfection in HUVECs exposed to OGD/R, indicating that miR-126
promoted the nuclear translocation of Nrf2. Additionally, miR-126
mimics reversed the OGD/R-induced decrease in the expression of
HO-1 (Fig. 2F). Overall, these
results suggested that miR-126 overexpression enhanced the
activation of the SIRT1/Nrf2 signaling pathway.
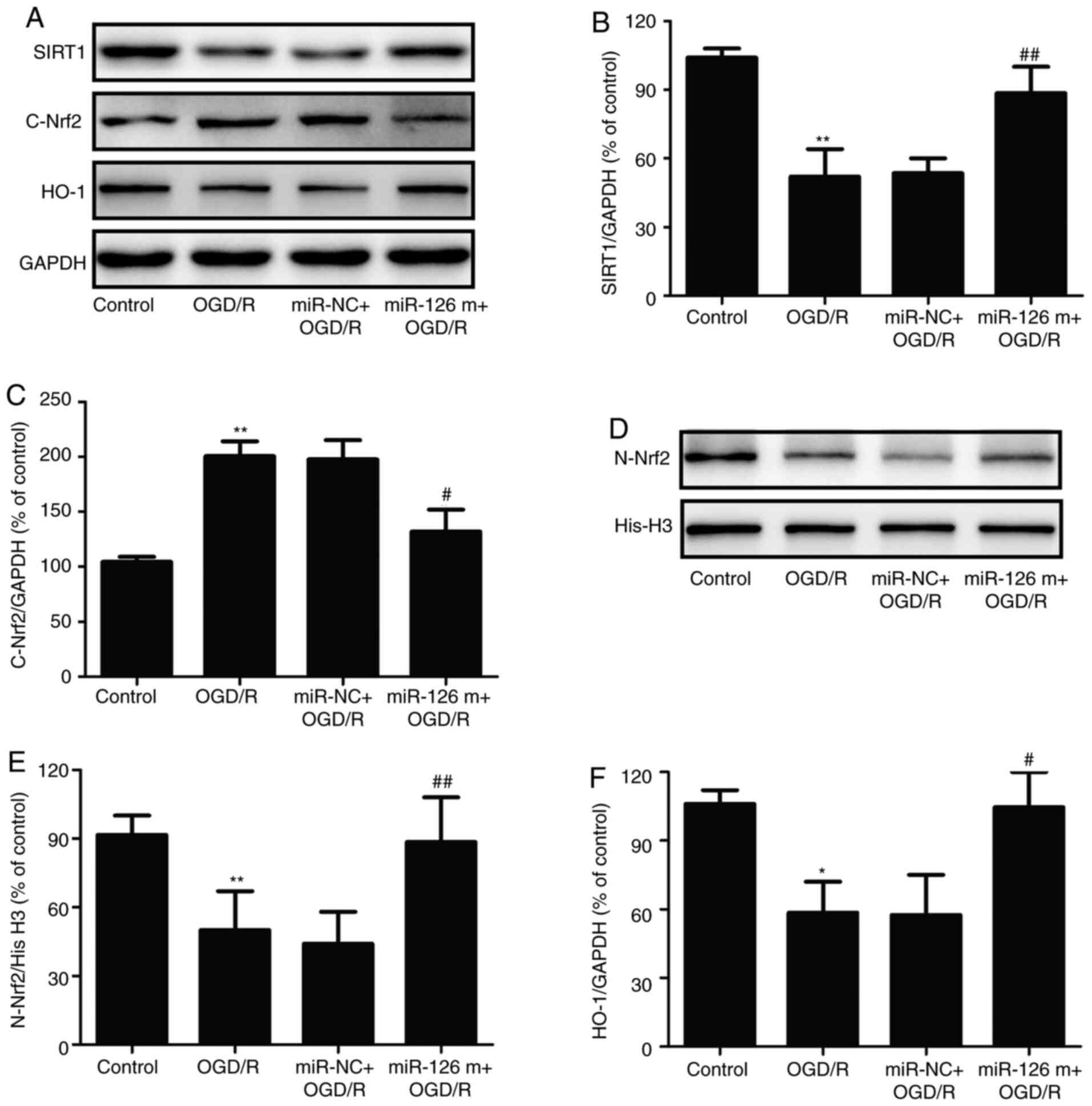 | Figure 2.Effects of miR-126 overexpression on
SIRT1 expression, Nrf2 protein distribution and HO-1 protein
expression in OGD/R-exposed HUVECs. HUVECs were transfected with
miR-126 m or miR-NC prior to OGD (4 h)/R (24 h) exposure. (A)
Protein expression was determined via western blotting.
Semi-quantification of (B) cytosolic SIRT1 and (C) Nrf2 normalized
to GAPDH. (D) Representative western blots of nuclear Nrf2. (E)
Quantification of nuclear Nrf2 normalized to histone H3. (F)
Quantification of HO-1 normalized to GAPDH. Data are presented as
the mean ± SD of at least three independent experiments. *P<0.05
and **P<0.01 vs. control group; #P<0.05 and
##P<0.01 vs. miR-NC + OGD/R group. miR-126 m, miR-126
mimics; miR-NC, miR-negative control; miR, microRNA; OGD,
oxygen-glucose deprivation; R, reoxygenation; HUVEC, human
umbilical vein endothelial cell; SIRT1, NAD-dependent protein
deacetylase sirtuin-1; Nrf2, nuclear factor erythroid 2-related
factor 2; HO-1, heme oxygenase 1. |
Inhibition of the SIRT1/Nrf2 signaling
pathway reverses the protective effect of miR-126 against OGD/R
injury in HUVECs
To determine the exact role of the SIRT1/Nrf2
signaling pathway in the protective effects of miR-126 on OGD/R
injury, HUVECs were pre-transfected with SIRT1-siRNA to inhibit the
SIRT1/Nrf2 signaling pathway. RT-qPCR results showed that
SIRT1-siRNA transfection significantly reduced the expression of
SIRT1 mRNA compared with Con-siRNA transfection (Fig. 3A). In addition, western blotting
results (Fig. 3B) demonstrated that
SIRT1-siRNA significantly reduced the miR-126 mimics-induced
upregulation of SIRT1 in OGD/R-treated HUVECs. Furthermore,
SIRT1-siRNA significantly reversed the miR-126 mimics-induced
nuclear translocation of Nrf2, as demonstrated by the increase in
Nrf2 expression in the cytoplasm (Fig.
3B) and the decrease in Nrf2 expression in the nucleus
(Fig. 3B). Additionally,
SIRT1-siRNA blocked the miR-126 mimics-induced up-regulation of
HO-1 expression (Fig. 3B). These
results indicated that transfection with SIRT1-siRNA attenuated the
miR-126 mimics-induced activation of the SIRT1/Nrf2 signaling
pathway.
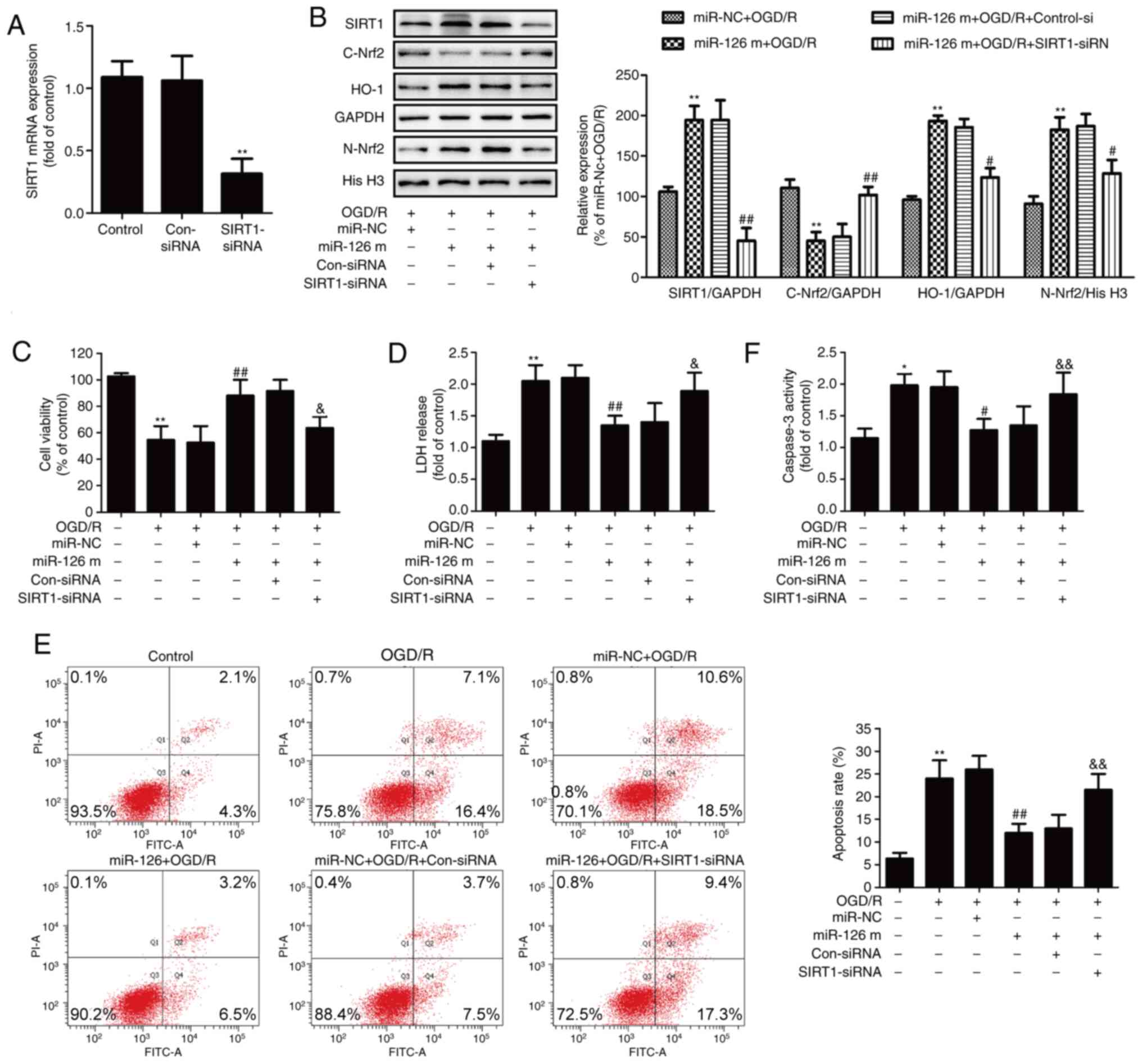 | Figure 3.Effects of SIRT1 siRNA on the
SIRT1/Nrf2 signaling pathway, cell injury and apoptosis in the
presence of miR-126 overexpression in OGD/R-exposed HUVECs. HUVECs
were pre-transfected with SIRT1-siRNA or Con-siRNA for 48 h, and
(A) the expression of SIRT1 mRNA was determined via reverse
transcription-quantitative PCR. HUVECs were pre-transfected with
SIRT1-siRNA or Con-siRNA for 30 min and then transfected with
miR-126 m or miR-NC before OGD (4 h)/R (24 h) exposure. (B) Protein
expression levels of SIRT1, nuclear Nrf2, cytosolic Nrf2 and HO-1
were determined via western blotting and bands were
semi-quantified. (C) Cell viability of each group was determined by
CCK-8 assay. (D) LDH release from HUVECs was measured using a LDH
Cytotoxicity assay kit. (E) The rate of apoptosis was detected by
Annexin V/PI double staining followed by flow cytometry. (F)
Caspase-3 activity was measured by Apo-ONE® Homogeneous
Caspase-3 assay kit. Data are presented as the mean ± SD of at
least three independent experiments. *P<0.05 and **P<0.01 vs.
control group; #P<0.05 and ##P<0.01 vs.
miR-NC + OGD/R group; &P<0.05 and
&&P<0.01 vs. miR-126 m + OGD/R + Con-siRNA
group. miR-126 m, miR-126 mimics; miR-NC, miR-negative control;
miR, microRNA; OGD, oxygen-glucose deprivation; R, reoxygenation;
HUVEC, human umbilical vein endothelial cell; SIRT1, NAD-dependent
protein deacetylase sirtuin-1; Nrf2, nuclear factor erythroid
2-related factor 2; siRNA, small interfering RNA; Con-siRNA,
negative control siRNA; LDH, lactate dehydrogenase. |
The results further revealed that SIRT1-siRNA
reversed the miR-126 mimics-induced increase in cell viability
(Fig. 3C) and the miR-126
mimics-induced decrease in LDH release in OGD/R-treated HUVECs
(Fig. 3D). Flow cytometry analysis
showed that SIRT1-siRNA also inhibited the miR-126 mimics-induced
reduction of apoptosis (Fig. 3E).
In addition, transfection with miR-126 mimics resulted in a
decrease in caspase-3 activity in OGD/R-treated HUVECs, whereas
this effect was reversed by SIRT1-siRNA (Fig. 3F). Overall, these results indicated
that the SIRT1/Nrf2 signaling pathway mediated the neuroprotective
effect of miR-126 on OGD/R injury.
miR-126 mimics mitigate OGD/R-induced
oxidative stress by potentiating the SIRT1/Nrf2 signaling pathway
in HUVECs
Increasing evidence has revealed that oxidative
stress is critical for I/R-induced neuronal injury, which
eventually results in apoptosis (3). Therefore, the effects of miR-126
mimics on oxidative stress in HUVECs exposed to OGD/R and the role
of the SIRT1/Nrf2 signaling pathway in these processes were further
investigated. DCFH-DA staining revealed that miR-126 mimics
inhibited the OGD/R-induced up-regulation of ROS generation in
HUVECs (Fig. 4A and B). However,
SIRT1-siRNA pretreatment reversed the effects of miR-126 mimic
transfection on ROS production compared with Con-siRNA transfection
in OGD/R-treated HUVECs. Similarly, compared with miR-NC
transfection, transfection with miR-126 mimics reduced the level of
intracellular MDA, a marker of lipid peroxidation, whereas this
effect of miR-126 mimics was attenuated by SIRT1-siRNA (Fig. 4C). To investigate the mechanisms
that may be involved in the protective effects of miR-126, the
activities of the antioxidant enzymes SOD and GSH-Px were measured.
Following transfection of HUVECs with miR-126 mimics, intracellular
SOD (Fig. 4D) and GSH-Px (Fig. 4E) activities were increased.
However, this antioxidant ability of miR-126 was reversed by
transfection with SIRT1-siRNA. These results suggested that miR-126
overexpression attenuated oxidative stress induced by OGD/R via the
up-regulation of the SIRT1/Nrf2 signaling pathway in HUVECs.
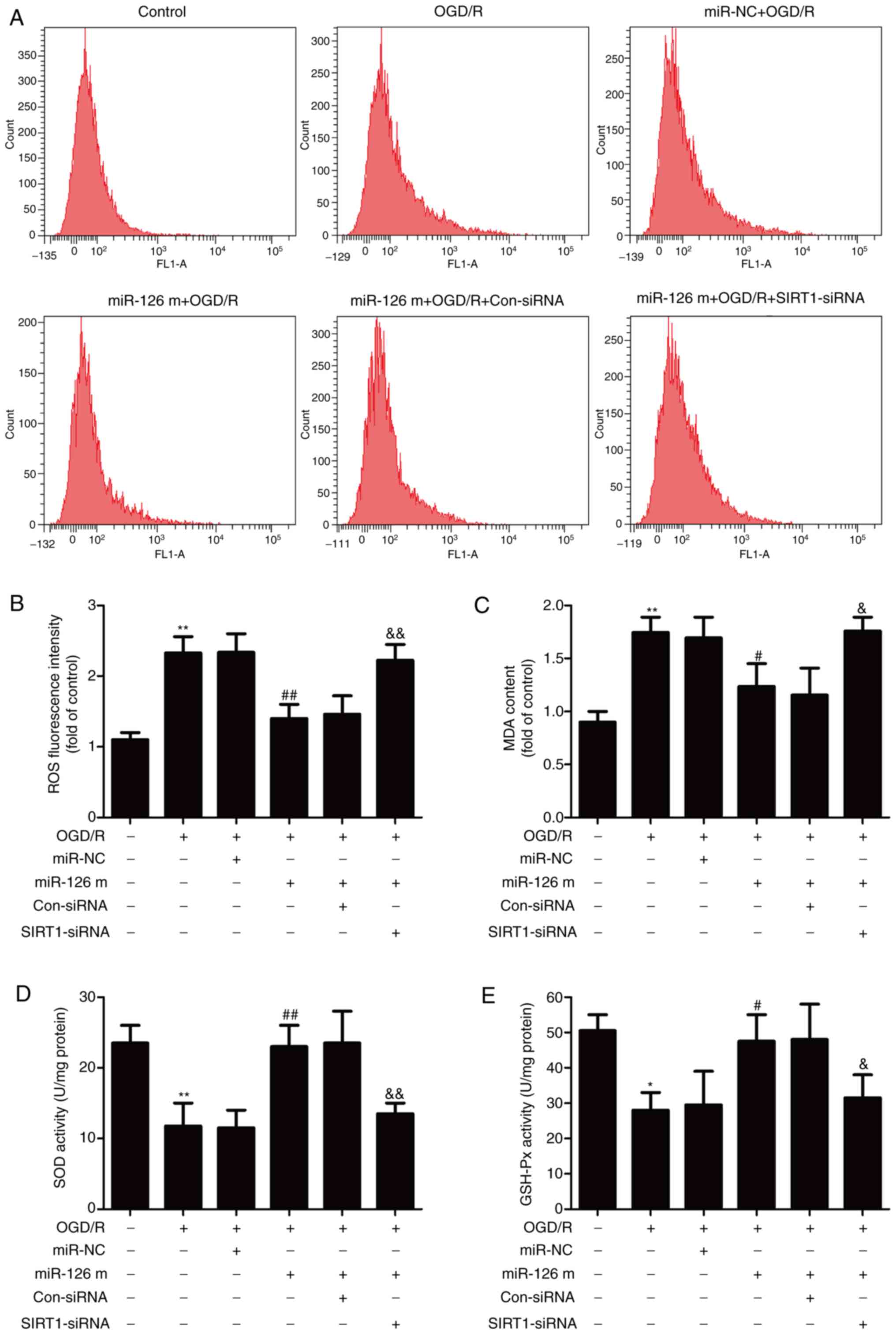 | Figure 4.Effects of miR-126 on oxidative
stress in HUVECs exposed to OGD/R in the presence or absence of
SIRT1 siRNA or Con-siRNA. HUVECs were pre-transfected with
SIRT1-siRNA or Con-siRNA for 30 min, and then transfected with
miR-126 m or miR-NC prior to OGD (4 h)/R (24 h) exposure. (A)
Intracellular ROS generation was assayed by
dichlorohydrofluorescein diacetate staining followed by flow
cytometry. (B) Quantitative analysis of ROS level. (C) MDA content
was measured using a Lipid Peroxidation MDA assay kit. (D) SOD
activity was determined by a Superoxide Dismutase assay kit. (E)
GSH-PX activity was detected using a Cellular Glutathione
Peroxidase assay kit. Data are presented as the mean ± SD of at
least three independent experiments. *P<0.05 and **P<0.01 vs.
control group; #P<0.05 and ##P<0.01 vs.
miR-NC + OGD/R group; &P<0.05 and
&&P<0.01 vs. miR-126 m + OGD/R + Con-siRNA
group. miR-126 m, miR-126 mimics; miR-NC, miR-negative control;
miR, microRNA; OGD, oxygen-glucose deprivation; R, reoxygenation;
HUVEC, human umbilical vein endothelial cell; SIRT1, NAD-dependent
protein deacetylase sirtuin-1; siRNA, small interfering RNA;
Con-siRNA, negative control siRNA; ROS, reactive oxygen species;
MDA, cell malondialdehyde; SOD, superoxide dismutase; GSH-Px,
glutathione peroxidase. |
miR-126 mimics suppress the
OGD/R-induced inflammatory response in a SIRT1-dependent
manner
To gain insight into the potential intracellular
mechanisms involved in the inhibitory effects of miR-126
overexpression on OGD/R injury, the levels of inflammatory-related
factors were investigated. As shown in Fig. 5, exposure to OGD/R resulted in the
up-regulation of pro-inflammatory cytokines, including TNF-α
(Fig. 5A), IL-1β (Fig. 5B) and IL-6 (Fig. 5C). However, transfection with
miR-126 mimics prevented the OGD/R-induced elevation of these
pro-inflammatory cytokines compared with miR-NC transfection.
Notably, the effect of transfection with miR-126 mimics was
reversed by transfection with SIRT1-siRNA compared with Con-siRNA
transfection. Additionally, transfection with miR-126 mimics led to
a marked inhibitory effect on the OGD/R-induced down-regulation of
anti-inflammatory cytokine IL-10 in HUVECs compared with miR-NC,
whereas this effect was attenuated by transfection with SIRT1-siRNA
(Fig. 5D). These results
demonstrated that miR-126 overexpression had anti-inflammatory
effects under OGD/R conditions via the promotion of the SIRT1/Nrf2
signaling pathway in HUVECs.
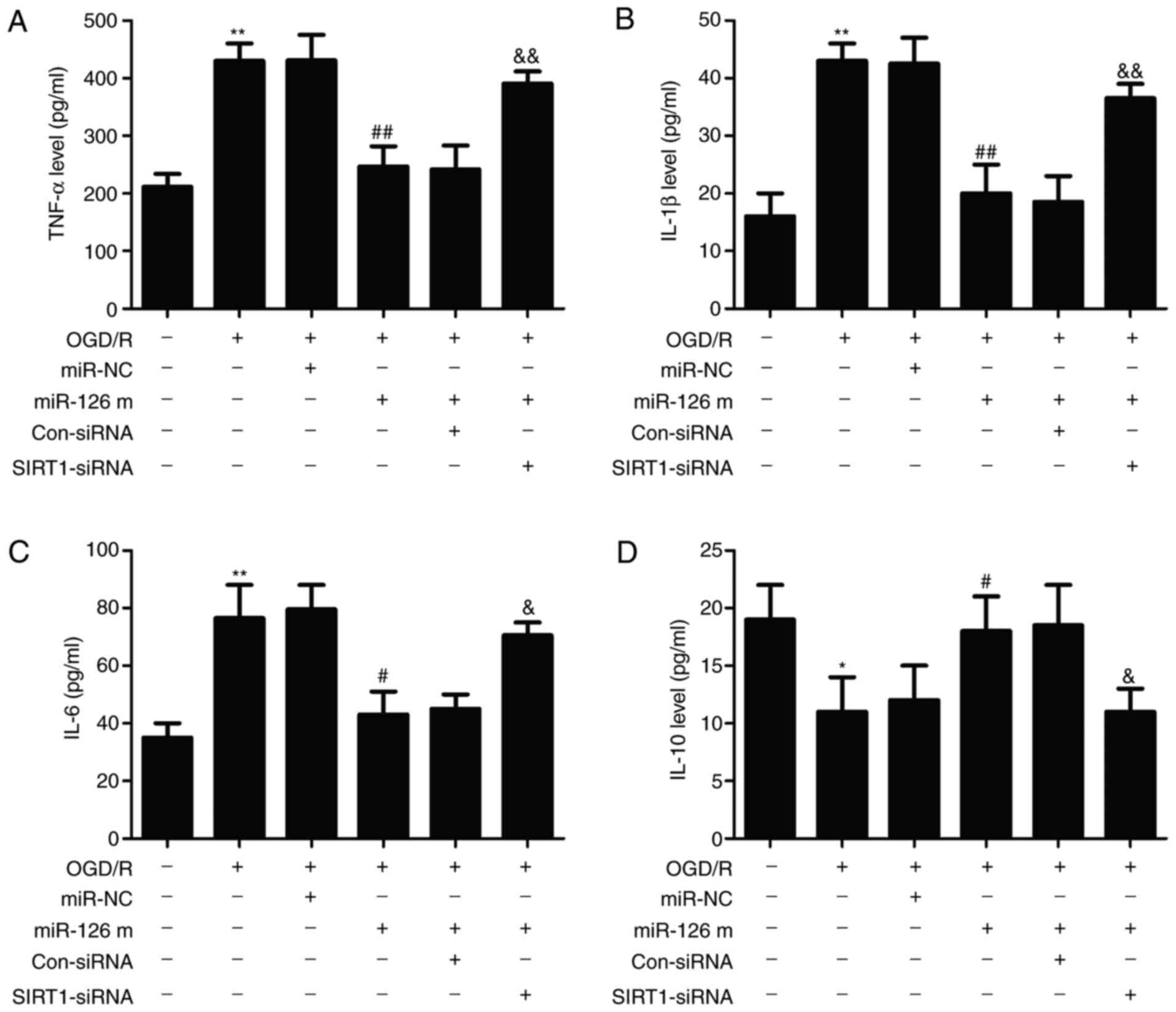 | Figure 5.Effects of miR-126 on the levels of
inflammatory-related cytokines in the presence of SIRT1-siRNA or
Con-siRNA in OGD/R-exposed HUVECs. HUVECs were pre-transfected with
SIRT1-siRNA or Con-siRNA for 30 min and then transfected with
miR-126 m or miR-NC prior to OGD (4 h)/R (24 h) exposure. The
levels of (A) TNF-α, (B) IL-1β, (C) IL-6 and (D) IL-10 were
measured using ELISA kits and the concentrations (µg/mg protein)
were calculated using a standard calibration curve. Data are
presented as the mean ± SD of at least three independent
experiments. *P<0.05 and **P<0.01 vs. control group;
#P<0.05 and ##P<0.01 vs. miR-NC + OGD/R
group; &P<0.05 and &&P<0.01
vs. miR-126 m + OGD/R + Con-siRNA group. miR-126 m, miR-126 mimics;
miR-NC, miR-negative control; miR, microRNA; OGD, oxygen-glucose
deprivation; R, reoxygenation; HUVEC, human umbilical vein
endothelial cell; SIRT1, NAD-dependent protein deacetylase
sirtuin-1; siRNA, small interfering RNA; Con-siRNA, negative
control siRNA; TNF-α, tumor necrosis factor-α; IL, interleukin. |
Discussion
The results of the present study demonstrated that
miR-126 expression was reduced in HUVECs exposed to OGD/R, and
miR-126 mimics reduced OGD/R-induced cytotoxicity and apoptosis.
Notably, miR-126 mimics activated the SIRT1/Nrf2 signaling pathway
under OGD/R conditions. Furthermore, the present study demonstrated
the protective effect of miR-126 overexpression against OGD/R
injury via promotion of the SIRT1/Nrf2 signaling pathway.
Furthermore, miR-126 attenuated OGD/R-induced oxidative stress and
inflammation by promoting the SIRT1/Nrf2 signaling pathway.
Therefore, these results indicated that overexpression of miR-126
had protective effects against OGD/R injury by inhibiting oxidative
stress and the inflammatory response by promoting the activation of
the SIRT1/Nrf2 signaling pathway in HUVECs.
According to previous studies, miR-126 is emerging
as a key player in the pathogenesis of ischemic stroke (11,25).
For instance, miR-126 expression is down-regulated following
cerebral ischemia, and miR-126 overexpression contributes to the
neurorestorative effects after stroke (26,27).
This was also observed in the present study. The results revealed
that OGD/R markedly reduced the expression levels of miR-126, and
overexpression of miR-126 induced by transfection with miR-126
mimics attenuated OGD/R-induced cytotoxicity and apoptosis in
HUVECs. Although, the present study did not investigate the effects
of miR-126 alone on cytotoxicity and apoptosis under normal
conditions, previous reports revealed that both miR-126 mimics and
miR-126 inhibitor have no effect on the activity and apoptosis of
HUVECs under normal conditions (28,29).
Taken together, these results indicated the protective effect of
miR-126 overexpression against I/R injury in the brain.
Previous studies have demonstrated that SIRT1 is
involved in the regulation of multiple cellular processes,
including cell survival, apoptosis, oxidative stress and
inflammation, and exerts important functions in ameliorating
cerebral I/R injury (30,31). As an important transcription factor,
Nrf2 serves as a downstream target of the SIRT1 signaling pathway
in increasing resistance to nerve damage (20,32). A
previous study revealed that SIRT1 served an important
physiological role during cerebral ischemia by activating Nrf2
(32). Notably, it was reported
that miR-126 regulated SIRT1 expression in a glucose intolerance
and peripheral artery disease mouse model (33). Consistent with these studies, the
present study revealed that miR-126 mimics increased the expression
of SIRT1 in OGD/R-treated HUVECs. However, it needs to be further
verified whether SIRT1 is a direct or indirect target of miR-126.
It is commonly known that once activated, Nrf2 translocates from
the cytoplasm to the nucleus, where this process can up-regulate
several antioxidant enzymes, particularly HO-1, resulting in
enhanced cell survival in numerous types of tissues (17). The results of the present study
further demonstrated that miR-126 mimics increased Nrf2 expression
in the nucleus, promoted HO-1 expression and decreased Nrf2
expression in the cytoplasm in OGD/R-treated HUVECs. These results
indicated that miR-126 overexpression promoted the activation of
the SIRT1/Nrf2 signaling pathway under I/R injury. Additionally,
the present results further demonstrated that inhibition of the
SIRT1/Nrf2 signaling pathway induced by transfection with
SIRT1-siRNA inhibited the protective effects of miR-126 mimics
against OGD/R injury in HUVECs. This was consistent with a previous
study that demonstrated that SIRT1 overexpression reduced
OGD/R-induced apoptosis, whereas SIRT1 knockdown increased
OGD/R-induced apoptosis (34).
Taken together, these results indicated that the SIRT1/Nrf2
signaling pathway served a significant role in the neuroprotective
effects of miR-126 against I/R injury.
Oxidative stress, which is primarily caused by
excess ROS, is widely involved in multiple pathophysiological
processes, including apoptosis, inflammation and nerve injury, and
is the core mechanism of neuronal damage following cerebral I/R
(35). Defensive antioxidants, such
as SOD and GSH-Px, can ameliorate the elevation of oxidants and
therefore protect brain tissues against ROS cytotoxicity (35). miR-126 has been demonstrated to
reduce oxidative stress and apoptosis in I/R injury (36). Similarly, the present study revealed
that miR-126 mimics reversed the OGD/R-induced increase in ROS
generation and MDA content (a major marker of lipid peroxidation
caused by oxidative stress), and the OGD/R-induced decrease in SOD
and GSH-Px activities in HUVECs. Thus, these results indicated that
miR-126 overexpression effectively prevented vascular I/R injury
caused by oxidative damage induced by ROS and the lipid
peroxidative effect. Notably, the SIRT1/Nrf2 signaling pathway was
revealed to serve a critical role in cellular antioxidant defense
mechanisms (20,37). It has been reported that
up-regulation of the SIRT and Nrf2 signaling pathway leads to
cellular defense mechanisms against oxidative stress injury
triggered by cerebral ischemia (32,38).
Consistent with these studies, the present results demonstrated
that SIRT1-siRNA-induced inhibition of the SIRT1/Nrf2 signaling
pathway blocked the anti-oxidative ability of miR-126 in
OGD/R-treated HUVECs. These effects indicated that miR-126
overexpression attenuated oxidative stress, resulting in protection
against vascular I/R injury in a SIRT1/Nrf2 signaling
pathway-dependent manner.
The inflammatory response is important in the
progression of cerebral ischemia (39). A number of studies have demonstrated
that, during I/R, there is a marked elevation in the levels of
pro-inflammatory mediators, including TNF-α, IL-1β and IL-6, and a
marked reduction in the levels of anti-inflammatory cytokines,
including IL-4 and IL-10 in the brain (40,41).
The present results also revealed that OGD/R treatment increased
the levels of TNF-α, IL-1β and IL-6, and decreased the level of
IL-10 in HUVECs. Growing evidence has suggested that, in addition
to promoting the neuroprotective effect, miR-126 also has
anti-inflammatory effects (42,43).
However, to the best of our knowledge, the anti-inflammatory
effects of miR-126 in I/R injury in HUVECs have not yet been
investigated. The results of the present study found that miR-126
mimics reduced the OGD/R-induced inflammatory response. In
addition, numerous studies have reported that the SIRT1/Nrf2
signaling pathway is a promising anti-inflammatory pathway target
to reduce nerve injury (21,44).
Furthermore, the present results indicated that inhibition of the
SIRT1/Nrf2 signaling pathway reversed the inhibitory effect of
miR-126 mimics on the OGD/R-induced inflammatory response. These
results suggested that the SIRT1/Nrf2 signaling pathway mediated
the anti-inflammatory action of miR-126 under I/R conditions in
HUVECs.
However, there are a few limitations of the present
study. It is unclear whether miR-126 directly targets SIRT1/Nrf2
signaling molecules, which requires further investigation. In
addition, in vivo experiments and studies using cell lines
derived from the brain are required to elucidate the specific
mechanisms involved in the neuroprotection of miR-126 against
cerebral I/R injury.
In conclusion, the present findings demonstrated
that the amelioration of I/R injury induced by miR-126
overexpression could be the result of its anti-oxidative and
anti-inflammatory effects in HUVECs. Notably, the SIRT1/Nrf2
signaling pathway was identified to contribute to the
neuroprotective effects of miR-126 overexpression.
Acknowledgements
Not applicable.
Funding
No funding was received.
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
JL designed and performed the experiments and wrote
and revised the manuscript. CY performed the experiments and
analyzed data. YW conceived the study and corrected the manuscript.
All authors read and approved the final manuscript.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Broussalis E, Killer M, McCoy M, Harrer A,
Trinka E and Kraus J: Current therapies in ischemic stroke. Part A.
Recent developments in acute stroke treatment and in stroke
prevention. Drug Discov Today. 17:296–309. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Correction to: Guidelines for the early
management of patients with acute ischemic stroke: 2019 update to
the 2018 guidelines for the early management of acute ischemic
stroke: A guideline for healthcare professionals from the American
heart association/American stroke association. Stroke.
50:e440–e441. 2019.PubMed/NCBI
|
|
3
|
Galkin A: Brain ischemia/reperfusion
injury and mitochondrial complex I damage. Biochemistry (Mosc).
84:1411–1423. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
LeBaron TW, Kura B, Kalocayova B,
Tribulova N and Slezak J: A new approach for the prevention and
treatment of cardiovascular disorders. Molecular hydrogen
significantly reduces the effects of oxidative stress. Molecules.
24:20762019. View Article : Google Scholar
|
|
5
|
Enzmann G, Kargaran S and Engelhardt B:
Ischemia-reperfusion injury in stroke: Impact of the brain barriers
and brain immune privilege on neutrophil function. Ther Adv Neurol
Disord. Aug 18–2018.(Epub ahead of print). doi:
10.1177/1756286418794184. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Wen Z, Hou W, Wu W, Zhao Y, Dong X, Bai X,
Peng L and Song L: 6′-O-Galloylpaeoniflorin attenuates cerebral
ischemia reperfusion-induced neuroinflammation and oxidative stress
via PI3K/Akt/Nrf2 activation. Oxid Med Cell Longev.
2018:86782672018. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Chamorro A, Dirnagl U, Urra X and Planas
AM: Neuroprotection in acute stroke: Targeting excitotoxicity,
oxidative and nitrosative stress, and inflammation. Lancet Neurol.
15:869–881. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Yang J, Chen M, Cao RY, Li Q and Zhu F:
The role of circular RNAs in cerebral ischemic diseases: Ischemic
stroke and cerebral ischemia/reperfusion injury. Adv Exp Med Biol.
1087:309–325. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Dewdney B, Trollope A, Moxon J, Thomas
Manapurathe D, Biros E and Golledge J: Circulating MicroRNAs as
biomarkers for acute ischemic stroke: A systematic review. J Stroke
Cerebrovasc Dis. 27:522–530. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Gai HY, Wu C, Zhang Y and Wang D: Long
non-coding RNA CHRF modulates the progression of cerebral
ischemia/reperfusion injury via miR-126/SOX6 signaling pathway.
Biochem Biophys Res Commun. 514:550–557. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Chen J, Cui C, Yang X, Xu J, Venkat P,
Zacharek A, Yu P and Chopp M: MiR-126 affects brain-heart
interaction after cerebral ischemic stroke. Transl Stroke Res.
8:374–385. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Pan Q, Zheng J, Du D, Liao X, Ma C, Yang
Y, Chen Y, Zhong W and Ma X: MicroRNA-126 priming enhances
functions of endothelial progenitor cells under physiological and
hypoxic conditions and their therapeutic efficacy in cerebral
ischemic damage. Stem Cells Int. 2018:29123472018. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Ren Z, He H, Zuo Z, Xu Z, Wei Z and Deng
J: The role of different SIRT1-mediated signaling pathways in toxic
injury. Cell Mol Biol Lett. 24:362019. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Kitada M, Ogura Y, Monno I and Koya D:
Sirtuins and type 2 diabetes: Role in inflammation, oxidative
stress, and mitochondrial function. Front Endocrinol (Lausanne).
10:1872019. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Hattori Y and Ihara M: Sirt1. Nihon
Rinsho. 74:589–594. 2016.(In Japanese). PubMed/NCBI
|
|
16
|
Meng X, Tan J, Li M, Song S, Miao Y and
Zhang Q: Sirt1: Role under the condition of ischemia/hypoxia. Cell
Mol Neurobiol. 37:17–28. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Loboda A, Damulewicz M, Pyza E, Jozkowicz
A and Dulak J: Role of Nrf2/HO-1 system in development, oxidative
stress response and diseases: An evolutionarily conserved
mechanism. Cell Mol Life Sci. 73:3221–3247. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Ma R, Liang W, Sun Q, Qiu X, Lin Y, Ge X,
Jueraitetibaike K, Xie M, Zhou J, Huang X, et al: Sirt1/Nrf2
pathway is involved in oocyte aging by regulating Cyclin B1. Aging
(Albany NY). 10:2991–3004. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Arioz BI, Tastan B, Tarakcioglu E, Tufekci
KU, Olcum M, Ersoy N, Bagriyanik A, Genc K and Genc S: Melatonin
attenuates LPS-induced acute depressive-like behaviors and
microglial NLRP3 inflammasome activation through the SIRT1/Nrf2
pathway. Front Immunol. 10:15112019. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Shah SA, Khan M, Jo MH, Jo MG, Amin FU and
Kim MO: Melatonin stimulates the SIRT1/Nrf2 signaling pathway
counteracting lipopolysaccharide (LPS)-induced oxidative stress to
rescue postnatal rat brain. CNS Neurosci Ther. 23:33–44. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Nagib MM, Tadros MG, Al-Khalek HAA, Rahmo
RM, Sabri NA, Khalifa AE and Masoud SI: Molecular mechanisms of
neuroprotective effect of adjuvant therapy with phenytoin in
pentylenetetrazole-induced seizures: Impact on Sirt1/NRF2 signaling
pathways. Neurotoxicology. 68:47–65. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Chan YH, Liu TC, Liao CK, Cheng YF, Tsai
CH, Lu YC, Hu CJ, Lin HJ, Lee YL, Wu CC and Hsu CJ: Consumption of
betel quid contributes to sensorineural hearing impairment through
arecoline-induced oxidative stress. Sci Rep. 9:145542019.
View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Xu JQ, Liu P, Si MJ and Ding XY:
MicroRNA-126 inhibits osteosarcoma cells proliferation by targeting
Sirt1. Tumour Biol. 34:3871–3877. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Wang JQ, Gu WP, Deng QQ, Huang Q, Wang AM,
Liu SX, Tang HY, Liang Y, Yan JH and Ouyang S: Endothelial
progenitor cell miR-126 promotes homing of endothelial progenitor
cells within arterial thrombus in patients with cerebral infarction
and its molecular mechanism. Eur Rev Med Pharmacol Sci.
22:1078–1083. 2018.PubMed/NCBI
|
|
26
|
Geng W, Tang H, Luo S, Lv Y, Liang D, Kang
X and Hong W: Exosomes from miRNA-126-modified ADSCs promotes
functional recovery after stroke in rats by improving neurogenesis
and suppressing microglia activation. Am J Transl Res. 11:780–792.
2019.PubMed/NCBI
|
|
27
|
Chen J, Ning R, Zacharek A, Cui C, Cui X,
Yan T, Venkat P, Zhang Y and Chopp M: MiR-126 contributes to human
umbilical cord blood cell-induced neurorestorative effects after
stroke in type-2 diabetic mice. Stem Cells. 34:102–113. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Ge HY, Han ZJ, Tian P, Sun WJ, Xue DX, Bi
Y, Yang ZH and Liu P: VEGFA expression is inhibited by arsenic
trioxide in HUVECs through the upregulation of Ets-2 and miRNA-126.
PLoS One. 10:e01357952015. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Tang F and Yang TL: MicroRNA-126
alleviates endothelial cells injury in atherosclerosis by restoring
autophagic flux via inhibiting of PI3K/Akt/mTOR pathway. Biochem
Biophys Res Commun. 495:1482–1489. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
He Q, Li Z, Wang Y, Hou Y, Li L and Zhao
J: Resveratrol alleviates cerebral ischemia/reperfusion injury in
rats by inhibiting NLRP3 inflammasome activation through
Sirt1-dependent autophagy induction. Int Immunopharmacol.
50:208–215. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Koronowski KB and Perez-Pinzon MA: Sirt1
in cerebral ischemia. Brain Circ. 1:69–78. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Xue F, Huang JW, Ding PY, Zang HG, Kou ZJ,
Li T, Fan J, Peng ZW and Yan WJ: Nrf2/antioxidant defense pathway
is involved in the neuroprotective effects of Sirt1 against focal
cerebral ischemia in rats after hyperbaric oxygen preconditioning.
Behav Brain Res. 309:1–8. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Togliatto G, Trombetta A, Dentelli P,
Gallo S, Rosso A, Cotogni P, Granata R, Falcioni R, Delale T, Ghigo
E and Brizzi MF: Unacylated ghrelin induces oxidative stress
resistance in a glucose intolerance and peripheral artery disease
mouse model by restoring endothelial cell miR-126 expression.
Diabetes. 64:1370–1382. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Ren Q, Hu Z, Jiang Y, Tan X, Botchway BOA,
Amin N, Lin G, Geng Y and Fang M: SIRT1 protects against apoptosis
by promoting autophagy in the oxygen glucose
deprivation/reperfusion-induced injury. Front Neurol. 10:12892019.
View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Sun MS, Jin H, Sun X, Huang S, Zhang FL,
Guo ZN and Yang Y: Free radical damage in ischemia-reperfusion
injury: An obstacle in acute ischemic stroke after
revascularization therapy. Oxid Med Cell Longev. 2018:38049792018.
View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Wang W, Zheng Y, Wang M, Yan M, Jiang J
and Li Z: Exosomes derived miR-126 attenuates oxidative stress and
apoptosis from ischemia and reperfusion injury by targeting ERRFI1.
Gene. 690:75–80. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Han J, Liu X, Li Y, Zhang J and Yu H:
Sirt1/Nrf2 signalling pathway prevents cognitive impairment in
diabetic rats through anti-oxidative stress induced by miRNA-23b-3p
expression. Mol Med Rep. 17:8414–8422. 2018.PubMed/NCBI
|
|
38
|
Tao X, Sun X, Xu L, Yin L, Han X, Qi Y, Xu
Y, Zhao Y, Wang C and Peng J: Total flavonoids from rosa laevigata
michx fruit ameliorates hepatic ischemia/reperfusion injury through
inhibition of oxidative stress and inflammation in rats. Nutrients.
8:4182016. View Article : Google Scholar
|
|
39
|
Anrather J and Iadecola C: Inflammation
and stroke: An overview. Neurotherapeutics. 13:661–670. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Cao XL, Du J, Zhang Y, Yan JT and Hu XM:
Hyperlipidemia exacerbates cerebral injury through oxidative
stress, inflammation and neuronal apoptosis in MCAO/reperfusion
rats. Exp Brain Res. 233:2753–2765. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Tobin MK, Bonds JA, Minshall RD,
Pelligrino DA, Testai FD and Lazarov O: Neurogenesis and
inflammation after ischemic stroke: What is known and where we go
from here. J Cereb Blood Flow Metab. 34:1573–1584. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Zhang Q, Zeng S, Quan C and Lin X:
Induction function of miR-126 in survival and proliferation in
neural stem cells. Med Sci Monit. 21:3023–3027. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Hu J, Zeng L, Huang J, Wang G and Lu H:
miR-126 promotes angiogenesis and attenuates inflammation after
contusion spinal cord injury in rats. Brain Res. 1608:191–202.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Lee JY, Song H, Dash O, Park M, Shin NE,
McLane MW, Lei J, Hwang JY and Burd I: Administration of melatonin
for prevention of preterm birth and fetal brain injury associated
with premature birth in a mouse model. Am J Reprod Immunol.
82:e131512019. View Article : Google Scholar : PubMed/NCBI
|














