Introduction
Colorectal cancer (CRC) is one of the most common
types of malignant cancer of the digestive system worldwide,
displaying high incidence and mortality rates (1,2). In
2020, the estimated new cases of colon cancer in the United States
are 78,300 in men, and 69,650 in women, accounting for 8–9% of all
cancer cases (2). Currently, the
standard therapeutic strategy used for CRC is surgical resection
combined with radio-chemotherapy (3); however, recurrence following curative
resection is a major issue (4).
Additionally, treating CRC using chemotherapy may result in
cytotoxic side effects and the development of resistance,
highlighting the importance of the development of more effective
and novel therapeutics and adjuvants (5,6).
Melatonin has been highlighted as a potential alternative
therapeutic due to its low toxicity and high efficacy (7).
Melatonin (N-acetyl-5-methoxytryptamine) is a pineal
hormone and its release is regulated by the light/dark cycle
(8). In addition to its presence in
the pineal gland, melatonin also exists in other tissues, such as
the liver and gastrointestinal tract (9,10).
Melatonin regulates numerous physiological functions, including the
circadian rhythm (11),
antioxidative protection, and anti-inflammatory (12,13)
and antioncogenic activities (14,15).
In particular, melatonin displays a range of protective effects in
the gastrointestinal tract, such as enhancing the immune functions
of the gut (10) and altering the
composition of the intestinal microbiota (16,17).
Moreover, melatonin may inhibit the development and progression of
gastrointestinal cancer (18,19).
Previous studies have demonstrated that the expression levels of
melatonin in the blood of patients with CRC are disordered, which
may increase the incidence of CRC (20,21).
Riabykh et al (22), Farriol
et al (23) and Anisimov
et al (24) reported that
melatonin inhibited CRC cell proliferation via decreasing DNA
synthesis and promoting cell differentiation. Hong et al
(25) and León et al
(26) reported that melatonin
induced CRC cell apoptosis by altering cell cycle progression or
reducing endothelin-1 expression. Although the mechanisms
underlying melatonin-mediated inhibition of proliferation and
induction of apoptosis in CRC cells have been studied, the
identified mechanisms cannot fully explain the breadth of antitumor
effects mediated by melatonin.
Increasing evidence has demonstrated that
dysregulated expression of microRNAs (miRs/miRNAs) serves critical
roles in the tumorigenesis of several types of cancer (27–31).
Highly expressed miR-21 and miR-155 are associated with the
metastasis and poor prognosis of colon cancer (30). miR-195 functions as a tumor
suppressor in colon cancer (31).
miRNAs are single-stranded non-coding RNAs 18–24 nucleotides in
length (32). miRNAs affect the
stability of mRNA transcription and protein translation by
targeting the 3′-untranslated region (UTR) of their target mRNAs
(33–35). miRNAs may serve as oncogenes or
tumor suppressors, and are involved in cancer development and
numerous cellular processes (36–38).
miRNAs frequently reside in clusters that include >2–3 miRNA
genes with pairwise chromosomal distances of ≤3,000 nucleotides in
the genome (39). Members of miRNA
clusters are generally similar in sequence and are transcribed in
the same direction (40). miRNAs
are highly conserved and usually function synergistically (41,42).
Notably, miRNAs are involved in melatonin-induced
inhibition of proliferation in gastric cancer, prostate cancer and
gliomas (43–45); however, the functions and mechanisms
underlying melatonin and miRNAs that are involved in CRC are not
completely understood. The present study examined the effects of
melatonin on cell proliferation and apoptosis in CRC in
vitro and also investigated whether melatonin could inhibit the
tumor growth of tumor-bearing nude mice in vivo. In
addition, it investigated the relationship between miRNAs and
melatonin in CRC.
Materials and methods
Cell culture
HCT116, LoVo, SW480, SW620, HT-29 and DLD-1 human
CRC cell lines were purchased from ATCC. All cells were cultured in
DMEM (HyClone; Cytiva) supplemented with 10% FBS (Shanghai ExCell
Biology, Inc.) and 1% penicillin-streptomycin-amphotericin B
[MACGENE (Beijing) Biotechnology Ltd.] at 37°C in a humidified
incubator with 5% CO2.
Cell viability assay
Cell viability was assessed by performing MTT
(Sigma-Aldrich; Merck KGaA) and Cell Counting Kit-8 (CCK-8; APExBIO
Technology LLC) assays. Briefly, HCT116, LoVo, SW480, SW620, HT-29
and DLD-1 human CRC cell lines were seeded (8×103
cells/well) into 96-well plates and treated with melatonin (0.1–2.0
mM; cat. no. M5250-1G; Sigma-Aldrich; Merck KGaA) according to
previously described protocols (19,46–49).
Subsequently, cell viability was detected by performing MTT and
CCK-8 assays. For MTT assay, the plate was incubated for an
additional 4 h at 37°C, and crystal formazan was dissolved in DMSO
(0.2 ml/well). The absorbance of each well was measured using a
microplate reader at wavelengths of 570 nm. For the CCK-8 assay,
the 96-well plates were incubated for 4 h at 37°C, and the
absorbencies at each time point were measured at 450 nm by a
microplate reader. Melatonin at 2 mM was selected for subsequent
experiments, as this concentration significantly reduced cell
viability compared with other concentrations.
Colony formation assay
HCT116 and LoVo CRC cells were pretreated with 2 mM
melatonin for 48 h at 37°C. Subsequently, cells were seeded
(1×103 cells/well) into a 6-well plate. After 7 days,
the colonies were fixed with 4% paraformaldehyde for 30 min at room
temperature and then stained with 0.1% crystal violet (Beijing
Solarbio Science & Technology Co., Ltd.) for 20 min at room
temperature. Colonies were observed with scanner (50,51).
Colonies containing >50 cells were counted.
EdU labelling and staining
HCT116 and LoVo CRC cells (5×105
cells/well) were plated into 96-well plates. EdU (50 µM) from the
Cell-Light™ EdU Apollo488 kit (Guangzhou RiboBio Co., Ltd.) was
used for labelling for 24 h at 37°C. Following fixation with 4%
paraformaldehyde for 30 min at room temperature, cells were stained
with Apollo and Hoechst 33342 for 30 min at room temperature in the
dark. Stained cells were visualized using a DMI6000 B inverted
fluorescence microscope (Leica Microsystems GmbH). The number of
stained cells was determined according to the manufacturer's
protocol.
Cell apoptosis analysis
HCT116 and LoVo CRC cells were pretreated with 2 mM
melatonin for 48 h at 37°C. The apoptosis rate was evaluated using
the Annexin V-FITC/PI Apoptosis Detection kit (C1062; Beyotime
Institute of Biotechnology) according to the instructions from the
manufacturer. Following treatment, the cells were collected, washed
with PBS, and resuspended in 195 µl binding buffer. Then, 5 µl
Annexin V-FITC and 10 µl PI were added to the buffer and incubated
at room temperature for 20 min in the dark. Early and late
apoptotic cells were analyzed using a CytoFLEX S flow cytometer and
CytExpert 2.3 software (Beckman Coulter, Inc.).
Cell transfection
HCT116 and LoVo CRC cells were seeded at
5×105 cells/well in 6-well plates. The cells were
transfected using 5 µl Lipofectamine® 2000 RNAiMAX
(Invitrogen; Thermo Fisher Scientific, Inc.) for 20 min at room
temperature with a mixture comprising 500 µl FBS-free medium with
10 µl miR-34a inhibitor (MIMAT0000255; 100 nM; Guangzhou RiboBio
Co., Ltd.) or 10 µl of miR-449a inhibitor (MIMAT0001541; 100 nM;
Guangzhou RiboBio Co., Ltd.) or the negative control (Guangzhou
RiboBio Co., Ltd.) according to the manufacturer's instructions.
The mixture was added to each well for 6 h at 37°C. The mixture was
changed for fresh medium after 6 h. Cells were treated with
melatonin after transfection for 24 h.
RNA extraction and reverse
transcription-quantitative PCR (RT-qPCR)
Total RNA was extracted from the HCT116 and LoVo CRC
cells using TRIzol® reagent (Thermo Fisher Scientific,
Inc.). cDNA was synthesized with a RevertAid™ First Strand cDNA
Synthesis kit (Thermo Fisher Scientific, Inc.) with the following
thermocycling conditions: 42°C for 1 h; followed by 75°C for 5 min
and 4°C forever. Subsequently, qPCR was performed to determine the
expression levels of hsa-miR-125a, hsa-miR-365a, hsa-miR-34a,
hsa-miR-449a, hsa-miR-184, hsa-miR-143a, Bcl-2 and Notch1. qPCR was
performed with SYBR-Green Realtime PCR Master Mix (Toyobo Life
Science) on a CFX96™ Real-Time System (Bio-Rad Laboratories, Inc.)
with the following thermocycling conditions: 95°C for 10 min;
followed by 40 cycles at 95°C for 15 sec and 60°C for 60 sec. The
sequences of the primers used for qPCR are presented in Table SI. miRNA and mRNA expression levels
were quantified using the 2−ΔΔCq method and normalized
to the internal reference genes U6 and actin, respectively
(52).
Western blotting
HCT116 and LoVo CRC cells were lysed in RIPA buffer
(Beyotime Institute of Biotechnology) to obtain total protein, and
total protein concentration was quantified using a BCA protein
assay kit (Beyotime Institute of Biotechnology). An equal amount of
total protein (30 µg) was subjected to 10% (w/v) SDS-PAGE gel and
transferred onto PVDF membranes (EMD Millipore). These membranes
were blocked with 5% skimmed milk for 2 h at room temperature.
Subsequently, the membranes were incubated overnight at 4°C with
the primary antibodies targeted against: cleaved caspase-3
(1:1,000; cat. no. 9661; Cell Signaling Technology, Inc.), Notch1
(1:1,000; cat. no. 3447; Cell Signaling Technology, Inc.), cleaved
poly (ADP-ribose) polymerase 1 (PARP; 1:1,000; cat. no. ab203467;
Abcam), p53 (1:1,000; cat. no. 10442-1-AP; ProteinTech Group,
Inc.), Bcl-2 (1:1,000; cat. no. sc-7382; Santa Cruz Biotechnology,
Inc.) and β-actin (1:1,000; cat. no. HC201; TransGen Biotech Co.,
Ltd.). After washing, the membranes were incubated with Goat
Anti-Mouse IgG (H+L) HRP-conjugated (1:5,000; cat. no. HS201-01;
TransGen Biotech Co., Ltd.), Goat Anti-Rabbit IgG (H+L)
HRP-conjugated (1:5,000; cat. no. HS101-01; TransGen Biotech Co.,
Ltd.) secondary antibodies and Goat Anti-Rat IgG (H+L) Secondary
Antibody, HRP (1:5,000; cat. no. 31470; Thermo Fisher Scientific,
Inc.) for 1 h at room temperature. Protein bands were visualized
using Immobilon™ Western Chemiluminescent Horseradish Peroxidase
Substrate (EMDMillipore). Protein expression levels were
semi-quantified using ImageJ software (v1.46, National Institutes
of Health) with β-actin as the loading control.
Luciferase reporter assay
Human Bcl-2 and Notch1 3′UTR regions containing the
putative binding sites of miR-34a/449a were subcloned into the pGL3
promoter (Promega Corporation) using XbaI restriction digest sites
at the end of the oligonucleotides. HCT116 CRC cells
(1.5×104 cells/well) were seeded into a 24-well plate in
triplicate. CRC cells were co-transfected with 400 ng
pGL3-3′UTR-Promoter plasmid, 100 ng Renilla (Promega
Corporation) and 100 nM miR-34a or miR-449a inhibitor using
Lipofectamine 2000 (Invitrogen; Thermo Fisher Scientific, Inc.). At
48 h post-transfection, luciferase activities were determined using
a Dual Luciferase Reporter assay kit (Promega Corporation). Firefly
luciferase activities were normalized to Renilla luciferase
activities.
Tumorigenesis in xenograft models
Female BALB/c Nude mice (age, 4–6 weeks; weight,
20±2 g; n=12) were purchased from GemPharmatech Co., Ltd. All
animals were housed in cages at 24°C with 12-h light/dark cycles,
50–70% humidity, and free access to food and water. All experiments
were approved by the Ethics Committee on Animal Experiments of the
Medical School of Shandong University [approval no.
KYLL-2017(KS)-357]. Nude mice were injected subcutaneously into the
right shoulder with 6×106 HCT 116 cells suspended in 150
µl PBS. When tumors reached an average volume of 50 mm3,
the mice were randomly divided into two groups (n=6/group): i)
Control group (same volume of PBS as the melatonin group) and ii)
melatonin group (25 mg/kg; intraperitoneal injection daily for 23
days). The 25 mg/kg melatonin was selected for xenograft assays
according to previous studies (53–55).
The tumors were isolated, measured and histologically confirmed.
The perpendicular tumor dimensions (a, length and b, width) were
measured using Vernier caliper to calculate the volume (V;
mm3) according to the following formula: V=(a ×
b2)/2. Mice were anesthetized with an intraperitoneal
injection of 50 mg/kg sodium pentobarbital (1%). The tumors were
removed and the tissue specimens were used for immunofluorescence
analysis. After sampling, the mice were euthanized with an overdose
of sodium pentobarbital (100 mg/kg) (55).
HE staining
The tumor sections were cut at a thickness of 4 µm
using a microtome. The sections were dewaxed using xylene and
rehydrated in ethanol with gradient concentrations (100, 95, 90,
80, 70, 50 and 30%) for 5 min at room temperature. After being
washed with distilled water for 5 min, Hematoxylin (cat. no. C0107;
Beyotime Institute of Biotechnology) was added dropwise to stain
the tissue for 15 min. The stained tissue was placed in 1%
hydrochloric acid ethanol (Merck KGaA) for differentiation for 10
sec. Then, 0.25% Eosin dye (cat. no. C0109; Beyotime Institute of
Biotechnology) solution was added for counterstaining for 5 min and
tissues were dehydrated by soaking in gradient concentration
alcohol (70, 80, 90, 95 and 100% ethanol) for 5 min. All these
steps were performed at room temperature. The sections were
observed (magnification, ×20) and images captured under the optical
microscope. Each section was divided into three columns, and three
images captured of each column, nine images per section.
Immunofluorescence staining
Cancer tissues obtained from the mouse xenograft
model were fixed using 4% paraformaldehyde for 48 h at 4°C. The
tumor sections were cut at a thickness of 10 µm using a freezing
microtome. The sections were blocked and permeabilized in 5% goat
serum (cat. no. ZLI-9021; OriGene Technologies, Inc.) solution
containing 0.3% Triton X-100 for 2 h at room temperature. The
sections were incubated with a rabbit anti-Ki-67 primary antibody
(1:400; cat. no. ab16667; Abcam) overnight at 4°C. The sections
were washed in PBS three times and incubated with Dylight 594, Goat
Anti-Rabbit IgG (1:1,000; cat. no. A23420; Abbkine Scientific Co.,
Ltd.) secondary antibodies for 1 h at room temperature. TUNEL assay
was performed using the TransDetect® in situ
Fluorescein TUNEL Cell Apoptosis Detection kit (cat. no. FA201;
TRANS). Briefly, the sections were blocked and permeabilized in 5%
goat serum solution containing 0.3% Triton X-100 for 2 h at room
temperature. The sections were incubated with a mix of 1X labeling
solution and TdT (25:1) for 1 h at 37°C in the dark. Nuclei were
stained with DAPI (Sigma-Aldrich; Merck KGaA) for 15 min at room
temperature. Stained samples were observed (TUNEL magnification,
×20; Ki-67 magnification, ×10) using a confocal microscope (Leica
Microsystems GmbH). Each section was divided into three columns,
and three images captured of each column, nine images per
section.
Bioinformatics analysis
The sequences of the microRNAs were searched through
miRBase (http://www.mirbase.org/). The downstream
target genes of the microRNAs cluster were predicted with
TargetScan (http://www.targetscan.org/vert_71/).
Statistical analyses
Data are presented as the mean ± standard deviation
of at least three independent experiments. Statistical analyses
were performed using GraphPad Prism software (version 8; GraphPad
Software, Inc.). Comparisons between two groups were analyzed using
the unpaired Student's t-test. Comparisons among multiple groups
were analyzed using one-way ANOVA followed by Dunnett's post hoc
test. P<0.05 was considered to indicate a statistically
significant difference.
Results
Melatonin suppresses CRC cell
proliferation and viability
The effect of melatonin on CRC cell viability
(HCT116, LoVo, SW480, SW620, HT-29 and DLD-1) following treatment
for 24 or 48 h was assessed by performing an MTT assay. The results
indicated that melatonin (0.1–2.0 mM) significantly inhibited CRC
cell viability in a time- and dose-dependent manner. Compared with
the control group, SW480 and SW620 cell viability was inhibited by
treatment with 1 mM melatonin for 48 h, whereas DLD-1 cell
viability was inhibited by treatment with 2 mM melatonin for 48 h.
HCT116 and LoVo cell viability were significantly reduced compared
with SW480, SW620, HT-29 and DLD-1 CRC cell lines at 24 and 48 h
(Fig. 1A). The effect of melatonin
on HCT116 and LoVo cell proliferation was assessed by performing
EdU staining and counting the number of EdU+ cells
(Fig. 1B and C). Compared with the
control group, the number of EdU+ cells in the melatonin
group was significantly reduced. The colony formation ability of
HCT116 and LoVo cells following treatment with melatonin was also
assessed (Fig. 1D and E). The
number of colonies in the melatonin group was also significantly
reduced compared with the control group.
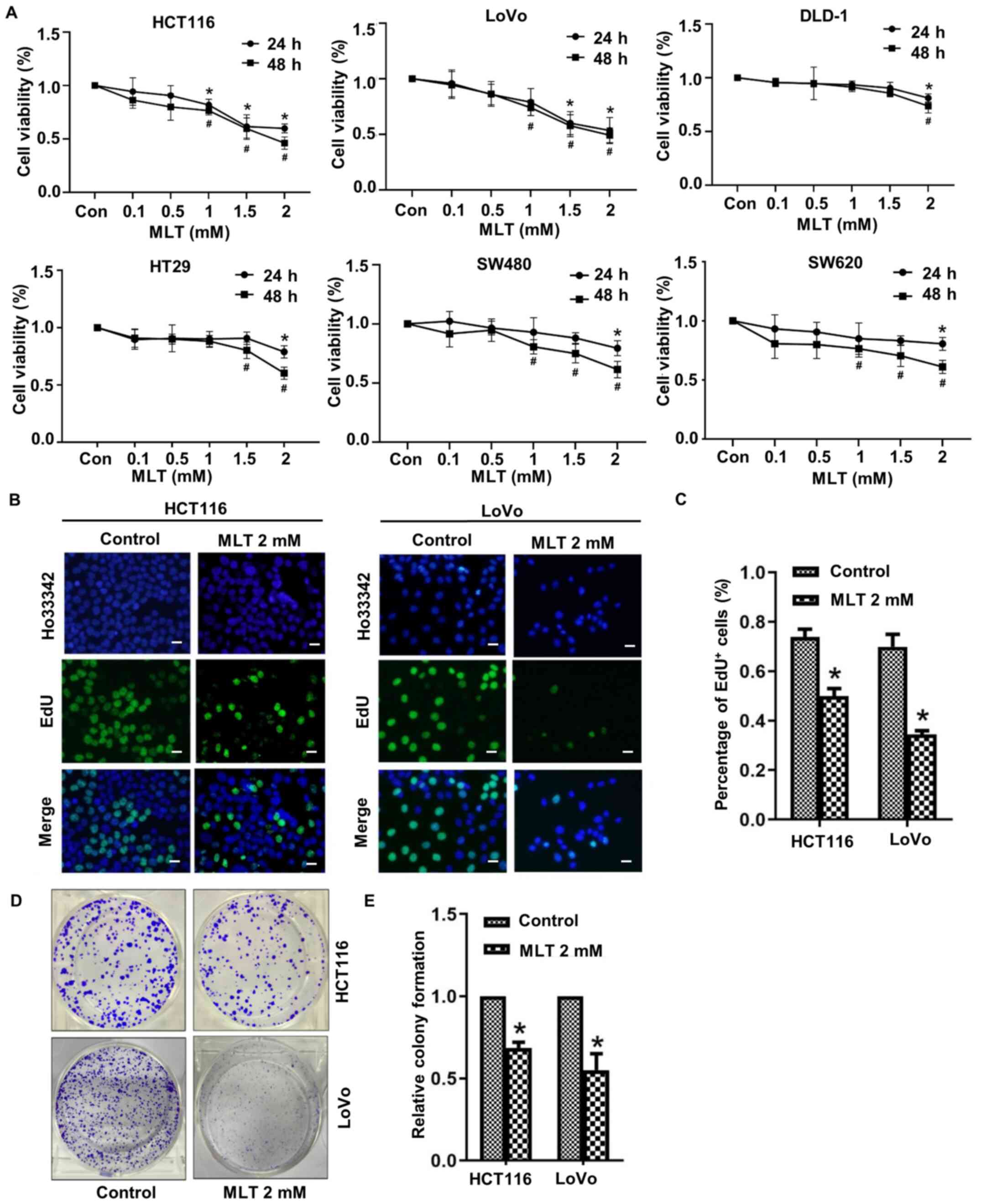 | Figure 1.Melatonin suppresses CRC cell
viability and proliferation. (A) HCT116, LoVo, SW480, SW620, HT-29
and DLD-1 CRC cells were treated with 0.1, 0.5, 1, 1.5 or 2 mM
melatonin for 24 or 48 h. Cell viability was assessed by performing
an MTT assay. *P<0.05 vs. 24 h control; #P<0.05
vs. 48 h control. Following treatment with melatonin for 48 h,
HCT116 and LoVo CRC cell proliferation was assessed by (B)
performing EdU staining (scale bar, 50 µm) and (C) quantified by
counting the number of EdU+ cells. Following treatment
with melatonin for 48 h, CRC cell proliferation was assessed by (D)
performing colony formation assays and (E) quantified. *P<0.05
vs. control. Data are presented as the mean ± standard deviation.
CRC, colorectal cancer; Con, control; MLT, melatonin. |
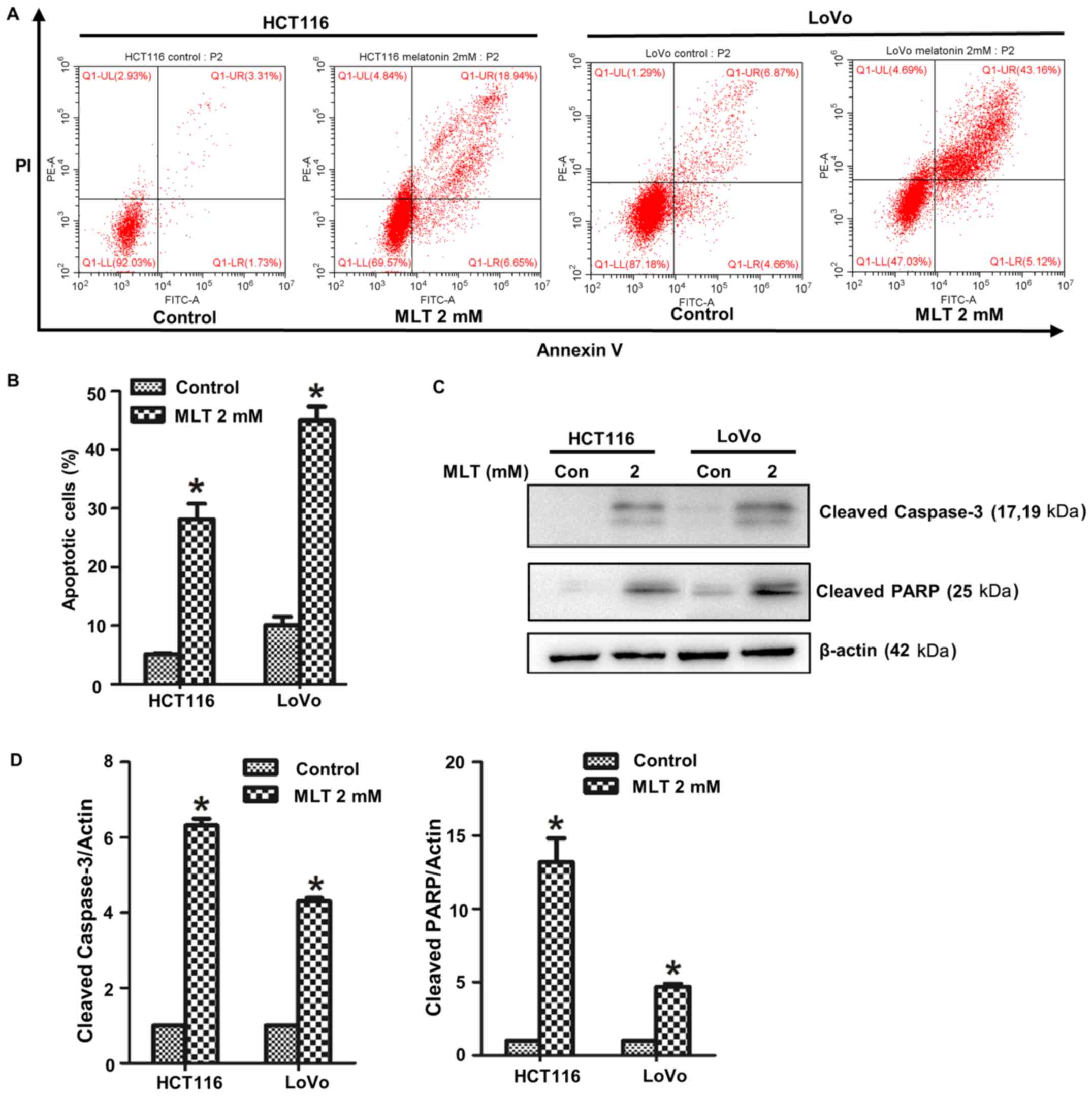
Melatonin induces CRC cell
apoptosis
To verify the effect of melatonin on HCT116 and LoVo
cell apoptosis, Annexin V/PI staining was conducted and the number
of apoptotic cells was determined via flow cytometry (Fig. 2A and B). Compared with the control
group, the number of apoptotic cells in the melatonin group was
significantly increased. The expression of apoptosis-related
proteins cleaved caspase-3 and cleaved PARP was determined via
western blotting (Fig. 2C and D).
The results indicated that the apoptotic rate of melatonin-treated
cells was significantly higher compared with the control group.
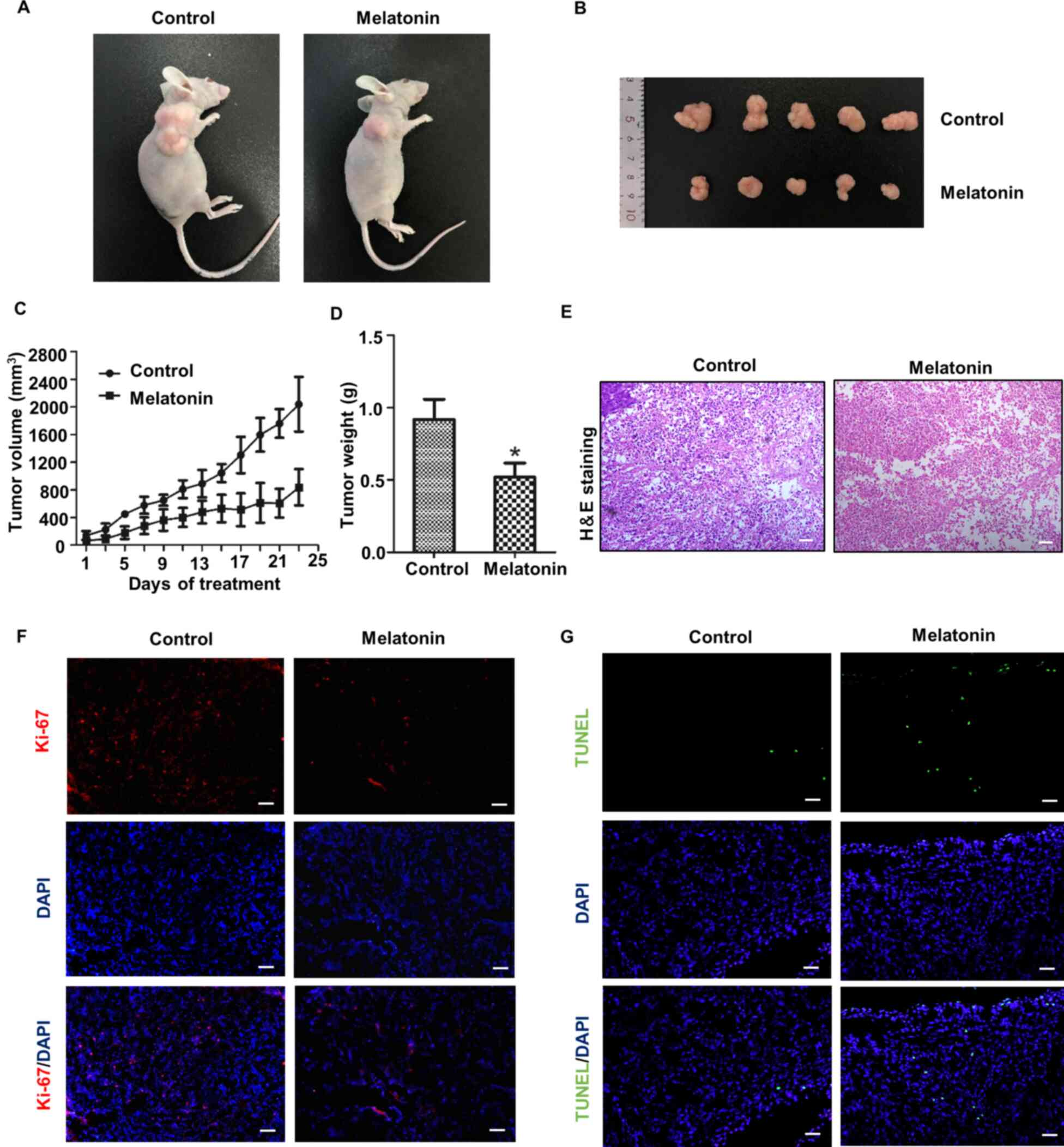
Melatonin inhibits tumor growth in
vivo
A nude mouse tumorigenesis model was established to
assess the inhibitory effect of melatonin on tumor growth in
vivo. Compared with the control group, the tumors in the
melatonin group were notably smaller (Fig. 3A). During the tumor-bearing period,
the tumor growth in the melatonin-treated group was clearly slower
compared with the control group, and the corresponding tumor volume
was smaller (Fig. 3B and C). The
tumor weight of the two groups was assessed at the end of the
experiments, and the results demonstrated that the average weight
of tumors in the melatonin group was significantly lower compared
with the control group (Fig. 3D).
Hematoxylin and eosin staining was performed and the results
indicated that the melatonin group displayed an increased number of
tissues with tumor cell necrosis compared with the control group
(Fig. 3E). Finally, tumor cell
proliferation was determined by performing Ki-67 staining. Tumor
cell proliferation was notably inhibited in the melatonin group
compared with the control group (Fig.
3F). TUNEL staining was also performed to assess tumor cell
apoptosis, which indicated that the melatonin group displayed
obviously increased levels of apoptosis compared with the control
group (Fig. 3G).
Melatonin affects CRC cell
proliferation and apoptosis by regulating the expression of the
miR-34a/449a cluster
Whether the aforementioned results were related to
miRNA expression was assessed. First, the expression of a series of
miRNAs (miR-125a, miR-365a, miR-34a, miR-449a, miR-184 and
miR-143a) in HCT116 cells treated with 2 mM melatonin for 48 h was
assessed via RT-qPCR. HCT116 cell viability and apoptosis were
significantly changed compared with other cell lines. All of the
aforementioned miRNAs have been reported to be associated with CRC
cell proliferation and apoptosis (36,38,56,57).
The results of the present study demonstrated that only miR-34a and
miR-449a expression levels were significantly altered by melatonin
treatment compared with the control group (Fig. 4A). Melatonin increased the
expression levels of miR-34a and miR-449a in a
concentration-dependent manner. miR-34a is a transcriptional target
of p53 (58). Therefore, the
expression of p53 in CRC cells treated with melatonin for 48 h was
assessed via western blotting. Compared with the control group,
melatonin markedly increased p53 expression levels in a
concentration-dependent manner (Fig.
S1). miR-34a and miR-449a belong to the same family, and work
together as a complex, termed the miR-34a/449a cluster (59). To further investigate whether the
increased expression of the miR-34a/449a cluster was related to
altered CRC cell proliferation and apoptosis, endogenous expression
of the miR-34a/449a cluster was downregulated by transfecting cells
with an miR-34a/449a cluster inhibitor. The transfection efficiency
of the miR-34a/449a cluster inhibitor is presented in Fig. 4B. The effect of the miR-34a/449a
cluster on CRC cell viability and proliferation was assessed by
performing CCK-8 and EdU staining assays, respectively. Compared
with the control group, CRC cell viability and proliferation were
significantly decreased in the melatonin group (Fig. 4C-E). However, in CRC cells
pretreated with inhibitor and then treated with melatonin, the
inhibitory effect of melatonin on CRC cell viability and
proliferation was significantly weakened to a certain degree.
Finally, the effect of the miR-34a/449a cluster on CRC cell
apoptosis was assessed via flow cytometry. Pretreatment with the
inhibitor significantly weakened the effect of melatonin on CRC
cell apoptosis (Fig. 4F and G). The
results suggested that melatonin inhibited viability and
proliferation, and promoted apoptosis in CRC cells by upregulating
the expression of the miR-34a/449a cluster.
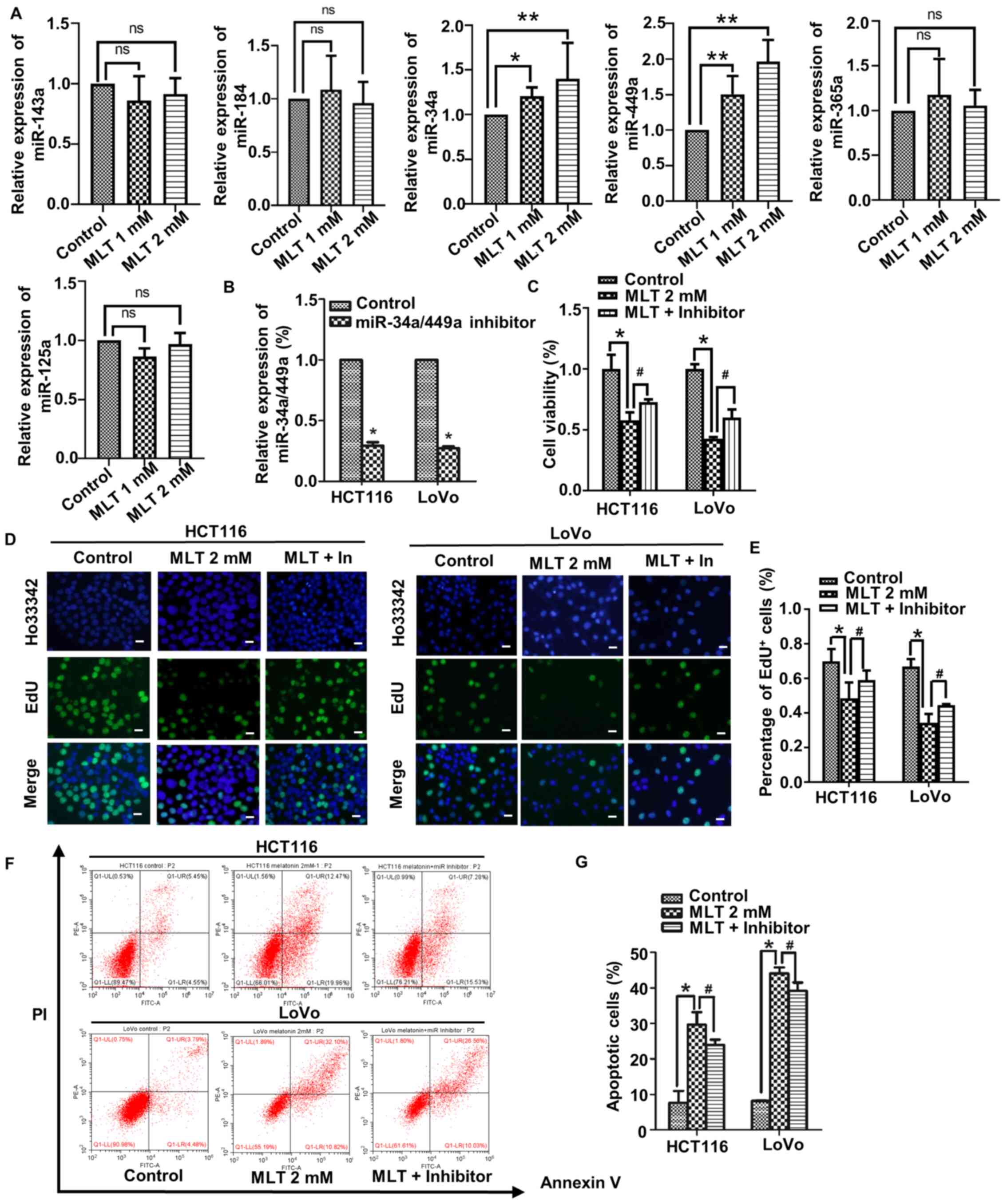 | Figure 4.Melatonin affects CRC cell
proliferation and apoptosis by regulating the expression of the
miR-34a/449a cluster. HCT116 cells were treated with 2 mM melatonin
for 48 h. (A) The expression levels of miR-125a, miR-365a, miR-34a,
miR-449a, miR-184 and miR-143a was determined via reverse
transcription-quantitative PCR. (B) The transfection efficiency of
miR-34a/449a inhibitor in HCT116 and LoVo cells. *P<0.05 vs.
control. (C) HCT116 and LoVo cell viability were detected using a
Cell Counting Kit-8 assay. CRC cell proliferation was (D)
determined by performing EdU staining (scale bar, 50 µm) and (E)
the number of EdU+ cells was quantified. CRC cell
apoptosis was (F) determined via flow cytometry and (G) quantified.
*P<0.05, **P<0.01 and #P<0.05 vs. MLT 2 mM.
Data are presented as the mean ± standard deviation. CRC,
colorectal cancer; miR, microRNA; ns, not significant; MLT,
melatonin. |
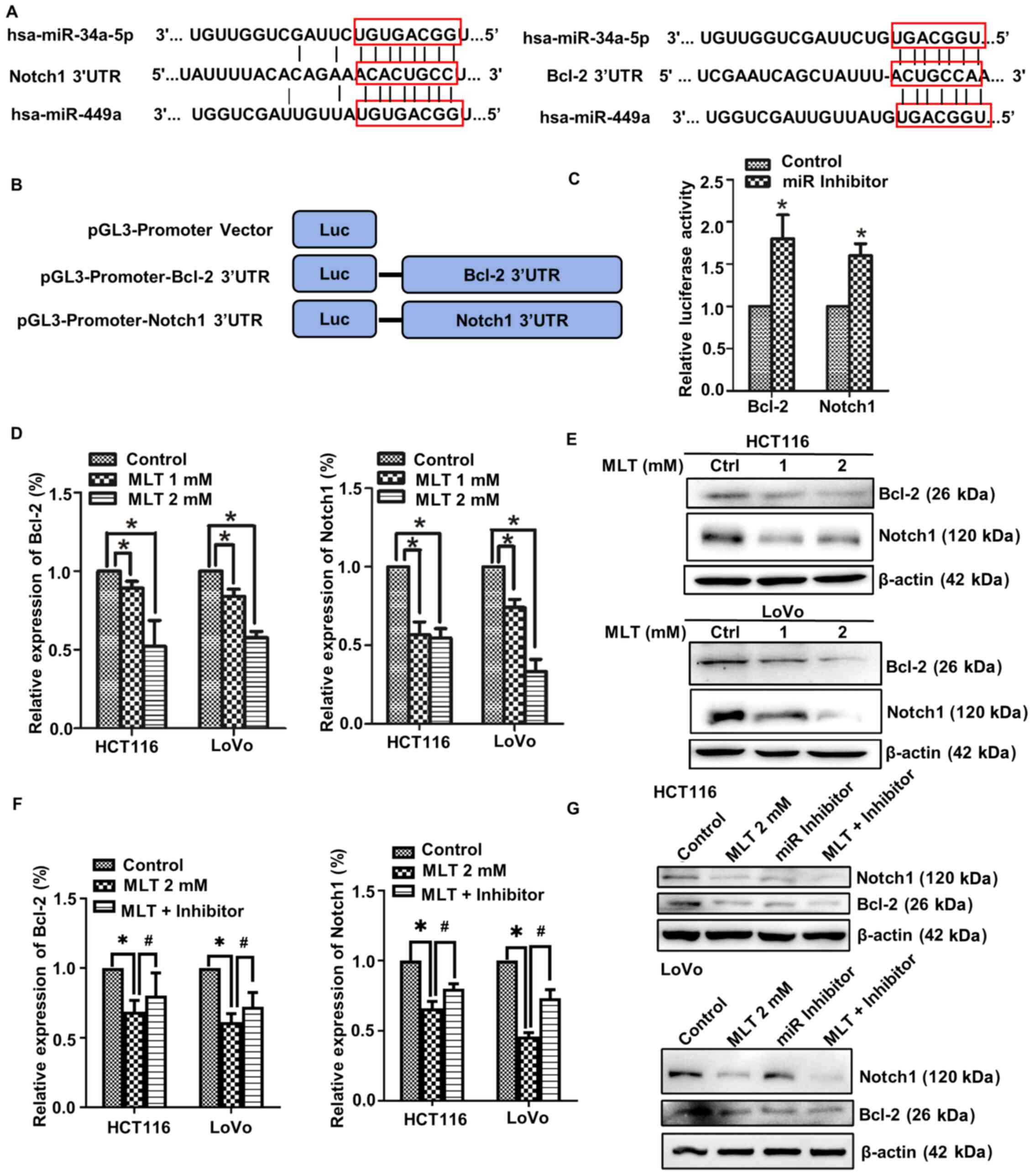
Bcl-2 and Notch1 are direct target
genes of the miR-34a/449a cluster
To further investigate how the miR-34a/449a cluster
exerted its regulatory function in melatonin-induced
antiproliferative and proapoptotic activities in CRC cells, the
downstream target genes of the miR-34a/449a cluster were determined
by performing bioinformatics analysis with TargetScan (http://www.targetscan.org/vert_71/). It was
predicted that the miR-34a/449a cluster bound to the 3′UTRs of
Bcl-2 and Notch1 (Fig. 5A).
pGL3-promoter-Bcl-2/Notch1 3′UTR vectors were constructed, and the
relative luciferase activity was assessed using a dual-luciferase
reporter assay. miR-34a/449a cluster inhibition significantly
increased the relative luciferase activity of pGL3-promoter-BCL2
3′UTR and pGL3-promoter-NOTCH1 3′UTR compared with the control
(Fig. 5B and C), suggesting that
Bcl-2 and Notch1 were direct target genes of the miR-34a/449a
cluster. RT-qPCR and western blotting were performed to measure the
mRNA and protein expression levels of Bcl-2 and Notch1 in
melatonin-treated CRC cells, respectively. Compared with the
control group, the mRNA and protein expression levels of Bcl-2 and
Notch1 were notably downregulated by melatonin treatment (Fig. 5D and E). The RT-qPCR results
demonstrated that miR-34a/449a cluster inhibition increased the
mRNA expression levels of Bcl-2 and Notch1 in melatonin-treated CRC
cells (Fig. 5F). The western
blotting results were similar compared with melatonin group, the
protein expression levels of Bcl-2 and Notch1 of miR inhibitor
group were markedly upregulated following transfection with the
miR-34a/449a cluster inhibitor (Fig.
5G). Collectively, the results indicated that the miR-34a/449a
cluster bound to the 3′UTRs of Bcl-2 and Notch1 to negatively
regulate their expression.
Discussion
CRC is one of the most common types of malignant
cancer of the digestive system worldwide and is associated with
high rates of morbidity and mortality (1,2). In
2020, the estimated new cases of colon cancer in the United States
are 78,300 in men, and 69,650 in women, accounting for 8–9% of all
cancer cases (2). In 50% of
patients with CRC, tumor recurrence is observed, even if the tumor
tissue is excised prior to tumor cell metastasis (60). In addition, the effect of
chemotherapeutic drugs on malignant colorectal tissue is reduced in
advanced phases (61). A
combinations of different agents may be used to overcome the
limitations of these therapeutics; however, this can also result in
increased side effects, off-target cytotoxicity and drug toxicity
(62). Therefore, improved
chemotherapeutic agents that display little to no unfavorable
off-target effects need to be identified. Melatonin is a natural
hormone that does not display toxicity and regulates numerous
physiological processes, including the circadian rhythm and
antioxidative processes (63,64).
An increasing number of studies have demonstrated that melatonin
also possesses anticancer properties in several types of cancer
(65,66), for example, melatonin inhibits
proliferation and invasion via repression of miRNA-155 in glioma
cells (45). Melatonin restrains
angiogenic factors in triple-negative breast cancer by targeting
miR-152-3p in in vivo and in vitro studies (55). Moreover, melatonin possesses
anticancer properties via various molecular mechanisms in CRC cells
(25,67–69),
for example, melatonin reduces endothelin-1 expression and
secretion in colon cancer cells through the inactivation of FoxO-1
and NF-κβ (26); Melatonin inhibits
colon cancer RKO cell migration by downregulating Rho-associated
protein kinase expression via the p38/MAPK signaling pathway
(69). To the best of our
knowledge, the present study demonstrated for the first time that
melatonin inhibited proliferation and viability, and induced
apoptosis in CRC cells in vitro and in vivo via
upregulating the expression of miR-34a/449a cluster, and
suppressing Bcl-2 and Notch1 expression.
In the present study, CRC cell (HCT116, LoVo, SW480,
SW620, HT-29 and DLD-1) viability following treatment with
melatonin (0.1–2 mM) for 24 and 48 h was assessed. Compared with
the control group, SW480 and SW620 cell viability was inhibited by
treatment with 1 mM melatonin for 48 h, whereas DLD-1 cell
viability was inhibited by treatment with 2 mM melatonin for 48 h.
The differences between the cell lines could be explained by
differences in melatonin concentrations, cell types and
experimental situations. Wei et al (70) reported that 2 mM melatonin
significantly suppressed LoVo cell proliferation following
treatment for 48 h via histone deacetylase 4 nuclear import
mediated by calcium/calmodulin dependent protein kinase IIα alpha
inactivation. León et al (26) reported that treatment of Caco-2 and
T84 CRC cells for 72 h with 1 mM melatonin induced cell cycle
arrest at the G2/M phase. Riabykh et al (22), Farriol et al (23) and Anisimov et al (24) reported that melatonin inhibited CRC
cell proliferation via decreasing DNA synthesis or promoting cell
differentiation. In the present study, compared with the control
group, HCT116 and LoVo cell viability was significantly inhibited
by 50% following treatment with 2 mM melatonin for 48 h via
upregulation of the expression of the miR-34a/449a cluster. The
aforementioned studies and the present study suggested that
melatonin displayed a direct antitumor effect on CRC cells.
Moreover, the results of the present study provided further
evidence for the anticancer effects of melatonin, indicating that
melatonin regulated the expression of miRNAs.
Although there are various mechanisms underlying
melatonin-mediated inhibition of cell proliferation and promotion
of cell apoptosis, the relationship between its effects is not
completely understood. Trivedi et al (71) investigated the relationship among
melatonin, colitis and malignant tumors using a colitis-associated
colon carcinogenesis mice model. Their study was aimed at
deciphering the effect of melatonin on autophagy and Nrf2 signaling
pathways in a mouse model of colitis-associated colon
carcinogenesis (CACC), and they focused on the relationship between
melatonin and inflammation. However, this mice model did not study
the relationship between melatonin and tumorigenesis directly. To
verify the antitumor effects of melatonin in vivo, a
xenograft mouse model was established and tumor cell proliferation
and apoptosis were assessed in the present study and the results
suggested that compared with the control group, melatonin inhibited
proliferation and induced apoptosis in CRC cells, suggesting that
melatonin may serve as a potential anti-CRC therapeutic.
miRNAs function as tumor oncogenes or suppressors,
and serve a key role in the development and progression of cancer
(38,72,73).
Melatonin results in upregulated miR-362-3p expression, and
suppressed TNF-α induced protein 8 and neuropilin 2expression
levels in breast cancer (74).
Melatonin inhibited glioma cell proliferation and invasion via
repression of miR-155 (45).
Melatonin also inhibited gastric cancer cell proliferation by
upregulating the expression of miR-16-5p (43). The aforementioned studies
highlighted the critical roles of miRNAs in melatonin-induced
attenuation of cancer growth. However, the association between
melatonin and miRNAs in CRC is not completely understood. It has
been reported that the members of miR-34 family are associated with
tumor metastasis and growth (75,76).
miR-34a, as a tumor suppressor, has been reported to serve a key
role in several types of solid tumors, including renal cell
carcinoma (77), glioblastoma
(78) and colon cancer (79). The expression of miR-34a was
downregulated by 36% in CRC tissues compared with healthy tissues
(80). In addition, Zhang et
al (79) and Wu et al
(57) suggested that miR-34a
suppressed CRC metastasis and invasion. Together, the
aforementioned studies suggest that miR-34a serves a key role in
CRC. However, whether melatonin exerts its antitumor effects by
regulating the expression of the miR-34a/449a cluster has not been
previously reported, therefore, whether this cluster was involved
in the therapeutic effects of melatonin was assessed in the present
study. Compared with the control group, melatonin significantly
increased the expression of the miR-34a/449a cluster. Furthermore,
melatonin inhibited proliferation and viability, and induced
apoptosis in CRC cells via upregulating the expression of the
miR-34a/449a cluster. To the best of our knowledge, the present
study was the first to suggest the important regulatory function of
miRNAs in melatonin-induced antiproliferative and proapoptotic
activities of CRC cells, providing further evidence to support the
critical role of miRNAs in melatonin-treated cancer.
In the present study, the bioinformatics based
target prediction suggested that Notch1 and Bcl-2 were direct
target genes of the miR-34a/449a cluster. Notch1 and Bcl-2 are
critical regulators of CRC cell proliferation, apoptosis and
development (81–83). Notch1 and Bcl-2 are regulated by
miRNAs in various types of cancer. In gastric cancer, miR-708
targeted Notch1 and subsequently suppressed cell proliferation
(84). In CRC cells, miR-34a
directly targeted Notch1, which was associated with tumor invasion
and metastasis (79). In renal cell
carcinoma, miR-34a inhibited cell proliferation by targeting Notch1
(77). Thus, whether
melatonin-mediated upregulation of the miR-34a/449a cluster,
resulting in downregulation of Notch1, ultimately inhibited CRC
cell proliferation was assessed. In human cervical cancer, Bcl-2 is
the target of miR-34a-5p and is associated with proliferation and
apoptosis (85). In prostate
carcinoma, miR-205 and miR-338-3p downregulate the expression of
the Bcl-2, which ultimately results in decreased apoptosis
(72). Numerous studies have
demonstrated that Bcl-2 is regulated by various miRNAs, which
subsequently affects cell apoptosis (37,38).
However, the relationship among melatonin, the miR-34a/449a cluster
and Bcl-2 in CRC is not completely understood. It was hypothesized
that melatonin upregulated the miR-34a/449a cluster by
downregulating Bcl-2 expression, which subsequently induced CRC
cell apoptosis. The results indicated that melatonin upregulated
the expression of the miR-34a/449a cluster, which resulted in
downregulated Notch1 and Bcl-2 expression levels compared with the
control group. The results indicated that the
miR-34a/449a/Notch1/Bcl-2 axis may serve as a critical mediator of
melatonin-treated CRC.
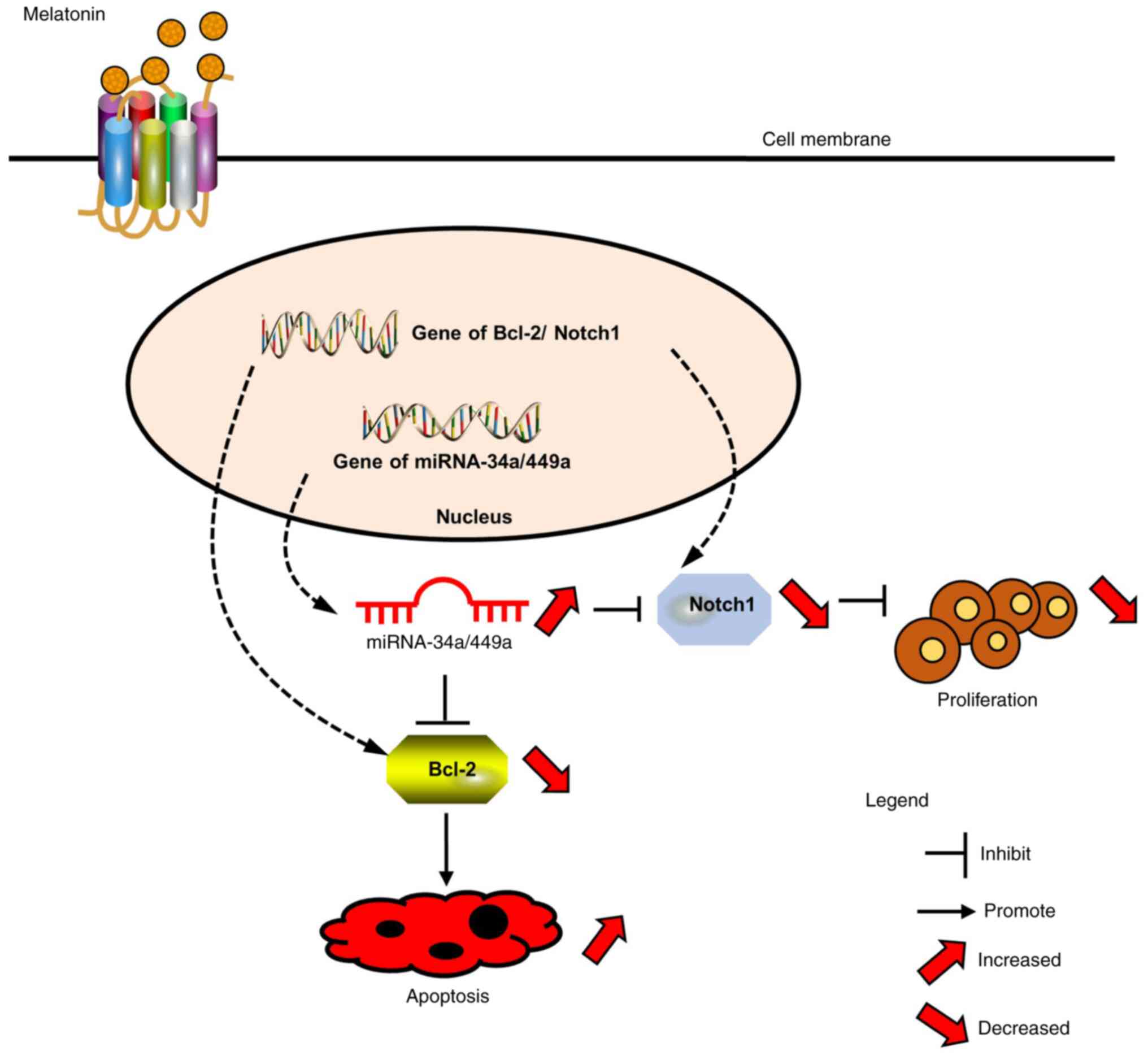
In summary, the present study demonstrated that
melatonin inhibited proliferation and viability, and induced
apoptosis of CRC cells via upregulating the expression of the
miR-34a/449a cluster (Fig. 6). To
the best of our knowledge, the present study was the first to
identify an association among melatonin, the miR-34a/449a cluster,
Notch1 and Bcl-2 in CRC. The results suggested that melatonin may
be useful as a single or adjuvant therapeutic for CRC.
Supplementary Material
Supporting Data
Acknowledgements
Not applicable.
Funding
The present study was supported by the Natural
Science Foundation of Shandong Province (grant no.
2018GSF118076).
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
GJ and HH designed the experiments. GJ, JD, XiaL, HH
and XinL acquired and analysed data for the work. GJ, WZ and XiaL
revised the manuscript for important intellectual content. WZ was
involved in acquiring and analyzing data for the work, and
designing the in vivo experiments. HH gave final approval of
the version to be published. GJ and HH agreed to be accountable for
the work in ensuring that questions related to the integrity of any
part of the work are appropriately investigated and resolved. All
authors read and approved the final manuscript.
Ethics approval and consent to
participate
The present study followed the Guide for the Care
and Use of Laboratory Animals (Eighth edition; 2011) in Animal Care
and Treatment. The present study was approved by the Ethics
Committee on Animal Experiments of the Medical School of Shandong
University (approval no. KYLL-2017(KS)-357).
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Chen W, Zheng R, Baade PD, Zhang S, Zeng
H, Bray F, Jemal A, Yu XQ and He J: Cancer statistics in China,
2015. CA Cancer J Clin. 66:115–132. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Siegel RLM, Miller KD and Jemal A: Cancer
statistics, 2020. CA Cancer J Clin. 70:7–30. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Zhang Y, Chen Z and Li J: The current
status of treatment for colorectal cancer in China: A systematic
review. Medicine (Baltimore). 96:e82422017. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Scholefield JH and Steele RJ; British
Society For Gastroenterology; Association of Coloproctology for
Great Britain and Ireland, : Guidelines for follow up after
resection of colorectal cancer. Gut. 51 (Suppl 5):V3–V5. 2002.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Cunningham D, Humblet Y, Siena S, Khayat
D, Bleiberg H, Santoro A, Bets D, Mueser M, Harstrick A, Verslype
C, et al: Cetuximab monotherapy and cetuximab plus irinotecan in
irinotecan-refractory metastatic colorectal cancer. N Engl J Med.
351:337–345. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Van Cutsem E, Peeters M, Siena S, Humblet
Y, Hendlisz A, Neyns B, Canon JL, Van Laethem JL, Maurel J,
Richardson G, et al: Open-label phase III trial of panitumumab plus
best supportive care compared with best supportive care alone in
patients with chemotherapy-refractory metastatic colorectal cancer.
J Clin Oncol. 25:1658–1664. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Wu H, Liu J, Yin Y, Zhang D, Xia P and Zhu
G: Therapeutic opportunities in colorectal cancer: Focus on
melatonin antioncogenic action. BioMed Res Int. 2019:97405682019.
View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Claustrat B, Brun J and Chazot G: The
basic physiology and pathophysiology of melatonin. Sleep Med Rev.
9:11–24. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Acuña-Castroviejo D, Escames G, Venegas C,
Díaz-Casado ME, Lima-Cabello E, López LC, Rosales-Corral S, Tan DX
and Reiter RJ: Extrapineal melatonin: Sources, regulation, and
potential functions. Cell Mol Life Sci. 71:2997–3025. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Bubenik GA: Gastrointestinal melatonin:
Localization, function, and clinical relevance. Dig Dis Sci.
47:2336–2348. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Reiter RJ; RJ R, : Melatonin: The chemical
expression of darkness. Mol Cell Endocrinol. 79:C153–C158. 1991.
View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Reiter RJ, Tan DX, Mayo JC, Sainz RM, Leon
J and Czarnocki Z: Melatonin as an antioxidant: biochemical
mechanisms and pathophysiological implications in humans. Acta
Biochim Pol. 50:1129–1146. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Min KJ, Jang JH and Kwon TK: Inhibitory
effects of melatonin on the lipopolysaccharide-induced CC chemokine
expression in BV2 murine microglial cells are mediated by
suppression of Akt-induced NF-κB and STAT/GAS activity. J Pineal
Res. 52:296–304. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Joo SS and Yoo YM: Melatonin induces
apoptotic death in LNCaP cells via p38 and JNK pathways:
Therapeutic implications for prostate cancer. J Pineal Res.
47:8–14. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Zha L, Fan L, Sun G, Wang H, Ma T, Zhong F
and Wei W: Melatonin sensitizes human hepatoma cells to endoplasmic
reticulum stress-induced apoptosis. J Pineal Res. 52:322–331. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Ren W, Wang P, Yan J, Liu G, Zeng B,
Hussain T, Peng C, Yin J, Li T, Wei H, et al: Melatonin alleviates
weanling stress in mice: Involvement of intestinal microbiota. J
Pineal Res. Dec 20–2017.(Epub ahead of print). doi:
10.1111/jpi.12448.
|
|
17
|
Yin J, Li Y, Han H, Chen S, Gao J, Liu G,
Wu X, Deng J, Yu Q, Huang X, et al: Melatonin reprogramming of gut
microbiota improves lipid dysmetabolism in high-fat diet-fed mice.
J Pineal Res. 65:e125242018. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Kannen V, Marini T, Zanette DL, Frajacomo
FT, Silva GE Jr, Silva WA Jr and Garcia SB: The melatonin action on
stromal stem cells within pericryptal area in colon cancer model
under constant light. Biochem Biophys Res Commun. 405:593–598.
2011. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Wang J, Guo W, Chen W, Yu W, Tian Y, Fu L,
Shi D, Tong B, Xiao X, Huang W, et al: Melatonin potentiates the
antiproliferative and pro-apoptotic effects of ursolic acid in
colon cancer cells by modulating multiple signaling pathways. J
Pineal Res. 54:406–416. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Anisimov VN: Light pollution, reproductive
function and cancer risk. Neuro Endocrinol Lett. 27:35–52.
2006.PubMed/NCBI
|
|
21
|
Anisimov VN, Vinogradova IA, Panchenko AV,
Popovich IG and Zabezhinski MA: Light-at-night-induced circadian
disruption, cancer and aging. Curr Aging Sci. 5:170–177. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Riabykh TP, Nikolaeva TG and Bodrova NB:
Effects of biorhythm regulator melatonin on DNA synthesis in
short-term cultures of human malignant tumors. Vestn Ross Akad Med
Nauk. 8:30–33. 2000.(In Russian).
|
|
23
|
Farriol M, Venereo Y, Orta X, Castellanos
JM and Segovia-Silvestre T: In vitro effects of melatonin on cell
proliferation in a colon adenocarcinoma line. J Appl Toxicol.
20:21–24. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Anisimov VN, Popovich IG and Zabezhinski
MA: Melatonin and colon carcinogenesis: I. Inhibitory effect of
melatonin on development of intestinal tumors induced by
1,2-dimethylhydrazine in rats. Carcinogenesis. 18:1549–1553. 1997.
View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Hong Y, Won J, Lee Y, Lee S, Park K, Chang
KT and Hong Y: Melatonin treatment induces interplay of apoptosis,
autophagy, and senescence in human colorectal cancer cells. J
Pineal Res. 56:264–274. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
León J, Casado J, Sergio M, Ruiz J, Zurita
MS, González-Puga C, Rejón JD, Gila A, Muñoz de Rueda P, Pavón EJ,
et al: Melatonin reduces endothelin-1 expression and secretion in
colon cancer cells through the inactivation of FoxO-1 and
NF-kappaβ. J Pineal Res. 56:415–426. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Yu X, Zheng H, Chan MT and Wu WK:
Modulation of chemoresponsiveness to platinum-based agents by
microRNAs in cancer. Am J Cancer Res. 7:1769–1778. 2017.PubMed/NCBI
|
|
28
|
Dacosta C and Bao Y: The role of MicroRNAs
in the chemopreventive activity of sulforaphane from cruciferous
vegetables. Nutrients. 9:9022017. View Article : Google Scholar
|
|
29
|
Cochetti G, Rossi de Vermandois JA, Maulà
V, Giulietti M, Cecati M, Del Zingaro M, Cagnani R, Suvieri C,
Paladini A and Mearini E: Role of miRNAs in prostate cancer: Do we
really know everything? Urol Oncol. 38:623–635. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Shibuya H, Iinuma H, Shimada R, Horiuchi A
and Watanabe T: Clinicopathological and prognostic value of
microRNA-21 and microRNA-155 in colorectal cancer. Oncology.
79:313–320. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Liu L, Chen L, Xu Y, Li R and Du X:
microRNA-195 promotes apoptosis and suppresses tumorigenicity of
human colorectal cancer cells. Biochem Biophys Res Commun.
400:236–240. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Lu TX and Rothenberg ME: MicroRNA. J
Allergy Clin Immunol. 141:1202–1207. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Wong TS, Man OY, Tsang CM, Tsao SW, Tsang
RK, Chan JY, Ho WK, Wei WI and To VS: MicroRNA let-7 suppresses
nasopharyngeal carcinoma cells proliferation through downregulating
c-Myc expression. J Cancer Res Clin Oncol. 137:415–422. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Kang L, Mao J, Tao Y, Song B, Ma W, Lu Y,
Zhao L, Li J, Yang B and Li L: MicroRNA-34a suppresses the breast
cancer stem cell-like characteristics by downregulating Notch1
pathway. Cancer Sci. 106:700–708. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Godlewski J, Nowicki MO, Bronisz A,
Williams S, Otsuki A, Nuovo G, Raychaudhury A, Newton HB, Chiocca
EA and Lawler S: Targeting of the Bmi-1 oncogene/stem cell renewal
factor by microRNA-128 inhibits glioma proliferation and
self-renewal. Cancer Res. 68:9125–9130. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Wang YB, Zhao XH, Li G, Zheng JH and Qiu
W: MicroRNA-184 inhibits proliferation and promotes apoptosis of
human colon cancer SW480 and HCT116 cells by downregulating C-MYC
and BCL-2. J Cell Biochem. 119:1702–1715. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Li M, Yang Y, Kuang Y, Gan X, Zeng W, Liu
Y and Guan H: miR-365 induces hepatocellular carcinoma cell
apoptosis through targeting Bcl-2. Exp Ther Med. 13:2279–2285.
2017. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Tong Z, Liu N, Lin L, Guo X, Yang D and
Zhang Q: miR-125a-5p inhibits cell proliferation and induces
apoptosis in colon cancer via targeting BCL2, BCL2L12 and MCL1.
Biomed Pharmacother. 75:129–136. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Rodriguez A, Griffiths-Jones S, Ashurst JL
and Bradley A: Identification of mammalian microRNA host genes and
transcription units. Genome Res. 14:1902–1910. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Tang H, Bian Y, Tu C, Wang Z, Yu Z, Liu Q,
Xu G, Wu M and Li G: The miR-183/96/182 cluster regulates oxidative
apoptosis and sensitizes cells to chemotherapy in gliomas. Curr
Cancer Drug Targets. 13:221–231. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Altuvia Y, Landgraf P, Lithwick G, Elefant
N, Pfeffer S, Aravin A, Brownstein MJ, Tuschl T and Margalit H:
Clustering and conservation patterns of human microRNAs. Nucleic
Acids Res. 33:2697–2706. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Weber MJ: New human and mouse microRNA
genes found by homology search. FEBS J. 272:59–73. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Zhu C, Huang Q and Zhu H: Melatonin
inhibits the proliferation of gastric cancer cells through
Regulating the miR-16-5p-Smad3 pathway. DNA Cell Biol. 37:244–252.
2018. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Sohn EJ, Won G, Lee J, Lee S and Kim SH:
Upregulation of miRNA3195 and miRNA374b mediates the
anti-angiogenic properties of melatonin in hypoxic PC-3 prostate
cancer cells. J Cancer. 6:19–28. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Gu J, Lu Z, Ji C, Chen Y, Liu Y, Lei Z,
Wang L, Zhang HT and Li X: Melatonin inhibits proliferation and
invasion via repression of miRNA-155 in glioma cells. Biomed
Pharmacother. 93:969–975. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Messner M, Huether G, Lorf T, Ramadori G
and Schwörer H: Presence of melatonin in the human
hepatobiliary-gastrointestinal tract. Life Sci. 69:543–551. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Claustrat B and Leston J: Melatonin:
Physiological effects in humans. Neurochirurgie. 61:77–84. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Hong Y, Won J, Lee Y, Lee S, Park K, Chang
KT and Hong Y: Melatonin treatment induces interplay of apoptosis,
autophagy, and senescence in human colorectal cancer cells. J
Pineal Res. 56:264–274. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Park SY, Jang WJ, Yi EY, Jang JY, Jung Y,
Jeong JW and Kim YJ: Melatonin suppresses tumor angiogenesis by
inhibiting HIF-1a stabilization under hypoxia. J Pineal Res.
48:178–184. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Xu M, Chen X, Lin K, Zeng K, Liu X, Pan B,
Xu X, Xu T, Hu X, Sun L, et al: The long noncoding RNA SNHG1
regulates colorectal cancer cell growth through interactions with
EZH2 and miR-154-5p. Mol Cancer. 17:1412018. View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Zhao SJ, Shen YF, Li Q, He YJ, Zhang YK,
Hu LP, Jiang YQ, Xu NW, Wang YJ, Li J, et al: SLIT2/ROBO1 axis
contributes to the Warburg effect in osteosarcoma through
activation of SRC/ERK/c-MYC/PFKFB2 pathway. Cell Death Dis.
9:3902018. View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Livak KJS and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
53
|
Yang CY, Lin CK, Tsao CH, Hsieh CC, Lin
GJ, Ma KH, Shieh YS, Sytwu HK and Chen YW: Melatonin exerts
anti-oral cancer effect via suppressing LSD1 in patient-derived
tumor xenograft models. Oncotarget. 8:33756–33769. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
54
|
Wang Q, Sun Z, Du L, Xu C, Wang Y, Yang B,
He N, Wang J, Ji K, Liu Y, et al: Melatonin sensitizes human
colorectal cancer cells to γ-ray ionizing radiation in vitro and in
vivo. Int J Mol Sci. 19:39742018. View Article : Google Scholar
|
|
55
|
Marques JH, Mota AL, Oliveira JG, Lacerda
JZ, Stefani JP, Ferreira LC, Castro TB, Aristizábal-Pachón AF and
Zuccari DA: Melatonin restrains angiogenic factors in
triple-negative breast cancer by targeting miR-152-3p: In vivo and
in vitro studies. Life Sci. 208:131–138. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
56
|
Hong YG, Xin C, Zheng H, Huang ZP, Yang Y,
Zhou JD, Gao XH, Hao L, Liu QZ, Zhang W, et al: miR-365a-3p
regulates ADAM10-JAK-STAT signaling to suppress the growth and
metastasis of colorectal cancer cells. J Cancer. 11:3634–3644.
2020. View Article : Google Scholar : PubMed/NCBI
|
|
57
|
Wu J, Wu G, Lv L, Ren YF, Zhang XJ, Xue
YF, Li G, Lu X, Sun Z and Tang KF: MicroRNA-34a inhibits migration
and invasion of colon cancer cells via targeting to Fra-1.
Carcinogenesis. 33:519–528. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
58
|
Cekaite P, Eide PW, Lind GE, Skotheim RI
and Lothe RA: MicroRNAs as growth regulators, their function and
biomarker status in colorectal cancer. Oncotarget. 7:6476–6505.
2015. View Article : Google Scholar
|
|
59
|
Lv J, Zhang Z, Pan L and Zhang Y:
MicroRNA-34/449 family and viral infections. Virus Res. 260:1–6.
2019. View Article : Google Scholar : PubMed/NCBI
|
|
60
|
Sørensen NM, Schrohl AS, Jensen V,
Christensen IJ, Nielsen HJ and Brünner N: Comparative studies of
tissue inhibitor of metalloproteinases-1 in plasma, serum and
tumour tissue extracts from patients with primary colorectal
cancer. Scand J Gastroenterol. 43:186–191. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
61
|
Schwartz RN: Management of early and
advanced colorectal cancer: Therapeutic issues. Am J Health Syst
Pharm. 65 (Suppl 4):S8–S14; quiz S22-S24. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
62
|
Half E and Arber N: Colon cancer:
Preventive agents and the present status of chemoprevention. Expert
Opin Pharmacother. 10:211–219. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
63
|
Cipolla-Neto J and Amaral FGD: Melatonin
as a hormone: New physiological and clinical insights. Endocr Rev.
39:990–1028. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
64
|
Chitimus DM, Popescu MR, Voiculescu SE,
Panaitescu AM, Pavel B, Zagrean L and Zagrean AM: Melatonin's
impact on antioxidative and anti-inflammatory reprogramming in
homeostasis and disease. Biomolecules. 10:12112020. View Article : Google Scholar
|
|
65
|
Zhang S, Qi Y, Zhang H, He W, Zhou Q, Gui
S and Wang Y: Melatonin inhibits cell growth and migration, but
promotes apoptosis in gastric cancer cell line, SGC7901. Biotech
Histochem. 88:281–289. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
66
|
Li Y, Li S, Zhou Y, Meng X, Zhang JJ, Xu
DP and Li HB: Melatonin for the prevention and treatment of cancer.
Oncotarget. 8:39896–39921. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
67
|
Batista AP, da Silva TG, Teixeira AA, de
Medeiros PL, Teixeira VW, Alves LC, Dos Santos FA and Silva EC:
Ultrastructural aspects of melatonin cytotoxicity on Caco-2 cells
in vitro. Micron. 59:17–23. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
68
|
León J, Casado J, Jiménez Ruiz SM, Zurita
MS, González-Puga C, Rejón JD, Gila A, Muñoz de Rueda P, Pavón EJ,
Reiter RJ, et al: Melatonin reduces endothelin-1 expression and
secretion in colon cancer cells through the inactivation of FoxO-1
and NF-κβ. J Pineal Res. 56:415–426. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
69
|
Liu Z, Zou D, Yang X, Xue X, Zuo L, Zhou
Q, Hu R and Wang Y: Melatonin inhibits colon cancer RKO cell
migration by downregulating Rho associated protein kinase
expression via the p38/MAPK signaling pathway. Mol Med Rep.
16:9383–9392. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
70
|
Wei JY, Li WM, Zhou LL, Lu QN and He W:
Melatonin induces apoptosis of colorectal cancer cells through
HDAC4 nuclear import mediated by CaMKII inactivation. J Pineal Res.
58:429–438. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
71
|
Trivedi PP, Jena GB, Tikoo KB and Kumar V:
Melatonin modulated autophagy and Nrf2 signaling pathways in mice
with colitis-associated colon carcinogenesis. Mol Carcinog.
55:255–267. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
72
|
Zhang X, Pan Y, Fu H and Zhang J:
microRNA-205 and microRNA-338-3p reduces cell apoptosis in prostate
carcinoma tissue and LNCaP prostate carcinoma cells by directly
targeting the B-cell lymphoma-2 (Bcl-2) gene. Med Sci Monit.
25:1122–1132. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
73
|
Hongdan L and Feng L: miR-3120-5p promotes
colon cancer stem cell stemness and invasiveness through targeting
Axin2. Biochem Biophys Res Commun. 496:302–308. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
74
|
Kim TH and Cho SG: Melatonin-induced KiSS1
expression inhibits triple-negative breast cancer cell
invasiveness. Oncol Lett. 14:2511–2516. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
75
|
Farooqi AA, Tabassum S and Ahmad A:
MicroRNA-34a: A versatile regulator of myriads of targets in
different cancers. Int J Mol Sci. 18:2082017. View Article : Google Scholar
|
|
76
|
Li XJ, Ren ZJ and Tang JH: MicroRNA-34a: A
potential therapeutic target in human cancer. Cell Death Dis.
5:e13272014. View Article : Google Scholar : PubMed/NCBI
|
|
77
|
Zhang C, Mo R, Yin B, Zhou L, Liu Y and
Fan J: Tumor suppressor microRNA-34a inhibits cell proliferation by
targeting Notch1 in renal cell carcinoma. Oncol Lett. 7:1689–1694.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
78
|
Yu X, Zhang W, Ning Q and Luo X:
MicroRNA-34a inhibits human brain glioma cell growth by
down-regulation of Notch1. J Huazhong Univ Sci Technolog Med Sci.
32:370–374. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
79
|
Zhang X, Ai F, Li X, Tian L, Wang X, Shen
S and Liu F: MicroRNA-34a suppresses colorectal cancer metastasis
by regulating Notch signaling. Oncol Lett. 14:2325–2333. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
80
|
Tazawa H, Tsuchiya N, Izumiya M and
Nakagama H: Tumor-suppressive miR-34a induces senescence-like
growth arrest through modulation of the E2F pathway in human colon
cancer cells. Proc Natl Acad Sci USA. 104:15472–15477. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
81
|
Lindner AU, Salvucci M, Morgan C, Monsefi
N, Resler AJ, Cremona M, Curry S, Toomey S, O'Byrne R, Bacon O, et
al: BCL-2 system analysis identifies high-risk colorectal cancer
patients. Gut. 66:2141–2148. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
82
|
Ishiguro H, Okubo T, Kuwabara Y, Kimura M,
Mitsui A, Sugito N, Ogawa R, Katada T, Tanaka T, Shiozaki M, et al:
NOTCH1 activates the Wnt/β-catenin signaling pathway in colon
cancer. Oncotarget. 8:60378–60389. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
83
|
Ramesh P and Medema JP: BCL-2 family
deregulation in colorectal cancer: Potential for BH3 mimetics in
therapy. Apoptosis. 25:305–320. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
84
|
Li X, Zhong X, Pan X and Ji Y: Tumor
suppressive microRNA-708 targets Notch1 to suppress cell
proliferation and invasion in gastric cancer. Oncol Res.
26:1317–1326. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
85
|
Wang X, Xie Y and Wang J: [ARTICLE
WITHDRAWN] Overexpression of microRNA-34a-5p inhibits proliferation
and promotes apoptosis of human cervical cancer cells by
downregulation of Bcl-2. Oncol Res. 26:977–985. 2018. View Article : Google Scholar : PubMed/NCBI
|