Introduction
Diabetes mellitus (DM) is one of the most important
global health threats (1).
Accelerated and aggressive atherosclerosis leading to severe
cardiovascular events, such as ischemic heart disease and stroke,
accounts for an important cause of morbidity and mortality due to
cardiovascular disease in patients with DM (2). Notably, metabolic disease is
characterized by hyperglycemia, and lowering blood glucose levels
by diet, exercise and medication is an important foundation of DM
management (3). However, tight
glucose control has been reported to limit early microvascular
diseases, such as retinopathy and nephropathy, but not
macrovascular atherosclerosis, according to the Diabetes Control
and Complication Trial study (4–6). In
addition, more than half the increased risk of cardiovascular
disease in DM cannot be fully explained by well-known risk factors
(7), such as hypertension and
abnormal cholesterol, thus indicating that other elements
underlying DM may contribute to macrovascular atherosclerosis
(7,8).
Methylglyoxal (MG) is a highly reactive dicarbonyl
compound, which is a key contributor to atherosclerosis. MG is
derived from the glucose metabolic process, and is a precursor of
most of the functionally important spontaneous protein and DNA
modifications (9,10). Circulating and tissue MG are
3–5-fold higher in diabetic models (11). In patients with DM and vascular
complications, plasma MG levels are further elevated (12,13).
While limited data are available on the alteration of aortic MG
levels during DM, abundant literature supports the concept that MG
is important in the pathogenesis of atherosclerosis (14,15). A
previous report demonstrated that exposure to exogenous MG or
inhibition of glyoxalase-1 to increase endogenous MG levels was
able to increase vascular endothelial adhesion and augment
atherogenesis in apolipoprotein E (ApoE) knockout mice
(16). Furthermore, Goto-Kakizaki
rats (a type 2 DM model) treated with MG in drinking water
exhibited aggravated endothelial dysfunction and oxidative stress
(17,18). Overall, these studies suggested a
potential benefit for targeting MG in diabetic individuals.
Glutathione (GSH) is a rate-limiting co-factor in
the glyoxalase elimination pathway for MG. High glucose levels have
been reported to decrease the intracellular levels and rate of
uptake of cysteine, which is a rate-limiting factor that maintains
GSH levels in human aortic vascular smooth muscle cells (19). Reduced GSH content was further
detected in the aortic tissue of diabetic rats with hyperglycemia
in vivo (20).
N-acetylcysteine (NAC) is an effective agent to replenish GSH in
deficient cells. Previously, it was demonstrated that DM imbalanced
the MG/GSH ratio in brains of a stroke model, whereas NAC corrected
this ratio, and decreased dicarbonyl levels and oxidative stress to
reduce cerebral injury in a diabetic ischemia/reperfusion model
(21,22). These findings suggested a preventive
role of NAC in endothelial dysfunction, which may be associated
with the correction of MG/GSH levels and oxidative stress (21,22).
Since oxidative stress and endothelial dysfunction have been
identified as common upstream events that mediate the atherogenic
effects of hyperglycemia (23,24),
the present study aimed to investigate the anti-atherosclerotic
role of NAC in DM and whether the mechanism involved decreased
oxidative stress and endothelial damage via GSH-dependent MG
elimination in the aorta.
Materials and methods
Materials
MG (cat. no. M0252), GSH (cat. no. G4251), NAC (cat.
no. A7250), streptozotocin (STZ; cat. no. S0130), the GSH synthesis
inhibitor buthionine sulfoximine (BSO; cat. no. B2640) and
guanidine hydrochloride (cat. no. G7294) were purchased from
Sigma-Aldrich (Merck KGaA). Rabbit anti-glutathione peroxidase-1
(GPX-1; cat. no. ab22604), rabbit anti-superoxide dismutase-1
(SOD-1; cat. no. ab51254), rabbit anti-endothelial nitric oxide
synthase (eNOS; cat. no. ab5589), anti-phosphorylated (p)-eNOS
(cat. no. ab215717) and HRP-conjugated goat-anti-rabbit IgG (cat.
no. ab150077) were purchased from Abcam. Rabbit anti-Akt (cat. no.
sc-8312) and anti-p-Akt (cat. no. sc-7985 R) were obtained from
Santa Cruz Biotechnology, Inc. Mouse anti-actin antibody (cat. no.
612656) was obtained from BD Biosciences. HRP-conjugated
sheep-anti-mouse IgG (cat. no. NA931) was purchased from Amersham
(Cytiva). Enhanced chemiluminescence (ECL) reagent (cat. no.
1705060) was acquired from Bio-Rad Laboratories, Inc. Nitric oxide
(NO; cat. no. A012), which was used for the nitrate reductase
method according to the instructions of the NO assay kit, and the
malondialdehyde (MDA) assay kit (cat. no. A003) were obtained from
Nanjing Jiancheng Bioengineering Institute.
Animal preparation
Male ApoE−/− mice (age, 5 weeks;
weight, 20–25 g; Jackson Laboratory, n=40) were fed at the Animal
Center of Huazhong University of Science and Technology (Wuhan,
China), following the Guide for the Care and Use of Laboratory
Animals published by the National Institutes of Health (25). Rearing temperature was between
22–25°C, relative humidity was 60%, and light-dark cycle was 12 h.
Mice were supplied with continuous access to drinking water and a
normal diet. After acclimatization for 1 week, the mice were
randomly divided into four groups: i) control group (n=10); ii) DM
group (n=10); iii) diabetic mice fed a high-lipid diet (HLD) (DM +
HLD group, n=10); and iv) diabetic mice fed a HLD and administered
NAC (DM + HLD + NAC group, n=10). Experimental diabetic mice were
achieved via an intraperitoneal injection of 50 mg/kg STZ for 5
consecutive days, whereas control mice were injected with citrate
buffer for 5 consecutive days. At day 7, those mice whose plasma
glucose was >300 mg/dl were deemed to be diabetic. In total, 20
diabetic mice were placed on a HLD containing 1.25% cholesterol and
10% coconut oil. A total of 10 mice from the DM + HLD group
received 2 mmol/l NAC in their drinking water, whereas the
remaining mice in DM + HLD group received normal drinking water.
All mice were fed for 12 weeks. After 12 weeks, blood was obtained
from the carotid artery under anesthesia with ketamine (125 mg/kg,
intraperitoneal) and xylazine (6.25 mg/kg, intraperitoneal). The
doses of ketamine and xylazine were selected according to previous
studies (26–29), and were appropriately modified to
anesthetize mice and obtain blood for blood lipid, NO and MDA
assays. Subsequently, the mice were sacrificed by cervical
dislocation under deep anesthesia. The aortas were harvested and
immersed in liquid nitrogen for high-performance lipid
chromatography (HPLC), protein carbonyl contents assay, western
blotting and atherosclerotic lesion evaluation.
Cell culture
Human umbilical vein endothelial cells (HUVECs) were
isolated from neonatal umbilical cords, as described in our
previous study (30). HUVECs were
cultured in M199 (Gibco; Thermo Fisher Scientific, Inc.)
supplemented with 10% fetal bovine serum (Beijing Solarbio Science
& Technology Co., Ltd.), 1% insulin-transferrin-sodium selenite
(Sigma-Aldrich; Merck KGaA) and 1% penicillin-streptomycin at 37°C
in an atmosphere containing 5% CO2. HUVECs at passages
2–4 were divided into four groups: i) control group; ii) MG group,
where HUVECs were incubated with 1 mM MG for 24 h; iii) NAC group,
where HUVECs were pretreated with 1 mM NAC overnight, and then
incubated with 2 mM NAC and 1 mM MG for 24 h; and iv) BSO group,
where HUVECs were pretreated with 1 mM NAC and 50 µM BSO overnight,
and then incubated with 2 mM NAC, 300 µM BSO and 1 mM MG for 24 h.
For the reactive oxygen species (ROS) assay, 5×104
HUVECs were cultured in 96-well plates. For western blot analyses,
HUVECs were cultured in 6-well plates at 1×106
cells/well.
Evaluation of atherosclerotic
lesions
For the evaluation of aortic lesions, 5-µm
cryosections of the thoracic aortic root were stained with oil red
O at 37°C for 20 min, and were assessed using the Paigen's method
with Image Pro Plus 7.0 software (Media Cybernetics, Inc.)
(31). For each animal, five
sections were used to assess the mean lesion area under a light
microscope with Image-Pro Plus 7.0 software.
Blood lipid measurement
An automatic biochemistry analyzer (Olympus AU2700;
Olympus Corporation) was used to measure the levels of total
cholesterol (TC), triglyceride (TG) and high-density lipoprotein
cholesterol (HDL-C), according to the manufacturer's protocols. The
levels of low-density lipoprotein cholesterol (LDL-C) were
calculated using the Friedewald formula (32).
HPLC for quantification of GSH and MG
in the aorta
Determination of GSH was performed as previously
described (22,33). Briefly, trichloroacetic acid
(TCA)-soluble acid supernatants of the aorta in PBS were derived
using 6 mmol/l iodoacetic acid (pH 7–8) and 1% 2,4-dinitrophenyl
fluorobenzene (pH 7.0) to produce S-carboxymethyl and
2,4-dinitrophenyl derivatives, respectively. GSH derivatives were
separated using buffer [solvent A, 4:1 methanol:water (v/v);
solvent B, 272 g sodium acetate trihydrate, 122 ml water and 378 ml
glacial acetic acid were mixed, and 200 ml of the resulting
solution was added to 800 ml solvent A] on a 250×4.6-mm Alltech
Lichrosorb NH2 10 µm anion-exchange column (LC-20AB; Shimadzu
Corporation) and detected at UV-365 nm at room temperature [sample
quantity, 50 µl; flow rate, 1 ml/min; solvent A/solvent B, 90/10;
running time 30 min; internal standards, GSH and oxidized
glutathione (GSSG)]. The aortic GSH contents, expressed as nmol/mg
protein, were quantified by comparison to standard derivatives in
the same manner.
Determination of MG was performed as previously
described (22,34). Briefly, aortic homogenates in PBS
were treated with 60% perchloric acid (29:1 v/v), and the acidic
supernatants were derivatized with 0.1 mol/l o-phenylenediamine
(100:1 v/v) for 24 h. MG derivatives were separated on a 250×4.6-mm
Chromegabond Ultra C-18 reversed phase column (Shimadzu
Corporation), and detected at UV-315 nm. Aortic MG contents,
expressed as nmol/mg protein, were quantified by comparison to MG
standard derivatives with o-phenylenediamine.
Assessment of protein carbonyls
Total protein carbonyls were determined in aortic
homogenates as previously described (35). Briefly, homogenates were incubated
with 10 mmol/l 2,4-dinitrophenylhydrazine in 2 mol/l hydrochloric
acid (1:2 v/v) and precipitated with 10% TCA, and then centrifuged
at 11,000 × g at room temperature for 3 min. The supernatants were
removed and the pellets were washed with 1 ml ethanol-ethyl acetate
(1:1) at room temperature (1:1 volume ratio) three times (10 min
each). Then, protein pellets were re-dissolved in 600 µl of 6 mol/l
guanidine hydrochloride containing 20 mmol/l potassium phosphate
(pH 2.3) and incubated for 15 min at 37°C, and supernatants were
obtained following centrifugation at 11,000 × g at room temperature
for 3 min. Absorbance of the supernatants was measured at 366 nm.
Protein carbonyl contents were quantified using an extinction
coefficient of 22,000 M−1 cm−1.
ELISA for determination of serum NO
and MDA levels
Whole blood was placed for 2 h at room temperature
and centrifuged to obtain serum at 1,500 × g for 10 min at 4°C.
Serum NO and MDA levels were detected using commercial kits,
according to the manufacturer's protocols. The kits were based on
colorimetric methods and the samples were evaluated using a
microplate reader.
Measurement of intracellular ROS
levels
Intracellular ROS levels were measured using
2′,7′-dichlorofluorescein diacetate (DCFH-DA), according to the
method described by Zhu et al (36); briefly, 10 µM DCFH-DA was added to
the medium. DCFH-DA is converted to dichlorofluorescein (DCF) in
proportion to ROS concentration and remains trapped inside the
cell. Prior to fluorescence analysis, cells were incubated with
DCFH-DA for 30 min at 37°C in the dark and washed three times with
1X PBS. Subsequently, DCF fluorescence was determined using
excitation/emission wavelengths of 488/610 nm. ROS levels were
digitally quantified using Image Pro Plus 7.0 software (Media
Cybernetics, Inc.).
Western blot analysis of p-Akt,
p-eNOS, GPX-1 and SOD-1
Aortic tissues or HUVECs were homogenized with RIPA
lysis buffer containing 50 mM Tris (pH 8.0), 150 mM sodium
chloride, 0.5% sodium deoxycholate, 0.1% SDS and a protease
inhibitor cocktail (cat. no. P9599; Sigma-Aldrich; Merck KGaA).
Total proteins (30 µg) were separated by SDS-PAGE on 15% gels at
120 V for 140 min for actin, SOD-1 and GPX-1, and on 10% gels at
120 V for 120 min for actin, Akt, p-Akt, eNOS and p-eNOS. Proteins
were then transferred onto PVDF membranes at 100 V for 1 h. The
membranes were blocked in 5% skimmed milk in TBS with Tween-20
buffer [20 mM Tris, 137 mM NaCl and 0.1% Tween-20 (pH 7.6)] for 1 h
at room temperature, and were then incubated with rabbit anti-Akt
and p-Akt antibodies (1:500), rabbit anti-eNOS and p-eNOS
antibodies (1:500), rabbit anti-GPX-1 antibody (1:2,000), rabbit
anti-SOD-1 antibody (1:2,000) and mouse anti-actin antibody
(1:5,000) at 4°C overnight with agitation, followed by incubation
with the corresponding HRP-conjugated secondary antibody (1:5,000)
for 2 h at room temperature. Protein expression was detected using
ECL (Bio-Rad Laboratories, Inc.), according to the manufacturer's
instructions. Protein expression levels were normalized to actin
(in the case of SOD-1 and GPX-1), total Akt (in the case of p-Akt)
or total eNOS (in the case of p-eNOS) with Image-Pro Plus 7.0
software.
Statistical analysis
All experiments were repeated four to five times.
Data are presented as the mean ± SEM. Significant differences were
assessed by one-way ANOVA followed by Tukey's post hoc test (for
multiple comparisons) using GraphPad Prism 5 software (GraphPad
Software, Inc.). P<0.05 was considered to indicate a
statistically significant difference.
Results
NAC inhibits atherosclerotic
development in STZ-induced diabetic ApoE−/− mice fed a
HLD
At 12 weeks, the atherosclerotic plaque area of the
aortic root in diabetic mice was larger compared with that in the
normal diet control group (Fig. 1).
The atherosclerotic lesion area induced by the addition of a HLD
was 3-fold larger than the area induced by DM alone (Fig. 1). Conversely, NAC treatment for 12
weeks significantly attenuated the atheroma by nearly half in
diabetic mice fed a HLD compared with that in the DM + HLD group
(Fig. 1).
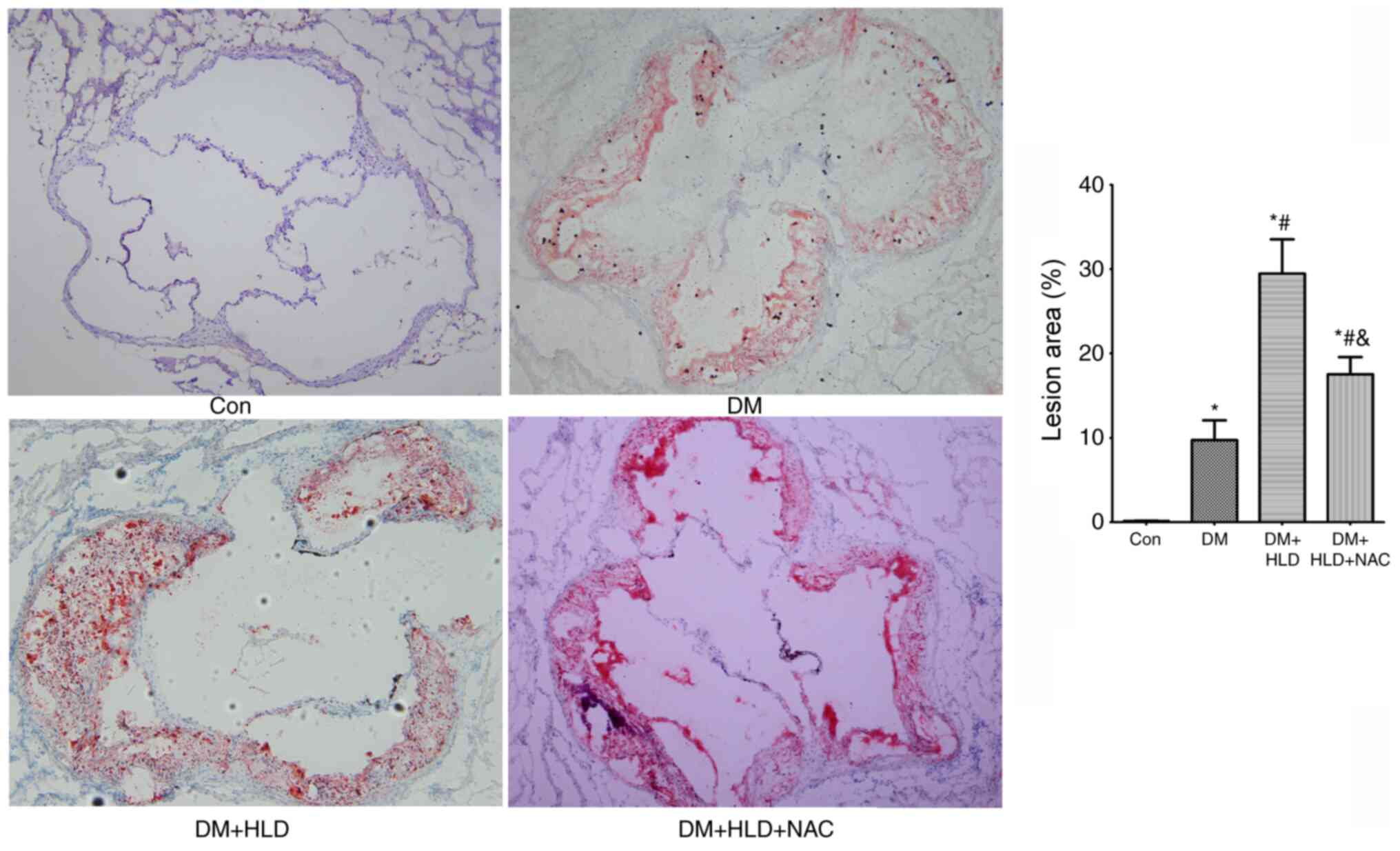 | Figure 1.Atherosclerotic lesion areas in the
aortic root were stained with oil red O (magnification, ×100) in
ApoE−/− mice (Con group), STZ-injected
ApoE−/− mice (DM group), STZ-injected
ApoE−/− mice fed a HLD (DM + HLD group), and
STZ-injected ApoE−/− mice fed a HLD and
administered NAC-containing water (DM + HLD + NAC group).
*P<0.05 vs. Con group; #P<0.05 vs. DM group;
&P<0.05 vs. DM + HLD group (n=5/group).
ApoE, apolipoprotein E; Con, control; DM, diabetes mellitus;
HLD, high-lipid diet; NAC, N-acetylcysteine; STZ,
streptozotocin. |
NAC does not alter blood lipid levels
or lipid profiles
High plasma LDL-C is a well-known risk factor for
the development and progression of atherosclerosis (37,38).
Low HDL-C is also a contributor to atherosclerotic lesions. To
ascertain if NAC affected cholesterol content to inhibit
atherosclerosis in DM, total lipids and their composition were
analyzed. The levels of TC, LDL-C and TG were elevated, whereas
HDL-C was decreased in diabetic mice at 12 weeks compared with the
lipid profiles in normal control mice. Mice fed a HLD for 12 weeks
exhibited further increased TC, LDL-C and TG levels compared with
DM group; however, this diet did not alter the levels of HDL-C in
diabetic mice. NAC treatment did not affect total lipid levels or
cholesterol composition compared with the observations in diabetic
mice fed a HLD (Fig. 2). These
results indicated that alteration of lipid or lipid profiles was
not associated with the mechanism underlying the
anti-atherosclerotic ability of NAC.
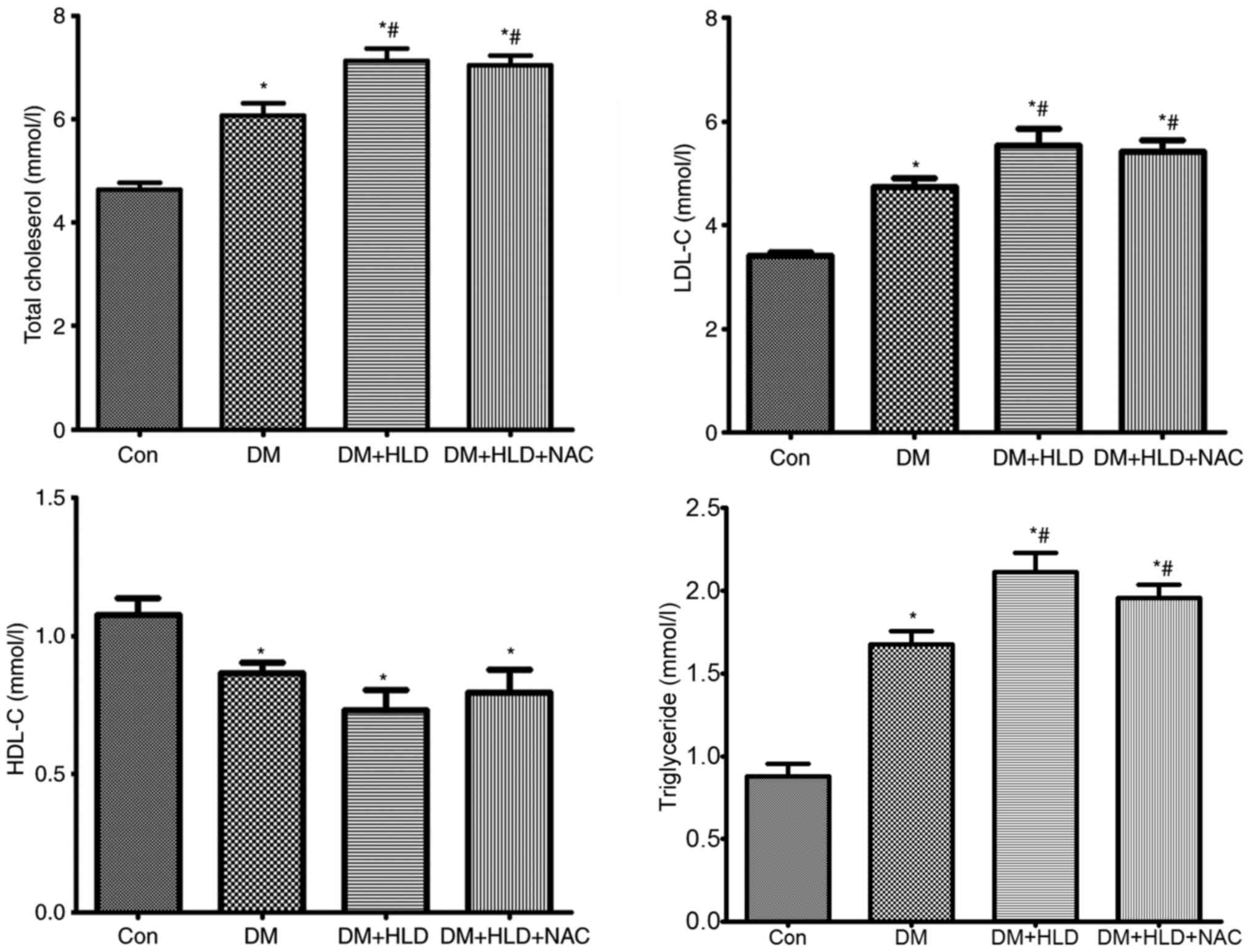 | Figure 2.Blood lipid levels (total
cholesterol, LDL-C, HDL-C and triglyceride) were assessed using an
automatic biochemistry analyzer in ApoE−/− mice
(Con group), STZ-injected ApoE−/− mice (DM
group), STZ-injected ApoE−/− mice fed a HLD (DM +
HLD group), and STZ-injected ApoE−/− mice fed a
HLD and administered NAC-containing water (DM + HLD + NAC group).
*P<0.05 vs. Con group; #P<0.05 vs. DM group
(n=8/group). LDL-C, low-density lipoprotein cholesterol; HDL-C,
high-density lipoprotein cholesterol; ApoE, apolipoprotein
E; Con, control; DM, diabetes mellitus; HLD, high-lipid diet; NAC,
N-acetylcysteine; STZ, streptozotocin. |
NAC corrects the upregulation of MG
and carbonyl proteins in diabetic ApoE−/− mice fed a
HLD
To ascertain whether NAC affected aortic MG and
dicarbonyl stress, MG and carbonyl protein levels were detected in
thoracic aorta samples. As expected, the MG and carbonyl protein
contents in the aorta of diabetic mice were increased ~3-fold
compared with those in the control group (Fig. 3A). Mice fed a HLD exhibited a slight
elevation in MG and carbonyl protein contents in the aorta compared
with those in the DM group; however, there were no significant
differences between the DM and DM + HLD groups. After treatment
with NAC for 12 weeks, DM + HLD + NAC mice exhibited reduced free
MG and protein carbonyl contents in the thoracic aorta compared
with those in the DM + HLD group (Fig.
3A). These results suggested that NAC corrected the increased
dicarbonyl stress detected in ApoE−/− mice in the
DM + HLD group.
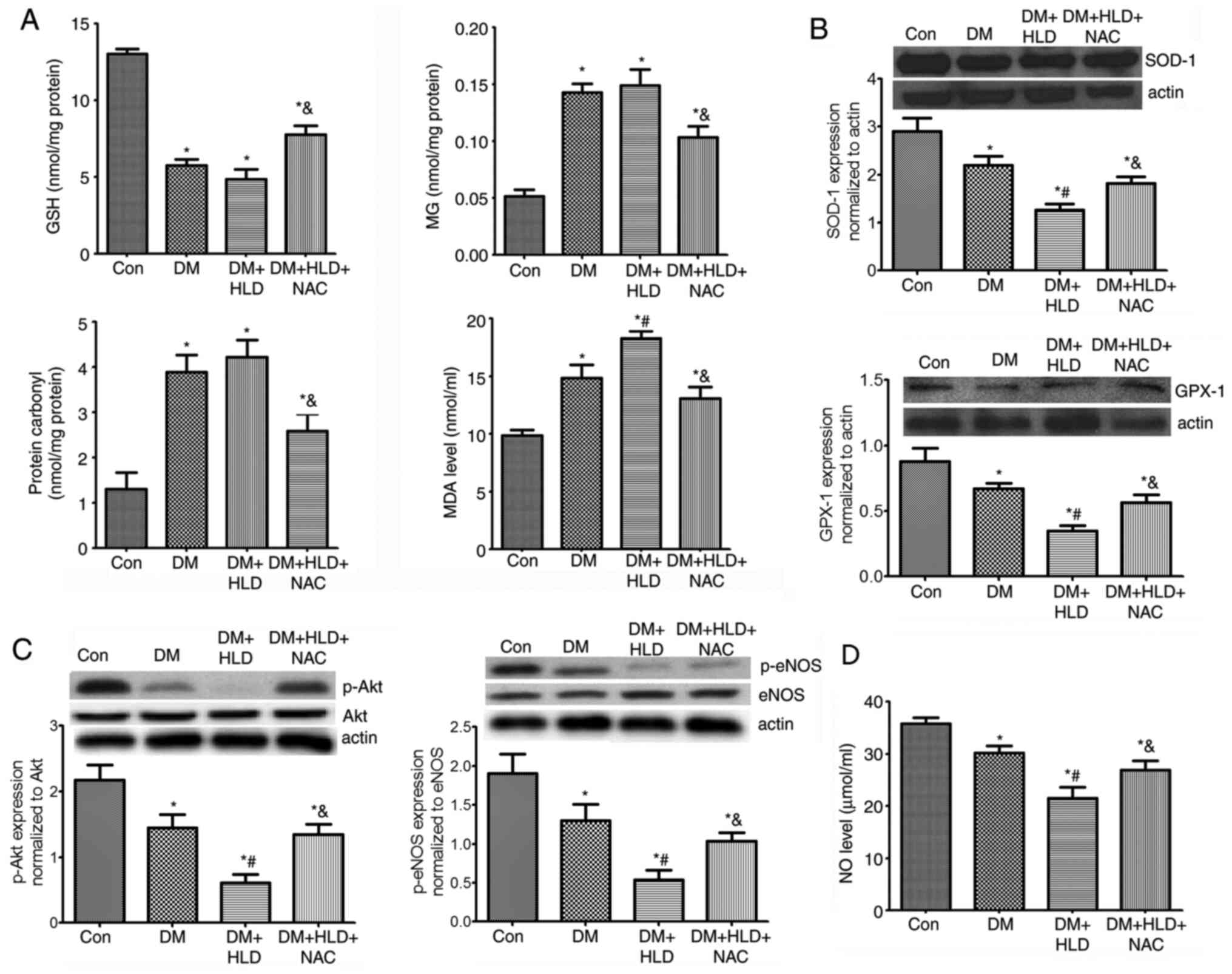 | Figure 3.Effects of NAC on oxidative stress,
p-Akt/p-eNOS protein expression and serum NO levels in diabetic
ApoE−/− mice fed a HLD. (A) Aortic protein
expression of GSH, MG, protein carbonyl contents and serum MDA
levels (n=5 repeats/group). (B) Aortic protein expression levels of
antioxidant enzymes (SOD-1 and GPX-1) (n=4 repeats/group). (C)
Aortic protein expression levels of p-Akt and p-eNOS (n=4
repeats/group). (D) Serum levels of NO (n=4 repeats/group) in
ApoE−/− mice (Con group), STZ-injected
ApoE−/− mice (DM group), STZ-injected
ApoE−/− mice fed a HLD (DM + HLD group), and
STZ-injected ApoE−/− mice fed a HLD and
administered NAC-containing water (DM + HLD + NAC group).
*P<0.05 vs. Con group; #P<0.05 vs. DM group;
&P<0.05 vs. DM + HLD group. ApoE,
apolipoprotein E; Con, control; DM, diabetes mellitus; HLD,
high-lipid diet; NAC, N-acetylcysteine; STZ, streptozotocin; GSH,
glutathione; MG, methylglyoxal; p-, phosphorylated; eNOS,
endothelial nitric oxide synthase; NO, nitric oxide. |
NAC scavenges oxidative stress in
diabetic ApoE−/− mice fed a HLD
To determine the levels of oxidative stress in DM,
serum MDA, and aortic SOD-1 and GPX-1 expression levels were
detected in mice. Serum MDA levels in DM mice were 1.5-fold higher
than those in the control group. Mice fed a HLD exhibited a further
increase in serum MDA levels compared with those detected in the DM
group (Fig. 3A). DM markedly
decreased aortic SOD-1 and GPX-1 protein expression levels, which
were further aggravated by a HLD. NAC treatment decreased serum
MDA, but increased aortic SOD-1 and GPX-1 expression levels
compared with the findings in ApoE−/− mice in the
DM + HLD group (Fig. 3A and B).
These findings indicated that oxidative stress was increased in
ApoE−/− mice in the DM + HLD group. Conversely,
NAC scavenged oxidative stress partly by decreasing serum MDA and
promoting the expression levels of antioxidant enzymes in diabetic
ApoE−/− mice fed a HLD.
NAC improves the reduction in aortic
p-Akt/p-eNOS protein expression, and corrects serum NO levels in
diabetic ApoE−/− mice fed a HLD
To further ascertain the endothelial function
affected by MG-dicarbonyl and oxidative stress, alterations in
p-Akt/eNOS and serum NO levels were detected. The phosphorylation
of Akt was significantly decreased in the aorta of mice in the DM
group compared with that in the aorta of control mice. The levels
of p-Akt in the aorta were further decreased in the DM + HLD group.
After treatment with NAC in diabetic ApoE−/− mice
fed a HLD, p-Akt expression was significantly enhanced compared
with that detected in the DM + HLD group. The changes in p-eNOS
expression in aorta samples were similar to those in p-Akt
expression. NO levels were significantly decreased in the DM group
compared with the control mice, which were further decreased in the
DM + HLD group. Whereas, after treatment with NAC, NO levels
increased. These results suggested that DM may damage endothelial
function and decrease serum NO levels, which were corrected by
treatment with NAC for 12 weeks via promoting Akt/eNOS
phosphorylation (Fig. 3C and
D).
MG induces ROS expression in
HUVECs
To explore whether MG induced oxidative stress, ROS
levels and antioxidant protein expression levels (SOD-1 and GPX-1)
were detected in HUVECs cultured with MG. In addition, p-Akt and
p-eNOS protein expression levels were detected in HUVECs (Fig. 4). As expected, 1 mM MG significantly
increased ROS, and decreased SOD-1 and GPX-1 expression levels
in vitro compared with in the control group. MG treatment
also markedly reduced p-Akt and p-eNOS protein expression levels in
HUVECs. The results in HUVECs were consistent with those in the
aortas of mice, and suggested that MG induced oxidative stress by
increasing ROS, and decreasing SOD-1 and GPX-1. Furthermore,
excessive MG may result in endothelial dysfunction by destructing
the p-Akt/p-eNOS pathway.
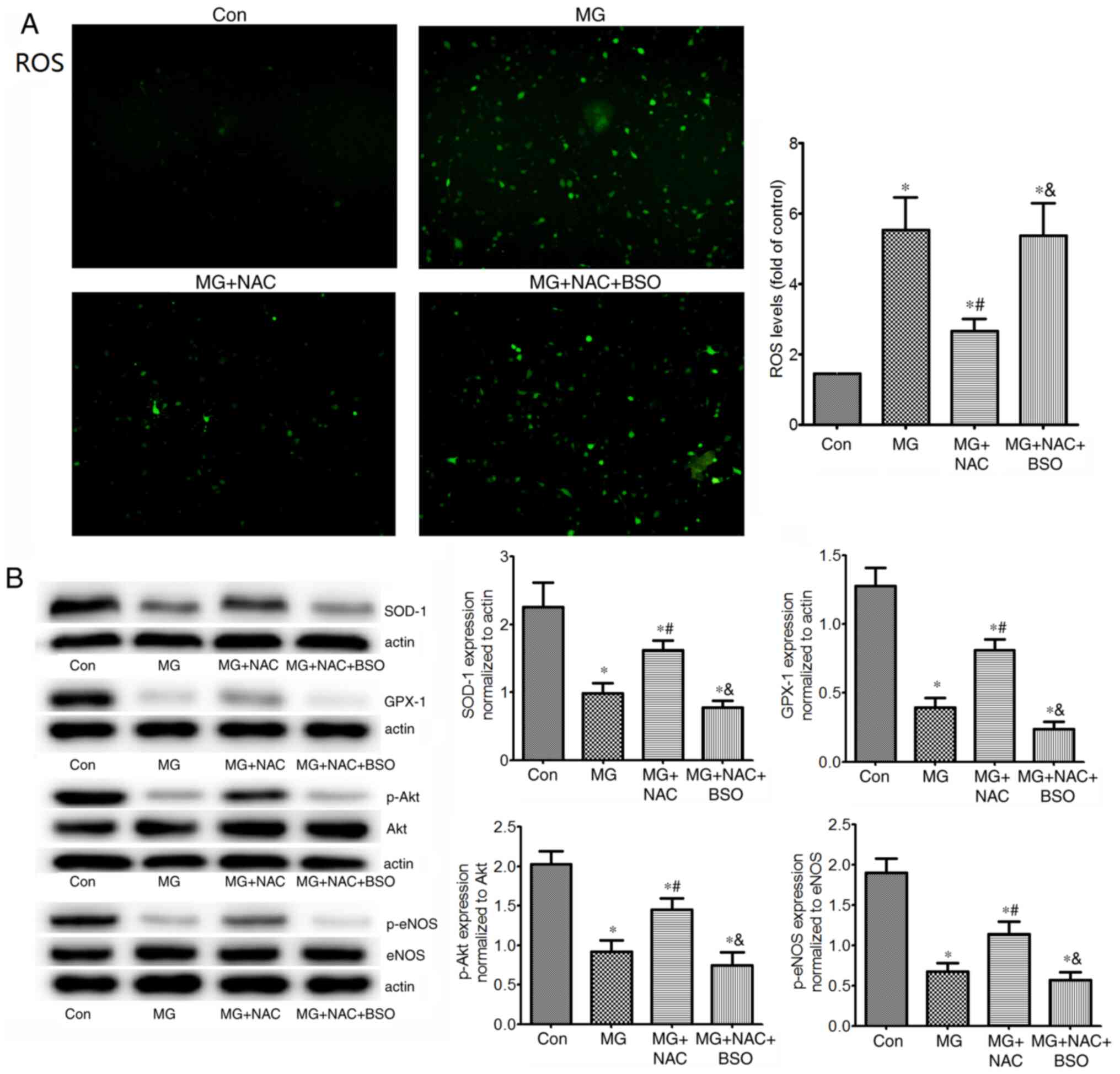 | Figure 4.Effects of NAC on ROS levels, SOD-1,
GPX-1 and p-Akt/p-eNOS protein expression in HUVECs. (A)
Intracellular ROS levels (magnification, ×100), and (B) protein
expression levels of antioxidant enzymes (SOD-1 and GPX-1), p-Akt
and p-eNOS in untreated HUVECs, or in HUVECs exposed to 1 mM MG,
with or without NAC, or with NAC + BSO (glutathione inhibitor).
Intracellular ROS levels were measured using
2′,7′-dichlorofluorescein diacetate. Protein expression levels were
assessed by western blotting (n=4 repeats/group). *P<0.05 vs.
control group; #P<0.05 vs. MG group;
&P<0.05 vs. MG + NAC group. HUVECs, human
umbilical vein endothelial cells; NAC, N-acetylcysteine; MG,
methylglyoxal; BSO, buthionine sulfoximine; ROS, reactive oxygen
species; SOD-1, superoxide dismutase 1; GPX-1, glutathione
peroxidase 1; p-, phosphorylated; eNOS, endothelial nitric oxide
synthase. |
Inhibition of GSH reverses the effects of NAC on
protection of HUVECs against MG-induced oxidative stress. GSH is a
rate-limiting enzyme in the elimination of MG. To determine the
role of GSH in the antioxidative effects of NAC, aortic GSH levels
were determined in mice. As expected, mice in the DM group
exhibited reduced levels of GSH in the aorta. In addition, a HLD
led to a slight reduction in aortic GSH compared with that in the
DM group; however, there was no significant difference between the
DM and DM + HLD groups. After treatment with NAC for 12 weeks,
diabetic ApoE−/− mice fed a HLD exhibited
enhanced aortic GSH levels compared with those in the DM + HLD
group (Fig. 3A). BSO is an
inhibitor of GSH (39). In
vitro, NAC significantly reduced MG-induced ROS elevation
compared with that in the MG group. However, administration of BSO
increased ROS to the levels of the MG group (Fig. 4A). These results indicated that NAC
inhibited MG-induced ROS by increasing GSH synthesis. In addition,
in cells treated with NAC, the protein expression levels of SOD-1,
GPX-1, p-Akt and p-eNOS were significantly increased compared with
those in the MG group (Fig. 4B).
Conversely, BSO reversed the effects of NAC on these proteins,
suggesting that the protective role of NAC on these proteins may be
due to increasing the levels of GSH.
Discussion
The present study demonstrated that mice in the DM
group exhibited a larger atherosclerotic area in the aorta compared
with that detected in control mice, which was associated with the
GSH-dependent ability to eliminate aortic MG. Additionally, the
elevation of MG in diabetic aortas was associated with an increase
in carbonyl and oxidative stress, as well as a decrease in serum NO
levels. NAC provides cysteine for GSH production, and may therefore
be effective at decreasing MG and oxidative stress, increasing
serum NO and preventing DM-induced atherosclerosis.
In the present study, MG levels in the aortas of
ApoE−/− mice injected with STZ were elevated
3-fold compared with those in the control mice. The increase in MG
levels in the aortas of experimental diabetic mice was consistent
with the elevated plasma MG levels that have previously been
described in this model (16).
Furthermore, GSH levels were markedly reduced in diabetic mice
aortas compared with those in the aortas of control mice, which
suggested that MG elimination was decreased in the aorta of
diabetic mice. Notably, neither MG nor GSH content in the aorta was
affected further by a HLD, suggesting that hyperglycemia rather
than hypercholesterolemia may be the major cause of alterations in
MG and GSH levels in the aorta. Although there are limited data to
support the alteration of MG in the aorta during DM, previous
studies have reported that hyperglycemia reduces GSH content in
aortic tissue (20,40), which is consistent with the present
results.
While anti-diabetic treatment can control the
increased production of MG caused by hyperglycemia, it is difficult
to correct for the decreased elimination of MG caused by low GSH
levels. NAC, a precursor of GSH, can increase MG elimination
(22). In the present animal study,
NAC treatment attenuated atherosclerosis in mice in the DM + HLD
group, indicating that aortic MG elevation may have an important
role in atherogenesis. NAC administration may therefore be
considered an effective anti-atherosclerotic method to decrease MG
levels and MG-dicarbonyl stress in the aorta.
Activation of receptor for advanced glycation end
products (RAGE) has been widely implicated in the
pro-atherosclerotic effects of MG, whereas RAGE deletion cannot
completely prevent vascular inflammation and damage (41). Increased oxidative stress has been
identified as a common upstream event that mediates the atherogenic
effects of hyperglycemia (42–44).
In line with this, diabetic ApoE−/− mice
exhibited higher serum MDA, and lower aortic SOD and GPX-1
expression levels compared with those in the control group. In
addition, a HLD aggravated oxidative stress. Conversely,
NAC-induced reduction of MG corrected the oxidative stress detected
in the aorta, suggesting that MG may induce aortic oxidative stress
in DM. This hypothesis was further validated by MG-treated HUVECs,
which exhibited an increase in ROS production and a decrease in
antioxidant levels. All of these effects were significantly
attenuated by pretreatment with NAC, but were blocked by inhibition
of GSH. These findings indicated that MG acted as a strong
stimulator that could induce an imbalance between ROS and
antioxidant enzymes, thus upregulating oxidative stress, whereas
NAC was dependent on GSH to reverse MG-induced dicarbonyl and
oxidative stress. Whereas NAC-restored GSH could act as an
antioxidant and/or induce elimination of MG, our previous findings
in platelets revealed that the protection offered by NAC in
STZ-treated mice was caused by its role as an eliminator of MG
instead of as an antioxidant, since GSSG levels were unchanged when
GSH and MG were significantly altered in platelets (45). In the present study, the GSSG levels
in the aorta were not detected. Whether NAC acts as an eliminator
of MG rather than as an antioxidant requires further
investigation.
Alterations in Akt/eNOS signaling, which can inhibit
ROS-induced endothelial damage and improve NO release, also serve
an important role in atherosclerosis (21,22).
The present study detected a reduction in aortic Akt
phosphorylation, and a subsequent decrease in eNOS phosphorylation
and serum NO levels in diabetic mice, suggesting endothelial
dysfunction in the DM group. These changes were further aggravated
by a HLD. NAC afforded protection to increase serum NO. In line
with the present results, a previous study revealed that MG
increased oxidative stress and/or AGEs formation alongside a
decrease in NO bioavailability, thus inducing or aggravating
endothelial dysfunction in normal Wistar rats or in Goto-Kakizaki
rats (7,18). Overall, based on the aforementioned
results, it may be hypothesized that MG-dicarbonyl stress induces
endothelial dysfunction by altering Akt and eNOS phosphorylation,
and aggravates endothelial dysfunction by increasing oxidative
stress. NAC may enhance the phosphorylation of Akt and increase NO
levels to reverse such dysfunction.
In the present study, lipids, particularly LDL-C,
were altered in mice in the DM and DM + HLD groups, whereas NAC had
no effect on blood lipid profiles. The present results differ from
those reported in other studies. For example, NAC has previously
been shown to reduce TC, TG, HDL and very-LDL levels without
affecting LDL-C levels in rats following a splenectomy (46). NAC also enhanced HDL-C levels in
patients with hyperlipidemia (47).
The possible reasons for these discrepancies may be different
experimental species, disease status and NAC dosage. Lipid
peroxidation by ROS has a critical role in atherogenesis,
particularly the early stage of atherosclerosis (48). However, the alteration of lipid
peroxidation was not measured in the present model and requires
further research in the future.
At present, no specific antioxidant treatment has
been recommended to prevent atherosclerotic progression (49). The present study confirmed that NAC
possessed anti-atherosclerotic potential in experimental DM via the
elevation of GSH in the aorta to inhibit dicarbonyl and oxidative
stress. This finding may lead to a novel antioxidant strategy in
the treatment of atherosclerosis in DM, in addition to the
treatment of established risk factors.
Acknowledgements
Not applicable.
Funding
The present study was supported by a grant from the
Hubei Natural Science Foundation of China (grant no.
WJ2017M117).
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
BW conceived and designed the experiments. SZ and MZ
performed the experiments. LL analyzed the data. XF performed the
experiments and drafted the paper. XF, SZ and BW confirm the
authenticity of all the raw data. All authors read and approved the
final manuscript.
Ethics approval and consent to
participate
The study was approved by the Ethics Committee of
Tongji Medical School, Huazhong University of Science and
Technology (Wuhan, China; approval no. S1297). Written informed
consent was obtained from patients prior to the use of umbilical
cord tissues.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
Glossary
Abbreviations
Abbreviations:
|
DM
|
diabetes mellitus
|
|
GPX-1
|
glutathione peroxidase-1
|
|
GSH
|
glutathione
|
|
GSSG
|
oxidized glutathione
|
|
HDL-C
|
high-density lipoprotein
cholesterol
|
|
HUVECs
|
human umbilical vein endothelial
cells
|
|
LDL-C
|
low-density lipoprotein
cholesterol
|
|
MG
|
methylglyoxal
|
|
MDA
|
malondialdehyde
|
|
NAC
|
N-acetylcysteine
|
|
NO
|
nitric oxide
|
|
p-eNOS
|
phosphorylated endothelial nitric
oxide synthase
|
|
ROS
|
reactive oxygen species
|
|
SOD-1
|
superoxide dismutase-1
|
|
STZ
|
streptozotocin
|
|
TC
|
total cholesterol
|
|
TG
|
triglyceride
|
References
|
1
|
Zimmet P, Alberti KG, Magliano DJ and
Bennett PH: Diabetes mellitus statistics on prevalence and
mortality: Facts and fallacies. Nat Rev Endocrinol. 12:616–622.
2016. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Bornfeldt KE: 2013 Russell Ross memorial
lecture in vascular biology: Cellular and molecular mechanisms of
diabetes mellitus-accelerated atherosclerosis. Arterioscler Thromb
Vasc Biol. 34:705–714. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Bairey Merz CN, Alberts MJ, Balady GJ,
Ballantyne CM, Berra K, Black HR, Blumenthal RS, Davidson MH, Fazio
SB, Ferdinand KC, et al American College of Cardiology Foundation;
American Heart Association; American College of Physicians Task
Force on Competence and Training (Writing Committee to Develop a
Competence and Training Statement on Prevention of Cardiovascular
Disease); American Academy of Neurology; American Association of
Cardiovascular and Pulmonary Rehabilitation; American College of
Preventive Medicine; American Diabetes Association; American
Society of Hypertension; Association of Black Cardiologists;
National Lipid Association; Preventive Cardiovascular Nurses
Association: ACCF/AHA/ACP 2009 competence and training statement: a
curriculum on prevention of cardiovascular disease: a report of the
American College of Cardiology Foundation/American Heart
Association/American College of Physicians Task Force on Competence
and Training (Writing Committee to Develop a Competence and
Training Statement on Prevention of Cardiovascular Disease):
developed in collaboration with the American Academy of Neurology;
American Association of Cardiovascular and Pulmonary
Rehabilitation; American College of Preventive Medicine; American
College of Sports Medicine; American Diabetes Association; American
Society of Hypertension; Association of Black Cardiologists;
Centers for Disease Control and Prevention; National Heart, Lung,
and Blood Institute; National Lipid Association; and Preventive
Cardiovascular Nurses Association, : J Am Coll Cardiol.
54:1336–1363. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Patel A, MacMahon S, Chalmers J, Neal B,
Billot L, Woodward M, Marre M, Cooper M, Glasziou P, Grobbee D, et
al ADVANCE Collaborative Group, : Intensive blood glucose control
and vascular outcomes in patients with type 2 diabetes. N Engl J
Med. 358:2560–2572. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Lachin JM, Genuth S, Cleary P, Davis MD
and Nathan DM; Diabetes Control and Complications
Trial/Epidemiology of Diabetes Interventions and Complications
Research Group, : Retinopathy and nephropathy in patients with type
1 diabetes four years after a trial of intensive therapy. N Engl J
Med. 342:381–389. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Fullerton B, Jeitler K, Seitz M, Horvath
K, Berghold A and Siebenhofer A: Intensive glucose control versus
conventional glucose control for type 1 diabetes mellitus. Cochrane
Database Syst Rev. (2):CD0091222014.
|
|
7
|
Schmidt AM and Stern D: Atherosclerosis
and diabetes: The RAGE connection. Curr Atheroscler Rep. 2:430–436.
2000. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Cushman WC, Evans GW, Byington RP, Goff DC
Jr, Grimm RH Jr, Cutler JA, Simons-Morton DG, Basile JN, Corson MA,
Probstfield JL, et al ACCORD Study Group, : Effects of intensive
blood-pressure control in type 2 diabetes mellitus. N Engl J Med.
362:1575–1585. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Rabbani N, Xue M and Thornalley PJ:
Methylglyoxal-induced dicarbonyl stress in aging and disease: First
steps towards glyoxalase 1-based treatments. Clin Sci (Lond).
130:1677–1696. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Turk Z: Glycotoxines, carbonyl stress and
relevance to diabetes and its complications. Physiol Res.
59:147–156. 2010.PubMed/NCBI
|
|
11
|
Watson AM, Soro-Paavonen A, Sheehy K, Li
J, Calkin AC, Koitka A, Rajan SN, Brasacchio D, Allen TJ, Cooper
ME, et al: Delayed intervention with AGE inhibitors attenuates the
progression of diabetes-accelerated atherosclerosis in diabetic
apolipoprotein E knockout mice. Diabetologia. 54:681–689. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Lapolla A, Flamini R, Dalla Vedova A,
Senesi A, Reitano R, Fedele D, Basso E, Seraglia R and Traldi P:
Glyoxal and methylglyoxal levels in diabetic patients: Quantitative
determination by a new GC/MS method. Clin Chem Lab Med.
41:1166–1173. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
McLellan AC, Thornalley PJ, Benn J and
Sonksen PH: Glyoxalase system in clinical diabetes mellitus and
correlation with diabetic complications. Clin Sci (Lond). 87:21–29.
1994. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Rabbani N, Godfrey L, Xue M, Shaheen F,
Geoffrion M, Milne R and Thornalley PJ: Glycation of LDL by
methylglyoxal increases arterial atherogenicity: A possible
contributor to increased risk of cardiovascular disease in
diabetes. Diabetes. 60:1973–1980. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Nigro C, Leone A, Raciti GA, Longo M,
Mirra P, Formisano P, Beguinot F and Miele C:
Methylglyoxal-glyoxalase 1 balance: the root of vascular damage.
Int J Mol Sci. 18:1882017. View Article : Google Scholar
|
|
16
|
Tikellis C, Pickering RJ, Tsorotes D, Huet
O, Cooper ME, Jandeleit-Dahm K and Thomas MC: Dicarbonyl stress in
the absence of hyperglycemia increases endothelial inflammation and
atherogenesis similar to that observed in diabetes. Diabetes.
63:3915–3925. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Wu L and Juurlink BH: Increased
methylglyoxal and oxidative stress in hypertensive rat vascular
smooth muscle cells. Hypertension. 39:809–814. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Sena CM, Matafome P, Crisóstomo J,
Rodrigues L, Fernandes R, Pereira P and Seiça RM: Methylglyoxal
promotes oxidative stress and endothelial dysfunction. Pharmacol
Res. 65:497–506. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Tachi Y, Okuda Y, Bannai C, Okamura N,
Bannai S and Yamashita K: High concentration of glucose causes
impairment of the function of the glutathione redox cycle in human
vascular smooth muscle cells. FEBS Lett. 421:19–22. 1998.
View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Tachi Y, Okuda Y, Bannai C, Bannai S,
Shinohara M, Shimpuku H, Yamashita K and Ohura K: Hyperglycemia in
diabetic rats reduces the glutathione content in the aortic tissue.
Life Sci. 69:1039–1047. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Sung HJ, Kim J, Kim Y, Jang SW and Ko J:
N-acetyl cysteine suppresses the foam cell formation that is
induced by oxidized low density lipoprotein via regulation of gene
expression. Mol Biol Rep. 39:3001–3007. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Wang B, Aw TY and Stokes KY: The
protection conferred against ischemia-reperfusion injury in the
diabetic brain by N-acetylcysteine is associated with decreased
dicarbonyl stress. Free Radic Biol Med. 96:89–98. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Soro-Paavonen A, Watson AM, Li J, Paavonen
K, Koitka A, Calkin AC, Barit D, Coughlan MT, Drew BG, Lancaster
GI, et al: Receptor for advanced glycation end products (RAGE)
deficiency attenuates the development of atherosclerosis in
diabetes. Diabetes. 57:2461–2469. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Kotur-Stevuljevic J, Memon L, Stefanovic
A, Spasic S, Spasojevic-Kalimanovska V, Bogavac-Stanojevic N,
Kalimanovska-Ostric D, Jelić-Ivanovic Z and Zunic G: Correlation of
oxidative stress parameters and inflammatory markers in coronary
artery disease patients. Clin Biochem. 40:181–187. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Institute of Laboratory Animal Resources
Commission on Life Sciences National Research Council, . Guide for
the Care and Use of Laboratory Animals. National Academy Press;
Washington, DC: pp. 8–78. 1996
|
|
26
|
Traslavina RP, King EJ, Loar AS, Riedel
ER, Garvey MS, Ricart-Arbona R, Wolf FR and Couto SS: Euthanasia by
CO2 inhalation affects potassium levels in mice. J Am
Assoc Lab Anim Sci. 49:316–322. 2010.PubMed/NCBI
|
|
27
|
Overmyer KA, Thonusin C, Qi NR, Burant CF
and Evans CR: Impact of anesthesia and euthanasia on metabolomics
of mammalian tissues: Studies in a C57BL/6J mouse model. PLoS One.
10:e01172322015. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Fuentes JM, Talamini MA, Fulton WB, Hanly
EJ, Aurora AR and De Maio A: General anesthesia delays the
inflammatory response and increases survival for mice with
endotoxic shock. Clin Vaccine Immunol. 13:281–288. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Yang XP, Liu YH, Rhaleb NE, Kurihara N,
Kim HE and Carretero OA: Echocardiographic assessment of cardiac
function in conscious and anesthetized mice. Am J Physiol.
277:H1967–H1974. 1999.PubMed/NCBI
|
|
30
|
Guan S and Wang B: Effects of fosinopril
and valsartan on expressions of ICAM-1 and NO in human umbilical
vein endothelial cells. Chin Med J (Engl). 116:923–927.
2003.PubMed/NCBI
|
|
31
|
Paigen B, Morrow A, Holmes PA, Mitchell D
and Williams RA: Quantitative assessment of atherosclerotic lesions
in mice. Atherosclerosis. 68:231–240. 1987. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Friedewald WT, Levy RI and Fredrickson DS:
Estimation of the concentration of low-density lipoprotein
cholesterol in plasma, without use of the preparative
ultracentrifuge. Clin Chem. 18:499–502. 1972. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Reed DJ, Babson JR, Beatty PW, Brodie AE,
Ellis WW and Potter DW: High-performance liquid chromatography
analysis of nanomole levels of glutathione, glutathione disulfide,
and related thiols and disulfides. Anal Biochem. 106:55–62. 1980.
View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Dhar A, Desai K, Liu J and Wu L:
Methylglyoxal, protein binding and biological samples: Are we
getting the true measure? J Chromatogr B Analyt Technol Biomed Life
Sci. 877:1093–1100. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Levine RL, Garland D, Oliver CN, Amici A,
Climent I, Lenz AG, Ahn BW, Shaltiel S and Stadtman ER:
Determination of carbonyl content in oxidatively modified proteins.
Methods Enzymol. 186:464–478. 1990. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Zhu ZX, Cai WH, Wang T, Ye HB, Zhu YT, Chi
LS, Duan YM, Sun CC, Xuan YH and Jin LT: bFGF-regulating MAPKs are
involved in high glucose-mediated ROS production and delay of
vascular endothelial cell migration. PLoS One. 10:e01444952015.
View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Kannel WB, Castelli WP, Gordon T and
McNamara PM: Serum cholesterol, lipoproteins, and the risk of
coronary heart disease. The Framingham study. Ann Intern Med.
74:1–12. 1971. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Keys A: Coronary heart disease in seven
countries. 1970. Nutrition. 13:250–252; discussion 249, 253. 1997.
View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Valdovinos-Flores C, Limón-Pacheco JH,
León-Rodríguez R, Petrosyan P, Garza-Lombó C and Gonsebatt ME:
Systemic L-buthionine-S-R-sulfoximine treatment increases plasma
NGF and upregulates L-cys/L-cys2 transporter and γ-glutamylcysteine
ligase mRNAs through the NGF/TrkA/Akt/Nrf2 pathway in the striatum.
Front Cell Neurosci. 13:3252019. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Nascimento NR, Costa-e-Forti A, Peter AA
and Fonteles MC: Free radical scavengers improve the impaired
endothelium-dependent responses in aorta and kidneys of diabetic
rabbits. Diabetes Res Clin Pract. 61:145–153. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Muniyappa R and Srinivas PR: Dicarbonyl
stress and atherosclerosis: Is it all RAGE? Diabetes. 63:3587–3589.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Serafini M and Del Rio D: Understanding
the association between dietary antioxidants, redox status and
disease: is the Total Antioxidant Capacity the right tool? Redox
Rep. 9:145–152. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Peluso I, Morabito G, Urban L, Ioannone F
and Serafini M: Oxidative stress in atherosclerosis development:
The central role of LDL and oxidative burst. Endocr Metab Immune
Disord Drug Targets. 12:351–360. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Matough FA, Budin SB, Hamid ZA, Alwahaibi
N and Mohamed J: The role of oxidative stress and antioxidants in
diabetic complications. Sultan Qaboos Univ Med J. 12:5–18. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Wang B, Yee Aw T and Stokes KY:
N-acetylcysteine attenuates systemic platelet activation and
cerebral vessel thrombosis in diabetes. Redox Biol. 14:218–228.
2018. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Sit M, Yilmaz EE, Tosun M and Aktas G:
Effects of N-acetyl cysteine on lipid levels and on leukocyte and
platelet count in rats after splenectomy. Niger J Clin Pract.
17:343–345. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Franceschini G, Werba JP, Safa O, Gikalov
I and Sirtori CR: Dose-related increase of HDL-cholesterol levels
after N-acetylcysteine in man. Pharmacol Res. 28:213–218. 1993.
View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Violi F, Loffredo L, Carnevale R,
Pignatelli P and Pastori D: Atherothrombosis and oxidative stress:
mechanisms and management in elderly. Antioxid Redox Signal.
27:1083–1124. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Pignatelli P, Menichelli D, Pastori D and
Violi F: Oxidative stress and cardiovascular disease: New insights.
Kardiol Pol. 76:713–722. 2018. View Article : Google Scholar : PubMed/NCBI
|














