Introduction
Retinoblastoma (RB) is a specific type of malignant
tumor that is often characterized by features of occult seizure,
rapid growth, and the ability for intracranial and distant
metastasis (1,2). RB is the most common ocular malignancy
diagnosed during childhood, and accounts for most intraocular
tumors found in children (3,4). The
mortality rate of patients with RB in developing countries is
50–70% (5,6). Children suffering from RB are also
susceptible to other diseases, including RB cell invasion,
malignant transformation of an intracranial neuroblastoma, and
development of a heterochronogenous tumor (7). In recent years, various novel surgical
techniques and individualized comprehensive therapies have
demonstrated efficacy in the clinical treatment of RB. However, the
survival rate of patients with RB remains very low, mainly because
the disease is often diagnosed in its later stages (8). Moreover, the traditional therapies for
RB, such as external beam radiotherapy and chemotherapy, often
produce side effects, including infection and blindness (9). Thus, in-depth research into the
molecular mechanisms underlying RB is of great clinical
significance for controlling the growth of the tumor and improving
the prognosis of patients.
Class A scavenger receptors (SRs) are essential cell
surface receptors that can bind various types of ligands, such as
nucleic acids and modified low-density lipoproteins (10). These receptors are known to have an
important role in host defense and are characterized as containing
a collagen-like domain (11). SR
class A member 5 (SCARA5), a member of the SR family, is mainly
expressed on epithelial cells associated with mucosal surfaces,
suggesting that it participates in the innate immune response
(11). Abnormally low expression
levels of SCARA5 have been reported in various types of cancer,
such as lung cancer, liver cancer, renal cancer and glioma
(12–15). Inhibition of SCARA5 expression by
Snail1 was previously shown to markedly attenuate the
epithelial-to-mesenchymal transition-associated migration of human
non-small cell lung carcinoma A549 cells (12). Another study reported that SCARA5
knockdown markedly enhanced human hepatocellular carcinoma
tumorigenesis and metastasis by activating the FAK signaling
pathway (13). Moreover, the
proliferation of renal cell carcinoma cells with SCARA5 knockdown
was promoted when Rho-associated coiled-coil forming protein kinase
2 (Rock2) was overexpressed (14).
In addition, the systemic treatment of tumor-bearing mice with a
SCARA5-cationic liposome complex not only reduced the growth of
subcutaneous human glioma tumors, but also markedly suppressed the
spontaneous formation of lung metastases (15). Consistent with these findings,
SCARA5 knockdown was shown to promote cell cycle progression in
hepatocellular carcinoma cells, whereas increased SCARA5 expression
inhibited cell cycle progression (16). These findings suggested that SCARA5
may have an important role in cancer progression. However, to the
best of our knowledge, its role in the progression and metastasis
of RB remains to be fully elucidated.
The present study investigated the precise role of
SCARA5 in RB tumor growth and metastasis, and clarified its
potential mechanism of action in RB. The results of the present
study may provide a novel therapeutic target for treating patients
with RB.
Materials and methods
Cell lines and cell culture
The human retinal pigment epithelial cell line
ARPE-19, and human RB cell lines HXO-Rb44, Y79, SO-RB50
and WERI-Rb-1, were purchased from the Institute of Biochemistry
and Cell Biology of the Chinese Academy of Sciences. The cell lines
(1×105 × cells) were cultured in RPMI-1640 medium
(Gibco; Thermo Fisher Scientific, Inc.) supplemented with 10% FBS
(Gibco; Thermo Fisher Scientific, Inc.), 100 U/ml penicillin and
100 µg/ml streptomycin at 37°C in a 5% CO2 incubator
with 95% humidity.
Vector construction and
transfection
The pcDNA3.1 vector carrying full length of SCARA5
and the pGCSIL-GFP plasmid carrying a short hairpin (sh) RNA
targeting SCARA5 were used for overexpression or knockdown of
SCARA5, respectively (both from GenScript). Y79 cells were seeded
in a six-well plate at a density of 3×105 cells/well
prior to transfection. The pcDNA3.1-SCARA5 and sh-SCARA5 plasmids
(2 µg) were transfected into Y79 cells when cell confluence reached
60–70% using Lipofectamine® 2000 (Invitrogen; Thermo
Fisher Scientific, Inc.) according to the manufacturer's
instructions. Empty pcDNA3.1 vector and pGCSIL-GFP plasmid carrying
a non-targeting control sh-RNA sequence (sh-control) were used as
controls for SCARA5 overexpression and knockdown, respectively. Y79
cells were cultured with different treatment were cultured in
RPMI-1640 medium (Gibco; Thermo Fisher Scientific, Inc.)
supplemented with 10% FBS (Gibco; Thermo Fisher Scientific, Inc.),
100 U/ml penicillin and 100 µg/ml streptomycin at 37°C in a 5%
CO2 incubator with 95% humidity for 48 h. Transfection
efficiency was detected at 48 h post-transfection using RT-qPCR and
western blotting, and the transfected Y79 cells were collected. The
short hairpin RNA (shRNA) used to knockdown SCARA5 expression was
designed and synthesized by Shanghai GenePharma Co., Ltd. The
SCARA5 and control shRNA sequences were as follows: SCARA5,
5′-GCUCCAUCUGUGAGGAUUCdTdT-3′; control,
5′-UUCCCGAACGUGUCACGUTT-3′.
Reverse transcription quantitative-PCT
(RT-qPCR)
Total RNA was extracted from cells or tumor tissues
using TRIzol® reagent (Invitrogen; Thermo Fisher
Scientific, Inc.) according to the manufacturer's instructions.
Subsequently, 2 µg total RNA was reverse transcribed into cDNA
using a PrimeScript RT Reagent kit (Invitrogen; Thermo Fisher
Scientific, Inc.) according to the manufacturer's instructions.
RT-qPCR was performed using an ABI PRISM 7900 Sequence Detector
(Applied Biosystems; Thermo Fisher Scientific, Inc.) and a SYBR
Green PCR kit (Applied Biosystems Biosystems; Thermo Fisher
Scientific, Inc.). GAPDH served as an internal control.
Thermocycling parameters for the amplification were as follows:
95°C, 10 min; followed by 95°C, 15 sec and 60°C, 1 min for 40
cycles. Relative expression levels of targeted genes were
calculated by 2−ΔΔCt method as previously described
(17). The primer sequences used
were as follows: SCARA5 forward, 5′-AAAGCTATGTACCTACACACCGT-3′ and
reverse, 5′-CCGCCGTTTGTGACATGGA-3′; GAPDH forward,
5′-TGTTCGTCATGGGTGTGAAC-3′ and reverse,
5′-ATGGCATGGACTGTGGTCAT-3′.
Western blotting
Total proteins were extracted from cells or tumor
tissues using RIPA lysis buffer containing 1% protease inhibitor
cocktail (cat. no. R0278-50ML, Sigma-Aldrich; Merck KGaA). The
total protein concentration in each extract was quantified using a
bicinchoninic acid protein assay kit. Subsequently, 20 µg protein
from each sample was mixed with protein loading buffer and heated
at 100°C for 5 min. The protein samples were then separated by
SDS-PAGE on 10–12% gels and the separated protein bands were
transferred onto PVDF membranes (EMD Millipore). Membranes were
subsequently blocked with 5% fat-free buffered milk for 2 h at room
temperature and were incubated with primary antibodies against PI3K
(1:1,000; rat monoclonal; cat. no. ABS1856; Sigma-Aldrich; Merck
KGaA), phosphorylated (p)-PI3K (1:1,000; cat. no. 04–1138; rat
monoclonal; Sigma-Aldrich; Merck KGaA), AKT (1:1,000; rabbit
polyclonal; cat. no. 4685; Cell Signaling Technology, Inc.), p-AKT
(1:1,000; rabbit polyclonal; cat. no. 4060; Cell Signaling
Technology, Inc.), SCARA5 (1:1,000; rabbit polyclonal; cat. no.
ab106439; Abcam), Bcl-2 (1:2,000 rabbit polyclonal; cat. no.
ab182858; Abcam), Bax (1:3,000; rabbit polyclonal; cat. no.
ab182734; Abcam), cleaved caspase-3 (1:1,000; rabbit polyclonal;
cat. no. ab49822; Abcam) and GAPDH (1:2,000; rabbit polyclonal;
cat. no. ab9485; Abcam) overnight at 4°C. Subsequently, the
membranes were incubated with an anti-rabbit IgG secondary antibody
(1:20,000; cat. no. ab6721; Abcam) or anti-rat IgG (1:20,000; cat.
no. ab125900; Abcam) for 2–3 h at room temperature; after which,
the immunostained protein bands were detected using an ECL reagent
(Thermo Fisher Scientific, Inc.) according to the manufacturer's
instructions. Densitometric analysis was performed by ImageJ V1.8.0
software (National Institutes of Health).
Immunofluorescence assay
Immunofluorescence assays were performed as
previously described (18).
Briefly, 1×103 cells were plated onto slides that had
been treated with polylysine and were incubated at 37°C for 4 h.
Subsequently, the cells were fixed with 10% formaldehyde and then
blocked with 5% bovine serum albumin (cat. no. 810784;
Sigma-Aldrich; Merck KGaA) in PBS for 2 h at room temperature.
Subsequently, the cells were stained with rabbit anti-human
polyclonal antibodies against SCARA5 (1:200; cat. no. ab106439;
Abcam) at 4°C overnight, and were incubated with a Cy3-conjugated
(red) anti-rabbit IgG antibody (1:1,000; cat. no. ab6939; Abcam) at
4°C for 2 h. After repeated rinsing in PBS, the cells were observed
under an inverted fluorescence microscope.
Cell proliferation assay
The cells were incubated for 5 h with
5-ethynyl-2′-deoxyuridine (EdU; Guangzhou Ribobio Co., Ltd.)
according to the manufacturer's protocol. Subsequently, cells were
washed three times with PBS and were then treated with 300 µl 1X
Apollo reaction cocktail for 30 min; after which, they were stained
with 100 µl EdU (5 µg/ml) for 30 min at room temperature, and their
DNA contents were detected with a fluorescence microscope. Cell
proliferation was also measured using a Cell Counting Kit 8 (CCK8)
kit (Sigma-Aldrich; Merck KGaA) according to the manufacturer's
instructions. Briefly, the cells were seeded onto 96-well plates
(Corning, Inc.) and cultured in medium at a concentration of
1×105 cells/ml. Subsequently, the cells were transfected
with vectors for 24, 48 and 72 h. CCK8 (10 µl) was then added to
the cells and the cells were cultured at 37°C for 4 h; after which,
cell proliferation was assessed with a microplate reader (Thermo
Fisher Scientific, Inc.) at a wavelength of 450 nm.
Transwell assay
Cell migration was evaluated using the Transwell
assay as previously described (19). Briefly, 5×104 cells
suspended in serum-free medium were placed into the upper chamber
of a Transwell plate (pore size, 8 µm; EMD Millipore), which was
not precoated with Matrigel, and medium containing 10% FBS was
added to the lower chamber. After 24 h, the cells which had
migrated into the lower chamber were fixed with 100% methanol and
stained with 0.1% crystal violet for 24 h at 37°C; after which,
they were observed and counted with an IX71 inverted light
microscope (Olympus Corporation). Each experiment was performed
three times.
Clone formation assay
Cells were seeded into the wells of 12-well plates
and cultured in complete medium for 2 weeks. The cells were then
fixed with 4% paraformaldehyde and stained with 0.1% crystal violet
for 15 min at 37°C. The number of colonies per well was then
counted.
Apoptosis assay
Apoptosis assays were performed using TUNEL and
Hoechst staining. TUNEL analysis was performed using a TUNEL
detection kit (cat. no. KGA702; Nanjing KeyGen Biotech Co., Ltd.)
according to the manufacturer's instructions. Briefly,
1×106 cells were suspended, permeabilized with 0.1%
Triton X-100 (Beyotime Institute of Biotechnology) for 30 min and
incubated in 50 mM TUNEL reaction mixture for 2 h at room
temperature. Slides were counterstained with DAPI for 10 min at
room temperature for nuclear staining. Images were captured using a
fluorescence microscope (Olympus Corporation). Apoptosis in tumor
tissues was also detected using a TUNEL assay according to the
manufacturer's instructions. Slides were stained with 10 µl
hematoxylin (cat. no. ab220365; Abcam) for 3 min at room
temperature for nuclear staining. Images were captured using a
XSP-36 light microscope (Boshida Optical Co., Ltd.). Each
experiment was performed three times. For Hoechst staining,
6×104 cells transfected cells were inoculated into the
wells of a 6-well plate. After removing the culture medium, the
culture plate was rinsed 3–5 times with PBS. Subsequently, the
cells in each well were fixed with 4% paraformaldehyde at 4°C for
20 min, and then rinsed with PBS. Approximately 1 ml Hoechst
staining buffer was then added to each well and the plate was
incubated for 20 min at room temperature. After being repeatedly
rinsed with PBS, the plate was observed under an inverted
fluorescence microscope, and the results were recorded for
subsequent statistical analysis.
Xenograft tumor experiment
A total of 15 male Balb/c nude mice (weight, 20–25
g; age, 3–4 weeks) were purchased from the Animal Center of Central
South University (Changsha, China). All of the nude mice were bred
in a specific-pathogen-free animal laboratory with 60–65% humidity
at 22–25°C under a 12-h light/dark cycle, and the experiment were
performed after 1 week of free access to food and water. Nude mice
were randomly divided into 3 groups with 5 in each group, including
Blank, Over-SCARA5, sh-SCARA5 groups. A xenograft tumor model was
created by subcutaneously injecting 3×106 cells
(untransfected, SCARA5-overexpressing or SCARA5-silenced Y79 cells)
into the flanks of nude mice. The size of each tumor was recorded
every 7 days, and images of each tumor were captured after 4 weeks.
At the end of the experiment, the nude mice were anesthetized by an
intraperitoneal injection of 3% sodium pentobarbital (50 mg/kg) and
in accordance with Hawkins P's animal experiment guidance, mice
were euthanized with 100% CO2, with a flow rate of 20%
chamber volume per minute (20).
During the present study, the Guide for the Care and Use of
Laboratory Animals (NRC 2011) of AAALAC was adhered to, in order to
minimize animal pain and discomfort (21). All study protocols involving animals
were approved by the Ethics Committee of Central South University
and were conducted at the Experimental Animal Center of Central
South University.
Immunohistochemistry
Tumor samples were fixed with 10% formalin for 24 h
at room temperature, embedded with paraffin and then cut into 4-µm
serial sections, which were used for immunohistochemistry.
Subsequently, deparaffinized and rehydrated sections were incubated
with 3% H2O2 in methanol for 10 min at room
temperature to block endogenous peroxidase activity. The sections
were then subjected to antigen retrieval for 10 min in a pressure
cooker containing sodium citrate, and were incubated in 5% normal
goat serum (cat. no. DXT-50197Z; Invitrogen; Thermo Fisher
Scientific, Inc.) for 20 min at 37°C, permeabilized in PBS-Triton
solution and incubated with a primary antibody against SCARA5
(1:200; cat. no. ab106439; Abcam) at 4°C overnight. The sections
were then incubated with a horseradish peroxidase-conjugated mouse
anti-rabbit secondary antibody (1:1,000; cat. no. ab6728; Abcam) at
37°C for 1 h.
For Ki67 staining, immunohistochemical analyses were
performed using anti-Ki67 (1:400; cat. no. SP6; Zytomed) which
incubated for 1 h at room temperature. The HRP-labeled goat
anti-rabbit IgG secondary antibody (1:1,000; cat. no. ab6721;
Abcam) was added to the sample, which was incubated for 10 min at
37°C. The sections were then counterstained with hematoxylin for 5
min at room temperature and finally dehydrated and covered with a
coverslip. The sections were observed under a XSP-36 light
microscope (Boshida Optical Co., Ltd.). Each experiment was
performed three times.
Statistical analysis
All statistical analyses were performed using
GraphPad Prism 6 software (GraphPad Software, Inc.) and results are
shown as the mean ± SEM. Unpaired Student's t-tests and one-way
ANOVA followed by the Student-Newman-Keuls post hoc test were
performed to compare the two groups and multiple groups,
respectively. P<0.05 was considered to indicate a statistically
significant difference.
Results
SCARA5 is downregulated in RB cell
lines
In order to determine the role played by SCARA5 in
the development and progression of RB, RT-qPCR was performed to
measure the mRNA expression levels of SCARA5 in RB cells. The
results revealed that SCARA5 mRNA was expressed at lower levels in
the RB cell lines (HXO-Rb44, Y79, SO-RB50 and WERI-Rb-1)
compared with in the healthy human retinal pigment epithelial cell
line (ARPE-19) (Fig. 1A).
Consistent with the difference in mRNA expression levels, western
blot analysis revealed that the protein expression levels of SCARA5
were also downregulated in the RB cell lines compared with those in
the healthy retinal pigment epithelial cell line (Fig. 1B). These results suggested that
SCARA5 may participate in RB development as a tumor suppressor. As
SCARA5 expression was markedly downregulated in the Y79 cell line
compared with that in the ARPE-19 cell line, and due to the
convenience to construct transfected Y79 cells, this cell line was
selected for subsequent experiments.
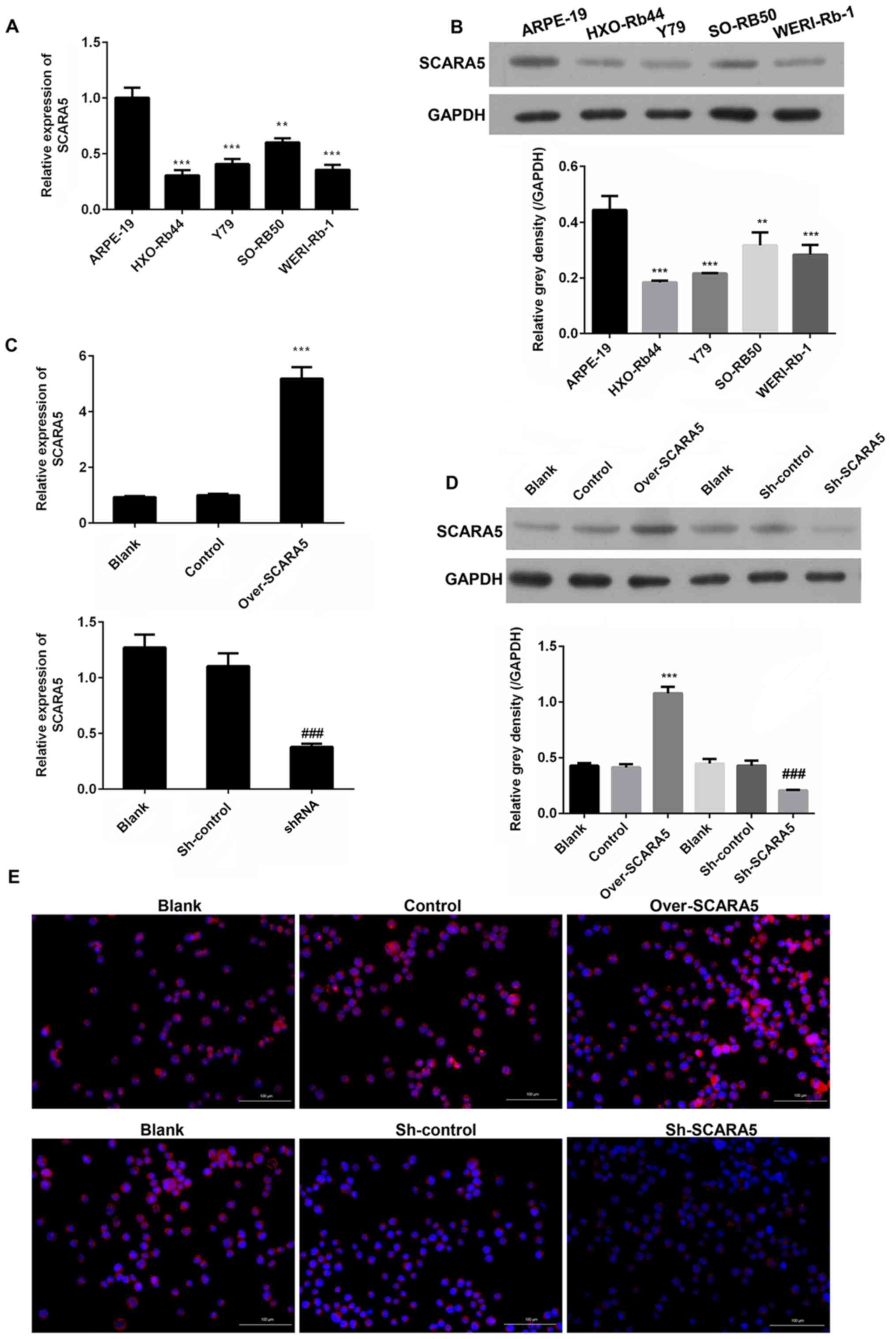 | Figure 1.Comparison of SCARA5 expression
levels in four human retinoblastoma cell lines
(HXO-Rb44, Y79, SO-RB50, WERI-Rb-1) and a human retinal
pigment epithelial cell line (ARPE-19). (A) RT-qPCR analysis of
SCARA5 expression. (B) Western blot analysis of SCARA5 expression.
**P<0.01, ***P<0.001 vs. ARPE-19. (C) RT-qPCR analysis of
SCARA5 expression in Y79 cells with SCARA5 overexpression or
knockdown. (D) Western blot analysis of SCARA5 expression in Y79
cells with SCARA5 overexpression or knockdown. Data are presented
as the mean ± SEM (n=3 with three repeats). ***P<0.001 vs.
Control; ###P<0.001 vs. sh-control. (E)
Immunofluorescence analysis of SCARA5 expression in Y79 cells with
SCARA5 overexpression or knockdown. Blue signal indicated the
nuclei stained by 4,6-diamino-2-phenyl indole (DAPI); Pink signal
was produced by the red immunofluorescence (Cy3) signal merging
with blue signal stained by DAPI. Scale bar, 100 µm. SCARA5,
scavenger receptor class A member 5; RT-qPCR, reverse
transcription-quantitative PCR; NC, negative control; shRNA/sh,
short hairpin RNA. |
Overexpression and knockdown of SCARA5
in Y79 cells
Y79 cells were used for transfection experiments
designed to investigate the role played by SCARA5 in RB
progression. Y79 cells were transfected with recombinant vectors
that induced overexpression or knockdown of SCARA5, and the
cellular expression levels of SCARA5 mRNA and protein were detected
by RT-qPCR and western blotting, respectively. As shown in Fig. 1C and D, when compared with the blank
control group, transfection with the SCARA5 vector significantly
upregulated the mRNA and protein expression levels of SCARA5,
whereas transfection with the SCARA5 knockdown vector produced the
opposite result. Consistent with these findings, immunofluorescence
analyses of SCARA5 protein expression revealed that SCARA5 protein
expression levels were upregulated or downregulated after the cells
had been transfected with the recombinant SCARA5 vector or SCARA5
knockdown vector, respectively (Fig.
1E). These results confirmed that SCARA5 was successfully
enhanced or suppressed by transfection with the respective
recombinant vectors.
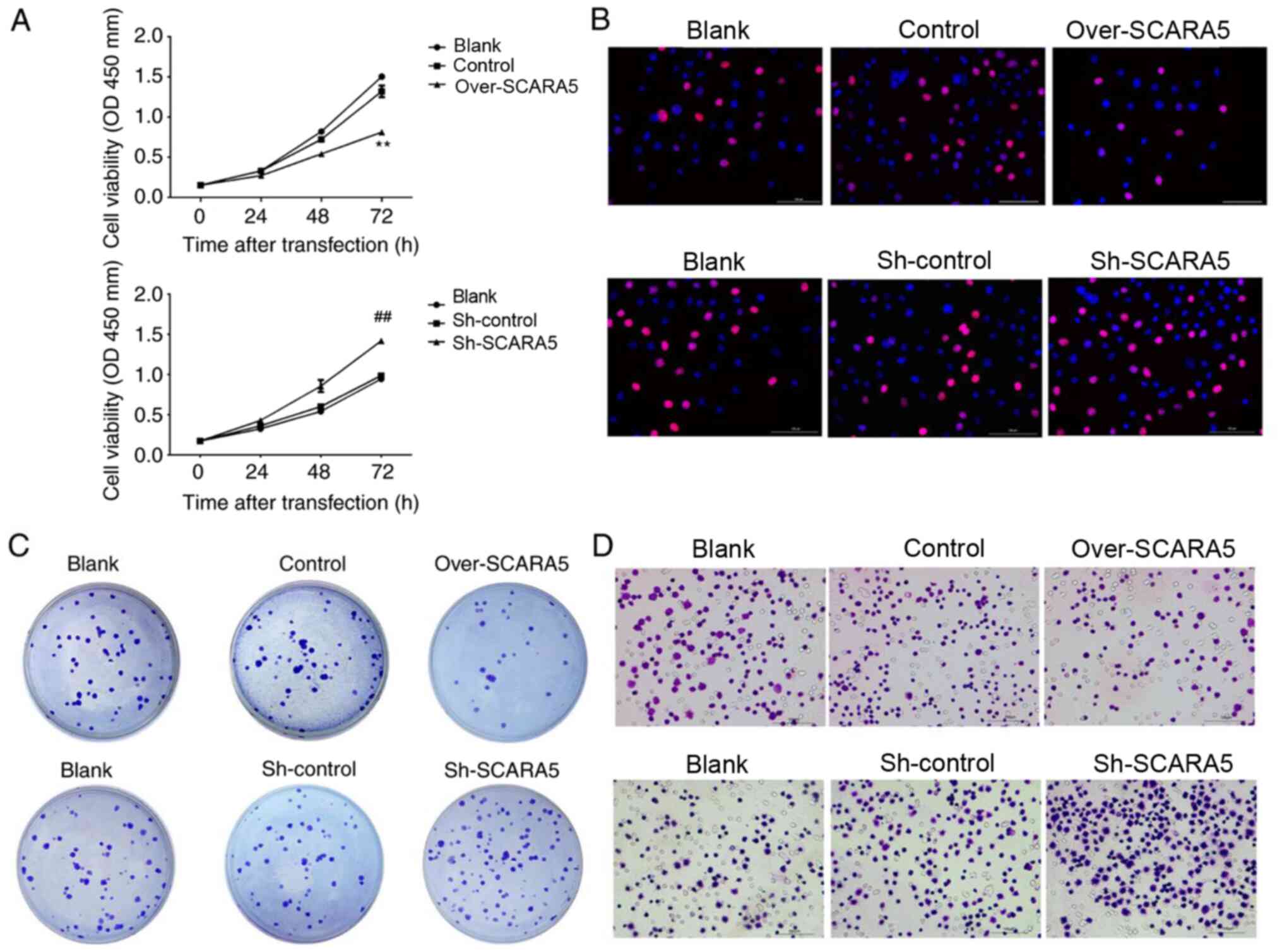
Overexpression of SCARA5 inhibits the
proliferation and migration of Y79 cells
To assess the biological effect of SCARA5 on RB cell
proliferation, CCK8 assays were performed to explore the effect of
SCARA5 overexpression or knockdown on RB cell proliferation. As
shown in Fig. 2A, when compared
with the control group, overexpression of SCARA5 significantly
inhibited the proliferation of Y79 cells, whereas knockdown of
SCARA5 significantly promoted the proliferation of Y79 cells at 72
h. Furthermore, Edu assay results confirmed that overexpression of
SCARA5 inhibited the growth rate of Y79 cells, when compared with
non-transfected Y79 cells (Fig.
2B). Consistent with these findings, the colony formation
assays revealed that compared with in the control group, the number
of cell colonies formed in the SCARA5 knockdown group was
increased. Conversely, SCARA5 overexpression decreased the number
of cell colonies that formed (Fig.
2C). The migratory abilities of Y79 cells were evaluated by
Transwell assay. The results revealed that overexpression of SCARA5
reduced the migratory capabilities of Y79 cells when compared with
control cells, whereas those migratory abilities were increased in
the knockdown cells when compared with the control cells (Fig. 2D). These results demonstrated that
SCARA5 suppressed the proliferation and migration of RB cells.
Overexpression of SCARA5 promotes the
apoptosis of Y79 cells
To assess the effect of SCARA5 on apoptosis of RB
cells, Hoechst 33342 and TUNEL staining were used to explore the
effect of SCARA5 overexpression or knockdown on Y79 cell apoptosis.
The present results demonstrated that the percentage of apoptotic
cells was increased in the SCARA5 overexpression group compared
with that in the control group (Fig.
3A). Conversely, SCARA5 knockdown decreased the proportion of
apoptotic cells, as evidenced by decreased fluorescence intensity
in the knockdown group compared with in the control group. In
addition, compared with in the control group, overexpression of
SCARA5 promoted apoptosis of Y79 cells whereas knockdown of SCARA5
suppressed cell apoptosis (Fig.
3B).
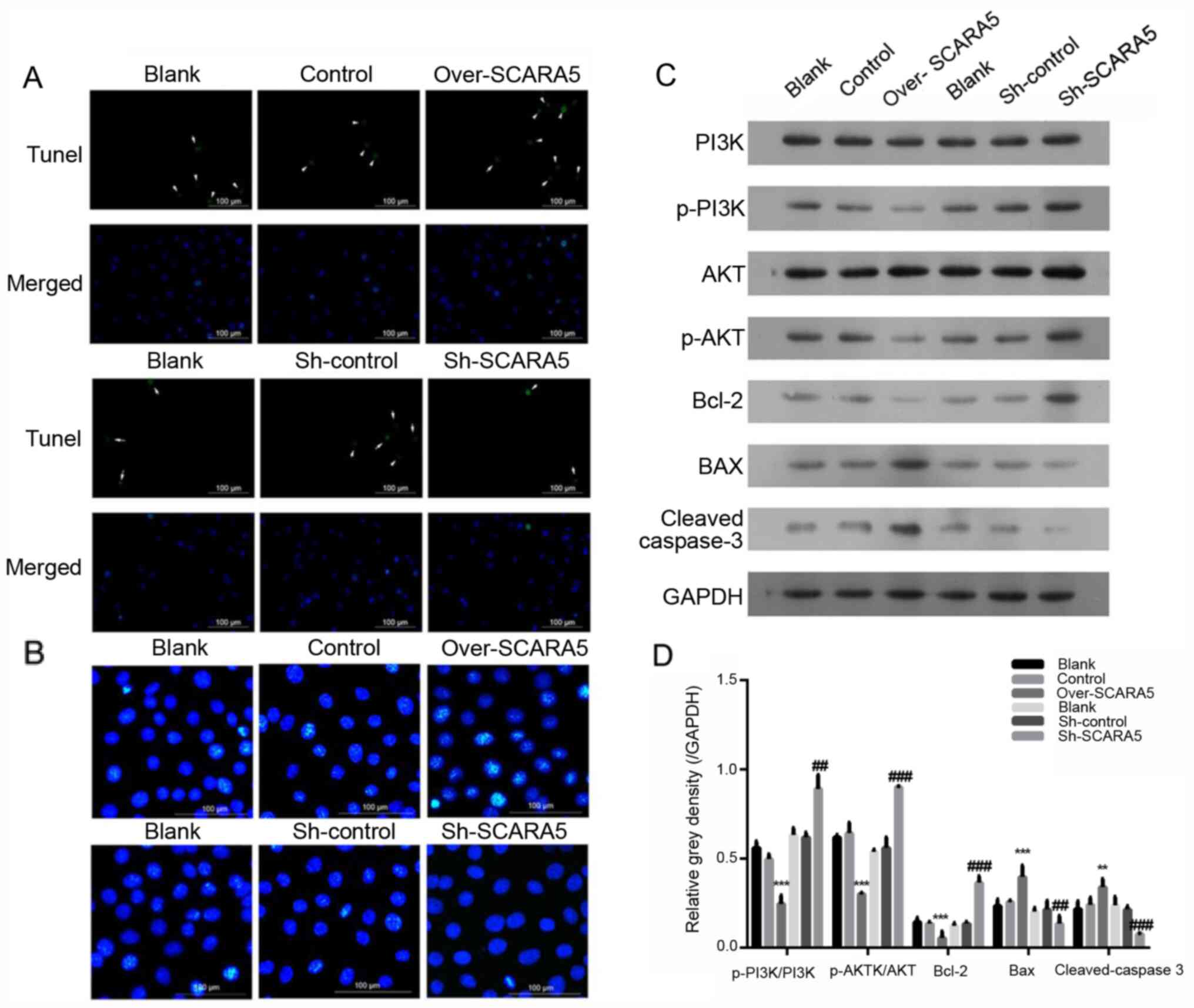 | Figure 3.Effect of SCARA5 overexpression on
Y79 cell apoptosis. (A) TUNEL analysis of apoptosis in Y79 cells.
The arrows indicate the apoptotic cells with green signal; (B)
Hoechst 33342 staining of apoptotic Y79 cells with SCARA5
overexpression or knockdown. The blue signals indicate the
apoptotic cells. Scale bar, 100 µm. (C and D) Western blot analysis
of PI3K, p-PI3K, AKT, p-AKT, Bax, Bcl-2 and cleaved caspase-3
expression in Y79 cells with SCARA5 overexpression or knockdown.
Data are presented as the mean ± SEM (n=3 with three repeats).
**P<0.01, ***P<0.001 vs. Control; ##P<0.01,
###P<0.001 vs. Sh-control. SCARA5, scavenger receptor
class A member 5; sh, short hairpin RNA; p-, phosphorylated. |
Overexpression of SCARA5 affects the
expression of proteins related to the PI3K/AKT pathway and
apoptotic signaling pathway in Y79 cells
The PI3K/AKT signaling pathway serves essential
roles in the apoptosis, proliferation and migration of tumor cells
(22). The expression levels of key
proteins involved in the PI3K/AKT pathway were detected in RB
cells. The results revealed that the expression levels of
p-PI3K/PI3K and p-AKT/AKT were decreased in the SCARA5
overexpression group compared with their expression levels in the
control group (Fig. 3C and D).
Meanwhile, when compared with the control group, the expression
levels of p-PI3K/PI3K and p-AKT/AKT were markedly increased in the
SCARA5 knockdown group (Fig. 3C and
D). Furthermore, the expression levels of key proteins involved
in the regulation of cell apoptosis were detected; the protein
expression levels of Bcl-2 were decreased, whereas the protein
expression levels of Bax and cleaved caspase-3 were increased in
the SCARA5 overexpression group compared with those in the control
group (Fig. 3C and D). Consistent
with these findings, knockdown of SCARA5 increased the expression
levels of Bcl-2, and decreased the expression levels of Bax and
cleaved-caspase 3 (Fig. 3C and D),
which was consistent with the results of the cell apoptosis
analysis.
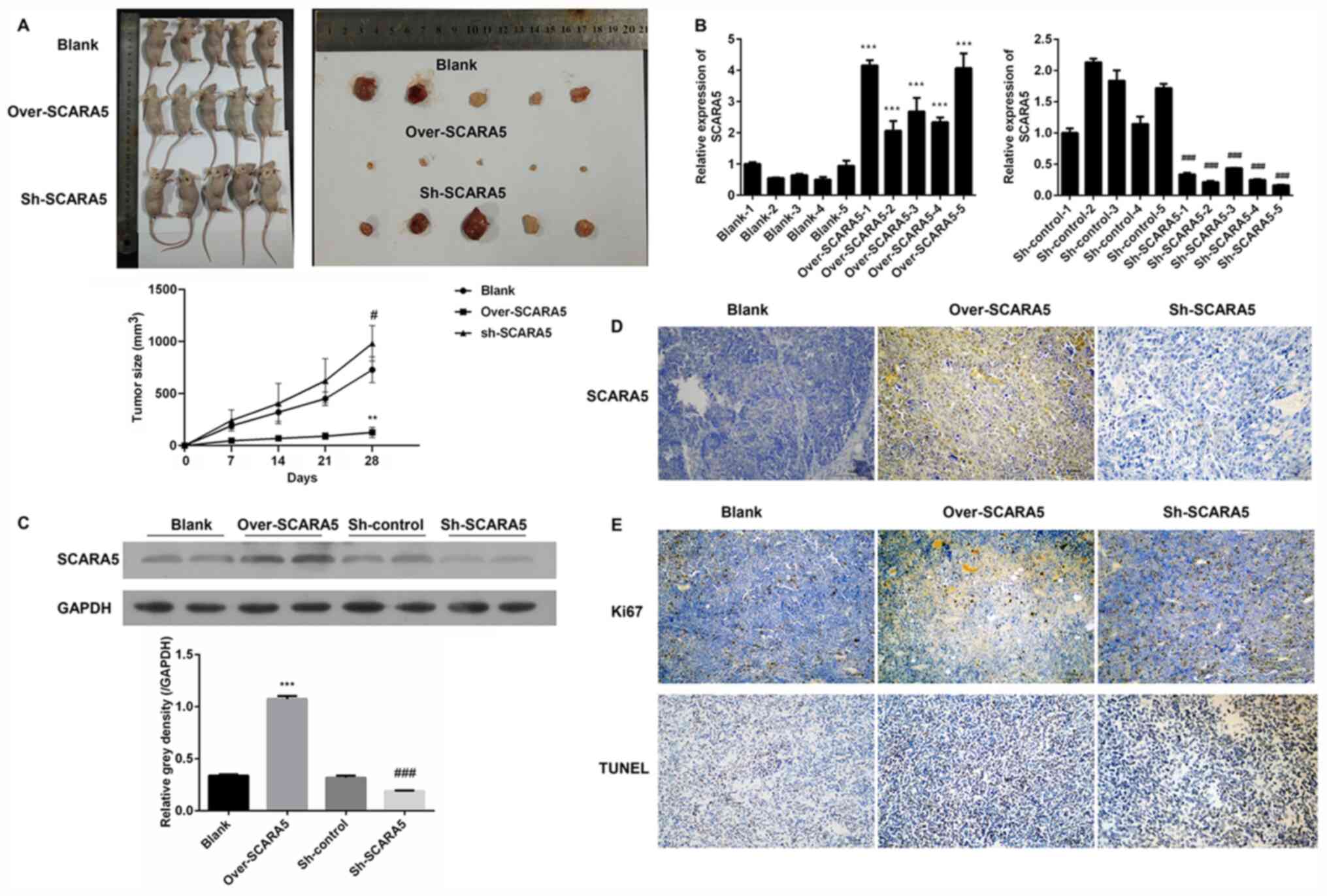
Effect of SCARA5 on RB carcinogenesis
in vivo
A xenograft tumor model was used to examine the role
played by SCARA5 in the development of RB in vivo. Y79 cells
transfected with the SCARA5 overexpression vector or SCARA5
knockdown vector were subcutaneously injected into nude mice. A
subsequent examination revealed that the subcutaneous tumors in the
SCARA5 overexpression group were smaller in size compared with
those in the control group and SCARA5 knockdown group,
demonstrating that overexpression of SCARA5 inhibited tumor growth
(Fig. 4A). Subsequently, the
expression levels of SCARA5 were examined by RT-qPCR, western
blotting and immunohistochemistry. As shown in Fig. 4B, mice injected with
SCARA5-overexpressing cells exhibited an increased expression of
SCARA5, whereas mice injected with the SCARA5 knockdown cells
exhibited decreased levels of SCARA5 expression. The changes in
SCARA5 protein expression levels were consistent with the changes
in mRNA expression, suggesting that the xenograft tumor model with
SCARA5 overexpression or suppression had been successfully
established (Fig. 4C and D). The
results of immunohistochemistry revealed that mice injected with
SCARA5-overexpressing cells had decreased numbers of Ki67-positive
cells and increased numbers of TUNEL-positive cells when compared
with the control and SCARA5 knockdown groups (Fig. 4E). These results were consistent
with the present in vitro results, suggesting that
overexpression of SCARA5 could inhibit proliferation and promote
apoptosis of Y79 Rb cells.
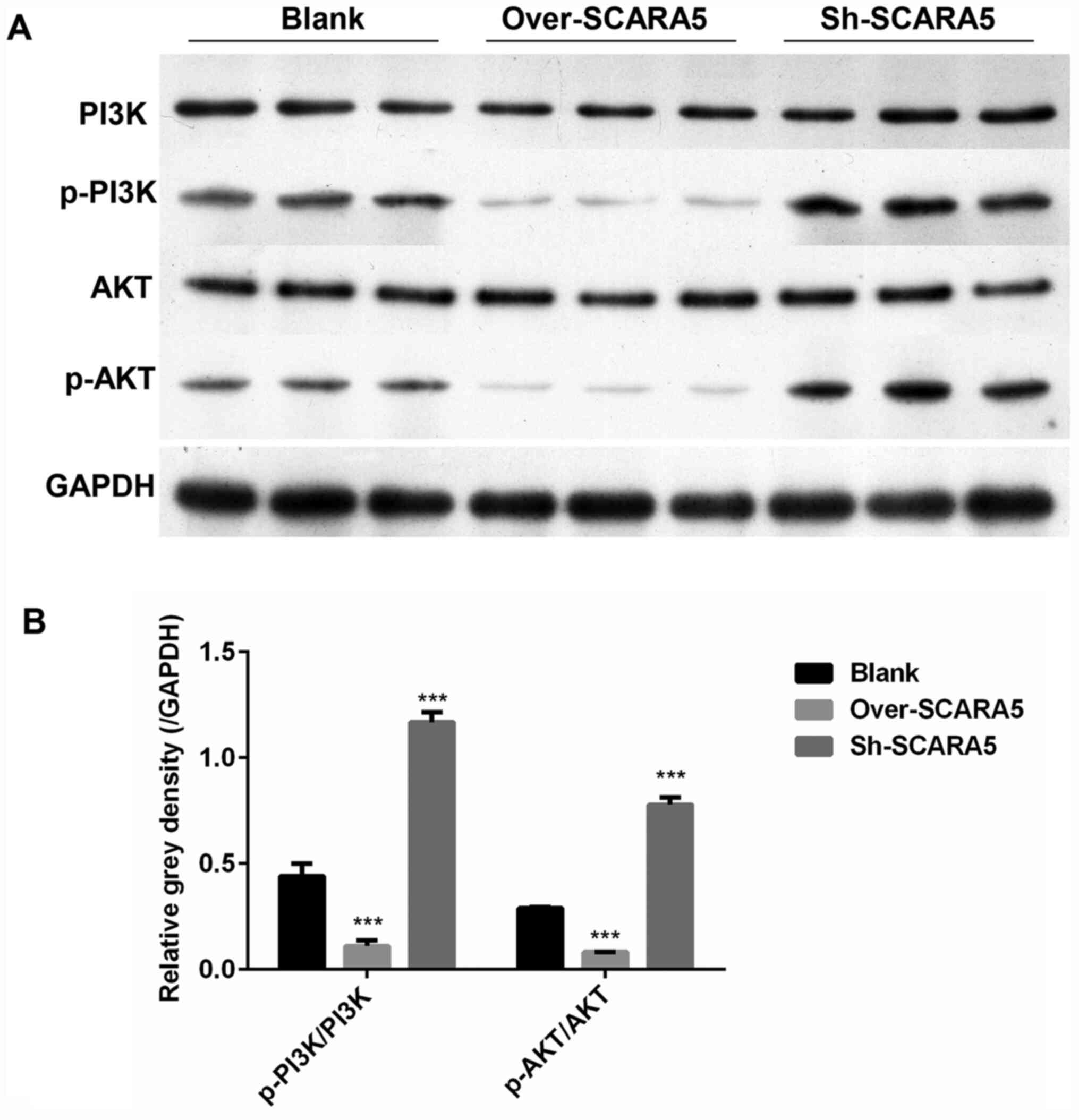
Overexpression of SCARA5 inhibits
p-PI3K and p-AKT expression in vivo
In order to determine how expression of the PI3K/AKT
signaling pathway was affected by SCARA5 in vivo, the in
vivo expression levels of several key proteins involved in the
PI3K/AKT pathway were detected. As shown in Fig. 5A and B, the protein expression
levels of p-PI3K/PI3K and p-AKT/AKT were decreased in the SCARA5
overexpression group compared with their levels in the control
group. Conversely, the protein expression levels of p-PI3K/PI3K and
p-AKT/AKT were markedly increased in the SCARA5 knockdown group.
These results were consistent with the present in vitro
results.
Discussion
RB is one of the most malignant tumors derived from
embryonic retinal cells. RB is almost exclusively diagnosed in
children and a number of children who survive the disease lose
their vision (23). Approximately
70–80% of children with RB are diagnosed at an age of 3 years or
younger, and RB is very uncommon in children >6 years. Notably,
the prevalence of RB is much higher in developing countries
compared with in developed countries (24). In addition, it has been reported
that the presence of human papilloma virus and poor socioeconomic
conditions are suspected risk factors for RB. Current treatment
methods include conservative treatment or removal of the eyeball
(25); however, the prognosis of
patients with RB remains unsatisfactory. Therefore, novel targets
for treating RB need to be identified.
SRs are an important pattern recognition receptor
family; this family includes a large subfamily of members
classified by their multidomain protein structure (26). SRs were initially thought to
contribute to various lipid metabolism disorders by recognizing
acetylated low-density lipoproteins (27). Numerous studies have focused on SR
class A members, due to the critical roles they play in innate
immunity (27–29). SCARA5 is a class A SR. A previous
study suggested that SCARA5 may participate in iron delivery
(30). Moreover, SCARA5 was
revealed to be highly expressed in white adipose tissue and was
suggested to serve an important role in the differentiation of
pluripotent stem cells into adipocytes (31). Numerous studies have revealed that
SCARA5 expression is markedly decreased in numerous types of cancer
cells; moreover, increased SCARA5 expression has been reported to
inhibit the growth of several types of cancer cells (12–14,16).
Consistent with these reports, the present study demonstrated that
overexpression of SCARA5 could inhibit the proliferation and
promote the apoptosis of RB cells. The invasive and migratory
capabilities of tumor cells are essential indicators of tumor cell
aggressiveness and metastatic ability (32). The majority of patients with RB
already show signs of distant metastasis and tissue invasion at the
time of initial diagnosis, and tumor metastasis is a major cause of
death among pediatric patients with RB (33). Therefore, a reduction in the
mortality rate of patients with RB might be achieved by reducing
tumor cell migration and/or invasion (34). The present study demonstrated that
overexpression of SCARA5 inhibited the migratory abilities of Y79
cells, indicating the crucial role of SCARA5 in RB metastasis.
It is well known that the PI3K/AKT signaling pathway
regulates a series of cellular processes, including proliferation,
invasion and metastasis (35). As
the initiator of the PI3K/AKT signaling pathway, P13K serves
important roles in cancer onset and development by regulating the
proliferation and survival of tumor cells (36). Aberrant P13K activity has been
reported to be associated with several processes that contribute to
tumor development, such as the induction of malignant
transformation, the migration and adhesion of tumor cells, and
degradation of the extracellular matrix (37). It has been demonstrated that PI3K
can regulate the activity of multifunctional proteins associated
with cell proliferation and apoptosis, which are critical
activities associated with cancer progression (38). In addition, AKT can activate or
inhibit numerous downstream signaling molecules, including Bcl-2
and Bax, by directly targeting the PI3K protein (39). It has been suggested that
suppression of PI3K/AKT signaling may aid in inhibiting the
proliferation of RB cells (40).
Thus, the PI3K/AKT pathway has been considered an attractive target
for treating RB. Previously, the PI3K inhibitor NVP-BEZ235 was
shown to effectively induce apoptosis of RB cells (41); furthermore, gingerol exhibited
potent anticancer effects in RB355 human RB cells by modulating
PI3K/AKT signaling (42). The
results of the present study revealed that the protein expression
levels of p-PI3K and p-AKT were decreased in RB cells that
overexpressed SCARA5, but were increased in cells with SCARA5
knockdown. Moreover, these changes were accompanied by altered
expression levels of the key apoptosis-associated proteins Bcl-2
and Bax. Thus, the present results indicated that the PI3K/AKT
signaling pathway may be involved in regulating the effects of
SCARA5 on the induction of apoptosis in RB cells. However, the
direct effects of the PI3K/AKT signaling pathway on the
proliferation and migration of human RB cells were not assessed.
Therefore, PI3K/AKT or inhibition of their phosphorylation in cell
or animal models should be studied to confirm the role of the
PI3K/AKT signaling pathway in the cell proliferation and migration
of human RB cells. In addition, further research is required to
investigate whether other crucial proteins associated with cell
proliferation and migration are affected by SCARA5 in RB cells.
In conclusion, the present study revealed that
SCARA5 expression was markedly downregulated in RB cells when
compared with its expression in healthy retinal pigment epithelial
cells. By performing cell transfection, it was demonstrated that
overexpression of SCARA5 could markedly inhibit the proliferation
and migration of Y79 cells, and promote apoptosis of Y79 cells.
Conversely, knockdown of SCARA5 promoted proliferation and
migration, but inhibited apoptosis of Y79 cells. Moreover, in
vivo studies conducted using a xenograft tumor model further
confirmed the in vitro findings and suggested that SCARA5
may function as a tumor suppressor gene in preventing the
occurrence and development of RB.
Acknowledgements
Not applicable.
Funding
No funding was received.
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
JT conceived and designed the study, and provided
administrative support. JW, SW and LC performed the experiments. JW
and SW analyzed and interpretated the data. JT and JW wrote the
manuscript. All authors read and approved the final manuscript.
Ethics approval and consent to
participate
All study protocols involving animals were approved
by the Ethics Committee of Central South University.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Gao Y and Lu X: Decreased expression of
MEG3 contributes to retinoblastoma progression and affects
retinoblastoma cell growth by regulating the activity of
Wnt/β-catenin pathway. Tumour Biol. 37:1461–1469. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Chantada G and Schaiquevich P: Management
of retinoblastoma in children: Current status. Paediatr Drugs.
17:185–198. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Zhou D, Liu P, Sun DW, Chen ZJ, Hu J, Peng
SM and Liu YL: USP22 down-regulation facilitates human
retinoblastoma cell aging and apoptosis via inhibiting TERT/P53
pathway. Eur Rev Med Pharmacol Sci. 21:2785–2792. 2017.PubMed/NCBI
|
|
4
|
Yang Q, Tripathy A, Yu W, Eberhart CG and
Asnaghi L: Hypoxia inhibits growth, proliferation, and increases
response to chemotherapy in retinoblastoma cells. Exp Eye Res.
162:48–61. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Shields CL and Shields JA: Retinoblastoma
management: Advances in enucleation, intravenous chemoreduction,
and intra-arterial chemotherapy. Curr Opin Ophthalmol. 21:203–212.
2010. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Shields CL and Shields JA: Basic
understanding of current classification and management of
retinoblastoma. Curr Opin Ophthalmol. 17:228–234. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Jabbour P, Chalouhi N, Tjoumakaris S,
Gonzalez LF, Dumont AS, Chitale R, Rosenwasser R, Bianciotto CG and
Shields C: Pearls and pitfalls of intraarterial chemotherapy for
retinoblastoma. J Neurosurg Pediatr. 10:175–181. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Zhang H, Zhong J, Bian Z, Fang X, Peng Y
and Hu Y: Long non-coding RNA CCAT1 promotes human retinoblastoma
SO-RB50 and Y79 cells through negative regulation of miR-218-5p.
Biomed Pharmacother. 87:683–691. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Lin P and O'Brien JM: Frontiers in the
management of retinoblastoma. Am J Ophthalmol. 148:192–198. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Peiser L and Gordon S: The function of
scavenger receptors expressed by macrophages and their role in the
regulation of inflammation. Microbes Infect. 3:149–159. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Jiang Y, Oliver P, Davies KE and Platt N:
Identification and characterization of murine SCARA5, a novel class
A scavenger receptor that is expressed by populations of epithelial
cells. J Biol Chem. 281:11834–11845. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Liu J, Hu G, Chen D, Gong AY, Soori GS,
Dobleman TJ and Chen XM: Suppression of SCARA5 by Snail1 is
essential for EMT-associated cell migration of A549 cells.
Oncogenesis. 2:e732013. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Huang J, Zheng DL, Qin FS, Cheng N, Chen
H, Wan BB, Wang YP, Xiao HS and Han ZG: Genetic and epigenetic
silencing of SCARA5 may contribute to human hepatocellular
carcinoma by activating FAK signaling. J Clin Invest. 120:223–241.
2010. View
Article : Google Scholar : PubMed/NCBI
|
|
14
|
Xu Z, Hong Z, Ma M, Liu X, Chen L, Zheng
C, Xi X and Shao J: Rock2 promotes RCC proliferation by decreasing
SCARA5 expression through β-catenin/TCF4 signaling. Biochem Biophys
Res Commun. 480:586–593. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Yan N, Zhang S, Yang Y, Cheng L, Li C, Dai
L, Dai L, Zhang X, Fan P, Tian H, et al: Therapeutic upregulation
of Class A scavenger receptor member 5 inhibits tumor growth and
metastasis. Cancer Sci. 103:1631–1639. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Liu H, Hu J, Wei R, Zhou L, Pan H, Zhu H,
Huang M, Luo J and Xu W: SPAG5 promotes hepatocellular carcinoma
progression by downregulating SCARA5 through modifying β-catenin
degradation. J Exp Clin Cancer Res. 37:2292018. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(−Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Onishi T, Matsuda S, Nakamura Y, Kuramoto
J, Tsuruma A, Sakamoto S, Suzuki S, Fuchimoto D, Onishi A, Chikaki
S, et al: Magnetically promoted rapid immunofluorescence staining
for frozen tissue sections. J Histochem Cytochem. 67:575–587. 2019.
View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Thayanithy V, O'Hare P, Wong P, Zhao X,
Steer CJ, Subramanian S and Lou E: A transwell assay that excludes
exosomes for assessment of tunneling nanotube-mediated
intercellular communication. Cell Commun Signal. 15:462017.
View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Hawkins P, Playle L, Golledge H, Leach M,
Banzett R, Coenen A, Cooper J, Danneman P, Flecknell R, Kirkden R,
et al: Newcastle consensus meeting on carbon dioxide euthanasia of
laboratory animals. 1–17. 2006.
|
|
21
|
National Research Council (US) Committee
for the Update of the Guide for the Care and Use of Laboratory
Animals, . Guide for the Care and Use of Laboratory Animals. 8th
ed. National Academies Press (US); Washington (DC): 2011
|
|
22
|
Martini M, De Santis MC, Braccini L,
Gulluni F and Hirsch E: PI3K/AKT signaling pathway and cancer: An
updated review. Ann Med. 46:372–383. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Ancona-Lezama D, Dalvin LA and Shields CL:
Modern treatment of retinoblastoma: A 2020 review. Indian J
Ophthalmol. 68:2356–2365. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Rossukon K and Duangnate R:
Retinoblastoma: Etiology, modeling, and treatment. Cancers (Basel).
12:23042020. View Article : Google Scholar
|
|
25
|
Sweid A, El Naamani K, Sajja KC, Hammoud
B, Knapp MD, Moylan DD, Joffe D, Morse CE, Habbal D, Weinberg JH,
et al: Incidence and predictors of ophthalmic artery occlusion in
intra-arterial chemotherapy for retinoblastoma. J Neurointerv Surg.
doi: http://dx.doi.org/10.1136/neurintsurg-2020-016759.
|
|
26
|
Chao H and Walsh CE: RNA repair for
haemophilia A. Expert Rev Mol Med. 8:1–8. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Pluddemann A, Neyen C and Gordon S:
Macrophage scavenger receptors and host-derived ligands. Methods.
43:207–217. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Peiser L, Mukhopadhyay S and Gordon S:
Scavenger receptors in innate immunity. Curr Opin Immunol.
14:123–128. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Areschoug T and Gordon S: Scavenger
receptors: Role in innate immunity and microbial pathogenesis. Cell
Microbiol. 11:1160–1169. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Li JY, Paragas N, Ned RM, Qiu A, Viltard
M, Leete T, Drexler IR, Chen X, Sanna-Cherchi S, Mohammed F, et al:
Scara5 is a ferritin receptor mediating non-transferrin iron
delivery. Dev Cell. 16:35–46. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Lee H, Lee YJ, Choi H, Seok JW, Yoon BK,
Kim D, Han JY, Lee Y, Kim HJ and Kim JW: SCARA5 plays a critical
role in the commitment of mesenchymal stem cells to adipogenesis.
Sci Rep. 7:148332017. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
De la Fuente IM and Lopez JI: Cell
motility and cancer. Cancers (Basel). 12:21772020. View Article : Google Scholar
|
|
33
|
Kashyap S, Singh L, Kumar N, Singh MK,
Pushker N, Bakhshi S, Sen S, Lomi N, Meel R and Chawla B: Combined
association of massive choroidal and optic nerve invasion as a
prognostic relevance in primary retinoblastoma: A 10-year study.
Asia Pac J Clin Oncol. 2020.(Epub ahead of print). View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Bock AJ, Nymoen DA, Brenne K, Kaern J and
Davidson B: SCARA3 mRNA is overexpressed in ovarian carcinoma
compared with breast carcinoma effusions. Hum Pathol. 43:669–674.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Cao Y, Xia F, Wang P and Gao M:
MicroRNA935p promotes the progression of human retinoblastoma by
regulating the PTEN/PI3K/AKT signaling pathway. Mol Med Rep.
18:5807–5814. 2018.PubMed/NCBI
|
|
36
|
Fransecky L, Mochmann LH and Baldus CD:
Outlook on PI3K/AKT/mTOR inhibition in acute leukemia. Mol Cell
Ther. 3:22015. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Faes S and Dormond O: PI3K and AKT:
Unfaithful partners in cancer. Int J Mol Sci. 16:21138–21152. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Follo MY, Manzoli L, Poli A, McCubrey JA
and Cocco L: PLC and PI3K/Akt/mTOR signalling in disease and
cancer. Adv Biol Regul. 57:10–16. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Ferenc P, Solar P, Kleban J, Mikes J and
Fedorocko P: Down-regulation of Bcl-2 and Akt induced by
combination of photoactivated hypericin and genistein in human
breast cancer cells. J Photochem Photobiol B. 98:25–34. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Wu S, Ai N, Liu Q and Zhang J: MicroRNA448
inhibits the progression of retinoblastoma by directly targeting
ROCK1 and regulating PI3K/AKT signalling pathway. Oncol Rep.
39:2402–2412. 2018.PubMed/NCBI
|
|
41
|
Xie C, Freeman MJ, Lu H, Wang X, Forster
CL, Sarver AL and Hallstrom TC: Retinoblastoma cells activate the
AKT pathway and are vulnerable to the PI3K/mTOR inhibitor
NVP-BEZ235. Oncotarget. 8:38084–38098. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Meng B, Ii H, Qu W and Yuan H: Anticancer
effects of gingerol in retinoblastoma cancer cells (RB355 Cell
Line) are mediated via apoptosis induction, cell cycle arrest and
upregulation of PI3K/Akt signaling pathway. Med Sci Monit.
24:1980–1987. 2018. View Article : Google Scholar : PubMed/NCBI
|