Introduction
Glaucoma is a chronic and progressive process
resulting in an optic neuropathy with characteristic visual field
deficits (1). Moreover, it affects
>2 million individuals annually in the USA and is expected to
drastically increase in the coming decades due to the aging
population (2). If glaucoma is not
treated in a timely manner, it can lead to blindness (3). Currently, filtering surgery is the
major treatment for glaucoma, but the surgery can easily fail
(4). For example, filtering bleb
scarring following glaucoma filtering surgery is the primary cause
of surgery failure (5,6). Dense collagen fibrillary connective
tissues abnormally thickened below the conjunctival epithelium of
filtering blebs are often observed in cases of failed filtering
surgery, accompanied by increased proliferation of fibroblasts,
thus causing the loss of the aqueous drainage function (7,8). At
present, anti-metabolic drugs such as fluorouracil and mitomycin
can inhibit the scar formation after surgery, but their
anti-metabolic effects may also lead to a series of postoperative
complications, such as ocular hypotension and filtering leakage
(9,10). Therefore, the development of safer
strategies for treating hypertrophic scars (HSs) after glaucoma
surgery is of great importance.
Long non-coding RNAs (lncRNAs) are a class of
non-coding RNA transcripts with a length of ~200 nucleotides
(11,12). lncRNAs are key mediators involved in
the progression of multiple diseases (13,14).
For instance, lncRNA XIST can promote the progression of
osteoarthritis (13) and lncRNA
TUG1 can represses apoptosis, autophagy and inflammatory response
by regulating miR-27a-3p/slit guidance ligand 2 in
lipopolysaccharide-treated vascular endothelial cells (14). Furthermore, previous studies have
revealed a close association between lncRNAs and HSs (15,16).
For example, overexpression of the lncRNA AC067945.2 could inhibit
the progression of HSs by regulating the Wnt and vascular
endothelial growth factor (VEGF) pathways (17). In addition, it has been reported
that the lncRNA family with sequence similarity 225 member B
(FAM225B) is upregulated in HSs (18). Thus, FAM225B may act as a mediator
in formation of HSs. Since the role of FAM225B in HSs remains
unknown, the present study aimed to investigate the function of
FAM225B in HSs.
Autophagy is a key cellular process, in which
cytoplasmic organelles contribute to lysosomal degradation
(19). In addition, activation of
autophagy may result in the dysfunction of cellular organelles
(20). Autophagy also affects cell
survival by regulating the apoptotic cascade. This process is
necessary for maintaining the cellular functions, which in turn
protect the organisms against inflammatory injuries (21,22).
Recent reports have revealed that autophagy can inhibit the
formation of HSs (23–25). These findings indicated that
autophagy serves a crucial role during the formation of HS;
however, the relationship between FAM225B and cell autophagy is yet
to be elucidated. Based on these previous results, the present
study aimed to investigate the association between FAM225B and cell
autophagy in HSs.
The current study aimed to investigate the effect of
FAM225B on the proliferation of scar fibroblasts. The findings of
the present study could provide a novel approach for treating HS
following glaucoma surgery.
Materials and methods
Isolation of scar fibroblasts
Scar fibroblasts were isolated from a patient (aged
63 years, female) who underwent glaucoma surgery in The Affiliated
Hospital of Beihua University in August 2019, as previously
described (26). The patient with
glaucoma was diagnosed as previously described (27). The clinical and pathological data of
patient were collected following their written informed consents.
The present study was approved by the Ethics Committee of The
Affiliated Hospital of Beihua University.
Cell culture
The scar fibroblasts were cultured in 90% RPMI-1640
medium supplemented with 10% FBS, 1% penicillin and 1% streptomycin
(all from Thermo Fisher Scientific, Inc.) in an incubator with 5%
CO2 at 37°C.
Tissue collection
HS tissues (n=5) and normal tissues (n=5) were
obtained from The Affiliated Hospital of Beihua University between
August 2019 and April 2020. HS and normal tissues were obtained
from patients who underwent glaucoma surgery. Patients with
glaucoma were diagnosed as previously described (27). Moreover, the patients who donated
the HS tissues and the normal tissues were the same. The
distributions of age and sex among the patients are listed in
Table I. The present study was
approved by the Institutional Ethical Committee of The Affiliated
Hospital of Beihua University. Written informed consent was
obtained from all participants.
 | Table I.Age and sex distribution among the
patients. |
Table I.
Age and sex distribution among the
patients.
| Patient | Age, years | Sex |
|---|
| 1 | 51 | Female |
| 2 | 27 | Female |
| 3 | 70 | Male |
| 4 | 51 | Female |
| 5 | 64 | Female |
The patient exclusion criteria were as follows: i)
Individuals who suffered from other diseases and are currently
under treatment; ii) pregnant and lactating women; iii) patients
allergic to probiotics or have used/are using antibiotics recently;
and iv) alcoholics (individuals who drank ≥5 bottles of beer at a
time, or for whom the alcohol content in the blood was at ≥0.08)
and smokers.
The patient inclusion criteria were as follows:
Individuals diagnosed with glaucoma, according to the latest
glaucoma diagnostic criteria formulated by the Ocular Fundus
Disease Cooperative Group of the People's Republic of China
(28), and patients who received
glaucoma surgery.
Reverse transcription-quantitative PCR
(RT-qPCR)
Total RNA was extracted from scar fibroblasts or
tissues using TRIzol® reagent (Takara Bio, Inc.)
according to the manufacturer's protocol. Subsequently, cDNA was
synthesized using PrimeScript RT reagent kit (Takara Bio, Inc.)
according to the manufacturer's instructions. The temperature and
duration of RT were: 37°C for 60 min and 85°C for 5 min. RT-qPCR
was performed using a SYBR® Premix Ex Taq™ II kit
(Takara Bio, Inc.) on a 7900HT system (Applied Biosystems; Thermo
Fisher Scientific, Inc.) in triplicate under the following
conditions: Initial denaturation for 10 min at 95°C; 40 cycles of
95°C for 15 sec and 60°C for 30 sec; and final extension for 1 min
at 60°C. The primer sequences (Shanghai GenePharma Co., Ltd.) used
were as follows: FAM225B forward, 5′-CCCTTGGATGCTTGGAGTGA-3′ and
reverse, 5′-GCGACGGTGCTGAATCTTGT-3′; p62 forward,
5′-GAACAGATGGAGTCGGATAACTG-3′ and reverse,
5′-CTGGGAGAGGGACTCAATCAG-3′; Beclin 1 forward,
5′-GGCACAATCAATAACTTCAGGC-3′ and reverse,
5′-GGCAGCTCCTTAGATTTGTCTG-3′; for microtubule associated protein 1
light chain 3 α (LC3) forward, 5′-ACCGCTGTAAGGAGGTACAGC-3′ and
reverse, 5′-GAAGCCGTCCTCGTCTTTCT-3′; autophagy related (ATG)7
forward, 5′-TTCCTCCTCTTGACATTTGCAG-3′ and reverse,
5′-TATCTTCGTCCTTTGACCTTGG-3′; and β-actin forward,
5′-GTCCACCGCAAATGCTTCTA-3′ and reverse, 5′-TGCTGTCACCTTCACCGTTC-3′.
The relative expression was quantified using the 2−ΔΔCq
method (29). The internal
reference gene β-actin was utilized for normalization.
Cell transfection
The small interfering RNAs (siRNAs) against FAM225B
(FAM225B siRNA1, FAM225B siRNA2 and FAM225B siRNA3; 10 nM) and
negative control siRNA (siRNA-NC) were purchased from Shanghai
GenePharma Co., Ltd., and were transfected into scar fibroblasts
using Lipofectamine® 2000 reagent (Thermo Fisher
Scientific, Inc.) at 37°C for 48 h according to the manufacturer's
instructions. Cells were incubated at 37°C for 6 h and the
transfection efficiency was determined using RT-qPCR. The siRNA
sequences were as follows: NC siRNA, 5′-GTCGCAGTACGCATACCTT-3′;
FAM225B siRNA1, 5′-GCATTGGCCTTGACCACAT-3′; FAM225B siRNA2,
5′-GCGTATGCAGACGTTGCTT-3′; and FAM225B siRNA3,
5′-GCTGCTAAGTGGCAGGTAA-3′.
For FAM225B overexpression, scar fibroblasts were
transfected with pcDNA3.1 vector (NC, 10 nM) or pcDNA3.1-FAM225B
overexpression plasmid (FAM225B-OE, 10 nM) using
Lipofectamine® 3000 (Invitrogen; Thermo Fisher
Scientific, Inc.) at 37°C for 48 h. The pcDNA3.1 vector and
FAM225B-OE plasmid were purchased from Shanghai GenePharma Co.,
Ltd.
Western blot analysis
Total proteins were isolated from scar fibroblasts
using a RIPA buffer (Shanghai GenePharma Co., Ltd.) and were
quantified with a BCA protein assay kit (Beyotime Institute of
Biotechnology). Total proteins (30 µg) were separated by 10%
SDS-PAGE, and then transferred onto PVDF (Bio-Rad Laboratories,
Inc.) membranes. Following blocking with 5% skimmed milk at room
temperature for 1 h, the membranes were incubated with primary
antibodies at 4°C overnight. Subsequently, membranes were incubated
with horseradish peroxidase-labeled goat anti-rabbit secondary
antibody (cat. no. ab7090; 1:5,000; Abcam) at room temperature for
1 h. Enhanced chemiluminescence reagent (Thermo Fisher Scientific,
Inc.) was used to visualize the protein bands. The membranes were
scanned with an Odyssey Imaging system and analyzed using the
Odyssey v2.0 software (Li-Cor Biosciences). The primary antibodies
used in the current study were as follows: Anti-p62 (cat. no.
ab109012; 1:1,000), anti-ATG7 (cat. no. ab52472; 1:1,000),
anti-Beclin 1 (cat. no. ab207612; 1:1,000), anti-Bax (cat. no.
ab32503; 1:1,000), anti-Bcl-2 (cat. no. ab182858; 1:1,000),
anti-cleaved caspase 3 (cat. no. ab49822; 1:1,000) and anti-β-actin
(cat. no. ab8226; 1:1,000; all from Abcam). β-actin served as an
internal control.
Cell Counting Kit 8 (CCK-8) assay
Cell viability was tested by CCK-8 kit according to
the manufacturer's instructions. Briefly, scar fibroblasts were
transfected with NC siRNA (10 nM), FAM225B siRNA1/siRNA2/siRNA3 (10
nM), FAM225B-OE (10 nM) or FAM225B-OE (10 nM) + 3-methyladenine
(3-MA; MedChemExpress) for 0, 24, 48 and 72 h at 37°C. Then, cells
were treated with 10 µl CCK-8 reagent (Beyotime Institute of
Biotechnology) and incubated for an additional 2 h at 37°C. The
absorbance was measured at 450 nm using a microplate reader.
Immunofluorescence
Following transfected with siRNA-NC (10 nM), FAM225B
siRNA2 (10 nM), FAM225B siRNA3 (10 nM), pcDNA3.1-NC (10 nM),
FAM225B-OE (10 nM) or FAM225B-OE (10 nM) + 3-MA at 37°C for 72 h,
scar fibroblasts were fixed with 4% paraformaldehyde at room
temperature for 20 min, washed with PBS and blocked with 2% BSA
(Beyotime Institute of Biotechnology) at room temperature for 30
min. Subsequently, cells were incubated with an EdU Assay/EdU
Staining Proliferation kit (iFluor 647; cat. no. ab222421; Abcam)
or anti-LC3 antibody (cat. no. ab63817; Abcam; 1:1,000) at 4°C
overnight. Subsequently, cells were incubated with a secondary
antibody [cat. no. ab6728; IgG (horseradish peroxidase); Abcam;
1:5,000] at room temperature for 1 h. The nuclei were stained with
DAPI (Beyotime Institute of Biotechnology) at room temperature for
5 min. Finally, cells were observed under a fluorescent microscope
at a magnification of ×200.
Transwell assay
For migration assays, the transfected cells
(density, 3×104 cells/well) were resuspended in
serum-free DMEM (100 µl; Thermo Fisher Scientific, Inc.) and were
then plated onto the upper chamber of the Transwell 24-well plates.
Moreover, the lower chamber of each well was supplemented with 600
µl DMEM containing 10% FBS. Following incubation for 24 h at 37°C,
the non-migrated cells on the upper surface of the membrane were
removed. Subsequently, the migrated cells on the lower surface were
fixed with 4% paraformaldehyde at room temperature for 15 min,
followed by staining with 0.5% crystal violet at room temperature
for 30 min. The number of migrated cells was counted under a light
microscope at a magnification of ×400.
Cell apoptosis assays
The early and late apoptosis (Annexin-V+
PI− plus Annexin-V+ PI+) of scar
fibroblasts were measured via flow cytometry. Scar fibroblasts were
seeded into 6-well plates at a density of 1×106
cells/well. The residue was resuspended in 100 µl binding buffer
following centrifugation (4°C) at 500 × g for 5 min. Subsequently,
5 µl Annexin V-FITC (BD Biosciences) and 5 µl PI (BD Biosciences)
at room temperature were added into the cell suspension for 15 min.
The cell apoptosis rate was measured using a flow cytometer (BD
Biosciences) and the results were analyzed using FACS (BD
Biosciences) with FlowJo (v10.6.2; FlowJo LLC).
Reactive oxygen species (ROS)
detection
ROS detection was performed using a ROS detection
kit (green fluorescence; Beyotime Institute of Biotechnology). Cell
suspensions were collected and stained with the ROS probe
2,7-dichlorofluorescein diacetate (Beyotime Institute of
Biotechnology) at room temperature for 15 min as previously
described (30). After 20 min of
incubation, cells were centrifuged at 300 × g (4°C for 10 min),
washed with PBS and resuspended. Finally, the relative ROS level
was measured via flow cytometry (FACSAria II; BD Biosciences), and
the data were analyzed using FlowJo 10.0 (BD Biosciences).
ELISA
The levels of superoxide dismutase (SOD) and
glutathione (GSH) in the supernatants of scar fibroblasts
(collected by centrifugation at 4°C for 10 min at 300 × g) were
detected using SOD ELISA kit [cat. no. CSB-EL022399; Multisciences
(Lianke) Biotech Co., Ltd.] and GSH ELISA kit [cat. no. xy-E10818;
Multisciences (Lianke) Biotech Co., Ltd.] according to the
manufacturer's instructions.
Statistical analysis
Data are presented as the mean ± SD. The CCK-8 assay
was performed in quintuplicate. Cell transfections, RT-qPCR,
immunofluorescence staining, flow cytometry, western blot analysis
and the Transwell migration assay were repeated three times.
GraphPad Prism software (version 7.0; GraphPad Software, Inc.) was
used to analyze the data. One-way ANOVA and Tukey's tests were
conducted to evaluate the differences for multiple comparisons.
P<0.05 was considered to indicate a statistically significant
difference.
Results
FAM225B knockdown significantly
inhibits the proliferation of scar fibroblasts
The morphology of scar fibroblasts isolated from a
patient was observed under a microscope, and fibrotic scarring was
observed (Fig. 1A). This morphology
was consistent with the characteristics of scar fibroblasts
(31). Moreover, the expression of
FAM225B was significantly upregulated in HS tissues, compared with
normal tissues (Fig. 1B). It was
also found that the expression of FAM225B in scar fibroblasts was
significantly downregulated following transfection with FAM225B
siRNAs (Fig. 1C). In addition,
FAM225B was more efficiently downregulated when scar fibroblasts
were treated with FAM225B siRNA2 or siRNA3 compared with FAM225B
siRNA1. Therefore, FAM225B siRNA2 and siRNA3 were selected to
knockdown FAM225B expression in the subsequent experiments. The
viability of scar fibroblasts was significantly decreased following
transfection with FAM225B siRNA2 or siRNA3 (Fig. 1D). Consistently, knockdown of
FAM225B significantly attenuated the proliferation of scar
fibroblasts (Fig. 1E). Overall,
these findings indicated that FAM225B knockdown significantly
attenuated the proliferation of scar fibroblasts.
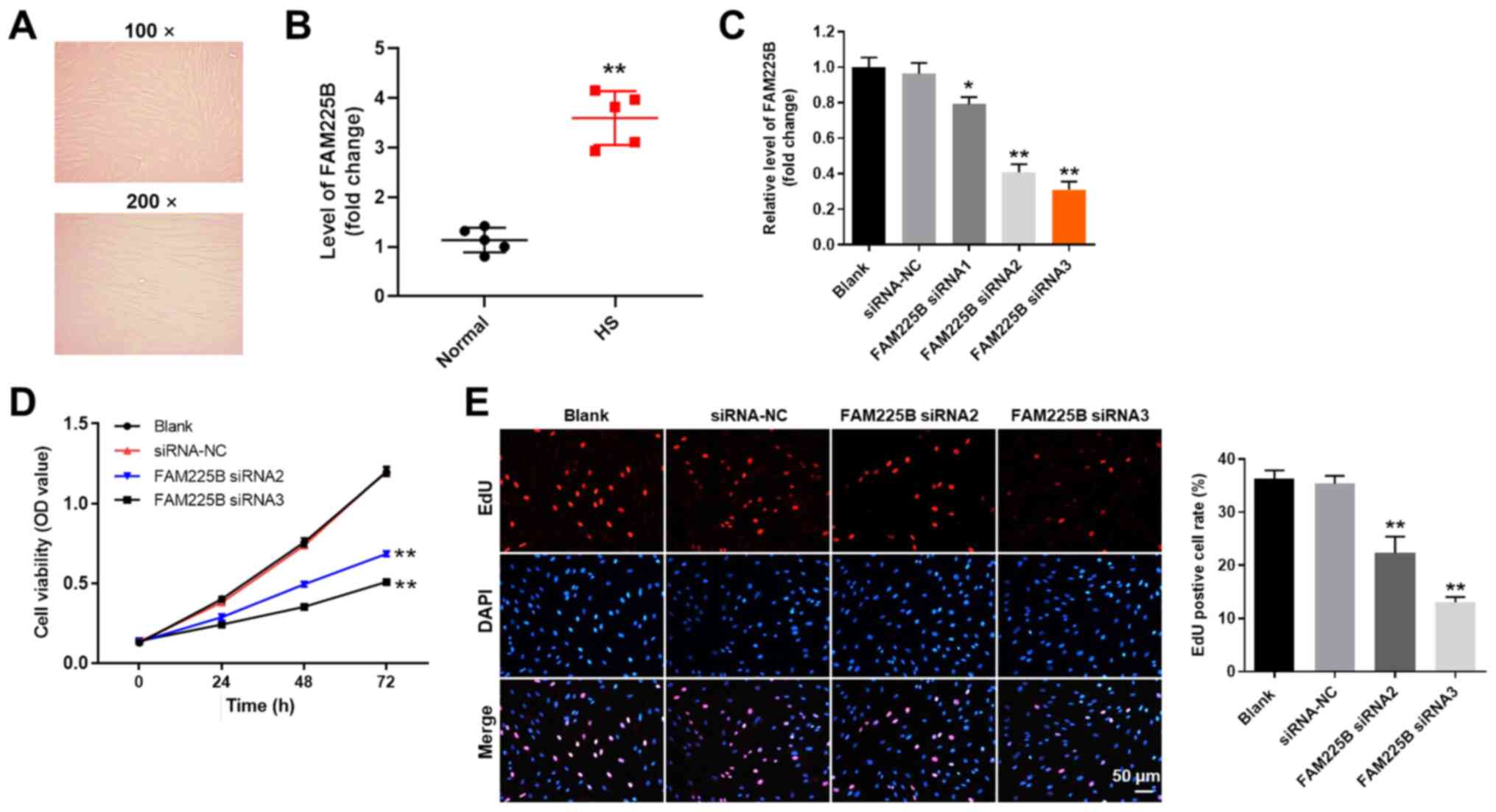 | Figure 1.FAM225B knockdown significantly
inhibits the proliferation of scar fibroblasts. (A) Scar fibroblast
morphology was observed under a microscope. (B) Expression of
FAM225B in HS or normal tissues was detected via RT-qPCR.
**P<0.01 vs. normal. (C) Scar fibroblasts were transfected with
FAM225B siRNA1, siRNA2 or siRNA3 for 24 h. Then, the efficiency of
cell transfection was detected via RT-qPCR. (D) Scar fibroblasts
were transfected with FAM225B siRNA2 or siRNA3 for 0, 24, 48 or 72
h. Then, cell viability was assessed using a Cell Counting Kit-8
assay. (E) Proliferation of scar fibroblasts was detected using EdU
staining (magnification, ×200). Red fluorescence indicates EdU,
while blue fluorescence indicates DAPI. *P<0.05, **P<0.01 vs.
Blank. RT-qPCR, reverse transcription-quantitative PCR; EdU,
5-Ethynyl-2′-deoxyuridine; NC, negative control; siRNA, small
interfering RNA; FAM225B, family with sequence similarity 225
member B; HS, hypertrophic scar; OD, optical density. |
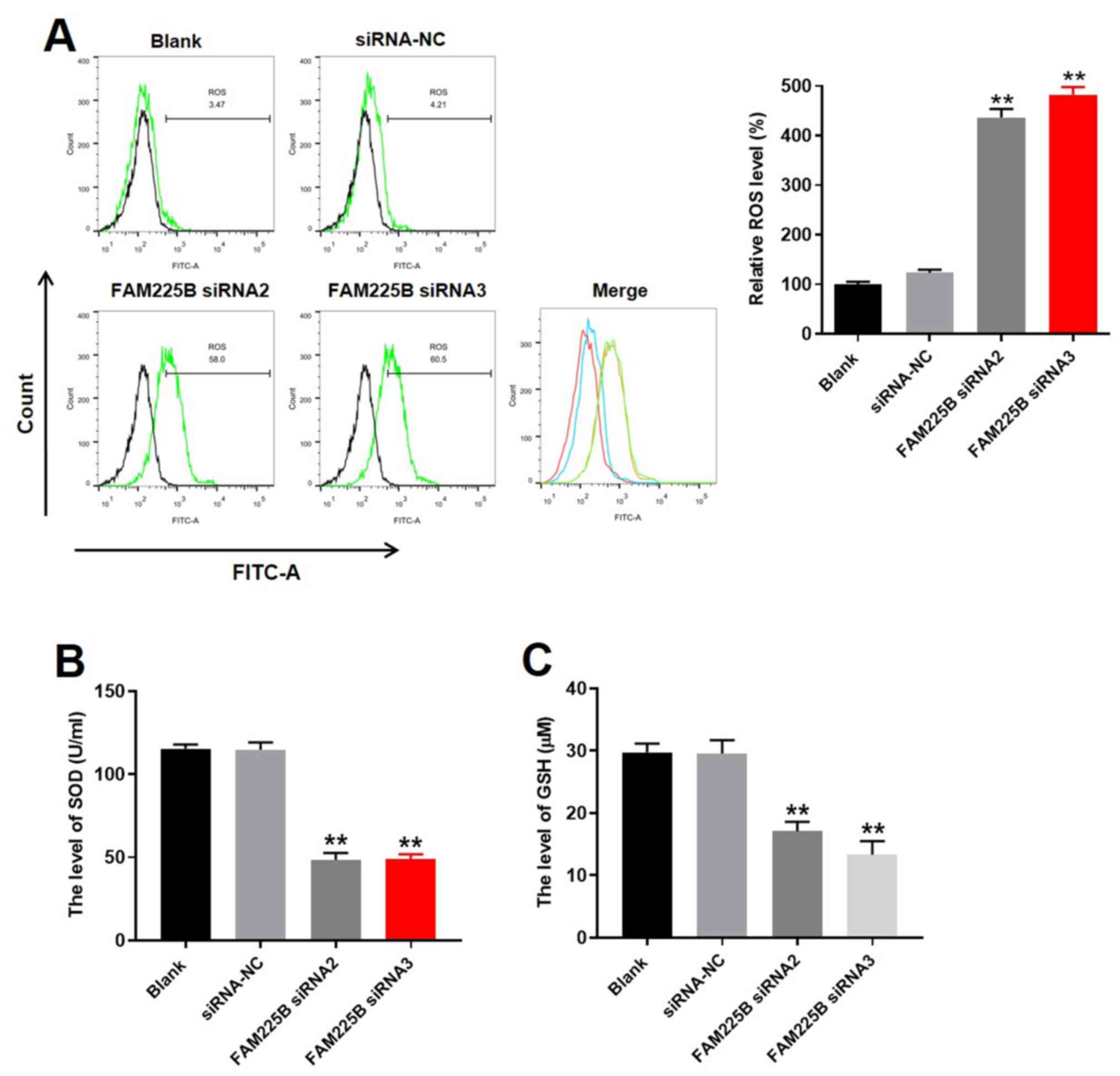
Knockdown of FAM225B notably induces
oxidative stress in scar fibroblasts
To evaluate the effect of FAM225B on the levels of
ROS in scar fibroblasts, flow cytometric analysis was conducted. As
expected, knockdown of FAM225B significantly increased ROS levels
in scar fibroblasts (Fig. 2A). In
addition, the levels of SOD and GSH in the supernatants of scar
fibroblasts were significantly decreased by FAM225B siRNA (Fig. 2B and C). Since the levels of ROS,
SOD and GSH are closely associated with cell injury (32), the aforementioned data suggested
that knockdown of FAM225B could notably induce oxidative stress in
scar fibroblasts.
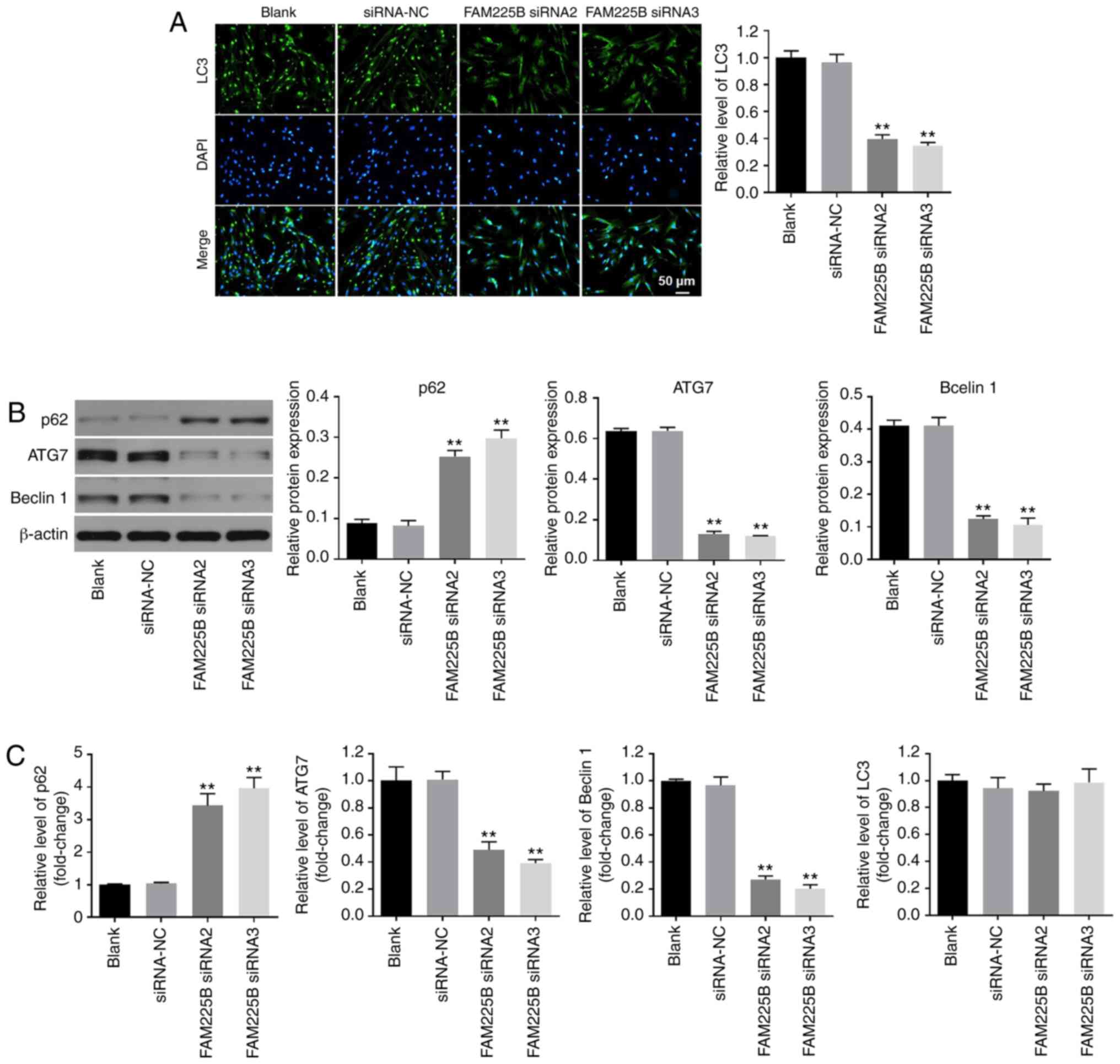
FAM225B siRNA attenuates the
progression of HS in vitro by inhibiting autophagy
In order to evaluate the association between FAM225B
and cell autophagy, immunofluorescence staining against LC3
(autophagy indicator) was performed. As presented in Fig. 3A, the expression of LC3 in scar
fibroblasts was significantly decreased following FAM225B
knockdown. In addition, knockdown of FAM225B significantly
upregulated p62 expression (Fig. 3B and
C). By contrast, the protein expression levels of ATG7 and
Beclin-1 in scar fibroblasts were significantly downregulated in
cells transfected with FAM225B siRNA (Fig. 3B and C). However, knockdown of
FAM225B had a very limited effect on the mRNA expression of LC3
(Fig. 3C). Since LC3, p62, ATG7 and
Beclin-1 are involved in the process of cell autophagy (33–35),
these factors can be considered as autophagy indicators. Thus, the
results indicated that FAM225B knockdown attenuated the progression
of HS in vitro by inhibiting autophagy.
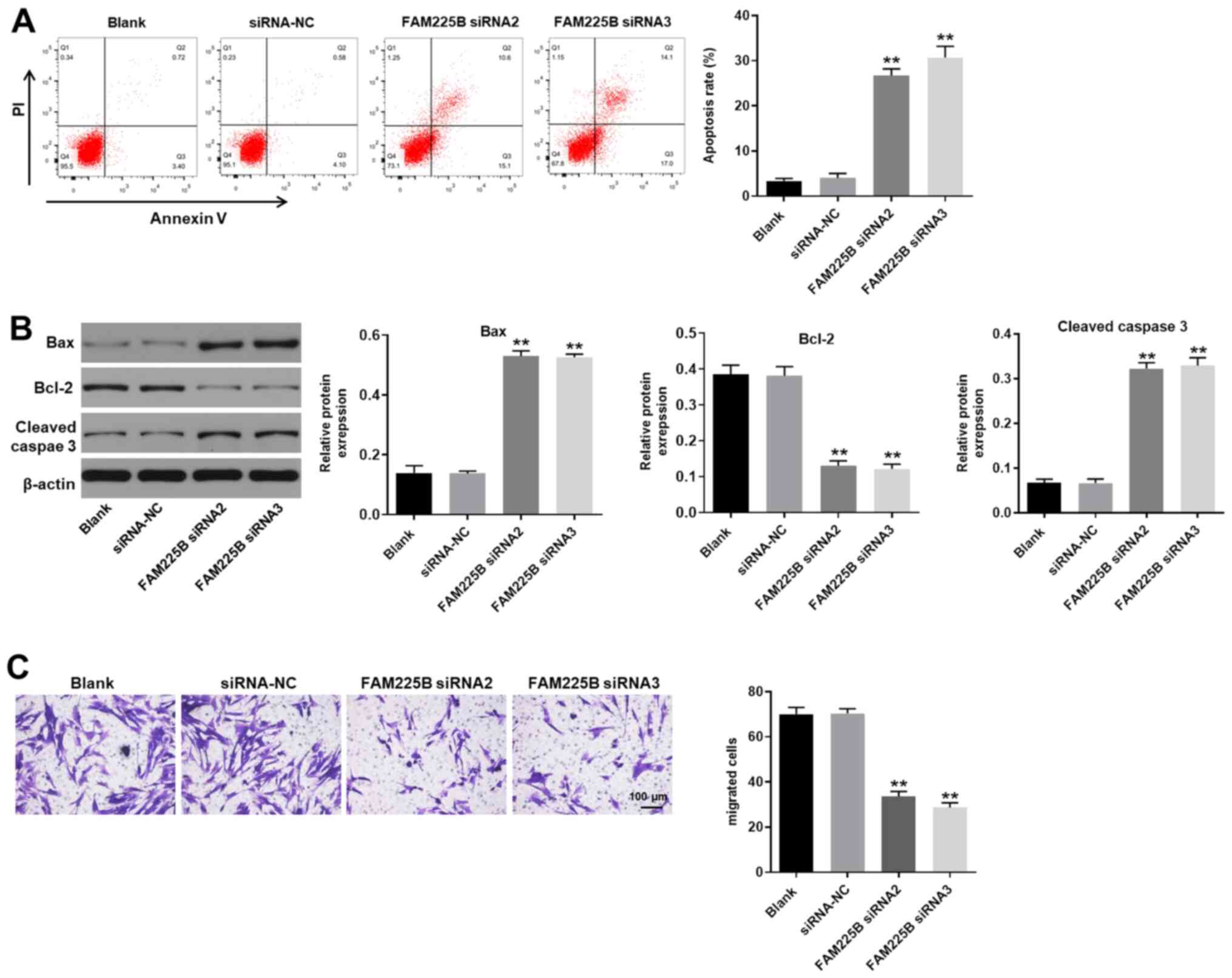
Knockdown of FAM225B significantly
induces apoptosis and inhibits migration of scar fibroblasts
To evaluate cell apoptosis, flow cytometric analysis
(Annexin V and PI staining) was performed. As presented in Fig. 4A, knockdown of FAM225B significantly
induced scar fibroblast apoptosis. Furthermore, FAM225B siRNA
significantly increased the expression levels of Bax and cleaved
caspase 3, and downregulated those of Bcl-2 in scar fibroblasts
(Fig. 4B). FAM225B knockdown also
significantly inhibited the migratory ability of scar fibroblasts
(Fig. 4C). Therefore, knockdown of
FAM225B markedly induced apoptosis and attenuated the migration of
scar fibroblasts.
Overexpression of FAM225B
significantly promotes the proliferation of scar fibroblasts by
inhibiting autophagy
In order to further confirm the association between
FAM225B and cell autophagy in scar fibroblasts, cells were
overexpressed with FAM225B, and then the efficiency of
overexpression was detected. As expected, pcDNA3.1-FAM225B
significantly increased the expression levels of FAM225B in scar
fibroblasts (Fig. 5A). In addition,
the viability of scar fibroblasts was significantly enhanced by
FAM225B overexpression, which was partially reversed following
treatment of fibroblasts with the autophagy inhibitor 3-MA
(Fig. 5B). Consistently, the
FAM225B overexpression-mediated increase of scar fibroblast
proliferation was significantly inhibited by 3-MA (Fig. 5C). Furthermore, overexpression of
FAM225B significantly enhanced the migratory ability of scar
fibroblasts, which was notably reversed by 3-MA (Fig. 5D). Collectively, these results
suggested that overexpression of FAM225B significantly promoted the
proliferation of scar fibroblasts by inhibiting autophagy.
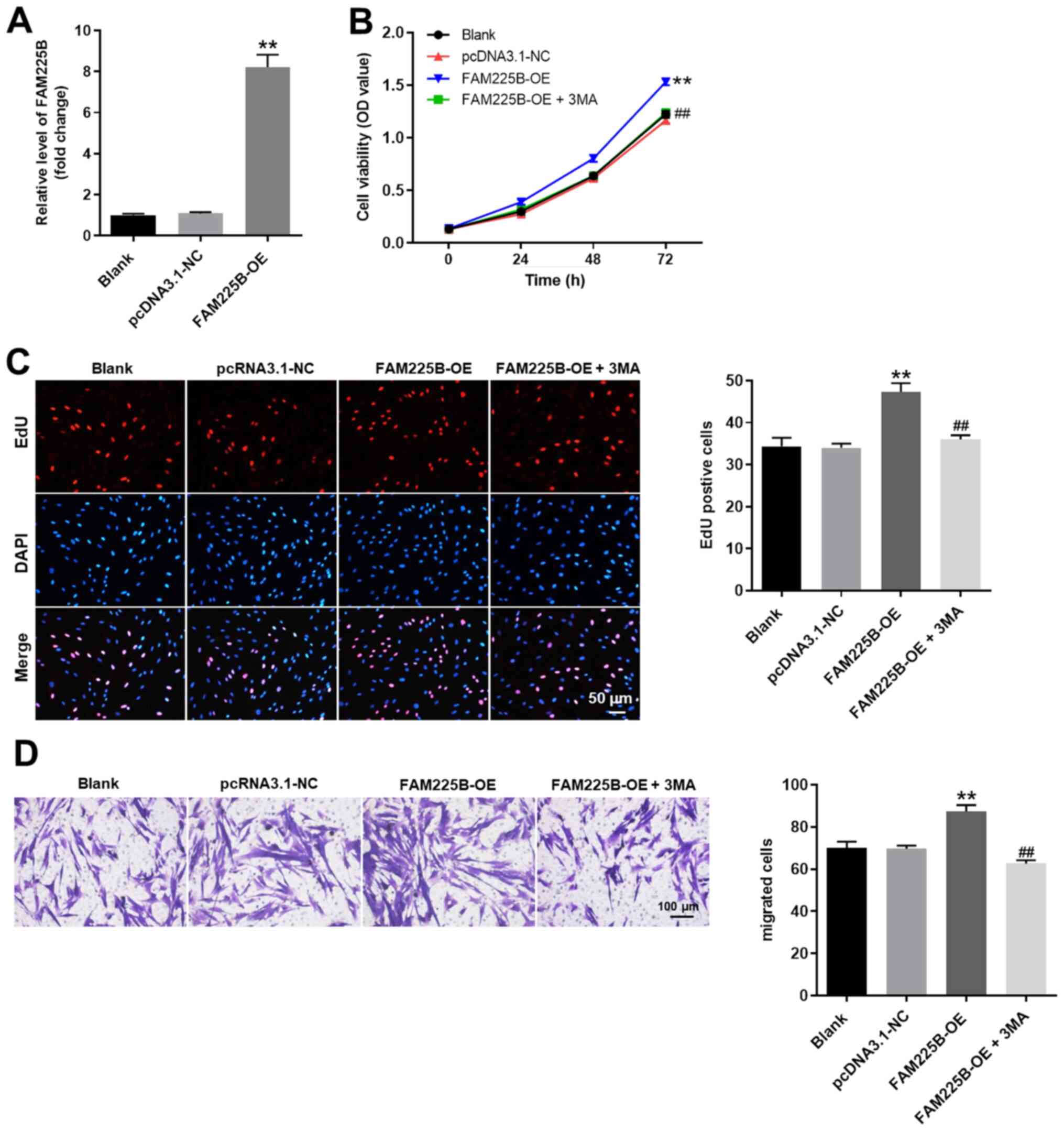 | Figure 5.OE of FAM225B significantly promotes
the proliferation of scar fibroblasts by inhibiting autophagy. (A)
Scar fibroblasts were transfected with pcDNA3.1-FAM225B (FAM225B
OE). Then, the efficiency of overexpression was detected via
reverse transcription-quantitative PCR. (B) Scar fibroblasts were
transfected with FAM225B OE or FAM225B OE + 3-MA. Then, cell
viability was assessed using a Cell Counting Kit-8 assay. (C)
Proliferation of scar fibroblasts was detected using EdU staining
(magnification, ×200). (D) Cell migration was detected with a
Transwell assay (magnification, ×400). **P<0.01 vs. Blank;
##P<0.01 vs. FAM225B OE. NC, negative control;
FAM225B, family with sequence similarity 225 member B; OE,
overexpression; OD, optical density; EdU,
5-Ethynyl-2′-deoxyuridine; 3-MA, 3-methyladenine. |
Discussion
Understanding the pathogenesis of HS may benefit the
selection of the appropriate HS treatment strategy. Several studies
have reported that non-coding RNAs, including lncRNAs, serve a key
role in HS (15,18,36).
For example, Tu et al (18)
revealed that FAM225B was upregulated in HS. The results of the
present study further supplemented the current knowledge regarding
the effect of FAM225B on the formation of HSs, suggesting that
FAM225B could serve an important role in HS progression.
Previous studies have revealed that autophagy serves
key roles in the pathogenesis of HSs as it can promote the
formation of HS fibroblasts (23,37).
For instance, activation of autophagy can inhibit proliferation of
scar fibroblasts by inducing cell apoptosis (38), and autophagy can inhibit the
proliferation of scar fibroblasts via the inactivation of the
PI3K/Akt/mTOR pathway (39). On the
contrary, the present data suggested that inhibition of autophagy
could decrease the proliferation of scar fibroblasts. This
discrepancy may be due to the different degrees of cell injury and
oxidative stress.
Beclin-1 is an important autophagy mediator, which
increases autophagy (40).
Furthermore, it has been reported that ATG7 promotes autophagy
(41), while p62 is negatively
associated with the autophagy process (42). The results of the current study were
consistent with the aforementioned findings, suggesting that
knockdown of FAM225B could inhibit the progression of HS by
inhibiting autophagy.
Apoptosis is closely associated with autophagy in
types of cancer, including hepatocellular carcinoma (43,44).
Importantly, autophagy is necessary for the onset of apoptosis,
which usually triggers the occurrence of cell apoptosis and injury
(45,46). Based on these aforementioned
findings, it can be concluded that knockdown of FAM225B induced
cell injury and increased the oxidative stress of scar fibroblasts
by inhibiting cell autophagy and inducing apoptosis. Moreover, a
previous study reported that Proscillaridin A could induce
apoptosis and autophagy in liver cancer cells (43). Thus, the association between
apoptosis and autophagy requires further investigation.
There are some limitations to the present research.
Firstly, this study focused only on the autophagy process. In
addition, the mechanism underlying the effect of FAM225B knockdown
on promoting cell autophagy remains unknown, and the microRNAs
sponged by FAM225B should be evaluated. Therefore, further studies
are required.
In conclusion, the present study demonstrated that
knockdown of FAM225B significantly attenuating the progression of
HS following glaucoma surgery by inhibiting autophagy. Therefore,
the current finding may provide a new perspective of developing
treatment strategy for the patients with HS after glaucoma
surgery.
Acknowledgements
Not applicable.
Funding
No funding was received.
Availability of data and materials
All data generated or analyzed during this study are
included in this published article.
Authors' contributions
XM and LL conceived and supervised the study,
designed experiments and reviewed the results. Both authors read
and approved the final version of the manuscript.
Ethics approval and consent to
participate
The present study was approved by the Institutional
Ethical Committee of The Affiliated Hospital of Beihua University.
Written informed consent was obtained from all participants.
Patient consent for publication
Not applicable.
Competing interests
These authors declared that they have no competing
interests.
References
|
1
|
Li X, Leng Y, Jiang Q, Wang Z, Luo P,
Zhang C, Chen L, Wang Y, Wang H, Yue X, et al: Eye drops of
metformin prevents fibrosis after glaucoma filtration surgery in
rats via activating AMPK/Nrf2 signaling pathway. Front Pharmacol.
11:10382020. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Swogger J, Conner IP, Rosano M, Kemmerer
M, Happ-Smith C, Wells A, Schuman JS and Yates CC: Injected versus
sponge-applied mitomycin C (MMC) during modified trabeculectomy in
New Zealand white rabbit model. Transl Vis Sci Technol. 9:232020.
View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Yaman D, Takmaz T, Yüksel N, Dincer SA and
Şahin FI: Evaluation of silent information regulator T (SIRT) 1 and
Forkhead Box O (FOXO) transcription factor 1 and 3a genes in
glaucoma. Mol Biol Rep. 47:9337–9344. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Cheng WS, Chen CL, Chen JT, Lin LT, Pao
SI, Chen YH and Lu DW: AR12286 alleviates TGF-β-related
myofibroblast transdifferentiation and reduces fibrosis after
glaucoma filtration surgery. Molecules. 25:44222020. View Article : Google Scholar
|
|
5
|
Yamamoto K, Kokubun T, Sato K, Akaishi T,
Shimazaki A, Nakamura M, Shiga Y, Tsuda S, Omodaka K, Saya H and
Nakazawa T: The DNA topoisomerase II inhibitor amsacrine as a novel
candidate adjuvant in a model of glaucoma filtration surgery. Sci
Rep. 9:192882019. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Yu J, Qiu LX, Qing GP, Zhao BW and Wang H:
Modified cortex mori capsules improving the successful rate of
functional filtering blebs after reclinical glaucoma filtering
surgery. World J Clin Cases. 7:3436–3445. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Zhang F, Liu K, Pan Z, Cao M, Zhou D, Liu
H, Huang Y and Duan X: Effects of rosiglitazone/PHBV drug delivery
system on postoperative fibrosis in rabbit glaucoma filtration
surgery model. Drug Deliv. 26:812–819. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Sun J, Liu X, Lei Y, Tang M, Dai Z, Yang
X, Yu X, Yu L, Sun X and Ding J: Sustained subconjunctival delivery
of cyclosporine A using thermogelling polymers for glaucoma
filtration surgery. J Mater Chem B. 5:6400–6411. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Holló G: Wound Healing and Glaucoma
Surgery: Modulating the scarring process with conventional
antimetabolites and new molecules. Dev Ophthalmol. 59:80–89. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Sharma A, Anumanthan G, Reyes M, Chen H,
Brubaker JW, Siddiqui S, Gupta S, Rieger FG and Mohan RR:
Epigenetic modification prevents excessive wound healing and scar
formation after glaucoma filtration surgery. Invest Ophthalmol Vis
Sci. 57:3381–3389. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Zhang H, Li J, Shao W and Shen N: lncRNA
SNHG9 is downregulated in osteoarthritis and inhibits chondrocyte
apoptosis by downregulating miR-34a through methylation. BMC
Musculoskelet Disord. 21:5112020. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Wang JH, Wu XJ, Duan YZ and Li F: Circular
RNA_CNST promotes the tumorigenesis of osteosarcoma cells by
sponging miR-421. Cell Transplant. 29:9636897209261472020.
View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Xu YT, Leng YR, Liu MM, Dong RF, Bian J,
Yuan LL, Zhang JG, Xia YZ and Kong LY: MicroRNA and long noncoding
RNA involvement in gout and prospects for treatment. Int
Immunopharmacol. 87:1068422020. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Dong Y, Fan G, Li Y and Zhou Q: TUG1
represses apoptosis, autophagy, and inflammatory response by
regulating miR-27a-3p/SLIT2 in LPS-treated vascular endothelial
cells. J Surg Res. 256:345–354. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Li M, Wang J, Liu D and Huang H:
High-throughput sequencing reveals differentially expressed lncRNAs
and circRNAs, and their associated functional network, in human
hypertrophic scars. Mol Med Rep. 18:5669–5682. 2018.PubMed/NCBI
|
|
16
|
Chen J, Zhou R, Liang Y, Fu X, Wang D and
Wang C: Blockade of lncRNA-ASLNCS5088-enriched exosome generation
in M2 macrophages by GW4869 dampens the effect of M2 macrophages on
orchestrating fibroblast activation. FASEB J. 33:12200–12212. 2019.
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Chen L and Li J, Li Q, Li X, Gao Y, Hua X,
Zhou B and Li J: Overexpression of lncRNA AC067945.2 down-regulates
collagen expression in skin fibroblasts and possibly correlates
with the VEGF and Wnt signalling pathways. Cell Physiol Biochem.
45:761–771. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Tu L, Huang Q, Fu S and Liu D: Aberrantly
expressed long noncoding RNAs in hypertrophic scar fibroblasts
in vitro: A microarray study. Int J Mol Med. 41:1917–1930.
2018.PubMed/NCBI
|
|
19
|
Yamoto M, Alganabi M, Chusilp S, Lee D,
Yazaki Y, Lee C, Li B and Pierro A: Lysosomal overloading and
necrotizing enterocolitis. Pediatr Surg Int. 36:1157–1165. 2020.
View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Li Y, Zhang J, Zhou K, Xie L, Xiang G,
Fang M, Han W, Wang X and Xiao J: Elevating sestrin2 attenuates
endoplasmic reticulum stress and improves functional recovery
through autophagy activation after spinal cord injury. Cell Biol
Toxicol. Aug 1–2020.(Epub ahead of print). doi:
10.1007/s10565-020-09550-4. View Article : Google Scholar
|
|
21
|
Gutjahr A, Papagno L, Vernejoul F, Lioux
T, Jospin F, Chanut B, Perouzel E, Rochereau N, Appay V, Verrier B
and Paul S: New chimeric TLR7/NOD2 agonist is a potent adjuvant to
induce mucosal immune responses. EBioMedicine. 58:1029222020.
View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Fu H, Chung KR, Liu X and Li H: Aaprb1, a
subtilsin-like protease, required for autophagy and virulence of
the tangerine pathotype of Alternaria alternata. Microbiol
Res. 240:1265372020. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Shi J, Xiao H, Li J, Zhang J, Li Y, Zhang
J, Wang X, Bai X, Tao K, Hu D and Guan H: Wild-type p53-modulated
autophagy and autophagic fibroblast apoptosis inhibit hypertrophic
scar formation. Lab Invest. 98:1423–1437. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Liu X, Xu Q, Long X, Liu W, Zhao Y,
Hayashi T, Hattori S, Fujisaki H, Ogura T, Tashiro SI, et al:
Silibinin-induced autophagy mediated by PPARα-sirt1-AMPK pathway
participated in the regulation of type I collagen-enhanced
migration in murine 3T3-L1 preadipocytes. Mol Cell Biochem.
450:1–23. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Kyun S, Yoo C, Park HY, Kim J and Lim K:
The effects of exogenous lactate administration on the
IGF1/Akt/mTOR pathway in rat skeletal muscle. Int J Environ Res
Public Health. 17:78052020. View Article : Google Scholar
|
|
26
|
Wang XC, Wang T, Zhang Y, Wang LL, Zhao RY
and Tan W: Tacrolimus inhibits proliferation and induces apoptosis
by decreasing survivin in scar fibroblasts after glaucoma surgery.
Eur Rev Med Pharmacol Sci. 22:2934–2940. 2018.PubMed/NCBI
|
|
27
|
Jammal AA, Berchuck SI, Thompson AC, Costa
VP and Medeiros FA: The effect of age on increasing susceptibility
to retinal nerve fiber layer loss in glaucoma. Invest Ophthalmol
Vis Sci. 61:82020. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Mu Y: Correlation analysis between TCM
syndromes of primary angle-closure glaucoma and type A personality.
Chin J Tradit Chin Ophthalmol. 10:23–30. 2020.
|
|
29
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) Method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Ye Q, Wu J, Wu P, Wang J, Niu W, Yang S,
Chen M, Rehman S and Zhu N: Enhancing peroxymonosulfate activation
of Fe-Al layered double hydroxide by dissolved organic matter:
Performance and mechanism. Water Res. 185:1162462020. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Wei Z, Wang Y, Jiang L, Ji N, Wang Y, Chen
F, Li T, Li J, Xu H, Zeng X and Chen Q: miR-223 regulates oral
squamous cell carcinoma metastasis through the Wnt/β-catenin
signaling pathway. Oral Oncol. 109:1049412020. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Liu YP, Wan J, Long F, Tian J and Zhang C:
circPVT1 facilitates invasion and metastasis by regulating
miR-205-5p/c-FLIP axis in osteosarcoma. Cancer Manag Res.
12:1229–1240. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Zhang Y, Chang B, Zhang J and Wu X: lncRNA
SOX2OT alleviates the high glucose-induced podocytes injury through
autophagy induction by the miR-9/SIRT1 axis. Exp Mol Pathol.
110:1042832019. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Perez Vidakovics MLA, Ure AE, Arrias PN,
Romanowski V and Gómez RM: Junin virus induces autophagy in human
A549 cells. PLoS One. 14:e02187302019. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Neubert P, Weichselbaum A, Reitinger C,
Schatz V, Schröder A, Ferdinand JR, Simon M, Bär AL, Brochhausen C,
Gerlach RG, et al: HIF1A and NFAT5 coordinate
Na+-boosted antibacterial defense via enhanced autophagy
and autolysosomal targeting. Autophagy. 15:1899–1916. 2019.
View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Nong Q, Li S, Wu Y and Liu D: lncRNA
COL1A2-AS1 inhibits the scar fibroblasts proliferation via
regulating miR-21/Smad7 pathway. Biochem Biophys Res Commun.
495:319–324. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Wang X, Chu J, Wen CJ, Fu SB, Qian YL, Wo
Y, Wang C and Wang DR: Functional characterization of TRAP1-like
protein involved in modulating fibrotic processes mediated by
TGF-β/Smad signaling in hypertrophic scar fibroblasts. Exp Cell
Res. 332:202–211. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Pang K, Li B, Tang Z, Yang W, Hao L, Shi
Z, Zhang J, Cai L, Li R, Liu Y, et al: Resveratrol inhibits
hypertrophic scars formation by activating autophagy via the
miR-4654/Rheb axis. Mol Med Rep. 22:3440–3452. 2020.PubMed/NCBI
|
|
39
|
Shi W, Wu Y and Bian D: p75NTR silencing
inhibits proliferation, migration and extracellular matrix
deposition of hypertrophic scar fibroblasts by activating autophagy
through inhibiting PI3K/Akt/mTOR pathway. Can J Physiol Pharmacol.
Jul 29–2020.(Epub ahead of print). doi: 10.1139/cjpp-2020-0219.
View Article : Google Scholar
|
|
40
|
De Risi M, Torromino G, Tufano M, Moriceau
S, Pignataro A, Rivagorda M, Carrano N, Middei S, Settembre C,
Ammassari-Teule M, et al: Mechanisms by which autophagy regulates
memory capacity in ageing. Aging Cell. 19:e131892020. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Lanzillotta C, Zuliani I, Vasavda C,
Snyder SH, Paul BD, Perluigi M, Di Domenico F and Barone E: BVR-A
deficiency leads to autophagy impairment through the dysregulation
of AMPK/mTOR axis in the brain-implications for neurodegeneration.
Antioxidants (Basel). 9:6712020. View Article : Google Scholar
|
|
42
|
Long J and Pi X: Polyphyllin I promoted
melanoma cells autophagy and apoptosis via PI3K/Akt/mTOR signaling
pathway. Biomed Res Int. 2020:51494172020. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Luo M, Liu Y, Liu N, Shao W, Ming L, Liu J
and Xie Y: Proscillaridin A inhibits hepatocellular carcinoma
progression through inducing mitochondrial damage and autophagy.
Acta Biochim Biophys Sin (Shanghai). Nov 17–2020.(Epub ahead of
print). doi: 10.1093/abbs/gmaa139. View Article : Google Scholar
|
|
44
|
Yang J, Wang B, Xu Q, Yang Y, Hou L, Yin
K, Guo Q, Hua Y, Zhang L, Li Y, et al: TMEM166 inhibits cell
proliferation, migration and invasion in hepatocellular carcinoma
via upregulating TP53. Mol Cell Biochem. Nov 16–2020.(Epub ahead of
print). doi: 10.1007/s11010-020-03979-1. View Article : Google Scholar
|
|
45
|
Yang Z, Tang Y, Lu H, Shi B, Ye Y, Xu G
and Zhao Q: Long non-coding RNA reprogramming (lncRNA-ROR)
regulates cell apoptosis and autophagy in chondrocytes. J Cell
Biochem. 119:8432–8440. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Urut F, Dede S, Yuksek V, Cetin S, Usta A
and Taspinar M: In vitro evaluation of the apoptotic, autophagic,
and necrotic molecular pathways of fluoride. Biol Trace Elem Res.
Nov 16–2020.(Epub ahead of print). doi: 10.1007/s12011-020-02491-3.
View Article : Google Scholar : PubMed/NCBI
|