In the fields of plastic surgery, hand and foot
surgery, and burn treatment, skin flaps are particularly important
for the closure of tissue defects. However, the ability to
attenuate or even abolish the necrosis that occurs on skin flaps,
mainly due to ischemia, remains challenging. Necrosis of the skin
flap is not only related to anatomical factors of the skin flap
itself (1–3), but also to some high-risk factors of
the patient, such as age and diabetes (4–7). For
example, Las et al (8)
reported that diabetes, excessive anesthesia time and postoperative
wound infection are risk factors for the failure of free flaps.
Bekara et al (5) showed that
an age older than 60 years, or the presence of diabetes and
arterial diseases are risk factors for complications of lower
extremity pedicle perforator flaps. In addition, Sanati-Mehrizy
et al (4) reported that
smoking and operation time are risk factors for free flap necrosis
and de Blacam et al (7)
found that venous congestion and increased age are risk factors for
pedicle flap failure. Current literature regarding the risk factors
of skin flap necrosis mainly focus on free skin flaps and lower
limb pedicled skin flaps (8–15), and
treatment methods that have been proposed are based on various
mechanisms such as surgical delay, chemical delay, extracorporeal
shock wave therapy, local thermal pretreatment, percutaneous
neuroelectric pretreatment, cold pretreatment, negative pressure
suction, targeted gene therapy, and drug injection (16–25).
Heat shock proteins (Hsps), which are highly evolutionarily
conserved from prokaryotes to human (26), are molecular chaperons that exhibit
a variety of biological activities including anti-oxidative,
anti-apoptotic and anti-inflammatory effects (27). In recent years, several Hsps have
been shown to prevent skin flap necrosis by reducing inflammation,
oxidative stress, apoptosis and regulating platelet activity
(28–32).
Hsp32, a member of the Hsp family also known as heme
oxygenase (HO)-1, is the most commonly studied molecule in the HO
family of proteins. However, clinical data concerning HO-1 in flap
surgery are scarce. This review summarizes the biological
characteristics of HO-1 and the functional significance of its
products. In particular, this review focuses on the role of carbon
monoxide (CO), one of the primary metabolites of HO-1, in flap
survival, elaborates on the protective effect of HO-1 on skin flaps
(Table I), and discusses the
feasibility and existing challenges of HO-1 in flap surgery.
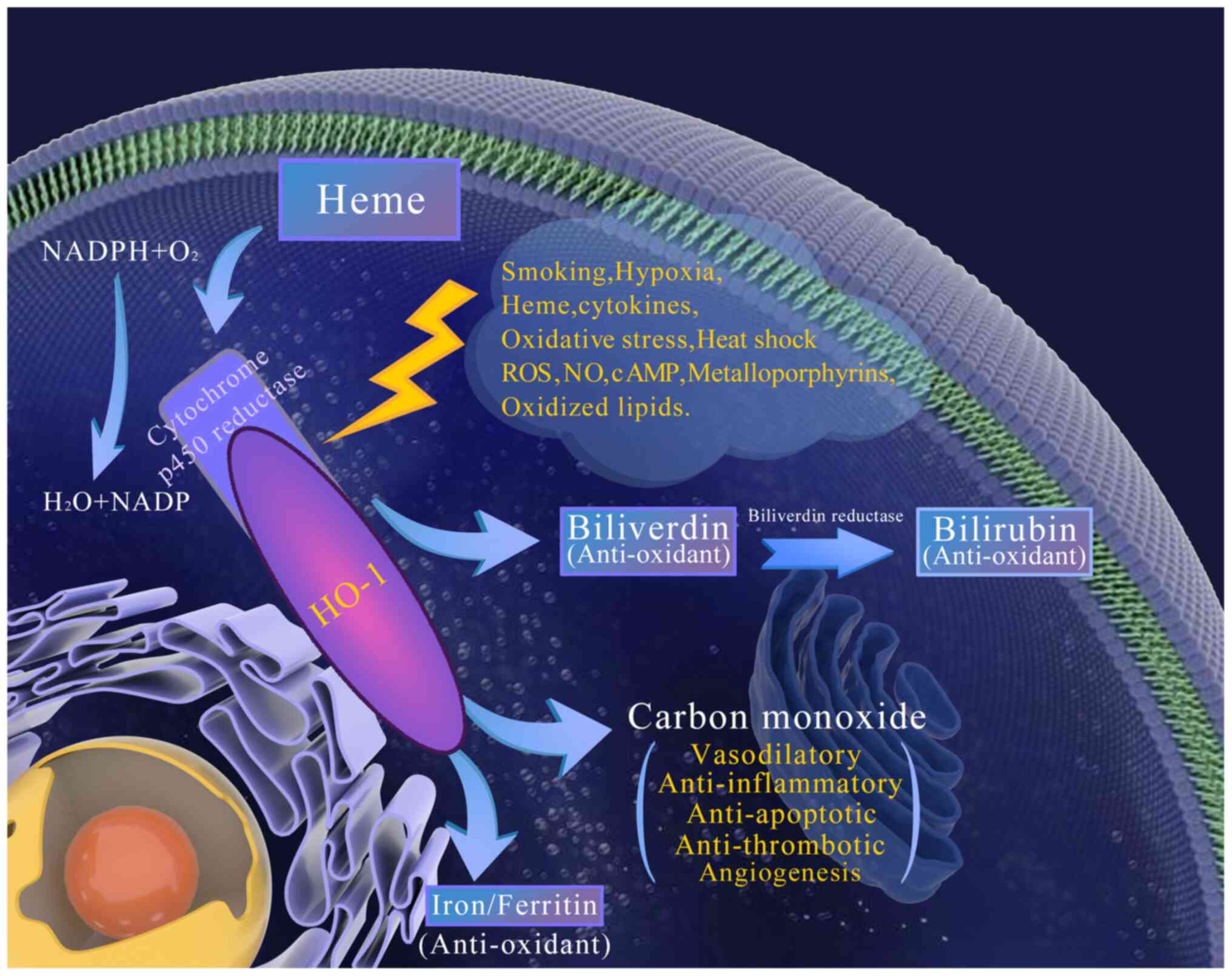
The HO family consists of three members: HO-1, HO-2
and HO-3. HO-1 is the rate-limiting enzyme of heme metabolism
(34), but the functions of HO-2
and HO-3 remain elusive. HO-1, with a molecular weight of 32 kDa,
is a stress protein that is either not expressed or has
exceptionally low expression under normal conditions. However, HO-1
can be induced by a variety of stimuli, including heavy metals,
heat shock, inflammatory stimuli, heme and its derivatives, stress,
hypoxia, and biological hormones (35–39).
Increased HO-1 expression under stress reduces protein oxidation
and lipid peroxidation, and attenuates cell and blood vessel
damage, thus playing a protective role (40,41).
For instance, Chen et al (40) reported that stress significantly
increases HO-1 expression in the heart, which in turn provides
cardioprotection. On the other hand, most metalloporphyrins,
including tin protoporphyrin and zinc protoporphyrin, are
inhibitors of HO-1 and compete with heme for the HO-1 binding site,
thereby inhibiting the biological effects of HO-1 (42,43).
The molecular weights of HO-2 and HO-3 are 36 kDa and 33 kDa,
respectively (44). It is known
that adrenal cortex hormones can induce the expression of HO-2, and
it is therefore speculated that HO-2 can regulate functions within
the nervous system. HO-3 has 90% primary structure homology with
HO-2 but does not have any enzymatic activity. It is speculated
that HO-3 regulates heme-dependent functions within cells (45).
HO-1 exists in microsomes and is the rate-limiting
enzyme for heme catabolism, which splits heme into biliverdin, CO
and iron. Biliverdin is further reduced to bilirubin under the
action of biliverdin reductase, which is an effective anti-oxidant
that protects cells from oxidative stress (46,47).
Another metabolite, CO, acts on soluble guanylate cyclase (sGC) to
increase the production of cyclic guanosine monophosphate (cGMP),
which serves as a second messenger to regulate a wide spectrum of
cellular events, including vasodilation (48,49) as
well as anti-inflammatory and anti-apoptotic activities (50) (Fig.
1).
Heme is a pro-oxidant molecule that participates in
the formation of oxidative free radicals leading to oxidative
damage (51). In a model of heme
overload using nude mice, excessive heme in the plasma promotes the
production of reactive oxygen species (ROS) and reduces the
effectiveness of nitric oxide (NO), thereby affecting the expansion
of blood vessels (52). HO-1
degrades heme, and therefore has anti-oxidant activity. Balla et
al (51) showed that the
combination of heme and ferritin, together with HO-1, prevents
oxidative stress. In another study performed in cultured alveolar
epithelial cells, HO-1 overexpression was found to lead to a nearly
3-fold increase in ferritin, which was accompanied by a
compensatory increase in transferrin receptors, subsequently
altering the distribution of iron in the cells and protecting cells
from iron toxicity induced by heme degradation (53).
The anti-inflammatory activity of CO and
CO-releasing molecules (CORM) has been well documented in various
animal models. Increased HO-1 expression or CO prevents impairment
of the blood-brain barrier (BBB), cerebral microvascular congestion
and neuroinflammation (54).
Mechanistically, this protection is achieved by the binding of CO
to hemoglobin to prevent its oxidation and the production of free
hemoglobin (54). In addition, a
carrageenan-induced inflammation model showed that CO inhibits
neutrophil migration and white blood cell adhesion (55). On the other hand, CORM-2 reduces the
accumulation of inflammatory cells, the expression of intercellular
adhesion molecule-1 (ICAM-1) and the activation of nuclear factor
κB (NF-κB) in septic mice (56).
In vitro experimental study confirmed that the mechanisms
behind the anti-inflammatory effect of CORM-2 are related to the
decreased expression of NF-κB-dependent vascular endothelial cell
adhesion molecules and secretion of inducible nitric oxide synthase
(iNOS) (56). Using an animal model
related to human sepsis (cecal ligation and puncture in rodents),
CORM-2 or CORM-3 was found to reduce the migration of
polymorphonuclear (PMN) leukocytes to purulent tissues, thereby
attenuating inflammation and saving animals from succumbing to
sepsis (57–59). In HO-1-deficient mice, treatment
with CORM-2 was found to ameliorate the severity of sepsis
associated with Enterococcus faecalis infection (60). CORM-3 was found to have a
bactericidal effect on Pseudomonas aeruginosa, thereby
reducing the mortality of mice with bacteremia (61). Additionally, CO exerts prominent
anti-inflammatory actions in part by reducing the release of tumor
necrosis factor α (TNF-α), interleukin (IL)-1, macrophage
inflammatory protein (MIP)-1 and IL-6 from activated macrophages
(62,63).
The beneficial effects of CO on the cardiovascular
system have been gradually discovered. Due to severe arterial
thrombosis, HO-1-deficient mice died within four days following an
aortic transplantation; however, treatment with CORM-2 was found to
improve the survival rate of HO-1-deficient mice (64). The underlying protective mechanisms
are related to the inhibition of platelet aggregation through the
activation of guanylate cyclase (65). Endogenous CO binds to and activates
sGC, subsequently increasing the concentration of cGMP and inducing
a vasodilatory response (50,66).
In a mouse model of myocardial infarction caused by coronary artery
ligation, intravenous infusion of CORM-3 prior to reperfusion was
found to reduce infarct size, fibrillation and tachycardia
(67,68). The mechanisms underlying
CORM-3-mediated cardiovascular protection involve potassium
channels, since a small amount of CORM-3 is lost when mitochondrial
ATP-dependent potassium channel inhibitors are used (67,68).
Another study also demonstrated that HO-1 promotes angiogenesis
through CO (69), and improves the
survival of tissues surrounding the blood supply by inhibiting
apoptosis of endothelial cells (70).
Many scholars have reported that CO plays an
important role in organ transplantation and preservation. CO in a
gaseous form or as a CORM can be used as a preservation adjuvant in
organ preservation solution. Using a model of allogeneic heart
transplantation in rodents, continuous inhalation of CO or CORM can
prolong the survival time of the transplanted heart (71,72).
The cardioprotective effect of CO gas is achieved by inhibiting
platelet aggregation and endothelial cell apoptosis, as well as
producing endogenous vasoconstrictors, thereby improving the
microcirculation of the graft (71,72).
Compared with the untreated control group, rabbit kidneys washed
with CORM-3 or CORM-1 and then stored in a cold solution for 24 h
showed higher rates of perfusion flow, glomerular filtration, and
sodium and glucose reabsorption (73). The above-mentioned mechanisms of
renal protection are thought to be related to CO-medicated
expansion of blood vessels, inhibition of cell apoptosis and
promotion of angiogenesis (73).
As previously mentioned, HO-1 generates biliverdin,
which is then converted to bilirubin. Both bilirubin and biliverdin
are effective oxidative inhibitors (74). Human biliverdin reductase (hBVR), a
Ser/Thr/Tyr kinase, is activated by insulin and free radicals
(75). hBVR is central to the
activation as well as the nuclear import and export of
extracellular signal-regulated kinase (ERK)1/2 (76). Insulin increases BVR tyrosine
phosphorylation and increases glucose uptake when BVR is knocked
down by small interfering RNA, suggesting an important function of
BVR in insulin signaling (77).
HO-1-derived bilirubin is an efficient scavenger of reactive oxygen
and nitrogen species (RONS) (78).
In macrophages, biliverdin induces the phosphorylation of
endothelial NO synthase (eNOS) at Ser1177, which generates eNOS. It
is well recognized that NO plays a crucial role in regulating a
wide spectrum of vascular functions, including vasorelaxation,
inhibition of leukocyte-endothelial adhesion, vascular smooth
muscle cell migration and proliferation, and platelet aggregation
(79).
In summary, each product generated by HO-1 plays an
important role in protecting cells and tissues through a variety of
mechanisms.
The survival rate of skin flaps is related to
ischemia-reperfusion injury and surgical trauma.
Ischemia-reperfusion injury, if not treated in a timely and
appropriate manner, may cause unfavorable outcomes for patients.
Both surgical trauma and ischemia-reperfusion may initiate
inflammatory responses, causing white blood cells to roll and
adhere to the capillary vein (80–82).
In the early reperfusion phase, the accumulation of activated
leukocytes and reactive oxygen metabolites aggravates the
inflammatory response and ultimately impairs endothelial integrity
(83). The initiation of the stress
response leads to increased HO-1, which significantly reduces the
adhesion of white blood cells to the endothelial surface and
lessens the impairment of the integrity of venous endothelium
(84). Rücker et al
(84) studied whether stress
conditioning-induced HO-1 could prevent an inflammatory response in
transferred osteomyocutaneous flaps. In all tissues analyzed,
control flaps presented with significant leukocyte adherence in
postcapillary venules, increased expression of intercellular
adhesion molecule-1 (ICAM-1), and disruption of endothelial
integrity. In contrast, stress conditioning induced considerable
HO-1 expression, which coincided with a significant reduction of
leukocyte adherence, ICAM-1 expression, and endothelial
hyperpermeability. Correspondingly, the inhibition of HO-1 by tin
protoporphyrin IX completely abolished the stress
conditioning-induced amelioration of the inflammatory response
(84). Hence, the protective effect
elicited by stress conditioning is mainly mediated by the induction
of HO-1, which reduces oxygen free radicals, ICAM-1, adherence of
white blood cells to the endothelial surface, endothelial
permeability and macromolecular extravasation, ultimately reducing
the overall inflammatory response.
Rats undergoing flap reconstruction surgery
experience lipid peroxidation and vascular damage, which is related
to ischemia-reperfusion injury (85). Ischemia-reperfusion injury involves
a complex oxidation process, which is closely related to flap
survival (86). Oxidative stress
induces an excessive activation of inflammatory processes, which
increases ROS. ROS are molecules known to cause tissue damage
through multiple mechanisms, including altering the structure and
chemistry of proteins, lipids, and nucleic acids. Hence, reducing
the release of ROS or removing excessive ROS protects tissues from
ischemia-reperfusion injury (87).
Lin et al (88) reported
that CORM-2 induces HO-1 expression, thereby reducing protein
kinase C (PKC)/amino acid-rich tyrosine kinase 2 (Pyk2)-dependent
production of ROS. Lin et al (28) showed that ginkgolide B reduced skin
flap necrosis and improved the survival of island perforator flaps
by inhibiting endoplasmic reticulum stress and oxidative stress. In
this study, they further showed that ginkgolide B activated nuclear
factor erythroid 2-related factor 2 (Nrf2) signaling. Following its
activation, Nrf2 is transported to the nucleus, where it binds to
antioxidant response elements located in the cis-regulatory
sequences of antioxidant-related enzymes and proteins and increases
their expression (89).
However, research has also shown that HO-1 may have
dual functions. For example, HO-1 has a dual role in cancer cells.
The levels of cellular iron and ROS are the determining factors for
the role of HO-1, in which excessive cellular iron and ROS tend to
push HO-1 from a protective role to that of a perpetrator. In
general, a moderate level of HO-1 activation exerts a
cytoprotective effect, while the over-activation of HO-1 becomes
cytotoxic due to the excessive increase of labile Fe2+
behind the buffering capacity of ferritin (97). Similarly, HO-1 also has a dual role
in nerve cells. Nrf2-dependent activation of HO-1 is generally
linked to protective effects in neurons and glial cells, while
Nrf2-independent activation of HO-1, which often involves AP-1 or
NF-κB, seems to exert neurotoxic effects. Indeed, HO-1 expression
is associated with neuronal damage and neurodegeneration,
especially in Alzheimer's and Parkinson's disease (98).
The application of CO, the catalytic product of
HO-1, within a clinical setting also has the following issues. For
example, the safety constraints of CO require clinical dosing to
maintain levels of carboxyhemoglobin (CO-Hb) that are under 14%,
which is in stark contrast to the preclinical frontrunner studies
with protocols that had sustained CO-Hb levels above 20% (99). Also, the pharmacokinetics of CO
inhalation protocols varied significantly by species. Rodent models
achieve therapeutic levels quickly, whereas the time to steady
state in humans is roughly three times longer based on respiratory
rate and cardiac output (100).
Similarly, the application of HO-1 or its
metabolites to the field of flap surgery will also face the
above-mentioned issues. Instead, perhaps it would be possible to:
i) use HO-1 inhibitors (such as tin protoporphyrin) to repress its
overexpression; ii) use an occasional monitoring system to achieve
feedback to control the level of CO in the body; or iii) search for
molecules that promote the release of CO or solutions that can
dissolve CO, and apply them locally to reduce systemic adverse
reactions.
HO-1 was first described in the late 1960s and its
functions have been gradually characterized over the following
decades, innovative observations have been made over the past 20
years uncovering the cellular cytoprotective capability of HO-1 in
a clinical setting. While studies have defined to some degree the
tissue protective roles of each of the by-products generated by
HO-1 (including CO) and attempted to harness HO-1 to improve
CO-delivery for clinical applications, there are still significant
limitations that prevent its use at the bed side (99). Previous studies have shown the
clinical relevance of HO-1, such as in ischemic stroke (101), skin health (102) and cardiovascular syndromes and
co-morbidities (103). However,
there are few studies on the application of HO-1 in the field of
human flap surgery. How to direct the clinical application of HO-1
and its related products in the field of flap surgery in order to
obtain more appreciable outcomes will be the focus of future
research.
The authors would like to thanks Dr Huanfa Yi from
the Second Department of the First Hospital of Jilin University,
who provided critical comments and invaluable advice on this
manuscript.
No funding was received.
Not applicable.
YZ drafted the manuscript. ZL assisted with the
literature search. MY assisted with drafting and revising the
manuscript. XG conceived and designed the review. YZ and XG are
responsible for confirming the authenticity of the raw data. All
authors read and approved the final manuscript and agree to be
accountable for the accuracy and referencing of the information
included in the review.
Not applicable.
Not applicable.
The authors declare that they have no competing
interests.
|
1
|
Taylor GI, Corlett RJ, Dhar SC and Ashton
MW: The anatomical (angiosome) and clinical territories of
cutaneous perforating arteries: Development of the concept and
designing safe flaps. Plast Reconstr Surg. 127:1447–1459. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Saint-Cyr M, Wong C, Schaverien M,
Mojallal A and Rohrich RJ: The perforasome theory: Vascular anatomy
and clinical implications. Plast Reconstr Surg. 124:1529–1544.
2009. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Callegari PR, Taylor GI, Caddy CM and
Minabe T: An anatomic review of the delay phenomenon: I.
Experimental studies. Plast Reconstr Surg. 89:397–407; discussion
417-398. 1992. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Sanati-Mehrizy P, Massenburg BB, Rozehnal
JM, Ingargiola MJ, Hernandez Rosa J and Taub PJ: Risk factors
leading to free flap failure: Analysis from the national surgical
quality improvement program database. J Craniofac Surg.
27:1956–1964. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Bekara F, Herlin C, Mojallal A, Sinna R,
Ayestaray B, Letois F, Chavoin JP, Garrido I, Grolleau JL and
Chaput B: A systematic review and meta-analysis of
perforator-pedicled propeller flaps in lower extremity defects:
Identification of risk factors for complications. Plast Reconstr
Surg. 137:314–331. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Wong AK, Joanna Nguyen T, Peric M, Shahabi
A, Vidar EN, Hwang BH, Niknam Leilabadi S, Chan LS and Urata MM:
Analysis of risk factors associated with microvascular free flap
failure using a multi-institutional database. Microsurgery.
35:6–12. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
de Blacam C, Colakoglu S, Ogunleye AA,
Nguyen JT, Ibrahim AM, Lin SJ, Kim PS and Lee BT: Risk factors
associated with complications in lower-extremity reconstruction
with the distally based sural flap: A systematic review and pooled
analysis. J Plast Reconstr Aesthet Surg. 67:607–616. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Las DE, de Jong T, Zuidam JM, Verweij NM,
Hovius SE and Mureau MA: Identification of independent risk factors
for flap failure: A retrospective analysis of 1530 free flaps for
breast, head and neck and extremity reconstruction. J Plast
Reconstr Aesthet Surg. 69:894–906. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Li B, Chang SM, Du SC, Zhuang L and Hu SJ:
Distally based sural adipofascial turnover flap for coverage of
complicated wound in the foot and ankle region. Ann Plast Surg.
84:580–587. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Lee ZH, Abdou SA, Ramly EP, Daar DA,
Stranix JT, Anzai L, Saadeh PB, Levine JP and Thanik VD: Larger
free flap size is associated with increased complications in lower
extremity trauma reconstruction. Microsurgery. 40:473–478. 2020.
View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Stranix JT, Lee ZH, Anzai L, Jacoby A,
Avraham T, Saadeh PB, Levine JP and Thanik VD: Optimizing venous
outflow in reconstruction of Gustilo IIIB lower extremity traumas
with soft tissue free flap coverage: Are two veins better than one?
Microsurgery. 38:745–751. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Gupta A, Lakhiani C, Lim BH, Aho JM,
Goodwin A, Tregaskiss A, Lee M, Scheker L and Saint-Cyr M: Free
tissue transfer to the traumatized upper extremity: Risk factors
for postoperative complications in 282 cases. J Plast Reconstr
Aesthet Surg. 68:1184–1190. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Reece EM, Bonelli MA, Livingston T,
Mulligan PS, Rockwood J, Wilson JR, Zoldos J and Champagne L:
Factors in free fasciocutaneous flap complications: A logistic
regression analysis. Plast Reconstr Surg. 136:54e–58e. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Innocenti M, Menichini G, Baldrighi C,
Delcroix L, Vignini L and Tos P: Are there risk factors for
complications of perforator-based propeller flaps for
lower-extremity reconstruction? Clin Orthop Relat Res.
472:2276–2286. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Dolan RT, Butler JS, Murphy SM and Cronin
KJ: Health-related quality of life, surgical and aesthetic outcomes
following microvascular free flap reconstructions: An 8-year
institutional review. Ann R Coll Surg Engl. 94:43–51. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Ellabban MA, Fattah IOA, Kader GA, Eldeen
OS, Mehana AE, Khodeer DM, Hosny H, Elbasiouny MS and Masadeh S:
The effects of sildenafil and/or nitroglycerin on random-pattern
skin flaps after nicotine application in rats. Sci Rep.
10:32122020. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Lee W, Oh W, Oh SM and Yang EJ:
Comparative effectiveness of different interventions of
perivascular hyaluronidase. Plast Reconstr Surg. 145:957–964. 2020.
View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Giatsidis G, Cheng L, Haddad A, Ji K,
Succar J, Lancerotto L, Lujan-Hernandez J, Fiorina P, Matsumine H
and Orgill DP: Noninvasive induction of angiogenesis in tissues by
external suction: Sequential optimization for use in reconstructive
surgery. Angiogenesis. 21:61–78. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Giatsidis G, Cheng L, Facchin F, Haddad A,
Lujan-Hernandez J, Lancerotto L, Nabzdyk CG, Matsumine H and Orgill
DP: Moderate-intensity intermittent external volume expansion
optimizes the soft-tissue response in a murine model. Plast
Reconstr Surg. 139:882–890. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Doğan F and Özyazgan İ: Flap
preconditioning by electrical stimulation as an alternative to
surgical delay: Experimental study. Ann Plast Surg. 75:560–564.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Menevşe GT, TeomanTellioglu A, Altuntas N,
Cömert A and Tekdemir I: Polidocanol injection for chemical delay
and its effect on the survival of rat dorsal skin flaps. J Plast
Reconstr Aesthet Surg. 67:851–856. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Tobalem M, Wettstein R, Pittet-Cuénod B,
Vigato E, Machens HG, Lohmeyer JA, Rezaeian F and Harder Y: Local
shockwave-induced capillary recruitment improves survival of
musculocutaneous flaps. J Surg Res. 184:1196–1204. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Kubulus D, Amon M, Roesken F, Rücker M,
Bauer I and Menger MD: Experimental cooling-induced preconditioning
attenuates skin flap failure. Br J Surg. 92:1432–1438. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Morris SF and Taylor GI: The time sequence
of the delay phenomenon: When is a surgical delay effective? An
experimental study. Plast Reconstr Surg. 95:526–533. 1995.
View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Akimoto M, Takeda A, Matsushita O, Inoue
J, Sakamoto K, Hattori M, Kounoike N and Uchinuma E: Effects of
CB-VEGF-A injection in rat flap models for improved survival. Plast
Reconstr Surg. 131:717–725. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Lindquist S and Craig EA: The heat-shock
proteins. Annu Rev Genet. 22:631–677. 1988. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Miller DJ and Fort PE: Heat shock proteins
regulatory role in neurodevelopment. Front Neurosci. 12:8212018.
View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Lin D, Wu H, Zhou Z, Tao Z, Jia T and Gao
W: Ginkgolide B improves multiterritory perforator flap survival by
inhibiting endoplasmic reticulum stress and oxidative stress. J
Invest Surg. 1–7. Dec 23–2019.(Epub ahead of print). View Article : Google Scholar
|
|
29
|
Kumar D, Jena GR, Ram M, Lingaraju MC,
Singh V, Prasad R, Kumawat S, Kant V, Gupta P, Tandan SK and Kumar
D: Hemin attenuated oxidative stress and inflammation to improve
wound healing in diabetic rats. Naunyn Schmiedebergs Arch
Pharmacol. 392:1435–1445. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Kankam HKN, Mehta S and Jain A: Thermal
preconditioning for surgery: A systematic review. J Plast Reconstr
Aesthet Surg. 73:1645–1664. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Park C, Lee H, Noh JS, Jin CY, Kim GY,
Hyun JW, Leem SH and Choi YH: Hemistepsin A protects human
keratinocytes against hydrogen peroxide-induced oxidative stress
through activation of the Nrf2/HO-1 signaling pathway. Arch Biochem
Biophys. 691:1085122020. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Jackson JW, Rivera-Marquez GM, Beebe K,
Tran AD, Trepel JB, Gestwicki JE, Blagg BSJ, Ohkubo S and Neckers
LM: Pharmacologic dissection of the overlapping impact of heat
shock protein family members on platelet function. J Thromb
Haemost. 18:1197–1209. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Matsunobu T, Satoh Y, Ogawa K and Shiotani
A: Heme oxygenase-1 expression in the guinea pig cochlea induced by
intense noise stimulation. Acta Otolaryngol Suppl. 18–23. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Harder Y, Amon M, Schramm R, Georgi M,
Banic A, Erni D and Menger MD: Heat shock preconditioning reduces
ischemic tissue necrosis by heat shock protein (HSP)-32-mediated
improvement of the microcirculation rather than induction of
ischemic tolerance. Ann Surg. 242:869–878; discussion 878-869.
2005. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Tian W, Bonkovsky HL, Shibahara S and
Cohen DM: Urea and hypertonicity increase expression of heme
oxygenase-1 in murine renal medullary cells. Am J Physiol Renal
Physiol. 281:F983–F991. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Rücker M, Schäfer T, Roesken F, Spitzer
WJ, Bauer M and Menger MD: Local heat-shock priming-induced
improvement in microvascular perfusion in osteomyocutaneous flaps
is mediated by heat-shock protein 32. Br J Surg. 88:450–457. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Chang YC, Lai CC, Lin LF, Ni WF and Tsai
CH: The up-regulation of heme oxygenase-1 expression in human
gingival fibroblasts stimulated with nicotine. J Periodontal Res.
40:252–257. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Contaldo C, Harder Y, Plock J, Banic A,
Jakob SM and Erni D: The influence of local and systemic
preconditioning on oxygenation, metabolism and survival in
critically ischaemic skin flaps in pigs. J Plast Reconstr Aesthet
Surg. 60:1182–1192. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Schürmann C, Seitz O, Klein C, Sader R,
Pfeilschifter J, Mühl H, Goren I and Frank S: Tight spatial and
temporal control in dynamic basal to distal migration of epithelial
inflammatory responses and infiltration of cytoprotective
macrophages determine healing skin flap transplants in mice. Ann
Surg. 249:519–534. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Chen YH, Lin SJ, Lin MW, Tsai HL, Kuo SS,
Chen JW, Charng MJ, Wu TC, Chen LC, Ding YA, et al: Microsatellite
polymorphism in promoter of heme oxygenase-1 gene is associated
with susceptibility to coronary artery disease in type 2 diabetic
patients. Hum Genet. 111:1–8. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Schillinger M, Exner M, Mlekusch W, Ahmadi
R, Rumpold H, Mannhalter C, Wagner O and Minar E: Heme oxygenase-1
genotype is a vascular anti-inflammatory factor following balloon
angioplasty. J Endovasc Ther. 9:385–394. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Whitington PF, Moscioni AD and Gartner LM:
The effect of tin (IV)-protoporphyrin-IX on bilirubin production
and excretion in the rat. Pediatr Res. 21:487–491. 1987. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Vreman HJ, Ekstrand BC and Stevenson DK:
Selection of metalloporphyrin heme oxygenase inhibitors based on
potency and photoreactivity. Pediatr Res. 33:195–200. 1993.
View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Elbirt KK and Bonkovsky HL: Heme
oxygenase: Recent advances in understanding its regulation and
role. Proc Assoc Am Physicians. 111:438–447. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Adach W, Błaszczyk M and Olas B: Carbon
monoxide and its donors-chemical and biological properties. Chem
Biol Interact. 318:1089732020. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Sorrenti V: Editorial of special issue
‘protective and detrimental role of heme oxygenase-1’. Int J Mol
Sci. 20:47442019. View Article : Google Scholar
|
|
47
|
Sha JY, Zhou YD, Yang JY, Leng J, Li JH,
Hu JN, Liu W, Jiang S, Wang YP, Chen C and Li W: Maltol
(3-Hydroxy-2-methyl-4-pyrone) slows d-galactose-induced brain aging
process by damping the Nrf2/HO-1-mediated oxidative stress in mice.
J Agric Food Chem. 67:10342–10351. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Schallner N, Romão CC, Biermann J, Lagrèze
WA, Otterbein LE, Buerkle H, Loop T and Goebel U: Carbon monoxide
abrogates ischemic insult to neuronal cells via the soluble
guanylate cyclase-cGMP pathway. PLoS One. 8:e606722013. View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Ndisang JF, Wu L, Zhao W and Wang R:
Induction of heme oxygenase-1 and stimulation of cGMP production by
hemin in aortic tissues from hypertensive rats. Blood.
101:3893–3900. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Motterlini R and Otterbein LE: The
therapeutic potential of carbon monoxide. Nat Rev Drug Discov.
9:728–743. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Balla G, Jacob HS, Balla J, Rosenberg M,
Nath K, Apple F, Eaton JW and Vercellotti GM: Ferritin: A
cytoprotective antioxidant strategem of endothelium. J Biol Chem.
267:18148–18153. 1992. View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Vinchi F, De Franceschi L, Ghigo A, Townes
T, Cimino J, Silengo L, Hirsch E, Altruda F and Tolosano E:
Hemopexin therapy improves cardiovascular function by preventing
heme-induced endothelial toxicity in mouse models of hemolytic
diseases. Circulation. 127:1317–1329. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
53
|
Fang R and Aust AE: Induction of ferritin
synthesis in human lung epithelial cells treated with crocidolite
asbestos. Arch Biochem Biophys. 340:369–375. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
54
|
Pamplona A, Ferreira A, Balla J, Jeney V,
Balla G, Epiphanio S, Chora A, Rodrigues CD, Gregoire IP,
Cunha-Rodrigues M, et al: Heme oxygenase-1 and carbon monoxide
suppress the pathogenesis of experimental cerebral malaria. Nat
Med. 13:703–710. 2007. View
Article : Google Scholar : PubMed/NCBI
|
|
55
|
Freitas A, Alves-Filho JC, Secco DD, Neto
AF, Ferreira SH, Barja-Fidalgo C and Cunha FQ: Heme
oxygenase/carbon monoxide-biliverdin pathway down regulates
neutrophil rolling, adhesion and migration in acute inflammation.
Br J Pharmacol. 149:345–354. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
56
|
Cepinskas G, Katada K, Bihari A and Potter
RF: Carbon monoxide liberated from carbon monoxide-releasing
molecule CORM-2 attenuates inflammation in the liver of septic
mice. Am J Physiol Gastrointest Liver Physiol. 294:G184–G191. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
57
|
Mizuguchi S, Stephen J, Bihari R, Markovic
N, Suehiro S, Capretta A, Potter RF and Cepinskas G: CORM-3-derived
CO modulates polymorphonuclear leukocyte migration across the
vascular endothelium by reducing levels of cell surface-bound
elastase. Am J Physiol Heart Circ Physiol. 297:H920–H929. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
58
|
Tsoyi K, Lee TY, Lee YS, Kim HJ, Seo HG,
Lee JH and Chang KC: Heme-oxygenase-1 induction and carbon
monoxide-releasing molecule inhibit lipopolysaccharide
(LPS)-induced high-mobility group box 1 release in vitro and
improve survival of mice in LPS- and cecal ligation and
puncture-induced sepsis model in vivo. Mol Pharmacol. 76:173–182.
2009. View Article : Google Scholar : PubMed/NCBI
|
|
59
|
Lancel S, Hassoun SM, Favory R, Decoster
B, Motterlini R and Neviere R: Carbon monoxide rescues mice from
lethal sepsis by supporting mitochondrial energetic metabolism and
activating mitochondrial biogenesis. J Pharmacol Exp Ther.
329:641–648. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
60
|
Chung SW, Liu X, Macias AA, Baron RM and
Perrella MA: Heme oxygenase-1-derived carbon monoxide enhances the
host defense response to microbial sepsis in mice. J Clin Invest.
118:239–247. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
61
|
Desmard M, Davidge KS, Bouvet O, Morin D,
Roux D, Foresti R, Ricard JD, Denamur E, Poole RK, Montravers P, et
al: A carbon monoxide-releasing molecule (CORM-3) exerts
bactericidal activity against pseudomonas aeruginosa and
improves survival in an animal model of bacteraemia. FASEB J.
23:1023–1031. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
62
|
Morse D, Pischke SE, Zhou Z, Davis RJ,
Flavell RA, Loop T, Otterbein SL, Otterbein LE and Choi AM:
Suppression of inflammatory cytokine production by carbon monoxide
involves the JNK pathway and AP-1. J Biol Chem. 278:36993–36998.
2003. View Article : Google Scholar : PubMed/NCBI
|
|
63
|
Otterbein LE, Bach FH, Alam J, Soares M,
Tao Lu H, Wysk M, Davis RJ, Flavell RA and Choi AM: Carbon monoxide
has anti-inflammatory effects involving the mitogen-activated
protein kinase pathway. Nat Med. 6:422–428. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
64
|
Chen B, Guo L, Fan C, Bolisetty S, Joseph
R, Wright MM, Agarwal A and George JF: Carbon monoxide rescues heme
oxygenase-1-deficient mice from arterial thrombosis in allogeneic
aortic transplantation. Am J Pathol. 175:422–429. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
65
|
Brüne B and Ullrich V: Inhibition of
platelet aggregation by carbon monoxide is mediated by activation
of guanylate cyclase. Mol Pharmacol. 32:497–504. 1987.PubMed/NCBI
|
|
66
|
Wang R: Resurgence of carbon monoxide: An
endogenous gaseous vasorelaxing factor. Can J Physiol Pharmacol.
76:1–15. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
67
|
Varadi J, Lekli I, Juhasz B, Bacskay I,
Szabo G, Gesztelyi R, Szendrei L, Varga E, Bak I, Foresti R, et al:
Beneficial effects of carbon monoxide-releasing molecules on
post-ischemic myocardial recovery. Life Sci. 80:1619–1626. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
68
|
Guo Y, Stein AB, Wu WJ, Tan W, Zhu X, Li
QH, Dawn B, Motterlini R and Bolli R: Administration of a
CO-releasing molecule at the time of reperfusion reduces infarct
size in vivo. Am J Physiol Heart Circ Physiol. 286:H1649–H1653.
2004. View Article : Google Scholar : PubMed/NCBI
|
|
69
|
Józkowicz A, Huk I, Nigisch A, Weigel G,
Dietrich W, Motterlini R and Dulak J: Heme oxygenase and angiogenic
activity of endothelial cells: Stimulation by carbon monoxide and
inhibition by tin protoporphyrin-IX. Antioxid Redox Signal.
5:155–162. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
70
|
Soares MP, Usheva A, Brouard S, Berberat
PO, Gunther L, Tobiasch E and Bach FH: Modulation of endothelial
cell apoptosis by heme oxygenase-1-derived carbon monoxide.
Antioxid Redox Signal. 4:321–329. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
71
|
Clark JE, Naughton P, Shurey S, Green CJ,
Johnson TR, Mann BE, Foresti R and Motterlini R: Cardioprotective
actions by a water-soluble carbon monoxide-releasing molecule. Circ
Res. 93:e2–e8. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
72
|
Sato K, Balla J, Otterbein L, Smith RN,
Brouard S, Lin Y, Csizmadia E, Sevigny J, Robson SC, Vercellotti G,
et al: Carbon monoxide generated by heme oxygenase-1 suppresses the
rejection of mouse-to-rat cardiac transplants. J Immunol.
166:4185–4194. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
73
|
Sandouka A, Fuller BJ, Mann BE, Green CJ,
Foresti R and Motterlini R: Treatment with CO-RMs during cold
storage improves renal function at reperfusion. Kidney Int.
69:239–247. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
74
|
Jansen T and Daiber A: Direct antioxidant
properties of bilirubin and biliverdin. Is there a role for
biliverdin reductase? Front Pharmacol. 3:302012.
|
|
75
|
Maines MD, Miralem T, Lerner-Marmarosh N,
Shen J and Gibbs PE: Human biliverdin reductase, a previously
unknown activator of protein kinase C betaII. J Biol Chem.
282:8110–8122. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
76
|
Gibbs PE, Tudor C and Maines MD:
Biliverdin reductase: More than a namesake-the reductase, its
peptide fragments, and biliverdin regulate activity of the three
classes of protein kinase C. Front Pharmacol. 3:312012. View Article : Google Scholar : PubMed/NCBI
|
|
77
|
Lerner-Marmarosh N, Shen J, Torno MD,
Kravets A, Hu Z and Maines MD: Human biliverdin reductase: A member
of the insulin receptor substrate family with
serine/threonine/tyrosine kinase activity. Proc Natl Acad Sci USA.
102:7109–7114. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
78
|
Jansen T, Hortmann M, Oelze M, Opitz B,
Steven S, Schell R, Knorr M, Karbach S, Schuhmacher S, Wenzel P, et
al: Conversion of biliverdin to bilirubin by biliverdin reductase
contributes to endothelial cell protection by heme
oxygenase-1-evidence for direct and indirect antioxidant actions of
bilirubin. J Mol Cell Cardiol. 49:186–195. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
79
|
Pae HO, Son Y, Kim NH, Jeong HJ, Chang KC
and Chung HT: Role of heme oxygenase in preserving vascular
bioactive NO. Nitric Oxide. 23:251–257. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
80
|
Eppihimer MJ and Granger DN:
Ischemia/reperfusion-induced leukocyte-endothelial interactions in
postcapillary venules. Shock. 8:16–25. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
81
|
Kunkel EJ, Jung U, Bullard DC, Norman KE,
Wolitzky BA, Vestweber D, Beaudet AL and Ley K: Absence of
trauma-induced leukocyte rolling in mice deficient in both
P-selectin and intercellular adhesion molecule 1. J Exp Med.
183:57–65. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
82
|
Menger MD and Vollmar B: Adhesion
molecules as determinants of disease: From molecular biology to
surgical research. Br J Surg. 83:588–601. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
83
|
Menger MD, Pelikan S, Steiner D and
Messmer K: Microvascular ischemia-reperfusion injury in striated
muscle: Significance of ‘reflow paradox’. Am J Physiol.
263:H1901–H1906. 1992.PubMed/NCBI
|
|
84
|
Rücker M, Schäfer T, Roesken F, Spitzer
WJ, Bauer M and Menger MD: Reduction of inflammatory response in
composite flap transfer by local stress conditioning-induced
heat-shock protein 32. Surgery. 129:292–301. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
85
|
Taleb S, Moghaddas P, Rahimi Balaei M,
Taleb S, Rahimpour S, Abbasi A, Ejtemaei-Mehr S and Dehpour AR:
Metformin improves skin flap survival through nitric oxide system.
J Surg Res. 192:686–691. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
86
|
Han HH, Lim YM, Park SW, Lee SJ, Rhie JW
and Lee JH: Improved skin flap survival in venous
ischemia-reperfusion injury with the use of adipose-derived stem
cells. Microsurgery. 35:645–652. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
87
|
Forrester SJ, Kikuchi DS, Hernandes MS, Xu
Q and Griendling KK: Reactive oxygen species in metabolic and
inflammatory signaling. Circ Res. 122:877–902. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
88
|
Lin CC, Hsiao LD, Cho RL and Yang CM:
Carbon monoxide releasing molecule-2-upregulated ROS-dependent heme
oxygenase-1 axis suppresses lipopolysaccharide-induced airway
inflammation. Int J Mol Sci. 20:31572019. View Article : Google Scholar
|
|
89
|
Shi Y, Liang XC, Zhang H, Sun Q, Wu QL and
Qu L: Combination of quercetin, cinnamaldehyde and hirudin protects
rat dorsal root ganglion neurons against high glucose-induced
injury through Nrf-2/HO-1 activation and NF-κB inhibition. Chin J
Integr Med. 23:663–671. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
90
|
Brouard S, Otterbein LE, Anrather J,
Tobiasch E, Bach FH, Choi AM and Soares MP: Carbon monoxide
generated by heme oxygenase 1 suppresses endothelial cell
apoptosis. J Exp Med. 192:1015–1026. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
91
|
Edmunds MC, Czopek A, Wigmore SJ and Kluth
DC: Paradoxical effects of heme arginate on survival of
myocutaneous flaps. Am J Physiol Regul Integr Comp Physiol.
306:R10–R22. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
92
|
Harder Y, Amon M, Schramm R, Rücker M,
Scheuer C, Pittet B, Erni D and Menger MD: Ischemia-induced
up-regulation of heme oxygenase-1 protects from apoptotic cell
death and tissue necrosis. J Surg Res. 150:293–303. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
93
|
Harder Y, Contaldo C, Klenk J, Banic A,
Jakob SM and Erni D: Improved skin flap survival after local heat
preconditioning in pigs. J Surg Res. 119:100–105. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
94
|
Harder Y, Amon M, Georgi M, Scheuer C,
Schramm R, Rücker M, Pittet B, Erni D and Menger MD: Aging is
associated with an increased susceptibility to ischaemic necrosis
due to microvascular perfusion failure but not a reduction in
ischaemic tolerance. Clin Sci (Lond). 112:429–440. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
95
|
Kubulus D, Roesken F, Amon M, Rücker M,
Bauer M, Bauer I and Menger MD: Mechanism of the delay phenomenon:
Tissue protection is mediated by heme oxygenase-1. Am J Physiol
Heart Circ Physiol. 287:H2332–H2340. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
96
|
Sun Y, Li QF, Zhang Y, Hu R and Jiang H:
Isoflurane preconditioning increases survival of rat skin
random-pattern flaps by induction of HIF-1α expression. Cell
Physiol Biochem. 31:579–591. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
97
|
Chiang SK, Chen SE and Chang LC: A dual
role of heme oxygenase-1 in cancer cells. Int J Mol Sci. 20:392018.
View Article : Google Scholar
|
|
98
|
Nitti M, Piras S, Brondolo L, Marinari UM,
Pronzato MA and Furfaro AL: Heme oxygenase 1 in the nervous system:
Does it favor neuronal cell survival or induce neurodegeneration?
Int J Mol Sci. 19:22602018. View Article : Google Scholar
|
|
99
|
Hopper CP, Meinel L, Steiger C and
Otterbein LE: Where is the clinical breakthrough of heme
oxygenase-1/carbon monoxide therapeutics? Curr Pharm Des.
24:2264–2282. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
100
|
Steiger C, Hermann C and Meinel L:
Localized delivery of carbon monoxide. Eur J Pharm Biopharm.
118:3–12. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
101
|
Bereczki D Jr, Balla J and Bereczki D:
Heme oxygenase-1: Clinical relevance in ischemic stroke. Curr Pharm
Des. 24:2229–2235. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
102
|
Szabo IL, Kenyeres A, Szegedi A and
Szollosi AG: Heme oxygenase and the skin in health and disease.
Curr Pharm Des. 24:2303–2310. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
103
|
Haines DD and Tosaki A: Role of heme
oxygenases in cardiovascular syndromes and co-morbidities. Curr
Pharm Des. 24:2322–2325. 2018. View Article : Google Scholar : PubMed/NCBI
|