Introduction
Colorectal cancer (CRC) is one of the most lethal
malignancies worldwide. It is the third leading cause of
cancer-associated death and the third most commonly diagnosed
cancer in both men and women (1).
Despite improvements in treatment options, such as surgery and
adjuvant therapeutic modalities in recent years, CRC still has a
low survival rate. Thus, there is an urgent requirement to identify
the important signaling pathways involved, to aid in the
development of novel effective treatment strategies for management
of CRC.
Non-coding (nc)RNAs have been shown to be involved
in various types of disease, including cancer, and an increasing
number of studies are identifying the associations between ncRNAs
and cancer (2–4). MicroRNAs (miRNAs/miRs) are a class of
small regulatory ncRNAs and are ~22 bp in length. They are
evolutionarily conserved across species and can negatively regulate
the expression levels of their targets by binding with the
3′-untranslated region (UTR) of mRNAs (5). Increasing evidence has shown that
miRNAs serve crucial roles in oncogenesis, progression and
prognosis of CRC (6–8).
Circular (circ)RNAs are a novel group of ncRNAs,
which have covalently closed continuous loops without a
polyadenylated tail or 5′ to 3′ polarity (9). circRNAs originate from exons, introns
or intergenic regions and are resistant to RNase R (10). It has been found that circRNAs are
conserved across multiple species and display tissue-specific and
developmental-stage specific expression levels (11); however, their functional
significance remains unknown. Previously, research has focused on
investigating the function of circRNAs. Studies have shown that
circRNAs exert functions in neurological diseases, cardiovascular
disorders, prion diseases and cancer (12,13).
circRNAs are principally localized in the cytoplasm, where they
exert their key roles in human diseases (14). Accumulating data have suggested that
circRNAs can function as miRNA sponges and interact with
RNA-binding proteins to widely regulate gene expression levels
(10). circAGFG1 promoted stemness
and metastasis in CRC by regulating YY1/CTNNB1 and sponging miRNAs
(15). circHIPK3 also inhibited
cancer proliferation, acting as a miR-124 sponge in multiple types
of cancer (16). These studies
demonstrate that circRNAs are novel targets for drug development
and clinical management in cancer.
In the present study, the role of circ-0000212, a
circRNA derived from the SFMBT2 gene, in CRC was investigated.
circ-0000212 expression was significantly higher in CRC and could
promote tumor progression by acting as a competing endogenous RNA
of miR-491 by modulating forkhead box P4 (FOXP4) expression levels.
Therefore, circ-0000212 may act as a suppressive factor in tumors
and thus serve as a novel target to treat CRC.
Materials and methods
Patient characteristics
Clinical data and miRNA expression data were
downloaded from The Cancer Genome Atlas (TCGA) database using the
TCGAbiolinks package (17). A total
of 500 samples were downloaded for RNA analysis, including 41
normal tissues and 459 cancer tissues, and 465 samples were
downloaded for miRNA analysis, including 8 normal tissues and 457
cancer tissues. TCGA data are publicly available and all written
informed consent was acquired as stated by TCGA. A total of 20
pairs of CRC and adjacent normal tissues were harvested from
patients with CRC, who received surgical treatment at The Second
Affiliated Hospital of Harbin Medical University (Harbin, China).
The median age of the patients was 64 years old, (age range, 48–81
years) and among the 20 cases, seven were female patients and 13
were male patients. All tissues were snap-frozen, then stored at
−80°C until use. The study adhered to the principles of the
Declaration of Helsinki. Written informed consent was provided by
all patients and the present study was approved by the
Institutional Review Board of The Second Affiliated Hospital of
Harbin Medical University (Heilongjiang, China).
Cell culture
The normal colon-derived epithelial FHC cell line,
293T cells, and the CRC cell lines, SW620, SW480, LoVo, HT29 and
HCT116 were purchased from the American Type Culture Collection.
The FHC cells were cultured in DMEM: F12 Medium, and 293T cells and
the CRC cell lines were cultured in DMEM (all from Gibco; Thermo
Fisher Scientific, Inc.). All media were supplemented with 10% FBS
(Thermo Fisher Scientific, Inc.) and 1% penicillin-streptomycin
(Gibco; Thermo Fisher Scientific, Inc.) at 37°C in a humidified
incubator with 5% CO2.
Reverse transcription-quantitative PCR
(RT-qPCR)
TRIzol® reagent (Invitrogen; Thermo
Fisher Scientific, Inc.) was used to extract RNA from the tissues
and cells and RT was performed using a RT kit according to the
manufacturer's protocol (Thermo Fisher Scientific, Inc.). qPCR was
performed with the SYBR Green mix kit (Takara Bio, Inc.). The
thermocycling conditions were as follows: 94°C for 2 min, followed
by 40 cycles at 94°C for 15 sec and 60°C for 1 min. NE-PER™ Nuclear
and Cytoplasmic Extraction Reagents (Thermo Fisher Scientific,
Inc.) were used to isolate the cytoplasmic and nuclear components,
respectively. The following primers were used: circ-0000212
forward, 5′-GTTTGGGAGGCTTCTTGGTAAT-3′ and reverse,
5′-AAAGCCAGTTTCCTCCAAGC-3′; SFMBT2 forward,
5′-TCTGCGCTACTGCGGTTAC-3′ and reverse
5′-ACCAGTCAAGTCACGTATGAGAA-3′; U6 forward,
5′-GCTTCGGCAGCACATATACTAAAAT-3′ and reverse,
5′-CGCTTCACGAATTTGCGTGTCAT-3′; miR-491 stem loop,
5′-CTCAACTGGTGTCGTGGAGTCGGCAATTCAGTTGAGCCTCATG-3′, forward,
5′-ACACTCCAGCTGGGAGTGGGGAACCCTTC-3′ and reverse,
5′-TGGTGTCGTGGAGTCG-3′; FOXP4 forward,
5′-ATCGGCAGCTGACGCTAAATGAGA-3′ and reverse,
5′-AAACACTTGTGCAGGCTGAGGTTG-3′; actin forward,
5′-GGCGGCAACACCATGTACCCT-3′ and reverse,
5′-AGGGGCCGGACTCGTCATACT-3′. U6 and actin were used as the loading
controls.
Plasmids, synthesized nucleotide
sequences and transfection
An overexpression plasmid (pcDNA3.1) containing the
FOXP4 transcript, small interfering (si)RNA targeting circ-0000212,
miR-491 mimics and miR-491 inhibitors were purchased from Shanghai
GenePharma Co., Ltd. The following siRNA sequence targeting
circ-0000212 was used: 5′-GGGAGGCTTCTTGGTAATTTT-3′. The negative
control used for overexpression of FOXP4 was empty pcDNA3.1
plasmid. The siRNA negative control sequence was as follows:
5′-TTUUGAACCAAGAAGCCUCCC-3′. The miR-491 mimic sequence was
5′-AGUGGGGAACCCUUCCAUGAGG-3′ and negative control used for miR-491
mimics was 5′-UUCUCCGAACGUGUCACGUTT-3′. The miR-491 inhibitor
sequence was 2′-methoxy modified: 5′-mCm CmU mCm AmU mGm GmA mAm
GmG mGm UmU mCm CmC mCm AmC mU-3′, and the negative control for the
miR-491 inhibitor was 5′-CAGUACUUUUGUGUAGUACAA-3′. The miR-491
inhibitor was 2′-methoxy modified because this modification could
more effectively inhibit the function of miRNA (18). The cells were seeded 24 h prior to
transfection, at 50–60% confluence, then transfected using
Lipofectamine® 2000 (Invitrogen; Thermo Fisher
Scientific, Inc.) according to the manufacturer's protocol. The
cells were transfected with 50 nM siRNA, 2.5 µg plasmid, 30 nM
miRNA mimics or 30 nM miRNA inhibitor, according to the
manufacturer's protocol. Subsequent experiments were performed 48 h
post transfection.
Bioinformatics analysis
TargetScan online database (Release 7.2, http://www.targetscan.org/vert_72/) was used to
identify potential mRNA targets of miRNA-491. CircBase (http://www.circbase.org/) and starBase (http://starbase.sysu.edu.cn/index.php)
were used to identify potential circRNAs that could bind with
miR-491. The sequence of miR-491 is 5′-ggaguaccuuccCAAGGGGUGa-3′
and the binding site of hsa-circ-0000212 is
5′-ggagcaagugcuGCUCCCCACa-3′ (upper-case letters refer to binding
site).
Cell Counting Kit-8 (CCK-8)
proliferation assay
A total of 2×103 cells were seeded in
each well of a 96-well plate containing 100 µl culture medium. The
cells were cultured for 24, 48 and 72 h, and 10 µl CCK-8 solution
(Abcam) was then added to each well and cultured for 2 h in the
incubator. The absorbance was measured at 460 nm using a microplate
reader.
Colony formation assay
A total of 500 cells were seeded in each well of a
6-well plate and incubated at 37°C for 7 days. To maintain the
effect of the siRNA, miRNA mimic or inhibitor, the cells were
re-transfected on day 3. The colonies were fixed with 4%
paraformaldehyde at room temperature for 15 min, then stained with
0.01% crystal violet solution at room temperature for 30 min. The
number of cell colonies was then analyzed. A colony was defined to
consist of ≥50 cells and colonies were detected under a light
microscope (magnification, ×4).
Western blot analysis
The cells were harvested and the total protein was
extracted using RIPA lysis buffer with protease-phosphatase
inhibitor (EMD Millipore). A Pierce BCA Protein assay kit (Thermo
Fisher Scientific, Inc.) was used to quantify protein expression.
Proteins (30 µg) were loaded on a 15% SDS-gel, resolved using
SDS-PAGE then transferred to PVDF membranes. Membranes were blocked
using 3% non-fat milk at room temperature for 2 h. The primary
antibodies used were anti-FOXP4 (1:1,000; cat. no. b17726) and
anti-β-actin (1:2,500; cat. no. ab8227) (both from Abcam), and
membranes were incubated with the antibodies overnight at 4°C.
Subsequently, a HRP-conjugated secondary antibody, goat anti-rabbit
IgG H&L (1:5,000; cat. no. b6721; Abcam), was incubated with
the membrane at room temperature for 1 h. The membranes were
visualized using an ECL kit (Bio-Rad Laboratories, Inc.) according
to the manufacturer's protocol.
Luciferase reporter assay
The 293T cells were seeded in 96-well plates and
co-transfected with either pRL-CMV Renilla luciferase
reporter, firefly luciferase reporter, or miR-491 mimic. The
luciferase activities of Renilla and firefly reporter
vectors were quantified using a dual-luciferase reporter assay
according to the manufacturer's protocol (Promega Corporation)
after 48 h of incubation. The 3′ untranslated regions (UTRs) were
taken from FOXP4, and the binding site of circ-0000212 with miR-491
was taken from circ-0000212. The sequences containing the
miR-491-binding sites or point mutations in the binding sites were
inserted between SpeI and HindIII sites in the
firefly luciferase reporter vectors, in order to generate wild-type
and mutant luciferase reporter vectors, respectively. These
sequences were all obtained from Shanghai GenePharma Co., Ltd. The
pRL-CMV Renilla luciferase reporter was purchased from
Promega Corporation. Lipofectamine 2000 was used to perform
transfection according to the manufacturer's protocol. Briefly, the
293T cells were seeded 24 h prior to transfection at 50–60%
confluence. The cells were transfected once they reached ~70%
confluence with 50 ng plasmids, and 20 nM miRNA mimics or miRNA
inhibitor, according to the manufacturer's protocol. Luciferase
activity was measured 48 h post-transfection and Renilla
luciferase activity was used as a control.
RNA immunoprecipitation (RIP)
assay
A RIP assay was performed using a Magna RIP kit (EMD
Millipore) with samples from SW620 cells, according to the
manufacturer's protocol. The anti-AGO2 (cat. no. b156870) and IgG
antibodies (cat. no. b133470) were purchased from Abcam.
Actinomycin D and RNase R
treatment
Actinomycin D (2 mg/ml; Sigma-Aldrich; Merck KGaA)
was added to inhibit transcription. The RNA from SW620 cells (2 µg)
was treated at 37°C for 30 min with 3 U/µg RNaseR (LGC Biosearch
Technologies). After treatment with Actinomycin D and RNase R, the
RNA expression levels of SFMBT2 and circ-0000212 were determined
using RT-qPCR.
Statistical analysis
All the experiments were repeated at least three
times. Statistical analysis was performed using SPSS version 20.0
(IBM Corp.). A Student's t-test (paired or unpaired depending on
the data) or Fisher's exact test was used to compare differences
between two groups. Differences between multiple groups were
compared using a one-way ANOVA with a post hoc Tukey's test.
Kaplan-Meier curves with a log-rank test were used to assess
survival. P<0.05 was considered to indicate a statistically
significant difference.
Results
miR-491 is downregulated in CRC
tissues and associated with certain clinicopathological
characteristics of CRC
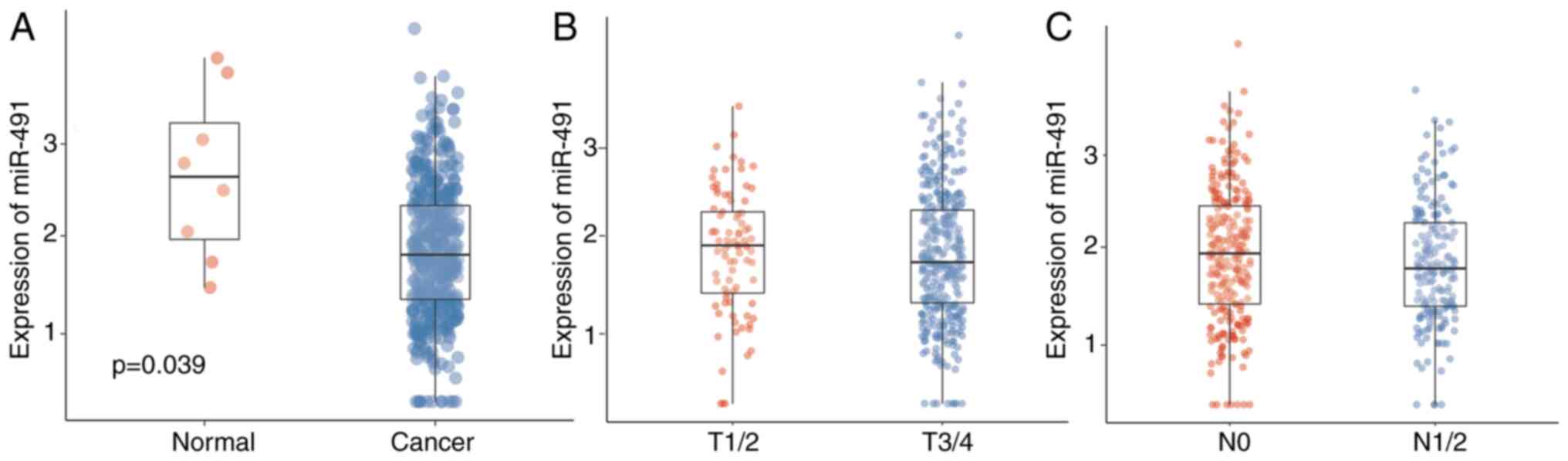
Data from TCGA was first analyzed to confirm the
expression levels of miR-491 and the potential role of miR-491 in
CRC. Table I shows the baseline
characteristics of the patients from TCGA. miR-491 was upregulated
in normal tissues compared with that in the cancer tissues
(Fig. 1A), indicating an inhibitory
effect on CRC. The expression levels of miR-491 in the cases with
different T stages was also analyzed; there were no significant
differences between the different T stages; however, late T stage
had a lower expression level of miR-491 (Fig. 1B). Similarly, there was only a trend
that patients with advanced N stage had lower miR-491 expression
levels (Fig. 1C). Patients were
further divided into two groups based on the miR-491 expression
level, and it showed that lower miR-491 expression level was
associated with advanced tumor and N staging (Table II).
 | Table I.Baseline clinicopathological
characteristics of the cases obtained from TCGA. |
Table I.
Baseline clinicopathological
characteristics of the cases obtained from TCGA.
|
Characteristics | N (%) |
|---|
| Age, years |
|
|
<60 | 124 (27.0) |
|
≥60 | 332 (72.2) |
| Sex |
|
|
Female | 216 (47.2) |
|
Male | 242 (52.8) |
| Stage |
|
| I | 76
(17.0) |
| II | 178 (39.7) |
|
III | 129 (28.8) |
| IV | 65
(14.5) |
| T stage |
|
| T1 | 11
(2.4) |
| T2 | 78
(17.0) |
| T3 | 312 (68.1) |
| T4 | 56
(12.2) |
|
Tis | 1
(0.2) |
| N stage |
|
| N0 | 269 (58.7) |
|
N1/2 | 189 (41.2) |
| M Stage |
|
| M0 | 336 (83.8) |
| M1 | 65
(16.2) |
| Radiation
therapy |
|
| No | 379 (97.7) |
|
Yes | 9
(2.3) |
| Residual tumor |
|
| R0 | 330 (85.7) |
| R1 | 4
(1.0) |
| R2 | 26
(6.8) |
| RX | 25
(6.5) |
| Histological
type |
|
|
Adenocarcinoma | 391 (86.3) |
|
Mucinous adenocarcinoma | 62
(13.7) |
| Ethnicity |
|
|
American Indiana or Alaskan native | 1
(0.4) |
|
Asian | 11
(3.9) |
| Black
or African American | 59
(20.8) |
|
White | 213 (75.0) |
| Table II.Association between miR-491 or FOXP4
expression with the clinicopathological characteristics. |
Table II.
Association between miR-491 or FOXP4
expression with the clinicopathological characteristics.
|
| miR-491, n |
| FOXP4, n |
|
|---|
|
|
|
|
|
|
|---|
|
Characteristics | High | Low | P-value | High | Low | P-value |
|---|
| Age |
|
| 0.3684 |
|
| 0.2504 |
|
<60 | 56 | 66 |
| 73 | 51 |
|
|
≥60 | 163 | 155 |
| 172 | 157 |
|
| Sex |
|
| 0.297 |
|
| 0.889 |
|
Female | 111 | 100 |
| 115 | 100 |
|
|
Male | 109 | 122 |
| 131 | 109 |
|
| Stage |
|
| 0.0455a |
|
|
<0.001c |
| I | 46 | 27 |
| 39 | 36 |
|
| II | 83 | 86 |
| 75 | 103 |
|
|
III | 53 | 72 |
| 82 | 46 |
|
| IV | 34 | 31 |
| 44 | 20 |
|
| T stage |
|
| 0.0506 |
|
| 1 |
|
T1/2 | 51 | 34 |
| 48 | 40 |
|
|
T3/4 | 169 | 187 |
| 198 | 168 |
|
| N stage |
|
| 0.026a |
|
|
<0.001c |
| N0 | 140 | 117 |
| 125 | 143 |
|
|
N1/2 | 80 | 105 |
| 121 | 66 |
|
| M Stage |
|
| 0.768 |
|
| 0.016a |
| M0 | 158 | 162 |
| 172 | 162 |
|
| M1 | 34 | 31 |
| 44 | 20 |
|
| Radiation
therapy |
|
| 0.949 |
|
| 1 |
| No | 179 | 188 |
| 202 | 175 |
|
|
Yes | 5 | 4 |
| 5 | 4 |
|
| Residual tumor |
|
| 0.009b |
|
| 0.7435 |
| R0 | 146 | 168 |
| 170 | 159 |
|
| R1 | 2 | 2 |
| 2 | 2 |
|
| R2 | 21 | 5 |
| 15 | 9 |
|
| RX | 11 | 14 |
| 12 | 13 |
|
| Histological
type |
|
| 0.397 |
|
| 0.1008 |
|
Adenocarcinoma | 184 | 192 |
| 216 | 172 |
|
|
Mucinous adenocarcinoma | 34 | 27 |
| 27 | 35 |
|
| Ethnicity |
|
| 0.014a |
|
| 0.6216 |
|
American Indiand or Alaskan native | 1 | 0 |
| 1 | 0 |
|
|
Asian | 3 | 8 |
| 5 | 6 |
|
| Black
or African American | 35 | 24 |
| 32 | 27 |
|
|
White | 82 | 131 |
| 125 | 88 |
|
miR-491 inhibits CRC cell
proliferation
Based on the aforementioned findings, it was
hypothesized that miR-491 inhibited cell malignancy. To validate
this hypothesis, miR-491 expression levels were first detected in
different CRC cell lines, as well as in a normal colon epithelial
cell line. As shown in Fig. 2A, the
CRC cell lines, particularly SW620 cells, had a relatively low
expression level of miR-491 compared with that in the FHC cell
line, indicating a potential tumor suppressor role. miR-491 was
then transiently overexpressed in the SW620 and LoVo cell lines
using miR-491 mimic (Fig. 2B), and
the CCK-8 and colony formation assays were performed. miR-491
significantly inhibited the proliferation and viability of both the
SW620 and LoVo cells (Fig.
2C-E).
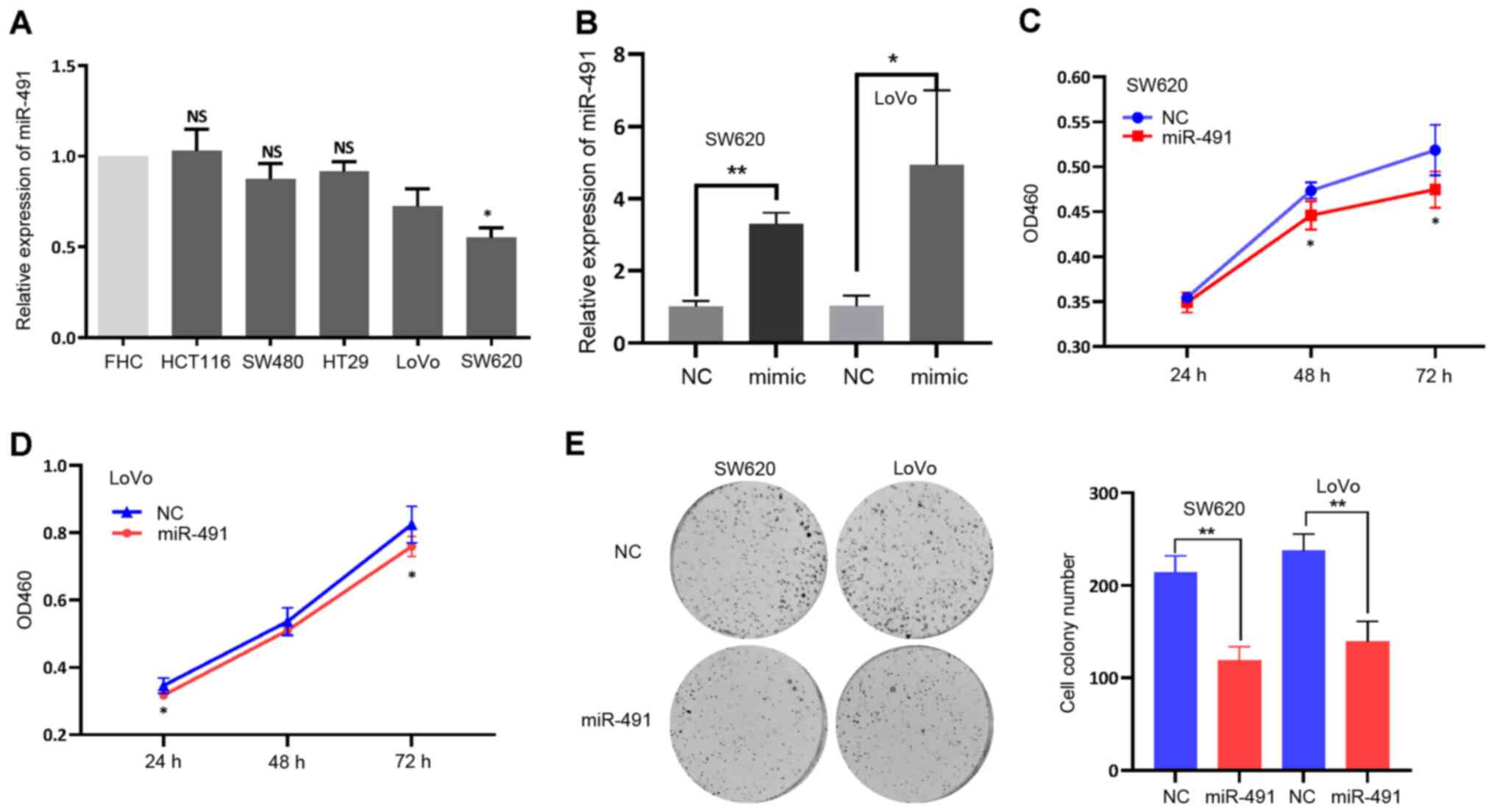 | Figure 2.miR-491 inhibits CRC cell
proliferation. (A) miR-491 expression in the different colorectal
cancer cell lines (HCT116, SW480, HT29, LoVo and SW620) and normal
colon cell line (FHC). *P<0.05 vs. FHC. (B) Expression of
miR-491 was increased after transfection of miR-491 mimic both in
LoVo and SW620 cells. miR-491 inhibited (C) SW620 and (D) LoVo cell
proliferation, based on the Cell Counting Kit-8 assays. (E) miR-491
suppressed SW620 and LoVo cell colony formation ability. Right
panel, representative figures; left panel, quantitative analysis.
*P<0.05, **P<0.01 vs. NC. Experiments were repeated three
times. miR, microRNA; CCK-8, Cell Counting Kit-8; NC, negative
control; OD, optical density; NS, not significant. |
miR-491 functionally targets
FOXP4
miRNAs exert their functions by inhibiting
downstream targets. From the data analysis of patients with CRC
from TCGA, it was found that while miR-491 was significantly
reduced in cancer tissues, the expression levels of numerous mRNAs
were significantly increased. Thus, the potential downstream
targets of miRNA regulation may be within the upregulated mRNAs.
FOXP4 was found to be upregulated in CRC tissues (adenocarcinoma
tissues) compared with that in normal tissues (Fig. 3A). Higher expression levels of FOXP4
were also associated with late tumor, N and M stages (Table II). Thus, the association between
miR-491 and FOXP4 expression levels with survival were analyzed,
and no association was found (Fig.
S1). Next, bioinformatics analysis was used to screen potential
targets of miR-491, as miRNAs could exert their functions by
inhibiting downstream targets. The results showed that FOXP4 could
bind with miR-491 and the binding sites were conserved across
different species (Fig. 3B;
Table SI). There were 2 binding
sites in the 3′-UTR of FOXP4 (Fig.
3C), indicating an interaction between miR-491 and FOXP4. To
further confirm binding between miR-491 and FOXP4, a luciferase
reporter assay was performed. The results showed that miR-491 mimic
could significantly inhibit the luciferase activity of the vector
containing the wild-type binding site compared with that in the
vector containing the mutant binding site (Fig. 3D). In addition, miR-491 reduced the
mRNA and protein expression levels of FOXP4 (Fig. 3E). It was then examined whether
FOXP4 was a functional target of miR-491. First, FOXP4 and miR-491
were overexpressed, and the results showed that FOXP4 could promote
cell colony formation ability, whereas overexpression of miR-491
reduced colony formation. Notably, FOXP4 reversed the inhibitory
effects on cell colony formation ability of miR-491 (Figs. 3F and S2A). Furthermore, similar results were
observed in the CCK-8 assay (Fig.
3G). These results confirmed that miR-491 could functionally
target FOXP4 in colon cancer.
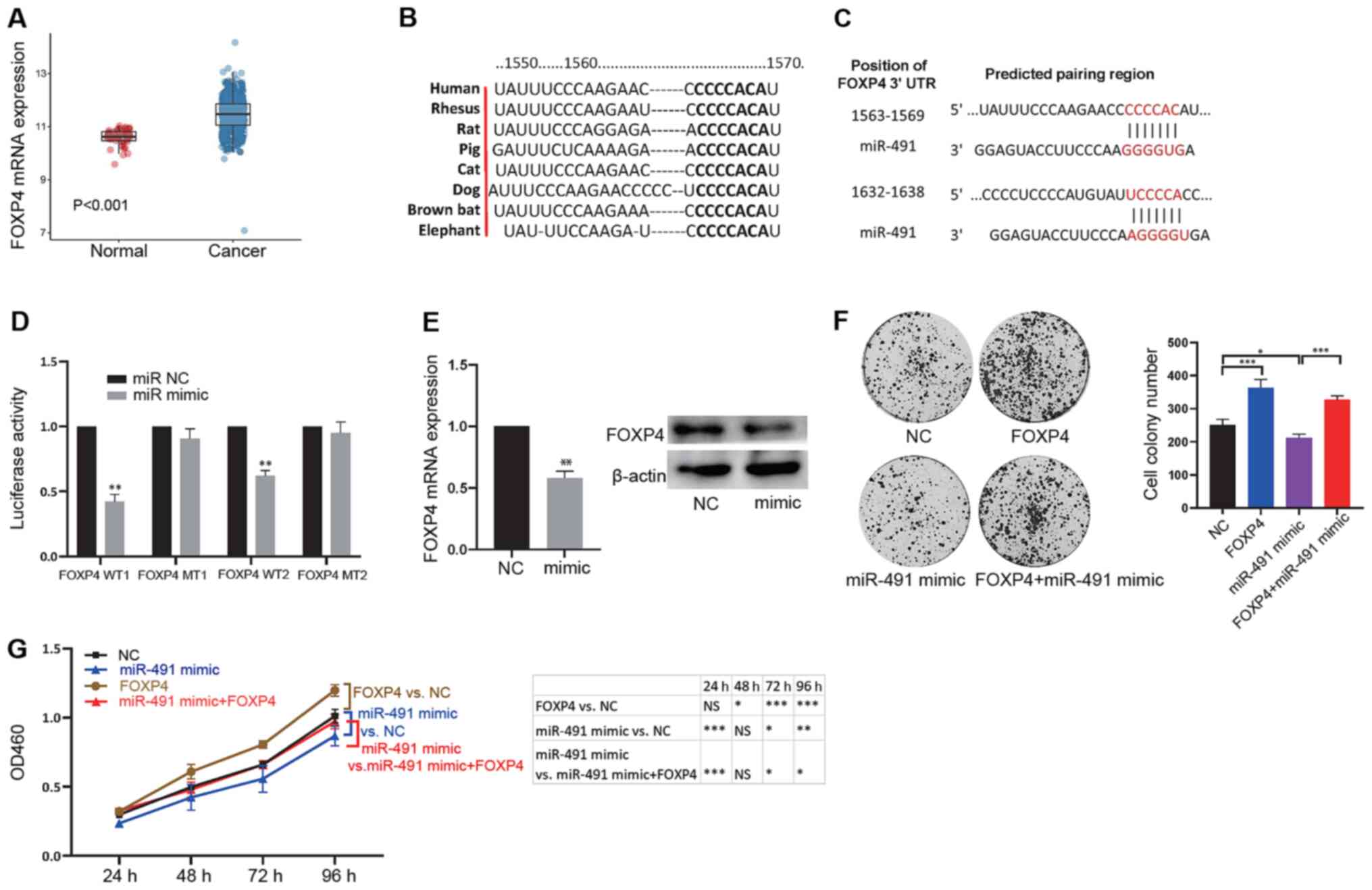 | Figure 3.miR-491 functionally targets FOXP4.
(A) FOXP4 expression was significantly increased in cancer tissues
compared with normal tissues. (B) The binding site between FOXP4
and miR-491 is conserved across different species. (C) Potential
binding sites between the 3′-UTR in FOXP4 with miR-491. (D) miR-491
mimic decreased luciferase activity of the vector containing WT
binding sites in 293T cells. *P<0.05 vs. miR NC. (E) miR-491
decreased FOXP4 mRNA (left) and protein (right) expression levels.
**P<0.01 vs. NC. (F) SW620 cells were divided into four groups:
NC, overexpression of FOXP4 (FOXP4), overexpression of miR-491
(miR-491 mimic), and concurrent overexpression of FOXP4 and miR-491
(FOXP4 + miR-491 mimic). Cell colony formation assays were
performed in the different groups of cells: Right, representative
figures; left, quantitative analysis. (G) SW620 Cells were divided
into four groups: NC, overexpression of FOXP4 (FOXP4),
overexpression of miR-491 (miR-491 mimic), and concurrent
overexpression of FOXP4 and miR-491 (FOXP4 + miR-491 mimic). Cell
Counting Kit-8 assays were performed in the different groups of
transfected cells. *P<0.05, **P<0.01, ***P<0.001, as
indicated. Experiments were repeated three times. miR-491,
microRNA-491; FOXP4, forkhead box P4; UTR, untranslated region; NC,
negative control; WT, wild-type; MT, mutant; OD, optical
density. |
Biological characteristics of
circ-0000212
An increasing number of studies have demonstrated
that circRNAs act as a novel class of miRNAs to regulate the target
expression level and therefore, may be involved in oncogenesis and
progression (19). To identify the
circRNAs that could sponge miR-491 in CRC, circBase and starBase
databases were used to identify potential circRNAs, that could bind
to and sponge miR-491 (Table SII).
As SFMBT2 is widely involved in cancer processes (20,21),
from the results, hsa-circ-0000212 was selected, which is derived
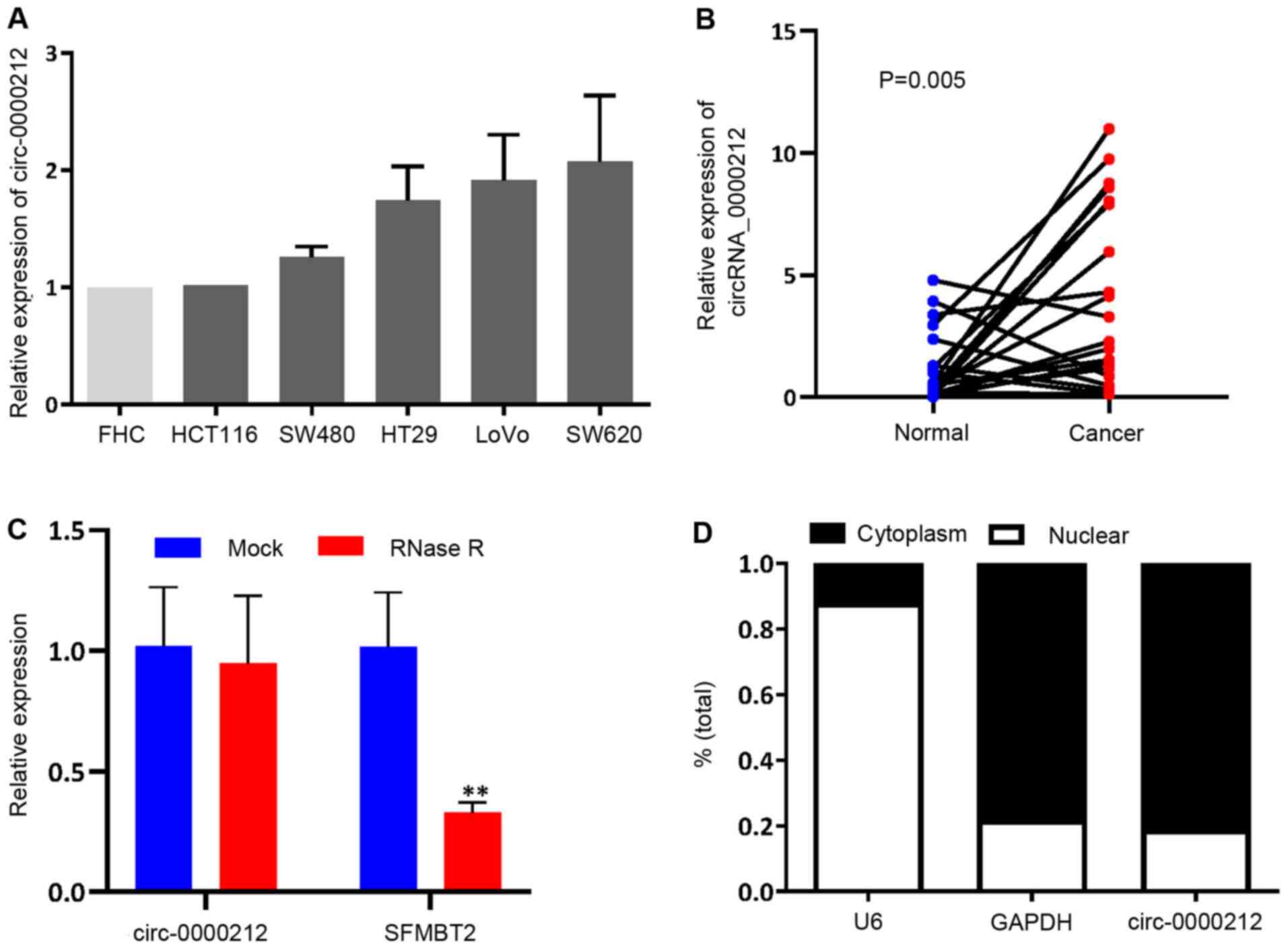
from the SFMBT2 gene (chromosome 10, 7405839-7423911). The
expression levels of circ-0000212 were assessed in the CRC cell
lines, and it was found that circ-0000212 was notably increased
when compared with that in the normal FHC cell line, although the
difference was not significant (Fig.
4A), opposite to the expression pattern of miR-491.
Furthermore, circ-0000212 was also detected in 20 paired cancer and
adjacent normal tissues, which revealed that circ-0000212 was
expressed at a higher level compared with that in the normal
adjacent tissues (Fig. 4B).
According to Fig. 4C, compared with
linear SFMBT2, circ-0000212 showed resistance to RNase R,
indicating that circ-0000212 possessed a loop structure (Fig. 4C). Cytoplasmic and nuclear RNA
analysis using RT-qPCR showed that circ-0000212 was primarily
distributed in the cytoplasm (Fig.
4D).
circ-0000212 acts as a sponge for
miR-491
Considering that circRNAs can sponge miRNAs, and
circ-0000212 was enriched in the cytoplasm, it was hypothesized
that circ-0000212 could act as a sponge for, and bind to, miR-491.
The potential binding site between circ-0000212 and miR-491 was
identified (Fig. 5A), then the
sequence was constructed and ligated downstream in the luciferase
reporter plasmid. It was hypothesized that miR-491 could decrease
the luciferase activity of circ-0000212. The luciferase reporter
vector for circ-0000212 was co-transfected with miR-491 mimic and
the results indicated that miR-491 markedly reduced the luciferase
activity compared with that in the vector with the mutant binding
sequence (Fig. 5B). Furthermore, a
RIP assay was performed for AGO2, and the levels of endogenous
circ-0000212 and miR-491 pulled-down from AGO2-expressing cells was
determined using RT-qPCR analysis. The results demonstrated that
circ-0000212 and miR-491 were highly enriched in the AGO2 pellet
compared with that in the input control (Fig. 5C). As a sponge of miR-491,
circ-0000212 could regulate the targets of miR-491. The cells were
than transfected with either si-circ-0000212 and miR-491 inhibitor
or si-circ-0000212 and miR-491 inhibitor at the same time. Notably,
si-circ-0000212 decreased the expression levels of FOXP4, a target
of miR-491, while inhibition of miR-491 increased the expression
levels of FOXP4 (Figs. 5D, S2B and C). Furthermore, si-circ-0000212
reversed the effects of miR-491 on the regulation of FOXP4
(Figs. 5D, S2B and C). Finally, the ability of
circ-0000212 to modulate cell proliferation in CRC was
investigated. si-circ-0000212 significantly reduced the
proliferative ability of the CRC cells (Figs. 5E, S2B
and C), and consistently, it could also reverse the ability of
the miR-491 inhibitor to increase cell proliferation (Figs. 5E and F, S2B and C). Taken together, circ-0000212
could act as sponge of miR-491.
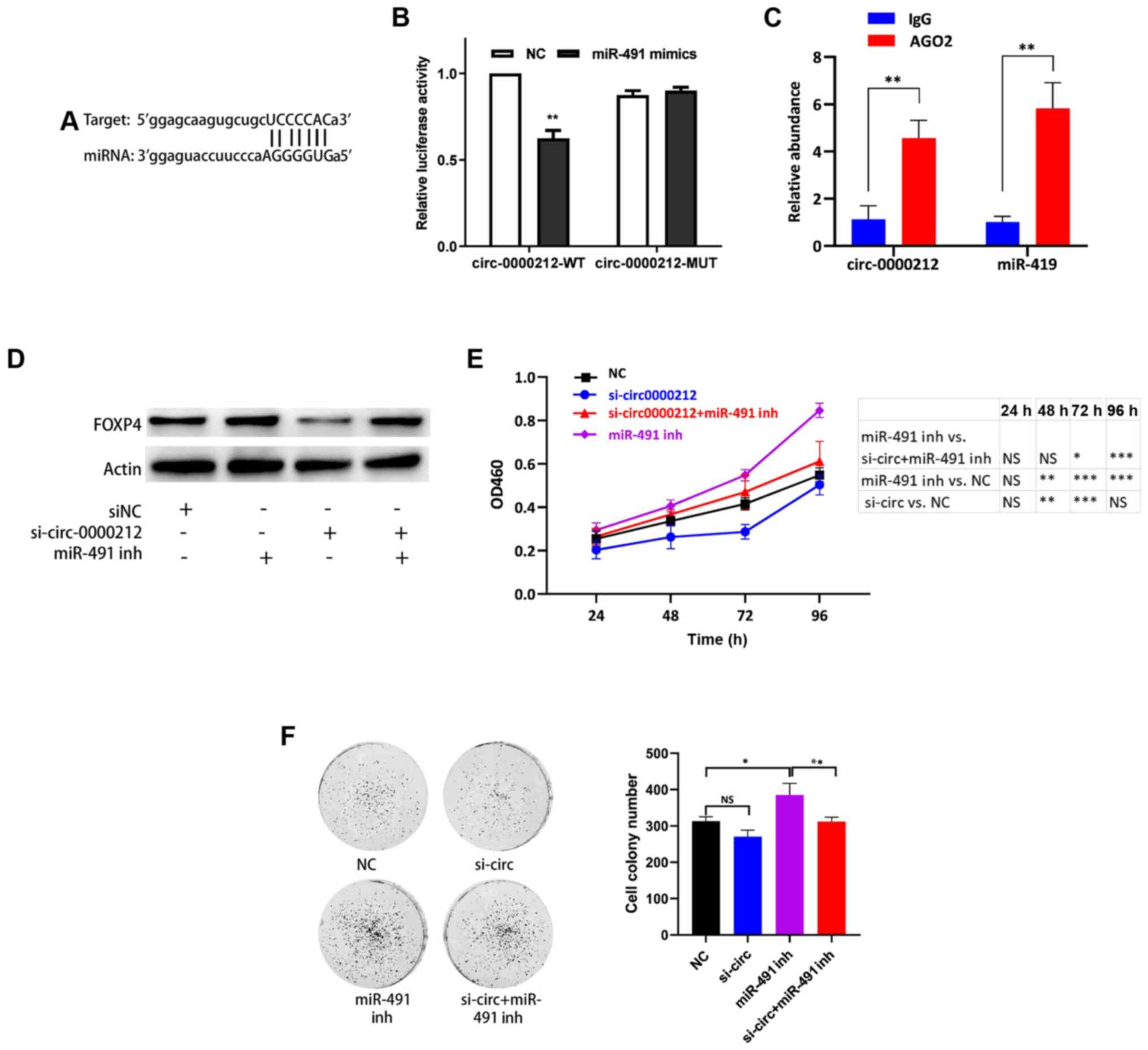 | Figure 5.circ-0000212 acts as a sponge for
miR-491. (A) Binding site between circ-0000212 and miR-491. (B) In
the 293T cells, transfection of the miR-491 mimic decreased
luciferase activity of the vector containing the wild type binding
site of circ-0000212. **P<0.01 vs. NC. (C) AGO2 RNA
immunoprecipitation assay to determine the quantity of circ-0000212
and miR-491. **P<0.01 vs. IgG. (D) Western blot analysis of the
expression levels of FOXP4 after transfection with si-circ-0000212
+ miR-491 inh in SW620 cells. (E) Cell Counting Kit-8 assay of
SW620 cells after transfection with si-circ-0000212 + miR-491 inh.
*P<0.05, **P<0.01, ***P<0.001. (F) Colony formation assay
of SW620 cells after transfection with si-circ-0000212 + miR-491
inh. *P<0.05, **P<0.01, as indicated. Experiments were
repeated three times. miR-491, microRNA-491; miR-491 inh, miR-491
inhibitor; si, small interfering; circ, circular RNA; OD, optical
density; NC, negative control; FOXP4, forkhead box P4. |
Discussion
Previous studies have shown that FOXP4 functions as
a tumor promoter in various types of cancer (22,23),
and is upregulated in cancer tissues (23). In the present study, the expression
levels of FOXP4 were increased in a large CRC cohort based on data
obtained from TCGA. Dysregulated miR-491 expression has been
identified in various types of tumors. In breast cancer, miR-491
was downregulated in cancer tissues and functioned as a tumor
suppressor by targeting TPX2 (24).
In glioma, miR-491 overexpression inhibited cell proliferation via
the Wnt3a/β-catenin pathway as well as invasion-mediated
epithelial-mesenchymal transition (25). miR-491 also suppressed cell
proliferation, motility and invasion in gastric cancer by targeting
HMGA2 (26), or SNAIL and FGFR4
(27). Furthermore, miR-491
inhibited cell proliferation, motility and progression in prostate
cancer (28) and nasopharyngeal
carcinoma (29). Even though
miR-491 has been investigated in CRC (30), there is limited research in this
area. In the present study, there was a difference in the sample
sizes between the normal and cancer tissues (8 vs. 457); however,
the data were normally distributed, and the two groups were of
equal variances. Therefore, the results are reliable. To the best
of our knowledge, for the first time, these results showed that the
miR-491 mimic suppressed cell proliferation by targeting FOXP4.
Based on the association between miR-491 and FOXP4 in CRC tissues,
the association between miR-491 and FOXP4 expression levels with
the clinicopathological characteristics in patients with CRC were
investigated. Both high miR-491 and low FOXP4 expression levels
were associated with pathological and N stages, highlighting its
potential as a diagnostic marker for patients with CRC. The
aforementioned results show that miR-491 may act as a tumor
suppressor in cancer. However, the results suggested that there was
no association between miR-491 and reduced invasion of the CRC
cells, as it was not associated with M stage, which has been shown
in other types of cancer (24).
This might be due to heterogeneity of different types of cancer and
cell lines; therefore, further investigation is required to
determine the role of miR-491 in different types of cancer.
An increasing number of studies has demonstrated
that circRNAs are more than junk or by-products of errors during
the splicing process. circRNAs can functionally regulate
transcriptional activity and also play roles as miRNA sponges and
therefore, modulate oncogenesis and cancer progression (31–34).
In addition, circRNAs may also function as protein scaffolds,
sequester proteins and function co-operatively to achieve
measurable effects (31).
Genome-wide research has discovered a large number
of new circRNAs with different functions in cancer. circRNAs can
act as tumor promoters. circCCDC66 and circHIAT1 increased tumor
proliferation and metastasis in various types of cancer (35,36).
Conversely, other circRNAs have also been shown to inhibit cancer
progression. circREPS2 and circ_0000353 suppresses gastric
progression (37) and metastasis of
non-small cell lung cancer (38),
respectively, via modulation of multiple signaling pathways.
In the present study, circ-0000212 was selected. It
is a circRNA derived from SFMBT2, which was shown to sponge
miR-491. SFMBT2 has been shown to be dysregulated in several types
of cancer and is involved in cell proliferation. It has been shown
that SFMBT2 promoted cell viability through the epigenetic
modulation of HOXB13 gene expression in prostate cancer cells
(39). Furthermore, SFMBT2
positively regulated chondrocyte proliferation via SOX9 (40). Notably, it was discovered that
circ-0000212 was upregulated in CRC tissues compared with that in
adjacent normal tissues. In addition, compared with linear SFMBT2,
circ-0000212 displayed stable expression in CRC cells and was
preferentially distributed in the cytoplasm. Functional experiments
found that circ-0000212 increased cell proliferation by sponging
miR-491 and modulated FOXP4 expression levels. These data suggest
that circ-0000212 may function as a tumor promoter in CRC via
regulation of a miR-491/FOXP4 signaling axis. Notably, the CRC cell
lines used in the present study had different expression levels of
miR-491 and circ-0000212. The HCT116, SW480, HT29, LoVo and SW620
cells are classified as stages IV, II, III, III, and III,
respectively. There was no significant association between the
expression levels and the cell lines characteristics. Thus, other
factors, such as gene mutation and chromosomal stability may affect
the expression levels of miR-491 and circ-0000212.
circRNAs are stable and present different
immunogenicity when synthesized in vitro, due to their
specific loop structure, and therefore could be used as promising
potential biomarkers for cancer diagnosis and prognosis (31,36).
Previously, research has focused on the application of circRNA
technologies to probe and operate intracellular processes (41), manipulate miRNA expression levels
(42), repress innate immune
responses (43), augment immune
functions (44) and serve as
biomarkers (45).
In conclusion, circ-0000212 may sponge miR-491 and
therefore modulate FOXP4 expression in CRC cells. The present study
lays the foundation for future studies, and circ-0000212 may serve
as a novel target for the development of treatments to manage
CRC.
Supplementary Material
Supporting Data
Supporting Data
Supporting Data
Acknowledgements
Not applicable.
Funding
This work was supported by the National Natural
Science Foundation of China (grant no. 8170110698), Heilongjiang
Province Natural Science Foundation (grant no. QC2017095),
Heilongjiang Postdoctoral Fund (grant no. LBH-Z18270) and China
Postdoctoral Science Foundation (grant no. 2019M651319).
Availability of data and materials
TCGA dataset is available from https://www.cancer.gov/about-nci/organization/ccg/research/structural-genomics/tcga.
The other datasets used and/or analyzed during the present study
are available from the corresponding author on reasonable
request.
Authors' contributions
QZ conceived the study. GW provided administrative
support and analyzed data. YT, performed the experiments, collected
the study materials and patient data. HW performed the experiments.
WZ analyzed and interpretated the data. All authors confirm the
authenticity of all the raw data. All authors wrote the manuscript,
and read and approved the final manuscript.
Ethics approval and consent to
participate
The study was approved by the Institutional Review
Board of The Second Affiliated Hospital of Harbin Medical
University (approval no. KY2017-031). All patients consent to
participate in the study.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Siegel RL, Miller KD and Jemal A: Cancer
statistics, 2019. CA Cancer J Clin. 69:7–34. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Adams BD, Parsons C, Walker L, Zhang WC
and Slack FJ: Targeting noncoding RNAs in disease. J Clin Invest.
127:761–771. 2017. View
Article : Google Scholar : PubMed/NCBI
|
|
3
|
Lekka E and Hall J: Noncoding RNAs in
disease. FEBS Lett. 592:2884–2900. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Lorenzi L, Avila Cobos F, Decock A,
Everaert C, Helsmoortel H, Lefever S, Verboom K, Volders PJ,
Speleman F, Vandesompele J and Mestdagh P: Long noncoding RNA
expression profiling in cancer: Challenges and opportunities. Genes
Chromosomes Cancer. 58:191–199. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Garzon R, Marcucci G and Croce CM:
Targeting microRNAs in cancer: Rationale, strategies and
challenges. Nat Rev Drug Discov. 9:775–789. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Corté H, Manceau G, Blons H and
Laurent-Puig P: MicroRNA and colorectal cancer. Dig Liver Dis.
44:195–200. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Yang L, Belaguli N and Berger DH: MicroRNA
and colorectal cancer. World J Surg. 33:638–646. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Acunzo M, Romano G, Wernicke D and Croce
CM: MicroRNA and cancer-a brief overview. Adv Biol Regul. 57:1–9.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Wilusz JE and Sharp PA: Molecular biology.
A circuitous route to noncoding RNA. Science. 340:440–441. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Qu S, Zhong Y, Shang R, Zhang X, Song W,
Kjems J and Li H: The emerging landscape of circular RNA in life
processes. RNA Biol. 14:992–999. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Zhou DN, Ye CS and Deng YF: CircRNAs:
Potency of protein translation and feasibility of novel biomarkers
and therapeutic targets for head and neck cancers. Am J Transl Res.
12:1535–1552. 2020.PubMed/NCBI
|
|
12
|
Liu J, Zhang X, Yan M and Li H: Emerging
role of circular RNAs in cancer. Front Oncol. 10:6632020.
View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Zhong Z, Huang M, Lv M, He Y, Duan C,
Zhang L and Chen J: Circular RNA MYLK as a competing endogenous RNA
promotes bladder cancer progression through modulating VEGFA/VEGFR2
signaling pathway. Cancer Lett. 403:305–317. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Barrett SP and Salzman J: Circular RNAs:
Analysis, expression and potential functions. Development.
143:1838–1847. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Zhang L, Dong X, Yan B, Yu W and Shan L:
CircAGFG1 drives metastasis and stemness in colorectal cancer by
modulating YY1/CTNNB1. Cell Death Dis. 11:5422020. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Zheng Q, Bao C, Guo W, Li S, Chen J, Chen
B, Luo Y, Lyu D, Li Y, Shi G, et al: Circular RNA profiling reveals
an abundant circHIPK3 that regulates cell growth by sponging
multiple miRNAs. Nat Commun. 7:112152016. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Colaprico A, Silva TC, Olsen C, Garofano
L, Cava C, Garolini D, Sabedot TS, Malta TM, Pagnotta SM,
Castiglioni I, et al: TCGAbiolinks: An R/bioconductor package for
integrative analysis of TCGA data. Nucleic Acids Res. 44:e712016.
View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Zellweger T, Miyake H, Cooper S, Chi K,
Conklin BS, Monia BP and Gleave ME: Antitumor activity of antisense
clusterin oligonucleotides is improved in vitro and in vivo by
incorporation of 2′-O-(2-methoxy)ethyl chemistry. J Pharmacol Exp
Ther. 298:934–940. 2001.PubMed/NCBI
|
|
19
|
Zhao ZJ and Shen J: Circular RNA
participates in the carcinogenesis and the malignant behavior of
cancer. RNA Biol. 14:514–521. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Gwak J, Jeong H, Lee K, Shin JY, Sim T, Na
J, Kim J and Ju BG: SFMBT2-mediated infiltration of preadipocytes
and TAMs in prostate cancer. Cancers (Basel). 12:27182020.
View Article : Google Scholar
|
|
21
|
Gwak J, Shin JY, Lee K, Hong SK, Oh S, Goh
SH, Kim WS and Ju BG: SFMBT2 (Scm-like with four mbt domains 2)
negatively regulates cell migration and invasion in prostate cancer
cells. Oncotarget. 7:48250–48264. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Ma T and Zhang J: Upregulation of FOXP4 in
breast cancer promotes migration and invasion through facilitating
EMT. Cancer Manag Res. 11:2783–2793. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Zhang G and Zhang G: Upregulation of FoxP4
in HCC promotes migration and invasion through regulation of EMT.
Oncol Lett. 17:3944–3951. 2019.PubMed/NCBI
|
|
24
|
Tan GZ, Li M, Tan X, Shi ML and Mou K:
miR-491 suppresses migration and invasion via directly targeting
TPX2 in breast cancer. Eur Rev Med Pharmacol Sci. 23:9996–10004.
2019.PubMed/NCBI
|
|
25
|
Meng Y, Shang FR and Zhu YL: miR-491
functions as a tumor suppressor through Wnt3a/β-catenin signaling
in the development of glioma. Eur Rev Med Pharmacol Sci.
23:10899–10907. 2019.PubMed/NCBI
|
|
26
|
Liu Z, Lü Y, Jiang Q, Yang Y, Dang C and
Sun R: miR-491 inhibits BGC-823 cell migration via targeting HMGA2.
Int J Biol Markers. 34:364–372. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Yu T, Wang LN, Li W, Zuo QF, Li MM, Zou QM
and Xiao B: Downregulation of miR-491-5p promotes gastric cancer
metastasis by regulating SNAIL and FGFR4. Cancer Sci.
109:1393–1403. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Xu Y, Hou R, Lu Q, Zhang Y, Chen L, Zheng
Y and Hu B: miR-491-5p negatively regulates cell proliferation and
motility by targeting PDGFRA in prostate cancer. Am J Cancer Res.
7:2545–2553. 2017.PubMed/NCBI
|
|
29
|
Cheng Q, Xu X, Jiang H, Xu L and Li Q:
Knockdown of long non-coding RNA XIST suppresses nasopharyngeal
carcinoma progression by activating miR-491-5p. J Cell Biochem.
119:3936–3944. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Hanisch C, Sharbati J, Kutz-Lohroff B,
Huber O, Einspanier R and Sharbati S: TFF3-dependent resistance of
human colorectal adenocarcinoma cells HT-29/B6 to apoptosis is
mediated by miR-491-5p regulation of lncRNA PRINS. Cell Death
Discov. 3:161062017. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Chen LL: The expanding regulatory
mechanisms and cellular functions of circular RNAs. Nat Rev Mol
Cell Biol. 21:475–490. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Zhou ZB, Huang GX, Fu Q, Han B, Lu JJ,
Chen AM and Zhu L: circRNA.33186 contributes to the pathogenesis of
osteoarthritis by sponging miR-127-5p. Mol Ther. 27:531–541. 2019.
View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Liu Z, Yu Y, Huang Z, Kong Y, Hu X, Xiao
W, Quan J and Fan X: CircRNA-5692 inhibits the progression of
hepatocellular carcinoma by sponging miR-328-5p to enhance DAB2IP
expression. Cell Death Dis. 10:9002019. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Meng S, Zhou H, Feng Z, Xu Z, Tang Y, Li P
and Wu M: CircRNA: Functions and properties of a novel potential
biomarker for cancer. Mol Cancer. 16:942017. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Hsiao KY, Lin YC, Gupta SK, Chang N, Yen
L, Sun HS and Tsai SJ: Noncoding effects of circular RNA CCDC66
promote colon cancer growth and metastasis. Cancer Res.
77:2339–2350. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Wang K, Sun Y, Tao W, Fei X and Chang C:
Androgen receptor (AR) promotes clear cell renal cell carcinoma
(ccRCC) migration and invasion via altering the
circHIAT1miR-195-5p/29a-3p/29c-3p/CDC42 signals. Cancer Lett.
394:1–12. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Guo X, Dai X, Liu J, Cheng A, Qin C and
Wang Z: Circular RNA circREPS2 Acts as a sponge of miR-558 to
suppress gastric cancer progression by regulating RUNX3/β-catenin
signaling. Mol Ther Nucleic Acids. 21:577–591. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Zhao WX, Tang YL, Wang WH and Bao MW:
Up-regulation of circ_0000353 impedes the proliferation and
metastasis of non-small cell lung cancer cells via adsorbing
miR-411-5p and increasing forkhead box O1. Cancer Biomark.
29:25–37. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Lee K, Na W, Maeng JH, Wu H and Ju BG:
Regulation of DU145 prostate cancer cell growth by Scm-like with
four mbt domains 2. J Biosci. 38:105–112. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Hussain S, Sun M, Guo Y, Mushtaq N, Zhao
Y, Yuan Y, Hussain N, Osoro E, Suleiman A, Sadiq M, et al: SFMBT2
positively regulates SOX9 and chondrocyte proliferation. Int J Mol
Med. 42:3503–3512. 2018.PubMed/NCBI
|
|
41
|
Litke JL and Jaffrey SR: Highly efficient
expression of circular RNA aptamers in cells using autocatalytic
transcripts. Nat Biotechnol. 37:667–675. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Jost I, Shalamova LA, Gerresheim GK,
Niepmann M, Bindereif A and Rossbach O: Functional sequestration of
microRNA-122 from hepatitis C virus by circular RNA sponges. RNA
Biol. 15:1032–1039. 2018.PubMed/NCBI
|
|
43
|
Liu CX, Li X, Nan F, Jiang S, Gao X, Guo
SK, Xue W, Cui Y, Dong K, Ding H, et al: Structure and degradation
of circular RNAs regulate PKR activation in innate immunity. Cell.
177:865–880.e21. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Chen YG, Kim MV, Chen X, Batista PJ,
Aoyama S, Wilusz JE, Iwasaki A and Chang HY: Sensing self and
foreign circular RNAs by intron identity. Mol Cell. 67:228–238.e5.
2017. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Bahn JH, Zhang Q, Li F, Chan TM, Lin X,
Kim Y, Wong DT and Xiao X: The landscape of microRNA,
Piwi-interacting RNA, and circular RNA in human saliva. Clin Chem.
61:221–230. 2015. View Article : Google Scholar : PubMed/NCBI
|