Introduction
Liver cancer is the third leading malignant tumor
worldwide, with a mortality rate of 8.2% and ~840,000 new cases
each year (1). As the most common
form of liver cancer, hepatocellular carcinoma (HCC) accounts for
90% of all cases of primary liver cancer (2). HCC is considered as a severe malignant
tumor in China due to 55% morbidity rate (3). Moreover, HCC morbidity has rapidly
increased on a global scale in the last five decades, partly due to
hepatitis B or C virus infection and cirrhosis associated with poor
lifestyle (4–6). At present, the best treatment
strategies available for HCC include surgical resection and liver
transplantation, but the survival rate (6.9%) of patients with HCC
is poor due to its high recurrence and metastasis (7–9). The
molecular mechanism underlying HCC carcinogenesis is not completely
understood. Therefore, improving the current understanding of the
molecular mechanism underlying HCC may aid with the development of
novel therapeutic strategies to improve the prognosis of the
disease.
Located on chromosome 7p14.1, secreted
frizzled-related protein 4 (sFRP-4) consists of a total of six
exons (10). sFRP-4 belongs to the
SFRP family and contains a cysteine-rich domain that is homologous
to the assumed Wnt-binding site of the frizzled proteins (11). Moreover, sFRP-4 can regulate the Wnt
signaling pathway by directly binding to Wnt, thus preventing Wnt
from binding to its receptor (12,13).
Increasing evidence has suggested that sFRP-4 could inhibit the
canonical Wnt signaling pathway, form silencing complexes and
suppress human cancer, including gastric (14), ovarian (15) and cervical cancer (16), as well as mesothelioma (17) and cemento ossifying fibroma
(18). Several studies also
reported that sFRP-4 protein was highly expressed in other types of
cancer, including prostate cancer (19), pancreatic ductal adenocarcinoma
(20), colon carcinoma (21) and colorectal carcinoma (22). Therefore, the aforementioned studies
suggested that sFRP-4 was differentially expressed in different
types of cancer. Our preliminary work demonstrated that sFRP-4
expression levels were upregulated in the serum obtained from
patients with HCC. In addition, the combined use of sFRP-4 and
α-fetoprotein (AFP; a standard serum marker of HCC) improved the
accuracy of HCC diagnosis (23).
However, the functions and mechanism underlying sFRP-4 in HCC
development were not investigated in our previous study. Therefore,
the present study explored the exact functions and mechanism
underlying sFRP-4 in HCC using HCC clinical tissues and cell
lines.
Furthermore, the Wnt signaling pathway can be
classified into two major types: Classical (β-catenin-dependent)
and non-classical (β-catenin-independent) (24,25).
As a key component of the classical pathway, β-catenin is strictly
regulated by a variety of protein complexes, including GSK-3β
(26). In the absence of Wnt
ligands, the Wnt/β-catenin signaling pathway is inactive and
β-catenin is degraded after sustained phosphorylation to maintain
low levels in the cytoplasm. Following Wnt induction, the
Wnt/β-catenin signaling pathway is activated and the degradation
process of β-catenin phosphorylation is inhibited, resulting in
intracellular accumulation of β-catenin (27,28).
As a key member of the Wnt/β-catenin signaling pathway, GSK-3β can
participate in the degradation of β-catenin, but when the
Wnt/β-catenin signaling pathway is activated, GSK-3β remains in an
inhibitory state (29). Abnormal
activation of the Wnt/β-catenin signaling pathway results in the
carcinogenesis and development of various types of cancer,
including gastric (30), colorectal
(31), esophageal (32) and pancreatic cancer (33), as well as HCC (34). Furthermore, it has been reported
that 30% of patients with HCC exhibited excessive activation of the
Wnt/β-catenin signaling pathway (35). Therefore, exploring the molecular
mechanism underlying the Wnt/β-catenin signaling pathway in HCC is
important for the identification of novel effective therapeutic
targets.
After performing preliminary research on the
clinical significance and mechanism underlying sFRP-4 in HCC, as
well as the regulatory mechanism underlying the Wnt/β-catenin
signaling pathway in HCC, it was hypothesized that sFRP-4 could
restrain HCC progression, potentially via suppression of the
Wnt/β-catenin signaling pathway. To verify the hypothesis, sFRP-4
expression levels in HCC tissues and cells were assessed.
Therefore, the results of the present study may aid with the
identification of novel therapeutic targets for HCC.
Materials and methods
Clinical tissue samples
Tissue samples (47 paired HCC and adjacent
non-cancerous tissues) were collected from patients (age range,
41–76 years) who were diagnosed with HCC at the General Hospital of
the Central Theater Command of the People's Liberation Army (Wuhan,
China) between June 2015 and December 2019, and 59.6% patients were
>60 years old. All patients had not received treatment with
anticancer drugs before surgical resection, and incomplete data
were excluded. After performing surgical resection, samples of HCC
tissue and samples of adjacent non-cancerous tissue (>3 cm away
from the tumor site; cirrhosis tissue was excluded) were
immediately labeled and frozen until further analysis. All cases
were diagnosed histopathologically. The clinical characteristics of
the patients are presented in Table
I. The present study was approved by the Ethics Committee of
the General Hospital of the Central Theater Command of the People's
Liberation Army (approval no. [2020]024-2). All patients provided
written informed consent.
 | Table I.Association between sFRP-4 expression
and the clinical characteristics of patients with hepatocellular
carcinoma. |
Table I.
Association between sFRP-4 expression
and the clinical characteristics of patients with hepatocellular
carcinoma.
|
|
| sFRP-4
expression |
|
|---|
|
|
|
|
|
|---|
| Characteristic | n (%) | High (n=20) | Low (n=27) | P-value |
|---|
| Age (years) |
|
|
| 0.959 |
|
>60 | 19 (40.4%) | 8 (40.0%) | 11 (40.7%) |
|
|
≤60 | 28 (59.6%) | 12 (60.0%) | 16 (59.3%) |
|
| Sex |
|
|
| 0.824 |
|
Male | 36 (76.6%) | 15 (75.0%) | 21 (77.8%) |
|
|
Female | 11 (23.4%) | 5 (25.0%) | 6 (22.2%) |
|
| Tumor diameter
(cm) |
|
|
| 0.057 |
|
>5 | 39 (83.0%) | 14 (70.0%) | 25 (92.6%) |
|
| ≤5 | 8 (17.0%) | 6 (30.0%) | 2 (7.4%) |
|
|
Differentiation |
|
|
| 0.133 |
|
Well | 7 (14.9%) | 5 (25.0%) | 2 (7.4%) |
|
|
Moderate | 25 (53.2%) | 11 (55.0%) | 14 (51.9%) |
|
|
Poor | 15 (31.9%) | 4 (20.0%) | 11 (40.7%) |
|
| TNM stage |
|
|
| 0.002a |
| I | 5 (10.6%) | 4 (20.0%) | 1 (3.7%) |
|
| II | 18 (38.3%) | 12 (60.0%) | 6 (22.2%) |
|
|
III | 22 (46.8%) | 4 (20.0%) | 18 (66.7%) |
|
| IV | 2 (4.3%) | 0 (0.0%) | 2 (7.4%) |
|
| Metastasis |
|
|
|
|
|
Positive | 26 (55.3%) | 6 (30.0%) | 20 (74.1%) | 0.003a |
|
Negative | 21 (44.7%) | 14 (70.0%) | 7 (25.9%) |
|
| AFP (IU) |
|
|
|
|
|
≤400 | 20 (42.6%) | 13 (65.0%) | 7 (25.9%) | 0.007a |
|
>400 | 27 (57.4%) | 7 (35.0%) | 20 (74.1%) |
|
Cell lines and cell culture
The cell lines used in the present study were
purchased from BeNa Culture Collection (Beijing Beina Chunglian
Biotechnology Research Institute). The following HCC cell lines
were used: HCCLM3 (cat. no. BNCC338460), Hep3B (cat. no.
BNCC337952) and Huh7 (cat. no. BNCC337690). The MIHA immortalized
normal human liver cell line (cat. no. BNCC340123) was also used.
MIHA cells were cultured in RPMI-1640 (Gibco; Thermo Fisher
Scientific, Inc.) supplemented with 10% FBS (Gibco; Thermo Fisher
Scientific, Inc.) and 100 U/ml streptomycin/penicillin (Invitrogen;
Thermo Fisher Scientific, Inc.). HCCLM3, Hep3B and Huh76 cells were
cultured in DMEM-H (Gibco; Thermo Fisher Scientific, Inc.)
supplemented with 10% FBS and 100 U/ml streptomycin/penicillin. All
cells were cultured at 37°C with 5% CO2.
Cell transfection and IM-12
treatment
sFRP-4 small interfering (si)RNA vector (si-sFRP-4;
forward: 5′-GGAGGAUCACAGAAAUGUAGA-3′ and reverse:
5′-UACAUUUCUGUGAUCCUCCUA-3′), sFRP-4 overexpression vector (OE;
sFRP-4-OE; forward: 5′-ATGCAAGCTTTTCCTCTCCATCCTAGTGGCG-3′, reverse:
5′-TCACGGGATCCTTCTTGGGACTGGCTGGTTT-3′), empty vector (pcDNA3.1) and
non-targeting siRNA (si-NC) were purchased from Guangzhou Funeng
Gene Co., Ltd. Cells in the blank group were cultured under normal
conditions. A total of 5×104 HCCLM3 and Huh7 cells were
transfected with 100 nM si-sFRP-4, 1 µg/ml sFRP-4-OE or negative
control (NC) vectors containing 1 µg/ml empty vector and 100 nM
si-NC using Lipofectamine RNAiMax (Thermo Fisher Scientific, Inc.)
at room temperature according to the manufacturer's protocol. After
48 h transfection, the transfection efficiency of si-sFRP-4 and
sFRP-4-OE was assessed via reverse transcription-quantitative PCR
(RT-qPCR). At 12 h post-transfection, cells were treated with 1 µM
IM-12 (cat. no. SML0084; Sigma-Aldrich; Merck KGaA) for 36 h at
37°C, and the control group was treated with 1% DMSO.
RT-qPCR
Total RNA was isolated from frozen tissues or
cultured cells using TRIzol® (Invitrogen; Thermo Fisher
Scientific, Inc.). RNA concentration was measured using a
spectrophotometer. Total RNA was reverse transcribed into cDNA
using SuperScript IV VILO Master Mix (cat. no. 11756050;
Invitrogen; Thermo Fisher Scientific, Inc.) at 50°C for 10 min and
85°C for 5 min. Subsequently, RT-qPCR was performed using PowerUp
SYBR™ Green Master Mix (cat. no. A25779; Applied Biosystems; Thermo
Fisher Scientific, Inc.) and an ABI 7500 Real Time PCR system
(Applied Biosystems; Thermo Fisher Scientific, Inc.). The
thermocycling conditions were as follows: Initial denaturation at
95°C for 4 min, followed by 40 cycles of denaturation at 95°C for
15 sec and annealing at 60°C for 60 sec. The following primers were
used for qPCR: GAPDH forward, 5′-CACCGTAGCCTTCCGAGTA-3′ and
reverse, 5′-GCCCTTGATGAGCTGTTGA-3′; and sFRP-4 forward,
5′-CAGAGGAGTGGCTGCAATGA-3′ and reverse, 5′-CTTGATGGGGCAGGATGTGT-3′.
sFRP-4 mRNA expression levels were quantified using the
2−∆∆Cq method (36) and
normalized to the internal reference gene GAPDH.
Immunohistochemistry (IHC)
analysis
sFRP-4 expression in 47 paired HCC and adjacent
non-cancerous tissues was assessed by performing IHC staining.
Tissues was imbedded in paraffin after 12 h of 10% neutral formalin
fixation at room temperature. Then, 5-µm thickness tissue sections
were gradually dewaxed and hydrated with xylene and descending
ethanol. Sections were then treated with 10 mM citrate buffer (cat.
no. AP-9003-125, Lab Vision Corp.) at 95°C for 5 min to repair
antigens via heat treatment. Then, slides washed in deionized water
three times for 2 min. Sections were incubated in 3% hydrogen
peroxide for 10 min at room temperature to inhibit endogenous
peroxidase activity. Subsequently, non-specific sites were blocked
with 5% goat serum (Chemicon International Inc.) for 30 min at room
temperature. Tissue sections were incubated with an anti-sFRP-4
primary antibody (1:500; cat. no. ab217180; Abcam) at 4°C
overnight. Following primary incubation, tissue sections were
incubated with a corresponding HRP-conjugated secondary antibody
(1:1,000; cat. no. ab6721; Abcam) at 37°C for 1 h. Protein
expression was visualized using the Pierce DAB Substrate kit (cat.
no. 34002; Thermo Fisher Scientific, Inc.). Finally, tissue
sections were counterstained with hematoxylin for 2 min at room
temperature. Stained sections were observed using an objective lens
of a computer-aided light microscope imager (magnification, ×200;
Openlab; PerkinElmer, Inc.). Images were scanned using ImageScope
Version 10 software (Aperio Technologies, Inc.).
Cell viability assay
HCCLM3 and Huh7 cell viability was assessed using
the Cell Counting Kit-8 (CCK-8) assay (cat no. CK04-13; Dojindo
Molecular Technologies, Inc.) according to the manufacturer's
protocol. After transfection for 24 h, HCCLM3 and Huh7 cells were
seeded (5×103 cells/well) into 96-well plates. Following
incubation for 0, 24, 48 or 72 h at 37°C with 5% CO2, 10
µl CCK-8 solution was added to each well and incubated for 2 h.
Subsequently, cell viability was assessed by measuring absorbance
at a wavelength of 450 nm using a microplate reader.
Cell proliferation assay
After 48-h transfection, HCCLM3 and Huh7 cell
proliferation was assessed by performing the BrdU ELISA assay using
the CytoSelect™ BrdU Cell Proliferation ELISA kit (cat. no.
CBA-251, Cell Biolabs, Inc.) according to the manufacturer's
protocol. HCCLM3 and Huh7 cells were seeded (1×104
cells/well) into 96-well plates for 48 h. Subsequently, 10 µl BrdU
solution (X10) was added to each well for 4 h at 37°C. Following
washing with PBS, cells were fixed and cellular DNA was denatured
by incubation with 100 µl Fix/Denature Solution for 30 min at room
temperature. Cells were then incubated with 100 µl anti-BrdU
antibody for 1 h at room temperature, followed by incubation with
100 µl secondary antibody for 1 h at room temperature. Cell
proliferation was assessed by measuring absorbance at a wavelength
of 450 nm using a microplate reader.
Cell apoptosis assay
HCCLM3 and Huh7 cell apoptosis was assessed via flow
cytometry using the Annexin V-FITC Apoptosis Staining/Detection kit
(cat. no. ab14085; Abcam) after transfection for 48 h. HCCLM3 and
Huh7 cells were seeded (5×104 cells/well) into a 6-well
plate and cultured at 37°C with 5% CO2. At 70–80%
confluence, cells were incubated in 500 µl binding buffer
containing 5 µl PI and 5 µl Annexin V-FITC at room temperature for
10 min in the dark. Apoptotic cells were analyzed via flow
cytometry on a FACSCalibur Flow Cytometer (BD Biosciences) using
the CellQuest Pro software (version 5.1; Becton, Dickinson and
Company) and the rate of apoptosis was calculated as the sum of
late apoptosis (Q1-UR) and early apoptosis (Q1-LR).
Western blotting
After 72-h transfection, total protein was isolated
from HCCLM3 and Huh7 cells using RIPA lysis buffer (Thermo Fisher
Scientific, Inc.) and the protein supernatant was collected after
centrifugation at 12,000 × g for 15 min at 4°C. Total protein
concentration was quantified using the BCA kit (Thermo Fisher
Scientific, Inc.). Then, 30 µg proteins per lane were separated via
8% SDS-PAGE and transferred to PVDF membranes, which were blocked
with 5% skimmed milk at room temperature for 1 h. Subsequently, the
membranes were incubated overnight at 4°C with the following
specific primary antibodies: Anti-GSK-3β (48 kDa; 1:1,000; cat. no.
ab131356; Abcam), anti-β-catenin (94 kDa; 1:4,000; cat. no.
ab16051; Abcam), anti-sFRP-4 (40 kDa; 1:3,000; cat. no. ab154167;
Abcam) and anti-GAPDH (36 kDa; 1:1,000; cat. no. ab8245; Abcam).
Following primary incubation, the membranes were incubated with the
corresponding HRP-conjugated secondary antibodies (1:5,000;
anti-mouse IgG, cat. no. ab197767 and anti-rabbit IgG, cat. no.
ab6721; Abcam) for 1 h at room temperature. Protein bands were
visualized using Pierce ECL Western Blotting Substrate (Thermo
Fisher Scientific, Inc.) and the iBrightCL1000 imaging system
(Thermo Fisher Scientific, Inc.). Protein expression was
semi-quantified using ImageJ software (version 1.48; National
Institutes of Health) with GAPDH as the loading control.
Statistical analysis
Statistical analyses were performed using GraphPad
Prism software (version 8.0.1; GraphPad Software, Inc.). Data are
presented as the mean ± SD from three independent experiments. The
paired Student's t test was used to analyze comparisons between two
groups. One way ANOVA followed by Bonferroni's post hoc test was
used to analyze comparisons among multiple groups. The
χ2 or Fisher's exact test was used to compare clinical
characteristics between patients with HCC with lower sFRP-4
expression and patients with HCC with higher sFRP-4 expression.
P<0.05 was considered to indicate a statistically significant
difference.
Results
sFRP-4 expression is decreased in
HCC
To identify the mechanism underlying sFRP-4 in HCC,
the expression of sFRP-4 in 47 paired HCC and adjacent
non-cancerous tissue samples was analyzed via RT-qPCR. sFRP-4
expression was significantly decreased by ~51% in HCC tissues
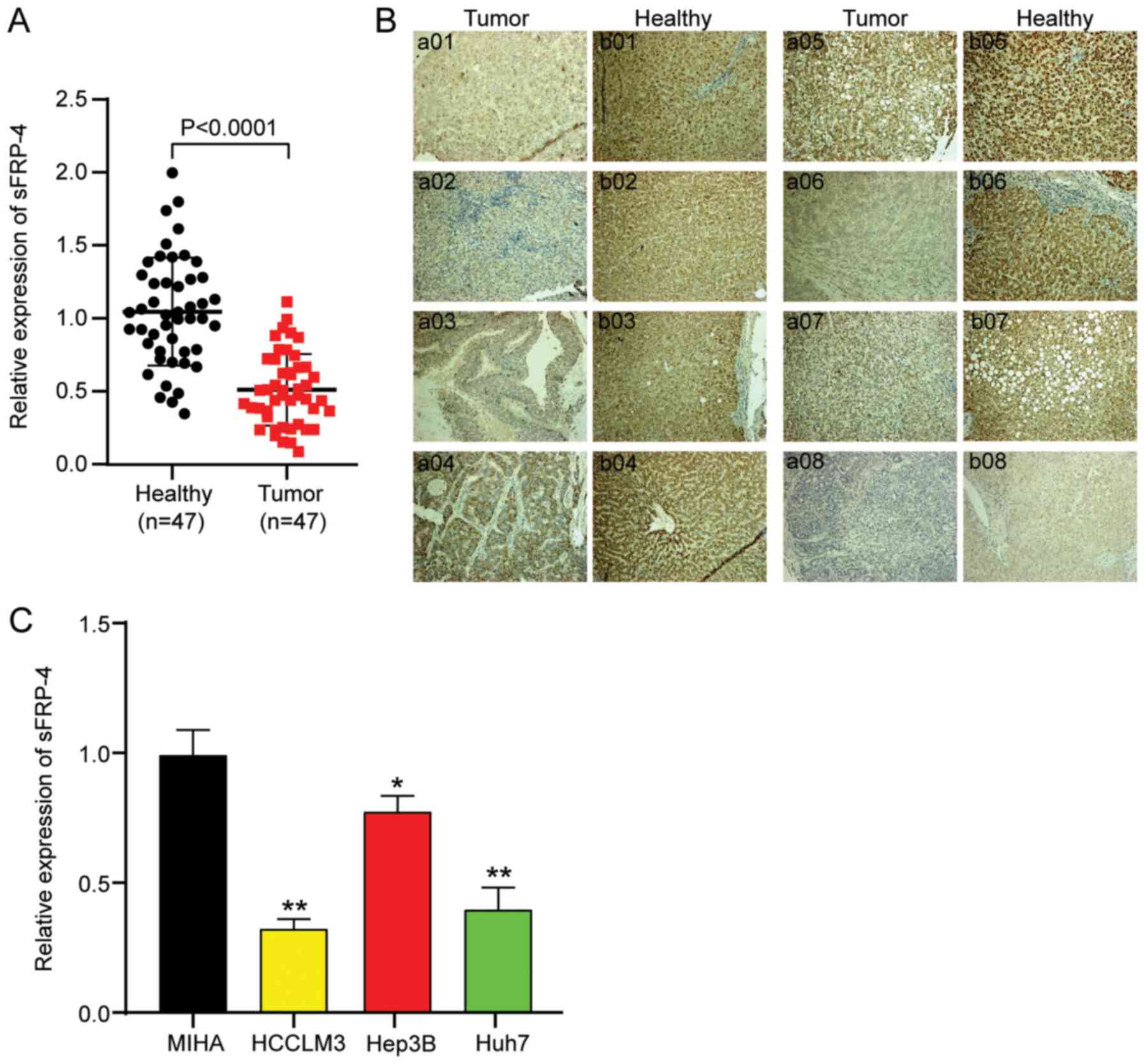
compared with adjacent non-cancerous tissues (Fig. 1A). Subsequently, IHC analysis was
performed to assess sFRP-4 expression in HCC tissues, and images
from eight randomly selected paired HCC and adjacent healthy
tissues are presented in Fig. 1B.
sFRP-4 expression in tumor tissues was notably lower compared with
adjacent healthy tissues. According to the mean value of sFRP4
(0.5) in HCC tumor tissues, as determined via RT-qPCR, patients
with HCC were divided into two groups: sFRP-4 low expression (≤0.5)
and sFRP-4 high expression (>0.5). The results demonstrated that
the tissue expression level of sFRP-4 was significantly negatively
associated with TNM stage, metastasis and AFP level in patients
with HCC (Table I). Moreover, the
expression levels of sFRP-4 in a normal liver cell line (MIHA) and
three HCC cell lines (including HCCLM3, Hep3B and Huh7) were
assessed. The results in HCC cell lines were consistent with HCC
tissues. Compared with the MIHA cell line, the expression of sFRP-4
in HCC cell lines was significantly decreased. Among the HCC cell
lines, sFRP-4 expression was downregulated to the lowest levels in
HCCLM3 (67%) and Huh7 (60%) cells compared with MIHA cells
(Fig. 1C). Therefore, HCCLM3 and
Huh7 cell lines were selected for subsequent experiments.
Collectively, the aforementioned results indicated that sFRP-4
expression was downregulated in HCC, which might exert
tumor-suppressive functions.
sFRP-4 restrains HCC cell viability
and proliferation, but accelerates HCC cell apoptosis
To investigate the influence of sFRP-4 on the
function of HCC cells, HCCLM3 and Huh7 cells were transfected with
sFRP-4-OE or si-sFRP-4. The transfection efficiencies of sFRP-4-OE
and si-sFRP-4 are presented in Fig.
S1. The RT-qPCR results demonstrated that sFRP-4-OE
successfully increased sFRP-4 expression to a level ~3 times higher
compared with the control group (Fig.
2A). By contrast, si-sFRP-4 significantly reduced the
expression of sFRP-4 by ~69% compared with the control group.
Subsequently, CCK-8, BrdU ELISA and flow cytometry assays were
performed to evaluate the effect of sFRP-4 on HCC viability,
proliferation and apoptosis, respectively. The CCK-8 assay results
indicated that HCCLM3 and Huh7 cell viability was significantly
decreased by sFRP-4-OE compared with the control group (Fig. 2B). However, HCCLM3 and Huh7 cell
viability was significantly increased by si-sFRP-4 compared with
the control group at 48 and 72 h. At 48 h post-transfection,
alterations in cell viability were more pronounced compared with
the control group (Fig. 2B), thus
the BrdU ELISA assay was performed to assess HCCLM3 and Huh7 cell
proliferation at 48 h post-transfection (Fig. 2C). Following transfection with
sFRP-4-OE, the absorbance of HCCLM3 and Huh7 cells at 450 nm was
reduced by ~34% compared with the control group. Moreover, sFRP-4
knockdown significantly increased the absorbance of HCCLM3 and Huh7
cells at 450 nm by ~58% compared with the control group. The
results demonstrated that sFRP-4 displayed an inhibitory effect on
HCC cell proliferation. Finally, flow cytometry was performed to
assess cell apoptosis. Compared with the control group, HCCLM3 and
Huh7 cell apoptosis was significantly increased by sFRP-4
overexpression, resulting in an apoptosis rate of 20.19%±1.39 and
23.84%±1.83, respectively (Fig. 2D and
E). However, compared with the control group, HCCLM3 and Huh7
cell apoptosis was significantly decreased by sFRP-4 knockdown,
resulting in an apoptosis rate of 1.32%±0.02 and 1.95%±0.12,
respectively. In summary, the aforementioned results indicated that
compared with the control group, sFRP-4 overexpression decreased
HCC cell viability and proliferation, and accelerated HCC cell
apoptosis, whereas sFRP-4 knockdown displayed the opposite
effect.
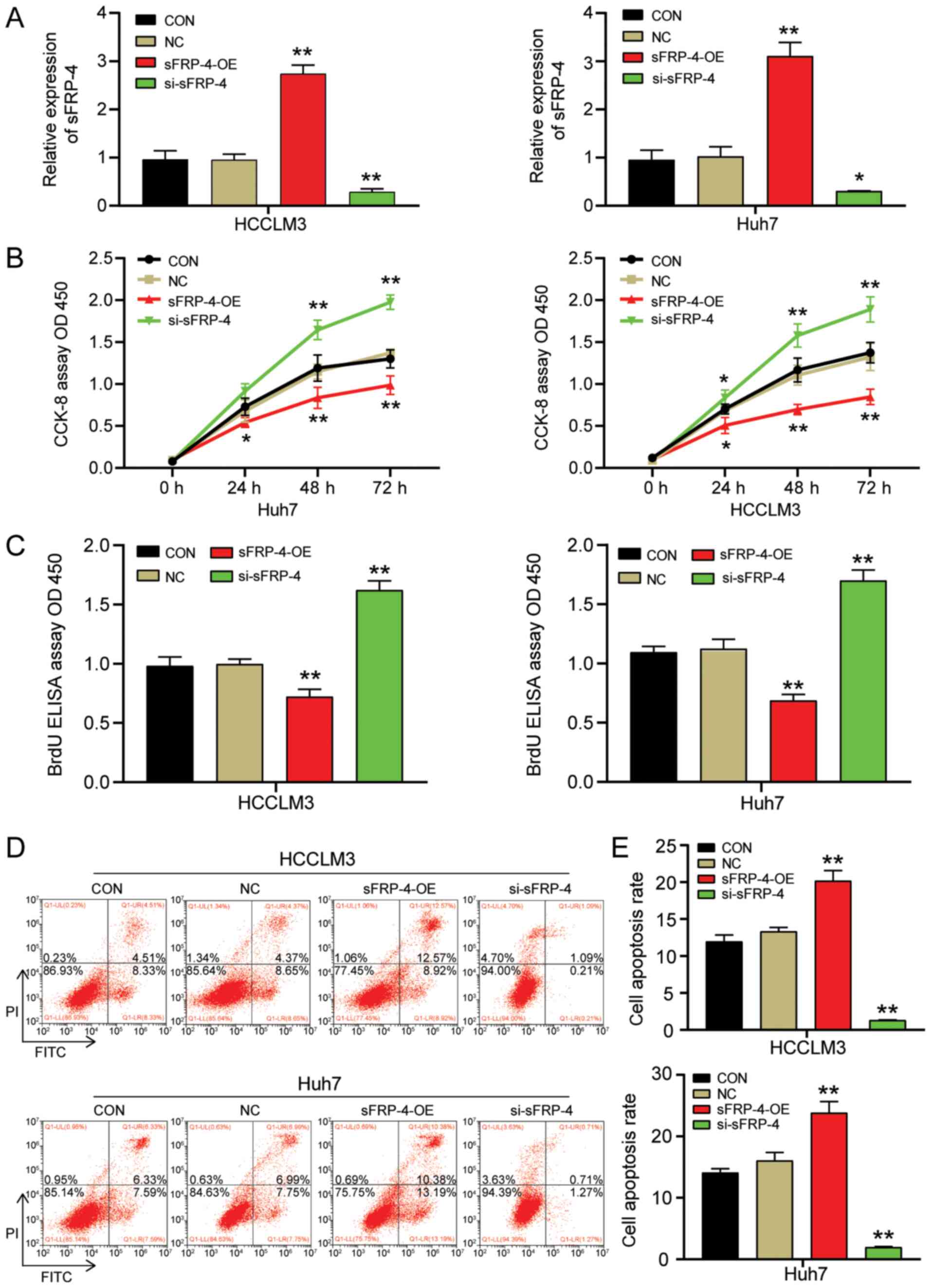 | Figure 2.sFRP-4 inhibits HCC cell viability
and proliferation, and promotes HCC cell apoptosis. (A)
Transfection efficiency of sFRP-4-OE and si-sFRP-4 in HCCLM3 and
Huh7 cells. (B) Effect of sFRP-4 on HCCLM3 and Huh7 cell viability,
as evaluated by performing CCK-8 assays. (C) Effect of sFRP-4 on
HCCLM3 and Huh7 cell proliferation, as evaluated by performing BrdU
ELISA assays. Effect of sFRP-4 on HCCLM3 and Huh7 cell apoptosis
was (D) evaluated via flow cytometry and (E) quantified. Data were
analyzed using one-way ANOVA followed by Bonferroni's post hoc
test. Data are presented as the mean ± SD from three independent
experiments. *P<0.05 and **P<0.001 vs. CON. sFRP-4, secreted
frizzled-related protein 4; HCC, hepatocellular carcinoma; OE,
overexpression; si, small interfering RNA; CCK-8, Cell Counting
Kit-8; CON, blank control; NC, negative control (empty vectors and
si-NC); OD, optical density. |
sFRP-4 inhibits the Wnt/β-catenin
signaling pathway in HCC
The effect of sFRP-4 on HCC carcinogenesis might
alter the associated signaling pathways. The Wnt/β-catenin
signaling pathway is one of the most classic carcinogenic signaling
pathways (37). To explore whether
sFRP-4 downregulation in HCC was mediated via regulation of the
Wnt/β-catenin signaling pathway, the expression levels of key
factors (GSK-3β and β-catenin) in the Wnt/β-catenin signaling
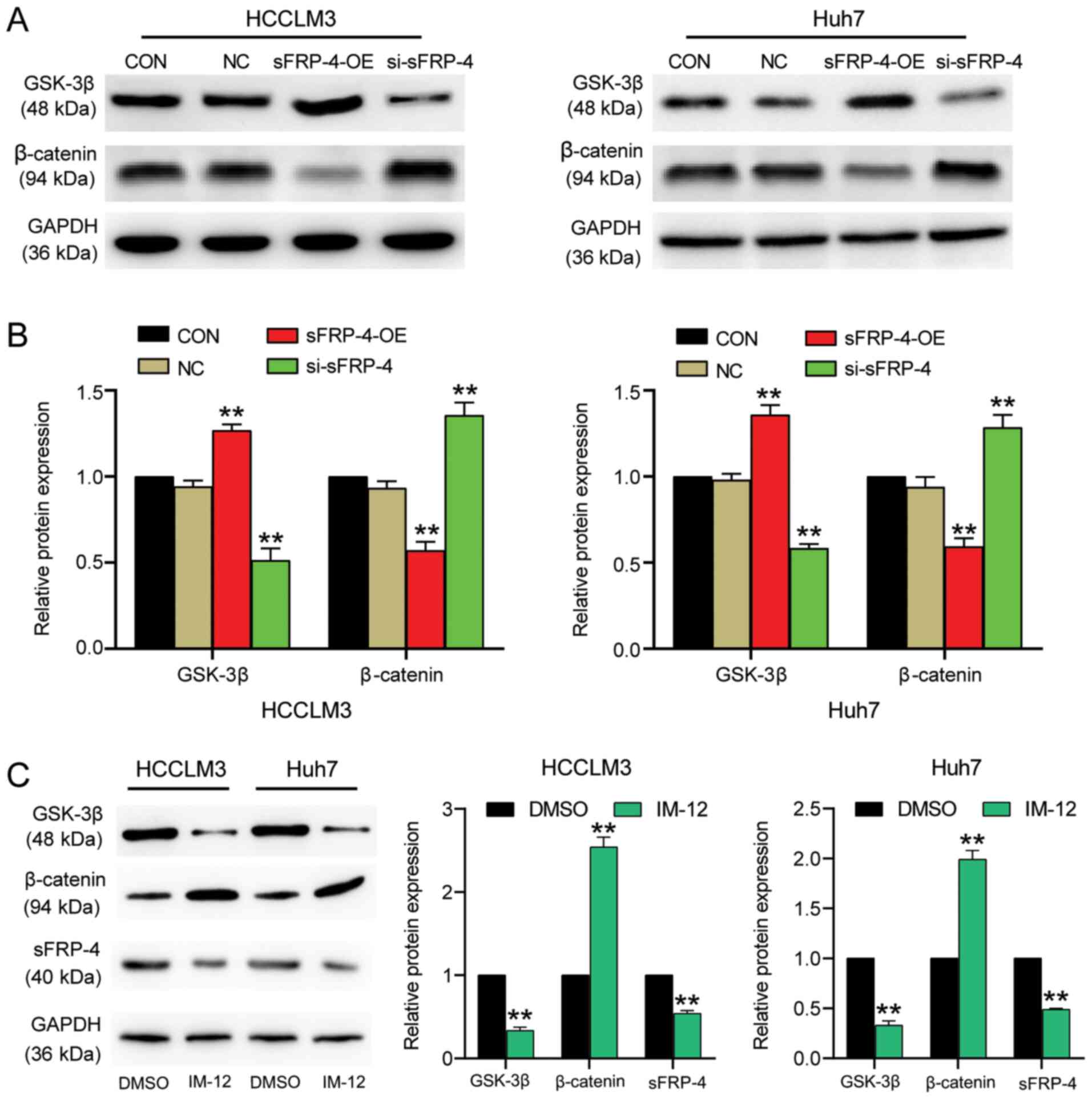
pathway were measured. The western blotting results demonstrated
that sFRP-4 knockdown significantly decreased GSK-3β protein
expression levels by >40% compared with the control group
(Fig. 3A and B). By contrast,
sFRP-4 knockdown significantly increased β-catenin levels by ~30%
compared with the control group. However, sFRP-4 overexpression
displayed the opposite effect, significantly increasing GSK-3β
expression levels by ~30% and decreasing β-catenin expression
levels by ~50% compared with the control group. The results
demonstrated that sFRP-4 increased GSK-3β expression and decreased
β-catenin expression, suggesting that sFRP-4 might suppress the
Wnt/β-catenin signaling pathway in HCC cells. To further evaluate
the effect of the Wnt/β-catenin signaling pathway on sFRP-4, IM-12,
a selective GSK-3β inhibitor, was used to activate the
Wnt/β-catenin signaling pathway in HCCLM3 and Huh7 cells (Fig. 3C). The western blotting results
demonstrated that GSK-3β protein expression levels were
significantly decreased by ~70%, whereas β-catenin protein
expression levels were significantly increased by >2-fold
following treatment with IM-12 compared with DMSO, which indicated
activation of the Wnt/β-catenin signaling pathway. Furthermore,
sFRP-4 expression was significantly decreased by ~50% in cells
treated with IM-12 compared with DMSO. The results suggested
negative feedback regulation of sFRP-4 expression by the
Wnt/β-catenin signaling pathway.
sFRP-4 inhibits HCC cell viability and
proliferation, and accelerates HCC cell apoptosis via the
Wnt/β-catenin signaling pathway
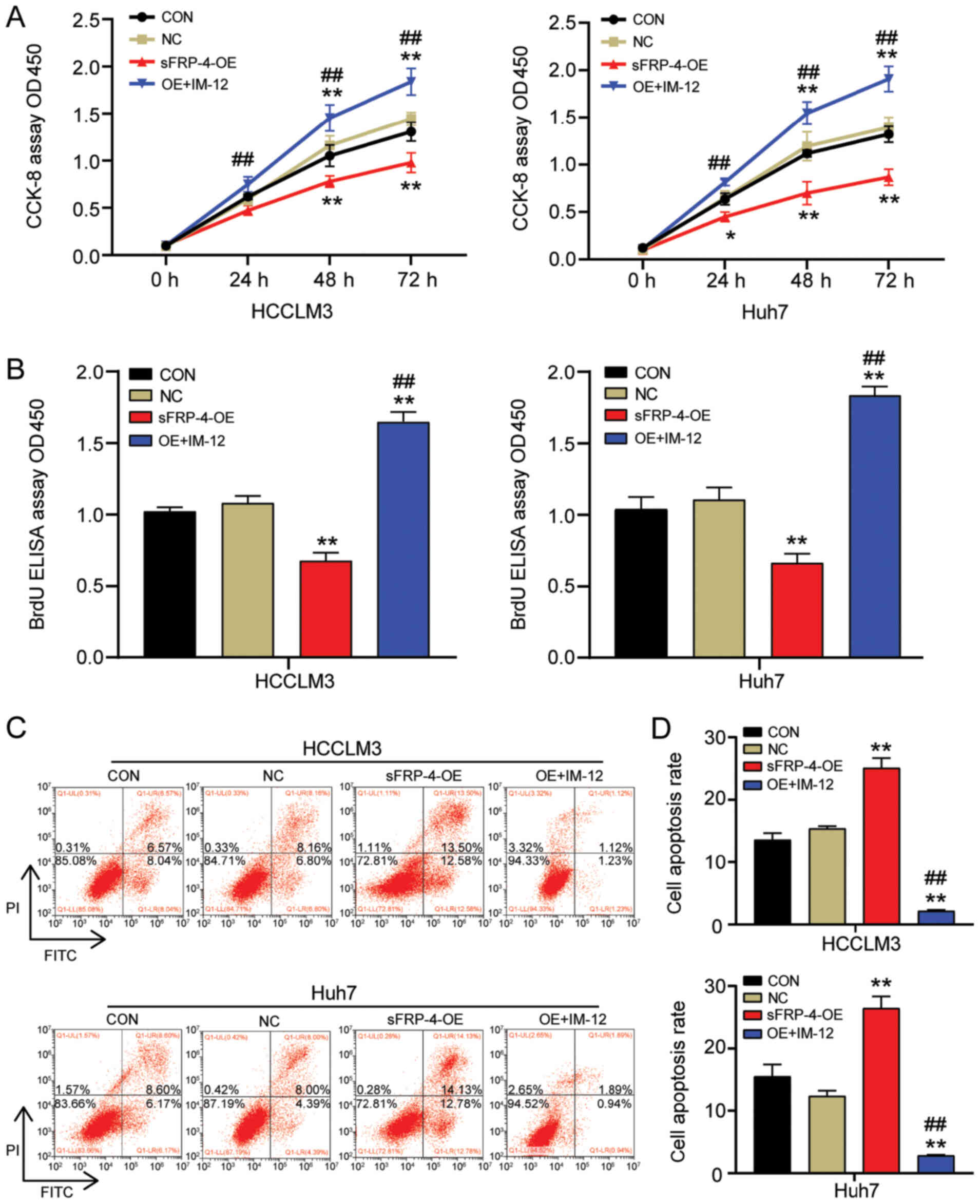
To further investigate whether sFRP-4 altered the
tumor phenotype of HCC cells via the Wnt/β-catenin signaling
pathway, the effect of sFRP-4 overexpression combined with IM-12
treatment on HCC cell viability, proliferation and apoptosis was
assessed by performing CCK-8, BrdU ELISA and flow cytometry assays,
respectively. The CCK-8 assay results demonstrated that IM-12
significantly reversed the suppressive effect of sFRP-4
overexpression on HCCLM3 and Huh7 cell viability (Fig. 4A). Similarly, the BrdU ELISA assay
results demonstrated that IM-12 significantly reversed the
inhibitory effects of sFRP-4 overexpression on HCCLM3 and Huh7 cell
proliferation (Fig. 4B). HCCLM3 and
Huh7 cell proliferation was significantly increased by 2.5-fold in
the sFRP-4-OE + IM-12 group compared with the sFRP-4-OE group.
Additionally, cell apoptosis was analyzed via flow cytometry. The
results demonstrated that IM-12 significantly reversed sFRP-4
overexpression-mediated induction of HCCLM3 and Huh7 cell apoptosis
(Fig. 4C and D). Compared with the
sFRP-4-OE group, the rate of apoptosis in HCCLM3 and Huh7 cells was
decreased by ~90% in the sFRP-4-OE + IM-12 group. The
aforementioned results further suggested that sFRP-4 inhibited the
malignant behavior of HCC cells by regulating the Wnt/β-catenin
signaling pathway.
Discussion
The present study demonstrated that sFRP-4
expression was significantly downregulated in HCC tissues and cells
compared with adjacent healthy tissues and MIHA cells,
respectively. Moreover, compared with the control group, sFRP-4
overexpression suppressed HCC cell viability and proliferation, but
facilitated HCC cell apoptosis. In addition, sFRP-4 overexpression
increased GSK-3β expression and decreased β-catenin expression
compared with the control group, which suggested that sFRP-4
inhibited the Wnt/β-catenin signaling pathway. IM-12 (a selective
of GSK-3β inhibitor) was used to investigate the inhibitory
function of sFRP-4. The results indicated that sFRP-4 inhibited the
malignant behavior of HCC cells by suppressing the Wnt/β-catenin
signaling pathway. Therefore, sFRP-4 may serve a negative
regulatory role during HCC carcinogenesis and development, which
indicated that sFRP-4 might serve as a promising therapeutic target
for HCC.
sFRPs are a class of Wnt regulatory proteins that
are often downregulated in a variety of different types of cancer,
such as gastric (38), oral
(39) and breast cancer (40), where they are associated with tumor
development and poor prognosis (41,40).
sFRP-4, a member of the sFRP family, is an underlying antagonist of
the Wnt signaling pathway (13).
Several studies have reported that sFRP-4 functions as a tumor
suppressor in various forms of cancer (42,43).
For instance, in mesothelioma and glioblastoma, the tumor
suppressor role of sFRP-4 has been demonstrated. Cell functional
experiments demonstrated that sFRP-4 overexpression displayed an
inhibitory effect on the malignant behavior of tumor cells
(41,44). Furthermore, in a study investigating
HCC, the sFRP-4 methylation level was evaluated in 12 HCC cell
lines and 19 HCC tissue samples. The results demonstrated that
sFRP-4 methylation was present in three HCC cell lines (HLF, CHC4
and CHC32) (45). The methylation
of sFRP family gene promoters led to transcriptional silencing of
the gene, thereby reducing its expression (46). Therefore, it was hypothesized that
sFRP-4 might be abnormally expressed in HCC. In our previous work,
it was demonstrated that compared with healthy patients, the
expression of sFRP-4 in the serum samples of patients with HCC was
obviously increased. After the diagnostic value of sFRP-4 in the
serum was evaluated, the study demonstrated that the combined use
of sFRP-4 and AFP (a standard serum marker of HCC) could enhance
the diagnostic accuracy of HCC (23). However, whether sFRP-4 is a tumor
suppressor or oncogene in HCC is not completely understood, thus
the mechanism underlying sFRP-4 in HCC is unclear. In the present
study, sFRP-4 expression was significantly downregulated in HCC
tissues and cells compared with adjacent healthy tissues and MIHA
cells, respectively. The IHC assay results further demonstrated
that the expression of sFRP-4 was notably reduced in HCC tissues
compared with healthy adjacent tissues. Therefore, the results of
the present study were contradictory to the results of our
aforementioned previous study that investigated serum expression
levels of sFRP-4. Although distinct methods were used to examine
sFRP4 expression in the two studies, including human antibody
arrays for serum detection, and RT-qPCR and IHC for tissue
detection, both results were convincing. Therefore, it was
hypothesized that the distinct sFRP4 expression in HCC tissues and
serum might derive from the multiple sources of serum sFRP4
according to the GeneCards description, including liver, lung,
spleen, bladder and reproductive organ, which suggested that the
high expression of sFRP4 in serum is the result of multiple body
tissues. However, it is possible that compensatory increases in
sFRP4 may be activated in other body tissues and eventually result
in the upregulation of sFRP4 in the serum of patients with HCC,
thus the aforementioned hypothesis requires further
investigation.
The present study also demonstrated that sFRP-4
knockdown not only enhanced HCC cell viability and proliferation,
but also suppressed HCC cell apoptosis compared with the control
group. However, sFRP-4 overexpression displayed the opposite
effect, which suggested that sFRP-4 might serve a tumor-suppressor
role during HCC carcinogenesis and development. Overall, the
results of the present study were consistent with the results of
previous studies investigating the role of sFRP-4 in glioblastoma
and mesothelioma (41,44).
In addition, numerous studies have reported that
excessive activation of the Wnt/β-catenin signaling pathway is
associated with different types of human cancer, including HCC
(47,48). The Wnt/β-catenin signaling pathway
is an intricate and precise regulation process, which is also known
as the canonical Wnt signaling pathway (37). The activation of the Wnt/β-catenin
signaling pathway can be identified as downregulation of GSK-3β and
stabilization of β-catenin, which is negatively regulated by GSK-3β
(49,50). Therefore, when the signaling pathway
is inactive, GSK-3β and other proteins constitute a degradation
complex that contributes to the phosphorylation of β-catenin,
resulting in the degradation of β-catenin. However, when the
signaling pathway is activated, this degradation complex is
dissociated and β-catenin degradation is suppressed, resulting in
accumulation of β-catenin in the cytoplasm (51–53).
Notably, as a mediator of the canonical Wnt signaling pathway,
β-catenin serves to activate a series of functional genes via
recruiting other transcriptional coactivators, such as Bcl-9 and
Pygopus (54,55). Therefore, the expression level of
β-catenin is closely related to the active state of the
Wnt/β-catenin signaling pathway (37).
Previous studies have also demonstrated that sFRP-4
overexpression in cancer could decrease the expression level of
β-catenin (15,41,56).
This finding was exemplified by a study on ovarian cancer.
Following treatment with human recombinant sFRP-4 treatment for 72
h in vitro, sFRP-4 protein expression levels were increased,
whereas β-catenin protein expression levels were decreased in the
nuclei of OVCAR3 cells, thus inhibiting the Wnt signaling pathway.
Furthermore, sFRP-4 inhibited OVCAR3 cell migration and EMT, and
promoted cell adhesion (15). In
addition, another study analyzed the protein expression levels of
downstream components (GSK-3β, phosphorylated β-catenin and
β-catenin) of the Wnt signaling pathway in prostate, breast and
ovarian cancer stem cells. The results indicated that following
demethylation of sFRP-4, the expression levels of sFRP-4, GSK-3β
and phosphorylated β-catenin were enhanced, whereas the expression
levels of the non-phosphorylated β-catenin were decreased,
resulting in inhibition of the Wnt signaling pathway (38).
In the present study, compared with the control
group, sFRP-4 overexpression significantly increased GSK-3β
expression and decreased β-catenin expression, whereas sFRP-4
knockdown significantly decreased GSK-3β expression and increased
β-catenin expression. In sFRP-4-overexpression HCC cells, IM-12 (an
inhibitor of GSK-3β, which is the key protein of the Wnt signaling
pathway) was used, and the results suggested that sFRP-4 inhibited
HCC cell viability and proliferation, and induced HCC cell
apoptosis by inhibiting the Wnt/β-catenin signaling pathway. In
addition, the results suggested that activation of the
Wnt/β-catenin signaling pathway displayed a negative feedback
effect on sFRP-4 expression, thereby counteracting the suppression
of sFRP-4 on the tumorous phenotype of HCC cells.
To conclude, the present study elucidated the tumor
suppressor role of sFRP-4 in HCC development. The results suggested
that sFRP-4 inhibited HCC cell viability and proliferation, and
promoted HCC cell apoptosis by inhibiting activation of the
Wnt/β-catenin signaling pathway. The results of the present study
might provide novel targets and theoretical foundations for the
development of HCC therapeutic strategies. However, the results of
the present study were limited as only isolated clinical HCC
tissues and cell lines were used; therefore, the results of the
present study should be verified using in vivo experiments,
and the sFRP-4 methylation level should be analyzed to further
investigate the mechanism underlying sFRP-4 in HCC.
Supplementary Material
Supporting Data
Acknowledgements
Not applicable.
Funding
The present study was supported by Hubei Provincial
Natural Science Foundation of China (grant no. 2019CFB101).
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
CX and ZR designed the study. QW and XZ performed
the majority of experiments. ZZ and BY performed the experiments
and wrote the manuscript. All authors read and approved the final
manuscript.
Ethics approval and consent to
participate
The present study was approved by the Ethics
Committee of General Hospital of the Central Theater Command of the
People's Liberation Army (approval no. [2020]024-2; Wuhan, China).
All patients provided written informed consent.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Bray F, Ferlay J, Soerjomataram I, Siegel
RL, Torre LA and Jemal A: Global cancer statistics 2018: GLOBOCAN
estimates of incidence and mortality worldwide for 36 cancers in
185 countries. CA Cancer J Clin. 68:394–424. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Llovet JM, Zucman-Rossi J, Pikarsky E,
Sangro B, Schwartz M, Sherman M and Gores G: Hepatocellular
carcinoma. Nat Rev Dis Primers. 2:160182016. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Xiang X, Zhong JH, Wang YY, You XM, Ma L,
Xiang BD and Li LQ: Distribution of tumor stage and initial
treatment modality in patients with primary hepatocellular
carcinoma. Clin Transl Oncol. 19:891–897. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Han J, Wang F, Lan Y, Wang J, Nie C, Liang
Y, Song R, Zheng T, Pan S, Pei T, et al: KIFC1 regulated by
miR-532-3p promotes epithelial-to-mesenchymal transition and
metastasis of hepatocellular carcinoma via gankyrin/AKT signaling.
Oncogene. 38:406–420. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Vilchez V, Turcios L, Marti F and Gedaly
R: Targeting Wnt/beta-catenin pathway in hepatocellular carcinoma
treatment. World J Gastroenterol. 22:823–832. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Saran U, Humar B, Kolly P and Dufour JF:
Hepatocellular carcinoma and lifestyles. J Hepatol. 64:203–214.
2016. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Ozakyol A: Global epidemiology of
hepatocellular carcinoma (HCC epidemiology). J Gastrointest Cancer.
48:238–240. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Serper M, Taddei TH, Mehta R, D'Addeo K,
Dai F, Aytaman A, Baytarian M, Fox R, Hunt K, Goldberg DS, et al:
Association of provider specialty and multidisciplinary care with
hepatocellular carcinoma treatment and mortality. Gastroenterology.
152:1954–1964. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Cheng D, Deng J, Zhang B, He X, Meng Z, Li
G, Ye H, Zheng S, Wei L, Deng X, et al: lncRNA HOTAIR
epigenetically suppresses miR-122 expression in hepatocellular
carcinoma via DNA methylation. EBioMedicine. 36:159–170. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Bukhari SA, Yasmin A, Zahoor MA, Mustafa
G, Sarfraz I and Rasul A: Secreted frizzled-related protein 4 and
its implication in obesity and type-2 diabetes. IUBMB Life.
71:1701–1710. 2019. View
Article : Google Scholar : PubMed/NCBI
|
|
11
|
Agostino M, Pohl S and Dharmarajan A:
Structure-based prediction of Wnt binding affinities for
Frizzled-type cysteine-rich domains. J Biol Chem. 292:11218–11229.
2017. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Feng Han Q, Zhao W, Bentel J, Shearwood
AM, Zeps N, Joseph D, Iacopetta B and Dharmarajan A: Expression of
sFRP-4 and beta-catenin in human colorectal carcinoma. Cancer Lett.
231:129–137. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Deshmukh A, Arfuso F, Newsholme P and
Dharmarajan A: Regulation of cancer stem cell metabolism by
secreted frizzled-related protein 4 (sFRP4). Cancers (Basel).
10:402018. View Article : Google Scholar
|
|
14
|
Wang H, Duan XL, Qi XL, Meng L, Xu YS, Wu
T and Dai PG: Concurrent hypermethylation of SFRP2 and
DKK2 activates the Wnt/β-catenin pathway and is associated
with poor prognosis in patients with gastric cancer. Mol Cells.
40:45–53. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Ford CE, Jary E, Ma SS, Nixdorf S,
Heinzelmann-Schwarz VA and Ward RL: The Wnt gatekeeper SFRP4
modulates EMT, cell migration and downstream Wnt signalling in
serous ovarian cancer cells. PLoS One. 8:e543622013. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Brebi P, Hoffstetter R, Andana A, Ili CG,
Saavedra K, Viscarra T, Retamal J, Sanchez R and Roa JC: Evaluation
of ZAR1 and SFRP4 methylation status as potentials biomarkers for
diagnosis in cervical cancer: Exploratory study phase I.
Biomarkers. 19:181–188. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
He B, Lee AY, Dadfarmay S, You L, Xu Z,
Reguart N, Mazieres J, Mikami I, McCormick F and Jablons DM:
Secreted frizzled-related protein 4 is silenced by hypermethylation
and induces apoptosis in beta-catenin-deficient human mesothelioma
cells. Cancer Res. 65:743–748. 2005.PubMed/NCBI
|
|
18
|
Pereira TDSF, Diniz MG, França JA, Moreira
RG, Menezes GHF, Sousa SF, Castro WH, Gomes CC and Gomez RS: The
Wnt/β-catenin pathway is deregulated in cemento-ossifying fibromas.
Oral Surg Oral Med Oral Pathol Oral Radiol. 125:172–178. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Sandsmark E, Andersen MK, Bofin AM,
Bertilsson H, Drabløs F, Bathen TF, Rye MB and Tessem MB: SFRP4
gene expression is increased in aggressive prostate cancer. Sci
Rep. 7:142762017. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Yang MW, Tao LY, Yang JY, Jiang YS, Fu XL,
Liu W, Huo YM, Li J, Zhang JF, Hua R, et al: SFRP4 is a prognostic
marker and correlated with Treg cell infiltration in pancreatic
ductal adenocarcinoma. Am J Cancer Res. 9:363–377. 2019.PubMed/NCBI
|
|
21
|
Nfonsam LE, Jandova J, Jecius HC, Omesiete
PN and Nfonsam VN: SFRP4 expression correlates with
epithelial mesenchymal transition-linked genes and poor overall
survival in colon cancer patients. World J Gastrointest Oncol.
11:589–598. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Huang D, Yu B, Deng Y, Sheng W, Peng Z,
Qin W and Du X: SFRP4 was overexpressed in colorectal carcinoma. J
Cancer Res Clin Oncol. 136:395–401. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Xu C, Zeng XH, Wang L, Tao SQ, Wu QX, Zhu
P, Deng GH and Wang YM: sFRP-4, a potential novel serum marker for
chronic hepatitis B-related hepatocellular carcinoma. Hepatobiliary
Pancreat Dis Int. 14:164–170. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Chen J, Rajasekaran M and Hui KM: Atypical
regulators of Wnt/β-catenin signaling as potential therapeutic
targets in Hepatocellular Carcinoma. Exp Biol Med (Maywood).
242:1142–1149. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Rao TP and Kuhl M: An updated overview on
Wnt signaling pathways: A prelude for more. Circ Res.
106:1798–1806. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Wang W, Li S, Liu P, Sideras K, van de
Werken HJG, van der Heide M, Cao W, Lavrijsen M, Peppelenbosch MP,
Bruno M, et al: Oncogenic STRAP supports hepatocellular carcinoma
growth by enhancing Wnt/β-catenin signaling. Mol Cancer Res.
17:521–531. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Zhan T, Rindtorff N and Boutros M: Wnt
signaling in cancer. Oncogene. 36:1461–1473. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Duchartre Y, Kim YM and Kahn M: The Wnt
signaling pathway in cancer. Crit Rev Oncol Hematol. 99:141–149.
2016. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Wang R, Wu Y, Huang W and Chen W:
MicroRNA-940 Targets INPP4A or GSK3beta and Activates the
Wnt/beta-Catenin pathway to regulate the malignant behavior of
bladder cancer Cells. Oncol Res. 26:145–155. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Peng Y, Zhang X, Ma Q, Yan R, Qin Y, Zhao
Y, Cheng Y, Yang M, Wang Q, Feng X, et al: miRNA-194 activates the
Wnt/β-catenin signaling pathway in gastric cancer by targeting the
negative Wnt regulator, SUFU. Cancer Lett. 385:117–127. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Ren Y, Tao J, Jiang Z, Guo D and Tang J:
Pimozide suppresses colorectal cancer via inhibition of
Wnt/beta-catenin signaling pathway. Life Sci. 209:267–273. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Zhang J, Hu SL, Qiao CH, Ye JF, Li M, Ma
HM, Wang JH, Xin SY and Yuan ZL: lncRNA-NEF inhibits proliferation,
migration and invasion of esophageal squamous-cell carcinoma cells
by inactivating wnt/β-catenin pathway. Eur Rev Med Pharmacol Sci.
22:6824–6831. 2018.PubMed/NCBI
|
|
33
|
Zhou P, Li Y, Li B, Zhang M, Liu Y, Yao Y
and Li D: NMIIA promotes tumor growth and metastasis by activating
the Wnt/β-catenin signaling pathway and EMT in pancreatic cancer.
Oncogene. 38:5500–5515. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Hu Z, Wang P, Lin J, Zheng X, Yang F,
Zhang G, Chen D, Xie J, Gao Z, Peng L and Xie C: MicroRNA-197
promotes metastasis of hepatocellular carcinoma by activating
Wnt/β-catenin signaling. Cell Physiol Biochem. 51:470–486. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Song J, Xie C, Jiang L, Wu G, Zhu J, Zhang
S, Tang M, Song L and Li J: Transcription factor AP-4 promotes
tumorigenic capability and activates the Wnt/β-catenin pathway in
hepatocellular carcinoma. Theranostics. 8:3571–3583. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Wang Q, Lv Q, Bian H, Yang L, Guo KL, Ye
SS, Dong XF and Tao LL: A novel tumor suppressor SPINK5 targets
Wnt/β-catenin signaling pathway in esophageal cancer. Cancer Med.
8:2360–2371. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Shin H, Kim JH, Lee YS and Lee YC: Change
in gene expression profiles of secreted frizzled-related proteins
(SFRPs) by sodium butyrate in gastric cancers: Induction of
promoter demethylation and histone modification causing inhibition
of Wnt signaling. Int J Oncol. 40:1533–1542. 2012.PubMed/NCBI
|
|
39
|
Pannone G, Bufo P, Santoro A, Franco R,
Aquino G, Longo F, Botti G, Serpico R, Cafarelli B, Abbruzzese A,
et al: WNT pathway in oral cancer: Epigenetic inactivation of
WNT-inhibitors. Oncol Rep. 24:1035–1041. 2010.PubMed/NCBI
|
|
40
|
Deshmukh A, Arfuso F, Newsholme P and
Dharmarajan A: Epigenetic demethylation of sFRPs, with emphasis on
sFRP4 activation, leading to Wnt signalling suppression and histone
modifications in breast, prostate, and ovary cancer stem cells. Int
J Biochem Cell Biol. 109:23–32. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Perumal V, Dharmarajan AM and Fox SA: The
Wnt regulator SFRP4 inhibits mesothelioma cell proliferation,
migration, and antagonizes Wnt3a via its netrin-like domain. Int J
Oncol. 51:362–368. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Pawar NM and Rao P: Secreted frizzled
related protein 4 (sFRP4) update: A brief review. Cell Signal.
45:63–70. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Bhuvanalakshmi G, Arfuso F, Millward M,
Dharmarajan A and Warrier S: Secreted frizzled-related protein 4
inhibits glioma stem-like cells by reversing epithelial to
mesenchymal transition, inducing apoptosis and decreasing cancer
stem cell properties. PLoS One. 10:e01275172015. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Bhuvanalakshmi G, Gamit N, Patil M, Arfuso
F, Sethi G, Dharmarajan A, Kumar AP and Warrier S: Stemness,
pluripotentiality, and Wnt antagonism: sFRP4, a wnt antagonist
mediates pluripotency and stemness in glioblastoma. Cancers
(Basel). 11:252018. View Article : Google Scholar
|
|
45
|
Takagi H, Sasaki S, Suzuki H, Toyota M,
Maruyama R, Nojima M, Yamamoto H, Omata M, Tokino T, Imai K and
Shinomura Y: Frequent epigenetic inactivation of SFRP genes in
hepatocellular carcinoma. J Gastroenterol. 43:378–389. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Yu J, Xie Y, Li M, Zhou F, Zhong Z, Liu Y,
Wang F and Qi J: Association between SFRP promoter hypermethylation
and different types of cancer: A systematic review and
meta-analysis. Oncol Lett. 18:3481–3492. 2019.PubMed/NCBI
|
|
47
|
Zhang T, Ma Z, Liu L, Sun J, Tang H, Zhang
B, Zou Y and Li H: DDX39 promotes hepatocellular carcinoma growth
and metastasis through activating Wnt/β-catenin pathway. Cell Death
Dis. 9:6752018. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Lin J, Lin W, Ye Y, Wang L, Chen X, Zang S
and Huang A: Kindlin-2 promotes hepatocellular carcinoma invasion
and metastasis by increasing Wnt/β-catenin signaling. J Exp Clin
Cancer Res. 36:1342017. View Article : Google Scholar : PubMed/NCBI
|
|
49
|
MacDonald BT, Tamai K and He X:
Wnt/beta-catenin signaling: Components, mechanisms, and diseases.
Dev Cell. 17:9–26. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Katoh M and Katoh M: WNT signaling pathway
and stem cell signaling network. Clin Cancer Res. 13:4042–4045.
2007. View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Cai J, Guan H, Fang L, Yang Y, Zhu X, Yuan
J, Wu J and Li M: MicroRNA-374a activates Wnt/β-catenin signaling
to promote breast cancer metastasis. J Clin Invest. 123:566–579.
2013.PubMed/NCBI
|
|
52
|
Fang L, Cai J, Chen B, Wu S, Li R, Xu X,
Yang Y, Guan H, Zhu X, Zhang L, et al: Aberrantly expressed
miR-582-3p maintains lung cancer stem cell-like traits by
activating Wnt/β-catenin signalling. Nat Commun. 6:86402015.
View Article : Google Scholar : PubMed/NCBI
|
|
53
|
Peng Y, Zhang X, Feng X, Fan X and Jin Z:
The crosstalk between microRNAs and the Wnt/beta-catenin signaling
pathway in cancer. Oncotarget. 8:14089–14106. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
54
|
Staal FJ and Clevers H: Tcf/Lef
transcription factors during T-cell development: Unique and
overlapping functions. Hematol J. 1:3–6. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
55
|
Kramps T, Peter O, Brunner E, Nellen D,
Froesch B, Chatterjee S, Murone M, Züllig S and Basler K:
Wnt/wingless signaling requires BCL9/legless-mediated recruitment
of pygopus to the nuclear beta-catenin-TCF complex. Cell.
109:47–60. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
56
|
Warrier S, Bhuvanalakshmi G, Arfuso F,
Rajan G, Millward M and Dharmarajan A: Cancer stem-like cells from
head and neck cancers are chemosensitized by the Wnt antagonist,
sFRP4, by inducing apoptosis, decreasing stemness, drug resistance
and epithelial to mesenchymal transition. Cancer Gene Ther.
21:381–388. 2014. View Article : Google Scholar : PubMed/NCBI
|