Introduction
Hepatocellular carcinoma (HCC) is the fifth most
common malignant tumor and is the second leading cause of
cancer-related death worldwide (1).
Despite the development of chemotherapy and active treatments, the
5-year survival rate of HCC (5–10%) remains low due to late
diagnosis, tumor recurrence and drug resistance (2–4).
Chemoresistance is the primary reason for cancer therapy failure
and remains a big challenge in clinical treatment (5). At present, doxorubicin (DOX) is the
first-line chemotherapy drug for transcatheter arterial embolic
therapies (TACE) in HCC (6).
However, the intrinsic or acquired resistance of HCC cells to DOX
attenuates the effects of TACE (7).
Therefore, understanding the mechanisms underlying DOX resistance
and developing suitable therapeutic targets for DOX resistance in
HCC is important.
Previous studies have demonstrated that most human
genome transcripts are transcribed into non-coding RNAs (ncRNAs),
including small ncRNAs and long ncRNAs (lncRNAs) (8,9).
lncRNAs are a class of ncRNA transcripts that are >200
nucleotides in length (10).
Increasing evidence has indicated that lncRNAs serve important
roles in the occurrence and development of tumors, such as HCC,
breast cancer cells and gastric cancer (10,11).
The effects and functions of lncRNAs in chemotherapy resistance
have also been extensively studied. Wu et al (12) reported that Keap1
regulation-associated lncRNA, as a competitive endogenous RNA,
enhanced sensitivity to 5-fluorouracil in HCC by binding to
microRNA (miR)-141 and regulating kelch like ECH associated protein
1. Moreover, downregulation of lncRNA NR2F1 antisense RNA 1
increased oxaliplatin sensitivity in HCC by targeting the
miR-363/ATP binding cassette subfamily C member 1 signaling pathway
(13). lncRNA ribosomal protein
L13a pseudogene 20 knockdown markedly inhibited cell proliferation,
enhanced apoptosis, suppressed tumor growth and increased
sensitivity to DOX (14). Moreover,
Li et al (15) reported that
lncRNA arsR promotes resistance to DOX in HCC by modulating the
PTEN/PI3K/Akt signaling pathway.
Long intergenic non-protein coding RNA
(linc)-regulator of reprogramming (ROR) was first discovered in
induced pluripotent stem cells (iPSCs). linc-ROR contributes to
epigenetic regulators involved in pluripotency and lineage
commitment (16,17). linc-ROR is frequently increased in
different types of cancer, such as, breast cancer, endometrial
cancer stem cells, pancreatic cancer and gastric cancer stem cells;
it is also correlated to poor prognosis and progression in cancer
(18–21). linc-ROR upregulation could promote
cell invasion and metastasis by regulating miR-145 and zinc finger
E-box binding homeobox 2, and inducing epithelial-mesenchymal
transition (EMT) in HCC (22).
Extracellular transfer of linc-ROR can promote cell survival during
hypoxic stress and chemoresistance (23,24).
However, the roles and functions of linc-ROR in drug resistance in
HCC are not completely understood.
The present study investigated the potential effect
of linc-ROR on DOX resistance in HCC, as well as the underlying
mechanisms.
Materials and methods
Cell culture
HCC cell lines (Hep3B, Huh7, SNU387 and SNU449) were
purchased from American Type Culture Collection. Huh7 and Hep3B
cells were cultured in DMEM (Gibco; Thermo Fisher Scientific, Inc.)
and MEM (Gibco; Thermo Fisher Scientific, Inc.), respectively.
SNU387 and SNU449 cells were cultured in RPMI-1640 (Gibco; Thermo
Fisher Scientific, Inc.). All culture medium was supplemented with
10% FBS (Gibco; Thermo Fisher Scientific, Inc.). Cells were
incubated at 37°C with 5% CO2. DOX was purchased from
Selleck Chemicals.
Cell transfection
linc-ROR small interfering RNA (siRNA), TWIST1 siRNA
(20 µM), TWIST1 overexpression (OE) vector (OE-TWIST1) (2 µg) and
empty vector, negative control (NC) siRNA (non-targeting) were
synthesized by Qiagen Benelux B.V. The control was the blank
control. HCC cells (2×105 cells/well) were transfected
with linc-ROR siRNA, TWIST1 siRNA, OE-TWIST1 or NC siRNA using
Lipofectamine® 2000 (Thermo Fisher Scientific, Inc.)
according to the manufacturer's protocol at 37°C for 6 h. The
primers for linc-ROR siRNA, TWIST1 siRNA and NC siRNA were as
follows: linc-ROR forward, 5′-GAUGGCACUAUGACUACAATT−3′ and reverse,
5′-UUGUAGUCAUAGUGCCAUCTT−3′; TWIST1 forward,
5′-GGUGUCUAAAUGCAUUCAUTT−3′ and reverse,
5′-AUGAAUGCAUUUAGACACCTT−3′; and NC siRNA forward,
5′-UUCUCCGAACGUGUCACGUTT−3′ and reverse,
5′-ACGUGACACGUUCGGAGAATT−3′.
Cell viability assay
Cells were seeded (5×103 cells/well) into
96-well plates. Following incubation for 12 h at 37°C, cells were
incubated in fresh culture medium containing different
concentrations of DOX (0–5 µg/ml) for 48 h at 37°C. Cell viability
was determined using the Cell Counting Kit-8 (CCK-8) assay (Dojindo
Molecular Technologies, Inc.). Then, 10 µl CCK-8 solution was
added, the cells were incubated for an additional 3 h at 37°C,
absorbance was measured at a wavelength of 450 nm using an MRX II
microplate reader (Dynex Technologies, Inc.).
EdU assay
To assess cell proliferation, HCC cells were seeded
(5×103 cells/well) into 96-well plates for 2 days.
Subsequently, cells were incubated with EdU for 60 min at 37°C,
followed by 4% paraformaldehyde for fixation and staining at room
temperature for 15–30 min for EdU-incorporated cells using a
ClickiT EdU Assay kit (Invitrogen; Thermo Fisher Scientific, Inc.).
Hoechst 33342 (Invitrogen; Thermo Fisher Scientific, Inc.) was used
to stain cell nuclei at room temperature for 30 min in the dark.
The proportion of EdU-positive cells was determined by randomly
counting cells under a fluorescent microscope in 5–10 fields of
view (magnification, ×100).
Reverse transcription-quantitative PCR
(RT-qPCR)
Total RNA was extracted using TRIzol®
(Invitrogen; Thermo Fisher Scientific, Inc.). Total RNA was reverse
transcribed into cDNA using the PrimeScript RT Reagent kit with
gDNA Eraser (Takara Biotechnology Co., Ltd.) according to the
manufacturer's protocol. Subsequently, qPCR was performed using
Premix Ex Taq II (Takara Biotechnology Co., Ltd.) and an ABI 7500
Fast system. The PCR conditions were as follows: Initial
denaturation at 95°C for 30 sec, followed by 40 cycles of 95°C for
30 sec, 60°C for 34 sec and 72°C for 30 sec. linc-ROR expression
levels were quantified using the 2−ΔΔCq method (25) and normalized to the internal
reference gene β-actin. RT-qPCR was performed in triplicate. The
following primers were used for qPCR: linc-ROR forward,
5′-ACCTGCAACACTCCAGCTAT-3′ and reverse, 5′-TGACCTGTTGACCCACCTTT-3′;
TWIST1 forward, 5′-GGUACAUCGACUUCCUCUATT−3′ and reverse,
5′-UAGAGGAAGUCGAUGUACCTT−3′; and β-actin forward,
5′-ATCAAGGAGAAGCTCTGCTACATC−3′ and reverse,
5′-TCAGACTCGGCTGGAAGAGA-3′.
Western blotting
Total protein was isolated from HCC cells using RIPA
lysis buffer (Beyotime Institute of Biotechnology). Protein
concentrations were determined using the Bradford protein assay
method (Thermo Fisher Scientific, Inc.). Proteins (40 µg) were
separated via 10% SDS-PAGE and transferred onto PVDF membranes
(Thermo Fisher Scientific, Inc.). After blocking with 5% TBS-Tween
20 (TBST; 0.1% Tween 20 containing 5% BSA (Sangon Biotech Co.,
Ltd.), for 1 h at 37°C. The membranes were incubated at 4°C
overnight with primary antibodies (all 1:1,000; Abcam) targeted
against the following: TWIST1 (cat. no. ab49254), E-cadherin (cat.
no. ab40772), vimentin (cat. no. ab92547). Following washing three
times with TBST, the membranes were incubated with a HRP-conjugated
secondary antibody (cat. no. ab7090; 1:2,000; Abcam) for 1 h at
37°C. Protein bands were visualized using an enhanced
chemiluminescence kit [Roche Diagnostics (Shanghai) Co., Ltd.].
GAPDH (Cell Signaling Technology, Inc.; cat. no. 5174S; 1:2,000)
was used as the loading control.
Immunofluorescence
HCC cells were seeded into 48-well plates at
3×103 cells/well. Cells were fixed with 4%
paraformaldehyde for 15 min at room temperature, washed three times
with PBS (3 min per wash), permeated with 0.5% Triton X-100
(prepared in PBS) for 20 min at room temperature, washed three
times with PBS (3 min per wash). Following absorption of PBS using
absorbent paper, blocked using 1% BSA for 30 min in the room
temperature, cells were incubated with FITC-conjugated primary
antibodies targeted against E-cadherin (cat. no. ab40772; 1:50;
Abcam) and vimentin (cat. no. ab92547; 1:50; Abcam) overnight at
4°C. The cells were incubated with FITC-conjugated secondary
antibody (cat. no. ab6785; 1:100; Abcam) at 4°C for 2 h.
Subsequently, cells were incubated with DAPI for 5 min at room
temperature in the dark to stain cell nuclei. Following washing
four times with PBST (0.1% Tween 20 containing 5% BSA) (5 min per
wash), stained cells were observed and imaged using a confocal
fluorescence microscope (magnification, ×100).
Statistical analysis
Comparisons between two groups were analyzed using
the Student's t-test followed by unpaired t-test. Comparisons among
multiple groups were analyzed using one-way ANOVA followed by
Tukey's post hoc test. StarBase (version 3; starbase.sysu.edu.cn/index.php) was used to analyze
the level of linc-ROR in liver HCC (LIHC). P<0.05 was considered
to indicate a statistically significant difference. All experiments
were performed in triplicate.
Results
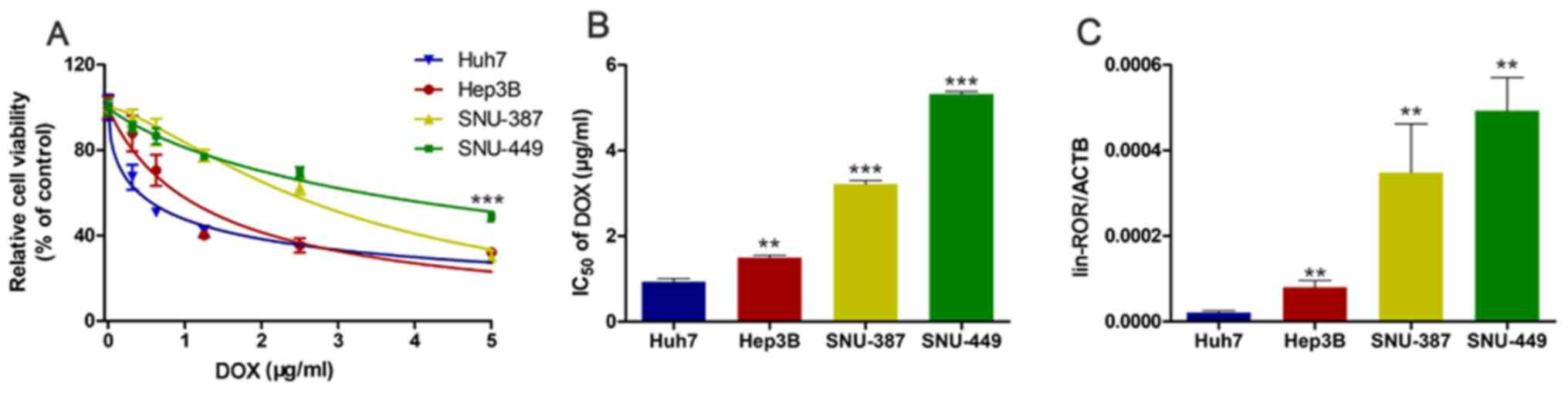
linc-ROR expression is negatively
associated with sensitivity to DOX in HCC cells
To investigate the relationship between linc-ROR
expression and DOX sensitivity, HCC cell viability following DOX
treatment was assessed. Among the HCC cell lines used in the
present study, the Huh7 cell line was the most DOX-sensitive,
whereas the SNU449 cell line was the most DOX-resistant (Fig. 1A). The IC50 of DOX in the
HCC cell lines, from highest to lowest, was SNU449, SNU387, Hep3B
and Huh7 cells (Fig. 1B).
Subsequently, linc-ROR expression levels in the four HCC cell lines
were measured. Among the HCC cell lines, SNU449 cells and Huh7
cells displayed the highest and lowest linc-ROR expression levels,
respectively (Fig. 1C), which
indicated that linc-ROR expression was negatively associated with
DOX sensitivity. Furthermore, starBase was used to analyze linc-ROR
expression levels in LIHC. linc-ROR expression levels were higher
in the 374 LIHC samples compared with the 50 healthy samples
(Fig. S1A).
linc-ROR knockdown increases
sensitivity to DOX in HCC cells
To investigate the effect of linc-ROR on DOX
sensitivity in HCC, interference oligonucleotides were synthesized
and transfected into HCC cells to knock down linc-ROR (Fig. 2A). Cell viability was assessed by
performing the CCK-8 assay. The results indicated that the linc-ROR
knockdown group was significantly more sensitive to DOX compared
with the control group (Fig. 2B).
The EdU assay results for cell proliferation were similar to the
CCK-8 assay results, indicating that linc-ROR knockdown
significantly enhanced DOX sensitivity compared with the DOX group
(Fig. 2C).
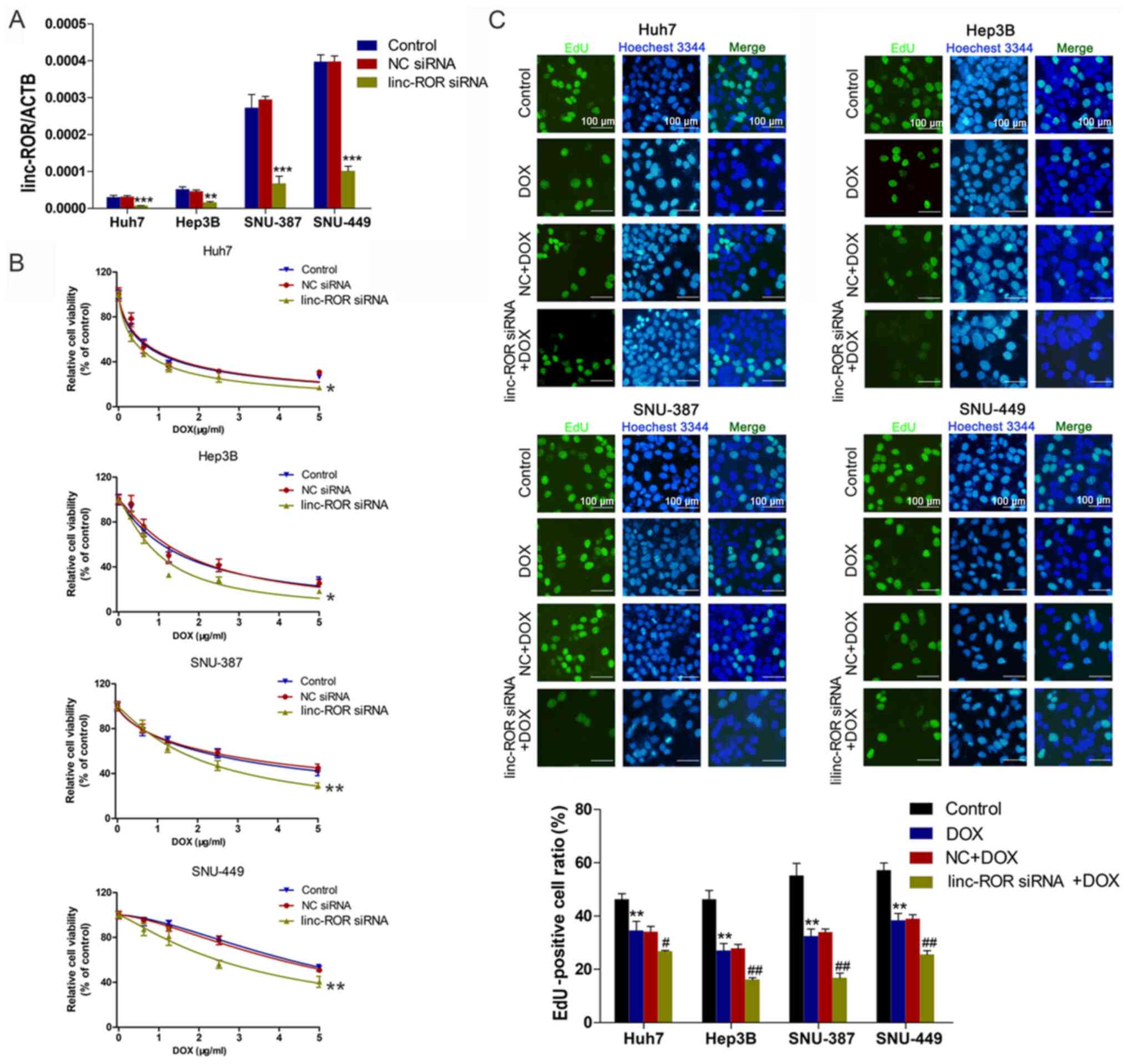 | Figure 2.Effect of linc-ROR on DOX sensitivity
in HCC cells. (A) Transfection efficiency of linc-ROR siRNA.
**P<0.01 and ***P<0.001 vs. control. (B) HCC cell viability
following transfection with linc-ROR siRNA or NC siRNA and
treatment with 0, 0.3125, 0.625, 1.25, 2.5 or 5 µg/ml DOX.
*P<0.05 and **P<0.01 vs. control (C) HCC cell proliferation
following treatment with DOX (IC50), NC siRNA + DOX or
linc-ROR siRNA + DOX (magnification, ×100). **P<0.01 vs.
control; #P<0.05 and ##P<0.01 vs. DOX.
linc-ROR, long intergenic non-protein coding RNA-regulator of
reprogramming; DOX; HCC, hepatocellular carcinoma; siRNA, small
interfering RNA; NC, negative control; ACTB, β-actin. |
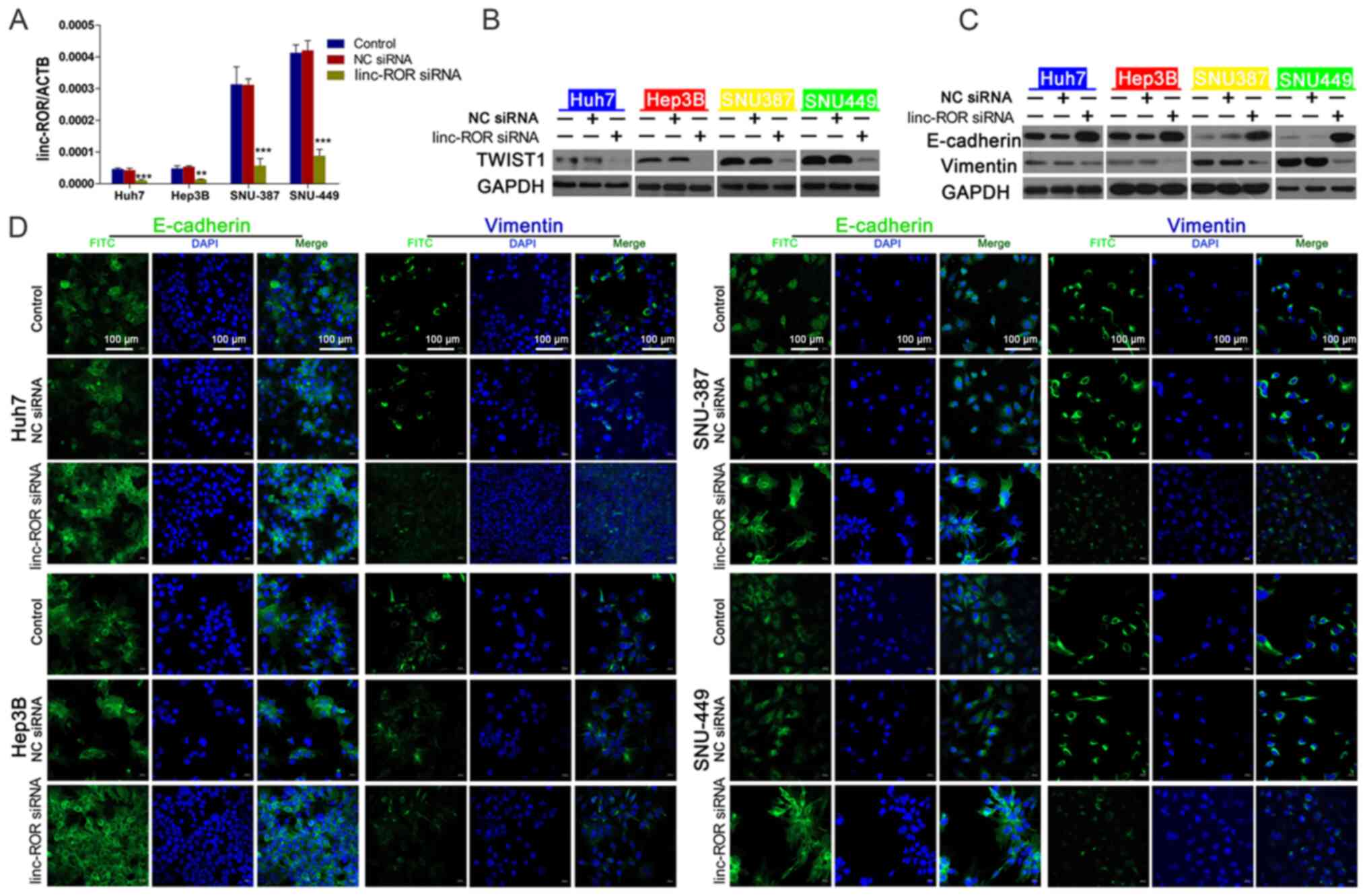
linc-ROR knockdown inhibits the EMT
signaling pathway by downregulating TWIST1
To explore whether the EMT signaling pathway
mediated linc-ROR-induced DOX resistance in HCC, the expression of
EMT-related proteins was assessed. The transfection efficiency of
linc-ROR siRNA was examined via RT-qPCR (Fig. 3A). Compared with NC siRNA, linc-ROR
knockdown markedly downregulated the expression levels of TWIST1,
an important promoter of EMT (26)
(Fig. 3B). E-cadherin (an epidermal
marker) (26) expression was
notably upregulated, whereas vimentin (a mesenchymal marker)
(26) expression was clearly
downregulated by linc-ROR knockdown compared with the NC siRNA
group (Fig. 3C). To explore the
effects of linc-ROR knockdown on the EMT signaling pathway,
immunofluorescence assays were performed to examine E-cadherin and
vimentin expression. The immunofluorescence assay results were
similar to the western blotting results, as linc-ROR knockdown
notably increased E-cadherin expression, but markedly decreased
vimentin expression compared with the NC siRNA group (Fig. 3D).
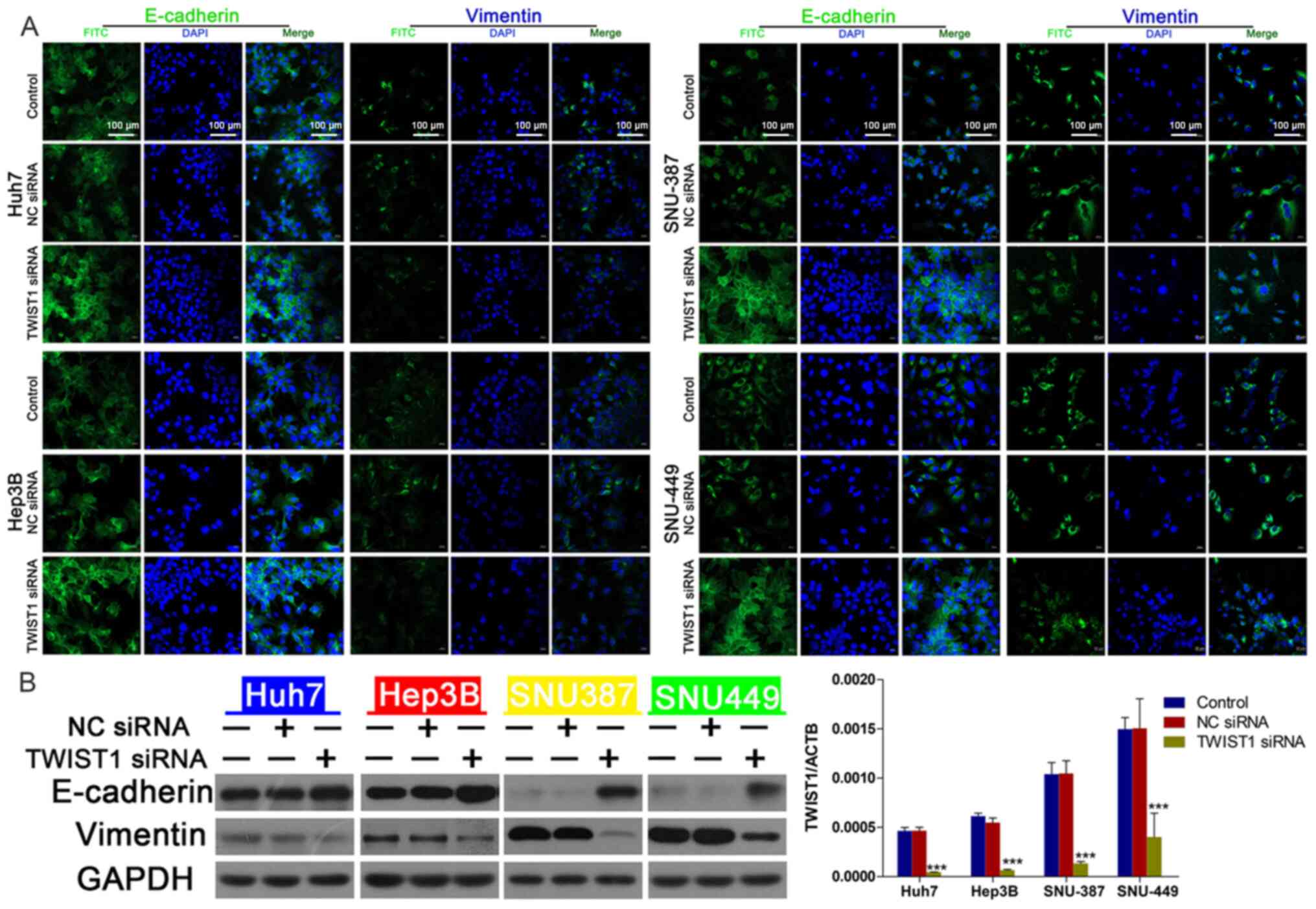
TWIST1 mediates linc-ROR
knockdown-regulated DOX sensitivity
To further investigate the effect of TWIST1 on
linc-ROR-mediated DOX resistance, the effect of TWIST1 knockdown on
HCC cell viability was assessed. The transfection efficiency of
TWIST1 knockdown was determined via western blotting (Fig. 4C). TWIST1-knockdown HCC cells were
significantly more sensitive to DOX compared with the control group
(Fig. 4A), EdU assay results
demonstrated that TWIST siRNA combined with DOX could reduce cell
proliferation compared with DOX group (Fig. 4B). Compared with the NC siRNA group,
TWIST1 knockdown also notably upregulated E-cadherin expression and
markedly downregulated vimentin expression, which was similar to
the results obtained for linc-ROR knockdown (Fig. 5B). The western blotting (Fig. 5B) and immunofluorescence (Fig. 5A) results for E-cadherin and
vimentin expression were consistent. Furthermore, there was no
significant difference in cell viability between the TWIST1 siRNA +
NC siRNA group and the TWIST1 siRNA + linc-ROR siRNA group
(Fig. 6). In addition, linc-ROR
knockdown-induced DOX sensitivity was inhibited by TWIST1
overexpression (Fig. S1B and C).
The expression of TWIST was determined by RT-qPCR (Fig. S1D).
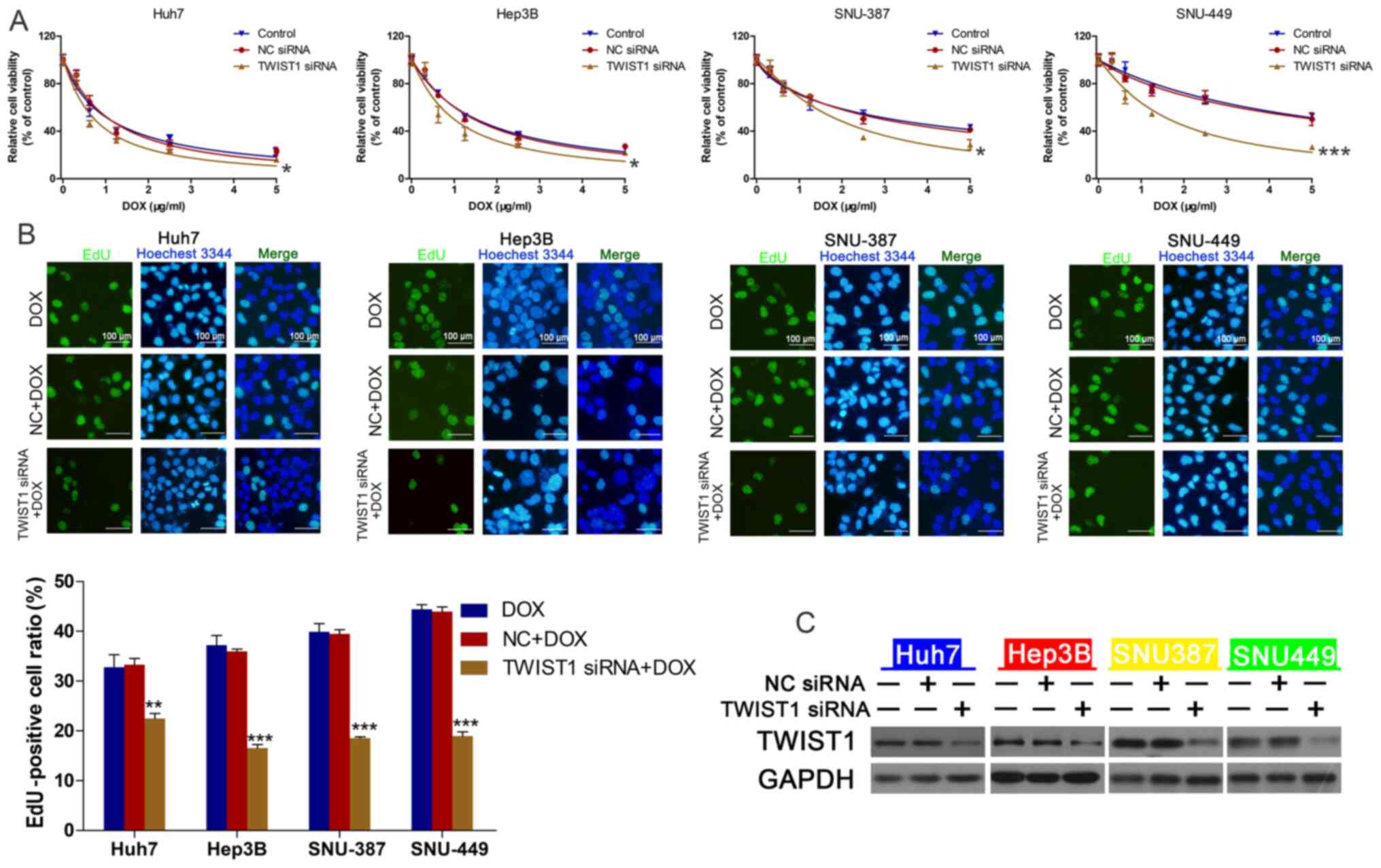 | Figure 4.TWIST1 knockdown regulates DOX
sensitivity in HCC cells. (A) HCC cell viability following
transfection with TWIST1 siRNA or NC siRNA and treatment with 0,
0.3125, 0.625, 1.25, 2.5 or 5 µg/ml DOX. *P<0.05 and
***P<0.001 vs. control. (B) HCC cell proliferation following
treatment with DOX (IC50), NC siRNA + DOX or linc-ROR
siRNA + DOX (magnification, ×100). **P<0.01 and ***P<0.001
vs. DOX. (C) Transfection efficiency of TWIST1 siRNA. TWIST1, twist
family bHLH transcription factor 1; DOX, doxorubicin; HCC,
hepatocellular carcinoma; siRNA, small interfering RNA; NC,
negative control; linc-ROR, long intergenic non-protein coding
RNA-regulator of reprogramming. |
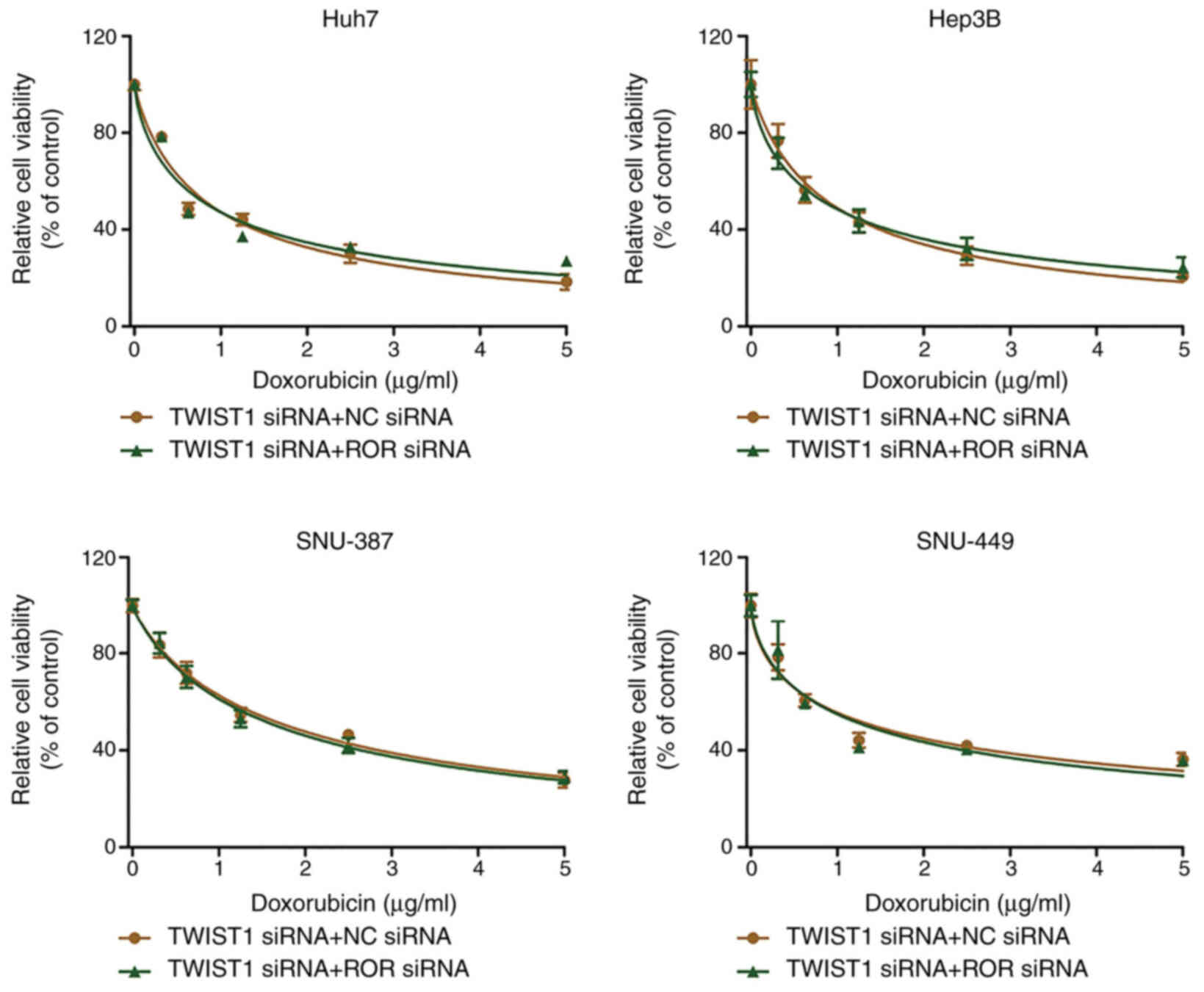 | Figure 6.TWIST1 knockdown reverses linc-ROR
knockdown-mediated effects on DOX sensitivity. HCC cells were
co-transfected with TWIST1 siRNA and linc-ROR siRNA or NC siRNA.
Subsequently, HCC cell viability was assessed following treatment
with 0, 0.3125, 0.625, 1.25, 2.5 or 5 µg/ml DOX. TWIST1, twist
family bHLH transcription factor 1; linc-ROR, long intergenic
non-protein coding RNA-regulator of reprogramming; DOX,
doxorubicin; HCC, hepatocellular carcinoma; siRNA, small
interfering RNA; NC, negative control. |
Discussion
HCC is an aggressive malignant tumor, and although
great progress in the treatment of HCC has been achieved, it still
has a high recurrence rate and mortality rate (27,28). A
primary pathological characteristic of HCC is chemoresistance
against a series of chemotherapeutic drugs, including cisplatin,
DOX and sorafenib (29,30). lncRNAs have been widely reported to
participate in regulating tumorigenesis and chemoresistance in HCC
cells (31,32). Moreover, linc-ROR participates in
the regulation of malignant biological properties in HCC, such as
proliferation and metastasis (22,33,34);
however, the effects of linc-ROR in mediating HCC chemoresistance
are not completely understood.
linc-ROR is a 2.6-kb lncRNA comprised of four exons
that was first described in iPSCs. linc-ROR is regulated by NANOG,
SOX2 and OCT4 (16). linc-ROR is
upregulated in numerous solid tumors, including ovarian, lung,
esophagus and renal cancer (35–38).
Furthermore, it has been reported that linc-ROR can regulate HCC
cell migration, proliferation and hypoxia signaling pathways in HCC
(22,23). Extracellular vesicle-mediated
transfer of linc-ROR could promote chemoresistance in HCC cells
(24). In the present study, the
Huh7 cell line, which displayed the lowest linc-ROR expression
levels among the HCC cell lines, was the most DOX-sensitive. By
contrast, the SNU449 cell line displayed the highest expression
levels of linc-ROR among the HCC cell lines and was the most
DOX-resistant. Furthermore, linc-ROR expression levels in LIHC were
analyzed using starBase. linc-ROR expression was higher in the 374
LIHC samples compared with the 50 healthy samples. However, the
specimens were not used to construct survival curves, so this
should be analyzed in a future study. Collectively, the present
study demonstrated that compared with the control group, linc-ROR
knockdown promoted sensitivity to DOX in HCC cells.
EMT is a complex process that is controlled by
various transcriptional regulatory agencies via different signaling
pathways, involving TWIST1, snail family transcriptional repressor
2 and snail family transcriptional repressor 1 (39). EMT also serves vital roles in the
chemoresistance of cancer cells (40). The TWIST family, consisting of
TWIST1 and TWIST2, serves a crucial role in the regulation of EMT
(41). TWIST1 leads to reduced
E-cadherin expression and increased mesenchymal marker vimentin
expression, resulting in HCC cell invasion (42,43).
TWIST is critically involved in drug resistance. For example,
TWIST1 downregulation reduced drug resistance by regulating ATP
binding cassette subfamily B member 1 and ATP binding cassette
subfamily C member 1 expression in colon cancer cells (44). Mesenchyme homeobox 2 and TWIST1
upregulation are associated with lung cancer chemoresistance and
prognosis (45). In HCC, the
platelet-derived growth factor D/miR-106a/TWIST1 signaling pathway
promotes gemcitabine resistance by regulating EMT (46). In the present study, linc-ROR
knockdown notably downregulated the expression levels of TWIST1 and
E-cadherin compared with the NC siRNA group. The human HCC cell
lines Huh7 and Hep3B, which display an epithelial phenotype,
displayed markedly higher E-cadherin expression levels compared
with SNU387 and SNU449 cells, which display a mesenchymal phenotype
(47). It has been previously
reported that cells with the mesenchymal phenotype are more
resistant compared with cells with the epithelial phenotype
(47). Furthermore, the results of
the present study indicated that linc-ROR regulated the expression
of TWIST1, thereby promoting the EMT signaling pathway. Moreover,
CCK-8 assays and western blotting were performed to examine cell
viability and EMT-related protein expression levels following
co-transfection of linc-ROR siRNA and OE-TWIST1, respectively. The
results demonstrated that the effects of linc-ROR siRNA on HCC cell
sensitivity to DOX were decreased after co-transfection of
OE-TWIST1 and lin-ROR siRNA, indicating that linc-ROR regulated
sensitivity to DOX via TWIST1 and EMT in HCC cells. A key
limitation of the present study was that the mechanism underlying
ROR-mediated regulation of TWIST1 was not investigated. It was
hypothesized that linc-ROR might regulate TWIST1 via the PI3K/Akt
and IL-6/STAT3 signaling pathways, but this should be investigated
further in future studies.
In conclusion, the results of the present study
provided novel mechanistic insight into chemotherapy resistance in
HCC. To the best of our knowledge, the present study demonstrated
linc-ROR/TWIST1 axis-mediated regulation of chemoresistance in HCC
for the first time. Therefore, targeting the linc-ROR/TWIST1 axis
might increase sensitivity and improve responses to traditional
therapeutic agents used for HCC.
Supplementary Material
Supporting Data
Acknowledgements
Not applicable.
Funding
The present study was supported by the Science and
Technology Department Public Welfare Project of Zhejiang Province
(grant no. LGF18H160023), the Administration of Traditional Chinese
Medicine of Zhejiang Province (grant no. 2015ZA012), The National
Natural Science Foundation of China (grant nos. 81572307, 81773096
and 81701630), the Major Project of Medical and Health Technology
Development Program in Zhejiang Province (grant no. 7211902), the
Zhejiang Provincial Medical and Health Research Project (grant nos.
2018KY126 and 2021KY030) and the Projects of Lishui Key Research
and Development Plan in Zhejiang Province (grant no.
2017ZDYF12).
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
WLW and DZ conceptualized the study. YZ and WDW
performed the experiments and confirmed the authenticity of the raw
data. QS and LY analyzed the data. WLW drafted the manuscript. All
authors read and approved the final version of the manuscript.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Siegel RL, Miller KD and Jemal A: Cancer
statistics, 2017. CA Cancer J Clin. 67:7–30. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Song P, Cai Y, Tang H, Li C and Huang J:
The clinical management of hepatocellular carcinoma worldwide: A
concise review and comparison of current guidelines from 2001 to
2017. Biosci Trends. 11:389–398. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Ozer Etik D, Suna N and Boyacioglu AS:
Management of hepatocellular carcinoma: Prevention, surveillance,
diagnosis, and staging. Exp Clin Transplant. 15 (Suppl 2):S31–S35.
2017.
|
|
4
|
Kulik LM: Advancements in hepatocellular
carcinoma. Curr Opin Gastroenterol. 23:268–274. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Clark T, Maximin S, Meier J, Pokharel S
and Bhargava P: Hepatocellular carcinoma: Review of epidemiology,
screening, imaging diagnosis, response assessment, and treatment.
Curr Probl Diagn Radiol. 44:479–486. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Nishikawa H, Kita R, Kimura T and Osaki Y:
Transcatheter arterial embolic therapies for hepatocellular
carcinoma: A literature review. Anticancer Res. 34:6877–6886.
2014.PubMed/NCBI
|
|
7
|
Pastorelli D, Cartei G, Zustovich F,
Marchese F, Artioli G, Zovato S, Binato S, Ceravolo R, Cingarlini
S, Salmaso F, et al: Gemcitabine and liposomal doxorubicin in
biliary and hepatic carcinoma (HCC) chemotherapy: Preliminary
results and review of the literature. Ann Oncol. 17 (Suppl
5):v153–v157. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Yang JX, Rastetter RH and Wilhelm D:
Non-coding RNAs: An introduction. Adv Exp Med Biol. 886:13–32.
2016. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Wei L, Wang X, Lv L, Liu J, Xing H, Song
Y, Xie M, Lei T, Zhang N and Yang M: The emerging role of microRNAs
and long noncoding RNAs in drug resistance of hepatocellular
carcinoma. Mol Cancer. 18:1472019. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Kondo Y, Shinjo K and Katsushima K: Long
non-coding RNAs as an epigenetic regulator in human cancers. Cancer
Sci. 108:1927–1933. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Weidle UH, Birzele F, Kollmorgen G and
Ruger R: Long non-coding RNAs and their role in metastasis. Cancer
Genomics Proteomics. 14:143–160. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Wu L, Pan C, Wei X, Shi Y, Zheng J, Lin X
and Shi L: lncRNA KRAL reverses 5-fluorouracil resistance in
hepatocellular carcinoma cells by acting as a ceRNA against
miR-141. Cell Commun Signal. 16:472018. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Huang H, Chen J, Ding CM, Jin X, Jia ZM
and Peng J: LncRNA NR2F1-AS1 regulates hepatocellular carcinoma
oxaliplatin resistance by targeting ABCC1 via miR-363. J Cell Mol
Med. 22:3238–3245. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Xiao J, Lv Y, Jin F, Liu Y, Ma Y, Xiong Y,
Liu L, Zhang S, Sun Y, Tipoe GL, et al: LncRNA HANR promotes
tumorigenesis and increase of chemoresistance in hepatocellular
carcinoma. Cell Physiol Biochem. 43:1926–1938. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Li Y, Ye Y, Feng B and Qi Y: Long
noncoding RNA lncARSR promotes doxorubicin resistance in
hepatocellular carcinoma via modulating PTEN-PI3K/Akt pathway. J
Cell Biochem. 118:4498–4507. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Loewer S, Cabili MN, Guttman M, Loh YH,
Thomas K, Park IH, Garber M, Curran M, Onder T, Agarwal S, et al:
Large intergenic non-coding RNA-RoR modulates reprogramming of
human induced pluripotent stem cells. Nat Genet. 42:1113–1117.
2010. View
Article : Google Scholar : PubMed/NCBI
|
|
17
|
Wang Y, Xu Z, Jiang J, Xu C, Kang J, Xiao
L, Wu M, Xiong J, Guo X and Liu H: Endogenous miRNA sponge
lincRNA-RoR regulates Oct4, Nanog, and Sox2 in human embryonic stem
cell self-renewal. Dev Cell. 25:69–80. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Hou P, Zhao Y, Li Z, Yao R, Ma M, Gao Y,
Zhao L, Zhang Y, Huang B and Lu J: LincRNA-ROR induces
epithelial-to-mesenchymal transition and contributes to breast
cancer tumorigenesis and metastasis. Cell Death Dis. 5:e12872014.
View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Zhou X, Gao Q, Wang J, Zhang X, Liu K and
Duan Z: Linc-RNA-RoR acts as a ‘sponge’ against mediation of the
differentiation of endometrial cancer stem cells by microRNA-145.
Gynecol Oncol. 133:333–339. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Gao S, Wang P, Hua Y, Xi H, Meng Z, Liu T,
Chen Z and Liu L: ROR functions as a ceRNA to regulate Nanog
expression by sponging miR-145 and predicts poor prognosis in
pancreatic cancer. Oncotarget. 7:1608–1618. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Wang S, Liu F, Deng J, Cai X, Han J and
Liu Q: Long noncoding RNA ROR regulates proliferation, invasion,
and stemness of gastric cancer stem cell. Cell Reprogram.
18:319–326. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Li C, Lu L, Feng B, Zhang K, Han S, Hou D,
Chen L, Chu X and Wang R: The lincRNA-ROR/miR-145 axis promotes
invasion and metastasis in hepatocellular carcinoma via induction
of epithelial-mesenchymal transition by targeting ZEB2. Sci Rep.
7:46372017. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Takahashi K, Yan IK, Haga H and Patel T:
Modulation of hypoxia-signaling pathways by extracellular linc-RoR.
J Cell Sci. 127:1585–1594. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Takahashi K, Yan IK, Kogure T, Haga H and
Patel T: Extracellular vesicle-mediated transfer of long non-coding
RNA ROR modulates chemosensitivity in human hepatocellular cancer.
FEBS Open Bio. 4:458–467. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Odero-Marah V, Hawsawi O, Henderson V and
Sweeney J: Epithelial-mesenchymal transition (EMT) and prostate
cancer. Adv Exp Med Biol. 1095:101–110. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Yee NS: Update in systemic and targeted
therapies in gastrointestinal oncology. Biomedicines. 6:342018.
View Article : Google Scholar
|
|
28
|
Kudo M: Systemic therapy for
hepatocellular carcinoma: 2017 update. Oncology. 93 (Suppl
1):S135–S146. 2017. View Article : Google Scholar
|
|
29
|
Nishida N, Kitano M, Sakurai T and Kudo M:
Molecular mechanism and prediction of sorafenib chemoresistance in
human hepatocellular carcinoma. Dig Dis. 33:771–779. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Lohitesh K, Chowdhury R and Mukherjee S:
Resistance a major hindrance to chemotherapy in hepatocellular
carcinoma: An insight. Cancer Cell Int. 18:442018. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Li C, Chen J, Zhang K, Feng B, Wang R and
Chen L: Progress and prospects of long noncoding RNAs (lncRNAs) in
hepatocellular carcinoma. Cell Physiol Biochem. 36:423–434. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Li CH and Chen Y: Targeting long
non-coding RNAs in cancers: Progress and prospects. Int J Biochem
Cell Biol. 45:1895–1910. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Chen Y, Shen Z, Zhi Y, Zhou H, Zhang K,
Wang T, Feng B, Chen Y, Song H, Wang R and Chu X: Long non-coding
RNA ROR promotes radioresistance in hepatocelluar carcinoma cells
by acting as a ceRNA for microRNA-145 to regulate RAD18 expression.
Arch Biochem Biophys. 645:117–125. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Pan Y, Li C, Chen J, Zhang K, Chu X, Wang
R and Chen L: The emerging roles of long noncoding RNA ROR
(lincRNA-ROR) and its possible mechanisms in human cancers. Cell
Physiol Biochem. 40:219–229. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Lou Y, Jiang H, Cui Z, Wang L, Wang X and
Tian T: Linc-ROR induces epithelial-to-mesenchymal transition in
ovarian cancer by increasing Wnt/β-catenin signaling. Oncotarget.
8:69983–69994. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Pan Y, Chen J, Tao L, Zhang K, Wang R, Chu
X and Chen L: Long noncoding RNA ROR regulates chemoresistance in
docetaxel-resistant lung adenocarcinoma cells via epithelial
mesenchymal transition pathway. Oncotarget. 8:33144–33158. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Sahebi R, Malakootian M, Balalaee B,
Shahryari A, Khoshnia M, Abbaszadegan MR, Moradi A and Javad Mowla
S: Linc-ROR and its spliced variants 2 and 4 are significantly
up-regulated in esophageal squamous cell carcinoma. Iran J Basic
Med Sci. 19:1131–1135. 2016.PubMed/NCBI
|
|
38
|
Shi J, Zhang W, Tian H, Zhang Q and Men T:
lncRNA ROR promotes the proliferation of renal cancer and is
negatively associated with favorable prognosis. Mol Med Rep.
16:9561–9566. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Chaffer CL, San Juan BP, Lim E and
Weinberg RA: EMT, cell plasticity and metastasis. Cancer Metastasis
Rev. 35:645–654. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Zheng HC: The molecular mechanisms of
chemoresistance in cancers. Oncotarget. 8:59950–59964. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Kang Y and Massagué J:
Epithelial-mesenchymal transitions: Twist in development and
metastasis. Cell. 118:277–279. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Wang Y, Sun B, Zhao X, Zhao N, Sun R, Zhu
D, Zhang Y, Li Y, Gu Q, Dong X, et al: Twist1-related miR-26b-5p
suppresses epithelial-mesenchymal transition, migration and
invasion by targeting SMAD1 in hepatocellular carcinoma.
Oncotarget. 7:24383–24401. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Wang D, Han S, Wang X, Peng R and Li X:
SOX5 promotes epithelial-mesenchymal transition and cell invasion
via regulation of Twist1 in hepatocellular carcinoma. Med Oncol.
32:4612015.PubMed/NCBI
|
|
44
|
Liu YR, Liang L, Zhao JM, Zhang Y, Zhang
M, Zhong WL, Zhang Q, Wei JJ, Li M, Yuan J, et al: Twist1 confers
multidrug resistance in colon cancer through upregulation of
ATP-binding cassette transporters. Oncotarget. 8:52901–52912. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Ávila-Moreno F, Armas-López L,
Álvarez-Moran AM, López-Bujanda Z, Ortiz-Quintero B,
Hidalgo-Miranda A, Urrea-Ramírez F, Rivera-Rosales RM,
Vázquez-Manríquez E, Peña-Mirabal E, et al: Overexpression of MEOX2
and TWIST1 is associated with H3K27me3 levels and determines lung
cancer chemoresistance and prognosis. PLoS One. 9:e1141042014.
View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Wang R, Li Y, Hou Y, Yang Q, Chen S, Wang
X, Wang Z, Yang Y, Chen C, Wang Z and Wu Q: The
PDGF-D/miR-106a/Twist1 pathway orchestrates epithelial-mesenchymal
transition in gemcitabine resistance hepatoma cells. Oncotarget.
6:7000–7010. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Xue F, Liu Y, Chu H, Wen Y, Yan L, Tang Q,
Xiao E, Zhang D and Zhang H: eIF5A2 is an alternative pathway for
cell proliferation in cetuximab-treated epithelial hepatocellular
carcinoma. Am J Transl Res. 8:4670–4681. 2016.PubMed/NCBI
|