Introduction
Cholesteatoma constitutes an acquired benign
epidermal non-permanent bone lesion that is locally destructive,
and patients often relapse (1). It
is characterized by squamous epithelium overgrowth with
hyper-keratin and bone erosion in the middle ear (1). Regardless of whether cholesteatomas
are congenital or acquired, cholesteatomas that are infected appear
to become enlarged more promptly and damage local structures, as
detected by clinical observation (2). Previous reports have stated that
infection may facilitate the aggressiveness of cholesteatoma, and
that the subsequent inflammatory response may be the most critical
factor in cholesteatoma-induced bone destruction (2–4).
The mucosal innate immune system actively
contributes to the host response to bacterial pathogen invasions
and cell stress signals via pathogen recognition involving
pathogen-associated molecular pattern recognition receptors (PRRs),
and the ensuing pro-inflammatory cytokine expression and production
(5–7). PRRs constitute several families, such
as Toll-like receptors (TLRs) (8)
and Nod-like receptors (NLRs) (9).
Notably, TLRs (2,10) and NLRs (11) are central for IL-1β release, which
plays a core function in inflammatory middle ear disease
pathogenesis; NLR family pyrin domain containing 3 (NLRP3)
(12) in particular is associated
with human middle ear cholesteatoma pathogenesis by modulating the
activity of IL-1β via the generation of inflammasomes.
Inflammasomes are a group of intracellular protein
complexes that identify multiple inflammation-triggering stimuli
and induce pro-inflammatory cytokine maturation, including the
maturation of IL-1β, as well as IL-18 (13). In the past decade, NLRP3 has been
reported to induce the generation of inflammasomes and be activated
in cholesteatoma (12). However, to
the best of our knowledge, little is currently known regarding its
function and whether other inflammasomes participate in
inflammatory reactions and the subsequent release of cytokines in
cholesteatoma. Besides NLRs, the hematopoietic IFN-inducible
nuclear proteins with a 200 amino acid repeat (HIN-200) family
member absent in melanoma 2 (AIM2) has previously been documented
to activate an inflammasome that is pivotal for inducing caspase-1
in multiple immune cells in response to cytoplasmic DNA (14–16).
AIM2 constitutes an IFN-inducible HIN-200 family
member with an amino-terminal pyrin domain (PYD) and a
carboxy-terminal HIN-200 oligonucleotide/oligosaccharide-binding
domain (15). Cytoplasmic DNA
recognition by the HIN-200 domain mediates the generation of the
AIM2 inflammasome by activating AIM2 oligomerization and
mobilization of apoptosis-associated speck-like protein containing
a CARD (ASC), which initiates caspase-1 and modulates the release
and bioactivity of the IL-1 family of cytokines (15,16).
In our previous study, elevated apoptotic cells and extracellular
DNA fragments were observed in the cholesteatoma epithelium
(17). Furthermore, AIM2 has been
demonstrated to regulate IL-1β production in human keratinocytes in
response to cytoplasmic double stranded (ds)DNA (18,19).
Therefore, the present study hypothesized that human cholesteatoma
epithelium may express AIM2, and that cholesteatoma keratinocytes
may release IL-1β by generating inflammasomes in response to
cytoplasmic DNA.
To test this hypothesis, the present study examined
the prospective expression of AIM2 in human cholesteatoma tissues,
and then investigated the mechanism underlying the AIM2
inflammasome-mediated secretion of IL-1β in cholesteatoma
keratinocytes. The present study aimed to provide a basis for
further evaluation into the prospective function of AIM2 in the
apoptosis of human cholesteatoma keratinocytes.
Materials and methods
Tissue samples
Cholesteatoma tissues were acquired from the medial
ear of nine subjects (six male and three female patients; age
range, 8–52 years; median age, 32 years) during tympanoplasty
between January 2018 and December 2018 in the Department of
Otolaryngology Head and Neck Surgery, Eye and ENT Hospital of Fudan
University (Shanghai, China). The inclusion criteria were as
follows: i) pathology consistent with cholesteatoma; ii) for
congenital cholesteatoma, intact tympanic membrane and no history
of otorrhea. Patients were divided into congenital (three cases;
age, 8–14 years) and acquired (six cases; age, 33–52 years) groups.
The subjects with congenital cholesteatoma were selected carefully
as per the criteria of Derlacki and Clemis (20). The cholesteatoma diagnosis was
confirmed by histopathological examination. Small biopsy-acquired
samples from the external canal skin of five individuals undergoing
tympanoplasty for chronic otitis media were utilized as the control
group. Signed informed consent was obtained from all participants;
if a patient was <18 years old, informed consent was obtained
from a parent or guardian. The study protocols were approved by the
Research Ethics Committee of the Eye and ENT Hospital of Fudan
University (approval no. 2017024).
Immunofluorescent staining for
cholesteatoma tissues
Fixation of the samples in 4% paraformaldehyde was
performed for 24 h at room temperature, before embedding in
paraffin. Serial segments were sliced (7-µm thick) and fixed on
3-aminopropyltriethoxysilane-coated glass slides.
Immunofluorescence was then performed in order to investigate the
expression levels of IL-18, ASC, AIM2, caspase-1 and IL-1β in
tissue sections. Briefly, blocking of the segments was performed
for 1 h in 10% normal goat serum (cat. no. ab7481; Abcam) after
deparaffinization and rehydration in graded alcohol series at room
temperature. After rinsing, segments were incubated overnight at
4°C with the working solution of antibodies against IL-1β (1:200;
cat. no. GTX100793), caspase-1 (1:200; cat. no. GTX101322), AIM2
(1:100; cat. no. GTX116487), ASC (1:200; cat. no. GTX102474) and
IL-18 (1:250; cat. no. GTX32675) (all from GeneTex, Inc.). The
segments were then incubated with Alexa 546-labeled (1:500; cat.
no. 715-025-150; Jackson ImmunoResearch) or Alexa 488-labeled
(1:500; cat. no. 111-545-144, Jackson ImmunoResearch) goat
anti-rabbit antibodies at room temperature for 1 h, and
subsequently stained with blue fluorescent DAPI (1:500; cat. no.
1217226; Invitrogen; Thermo Fisher Scientific, Inc.) at room
temperature for 15 min. Images were captured using the Axioscope
microscope (Carl Zeiss AG).
Cell culture
The isolation and characterization of cholesteatoma
keratinocytes was performed as previously described (15). Briefly, cholesteatoma tissue was
obtained and delivered to the laboratory following surgical
resection. Then, tissues were cut into small segments using
scissors, followed by 4°C overnight-digestion of the pieces using
collagenase IV (200 U/ml; Sigma-Aldrich; Merck KGaA). The digested
segments were rinsed twice using Hanks' Balanced Salt Solution, and
then centrifuged for 5 min at 350 × g at room temperature. The
supernatant was discarded, and 10 ml keratinocyte serum-free medium
(KSFM; Invitrogen; Thermo Fisher Scientific, Inc.) enriched with
500 U/ml penicillin/streptomycin (Invitrogen; Thermo Fisher
Scientific, Inc.) was added, followed by culture in a 5%
CO2 humidified atmosphere at 37°C. The KSFM with the
antibiotics was replaced every 3 days. The cell cultures between
the third and fourth passages were utilized in the present
study.
TUNEL assay
The in situ cell death detection kit (Roche
Diagnostics) was utilized in the TUNEL assay to examine DNA
fragmentation according to the manufacturer's instructions.
Briefly, the Lab-Tech chamber slide, acquired from Nalge Nunc
International (Thermo Fisher Scientific, Inc.), was used to culture
the cholesteatoma keratinocytes. After that, the cells were
incubated with 2 µg/ml poly(dA:dT; cat. no. tlrl-patn; InvivoGen)
for 24 h after priming with IFN-γ (25 ng/ml; cat. no. 300-02-20;
PeproTech) for 24 h, and then the medium was replaced. Another
group of cells was treated with only 2 µg/ml poly(dA:dT). After
stimulation, cells were fixed for 30 min using 4% paraformaldehyde
at room temperature, followed by permeabilization in PBS with 0.3%
Tween (PBST) for 5 min at room temperature. Subsequently, the cells
were rinsed using PBS three times, and the chamber slide and the
TUNEL reaction were mixed with terminal deoxynucleotidyl
transferase (TdT) and TMR-dUTP, and incubated in a humidified
environment for 1 h. The slides were then rinsed three times and
analyzed using an Axioscope microscope (Carl Zeiss AG). DAPI
(1:500) was used to perform the nuclear counterstaining at room
temperature for 15 min. The TUNEL ratio was calculated by assessing
five randomly selected fields of view (1:100).
Western blotting
Cholesteatoma keratinocytes were placed in a petri
plate (35-mm) at a density of 1×105 cells per plate.
Subsequently, varying titers of IFN-γ alone or poly(dA:dT) (2
µg/ml) after priming with IFN-γ (25 ng/ml) for 24 h were added.
After being stimulated for 24 h, the culture supernatants were
collected, and the cells were isolated using cell lysis buffer
(Beyotime Institute of Biotechnology) and prepared in order to
purify the total cellular protein. The protein concentration was
determined by BCA protein assay kit (Beyotime Institute of
Biotechnology). Equal amounts of total protein (30 µg) were
separated via SDS-PAGE (12% gel), and subsequently transferred onto
a PVDF membrane (Beyotime Institute of Biotechnology). The
membranes were washed with Tris-buffered saline-Tween-20 (20 mM
Tris, 500 mM NaCl and 0.1% Tween-20) and blocked at room
temperature for 1 h in western blocking buffer (Beyotime Institute
of Biotechnology). After that, the membrane-bound proteins were
conjugated with the indicated primary antibodies via overnight
incubation at 4°C. Subsequently, conjugation with the
peroxidase-labeled secondary antibodies (1:2,500; cat. no. GAR007;
Multi Sciences) was performed according to the manufacturer's
protocols. Anti-β-actin was used as the internal loading control.
Semi-quantification of the proteins was performed using ImageJ
software (version 1.63; National Institutes of Health). The
fold-changes were calculated as the ratios of band densities of
target protein to β-actin. The primary antibodies were as follows:
Anti-cleaved-caspase-1 (1:1,000; cat. no. GTX134551), anti-AIM2
(1:1,000; cat. no. GTX116487), anti-ASC (1:1,000; cat. no.
GTX102474), anti-caspase-1 (1:1,000; cat. no. GTX101322),
anti-gasdermin D (GSDMD; 1:500; cat. no. GTX116840), anti-IL-18
(1:1,000; cat. no. GTX32675), anti-IL-1β (1:1,000; cat. no.
GTX100793; all GeneTex, Inc.), anti-cleaved N-terminal GSDMD
(GSDMD-N; 1:1,000; cat. no. ab215203; Abcam) and anti-β-actin
(1:2,500; cat. no. AF0003; Beyotime Institute of
Biotechnology).
Statistical analysis
The data represent ≥3 independent assays, with each
experiment performed in triplicate, and are presented as the mean ±
standard deviation. One-way ANOVA followed by Bonferroni's multiple
comparison test were used for the statistical analysis. P<0.05
was considered to indicate a statistically significant
difference.
Results
Human cholesteatoma tissues express
AIM2
In order to investigate whether AIM2 is expressed by
human cholesteatoma tissues, the present study assessed the
expression of the AIM2 protein in cholesteatoma tissues using both
immunofluorescence and western blotting. AIM2 protein was expressed
in the cytoplasm of cells, primarily in granular cells, as well as
the prickle-cell sheets in the cholesteatoma epithelium (Fig. 1A). Moreover, the AIM2 expression
intensity was generally decreased in the basal and parabasal layer
(Fig. 1A). However, the western
blotting data revealed that the expression of AIM2 varied in these
specimens (Fig. 1B and C). The AIM2
content was significantly elevated in acquired cholesteatoma
compared with the congenital cholesteatoma (Fig. 1D).
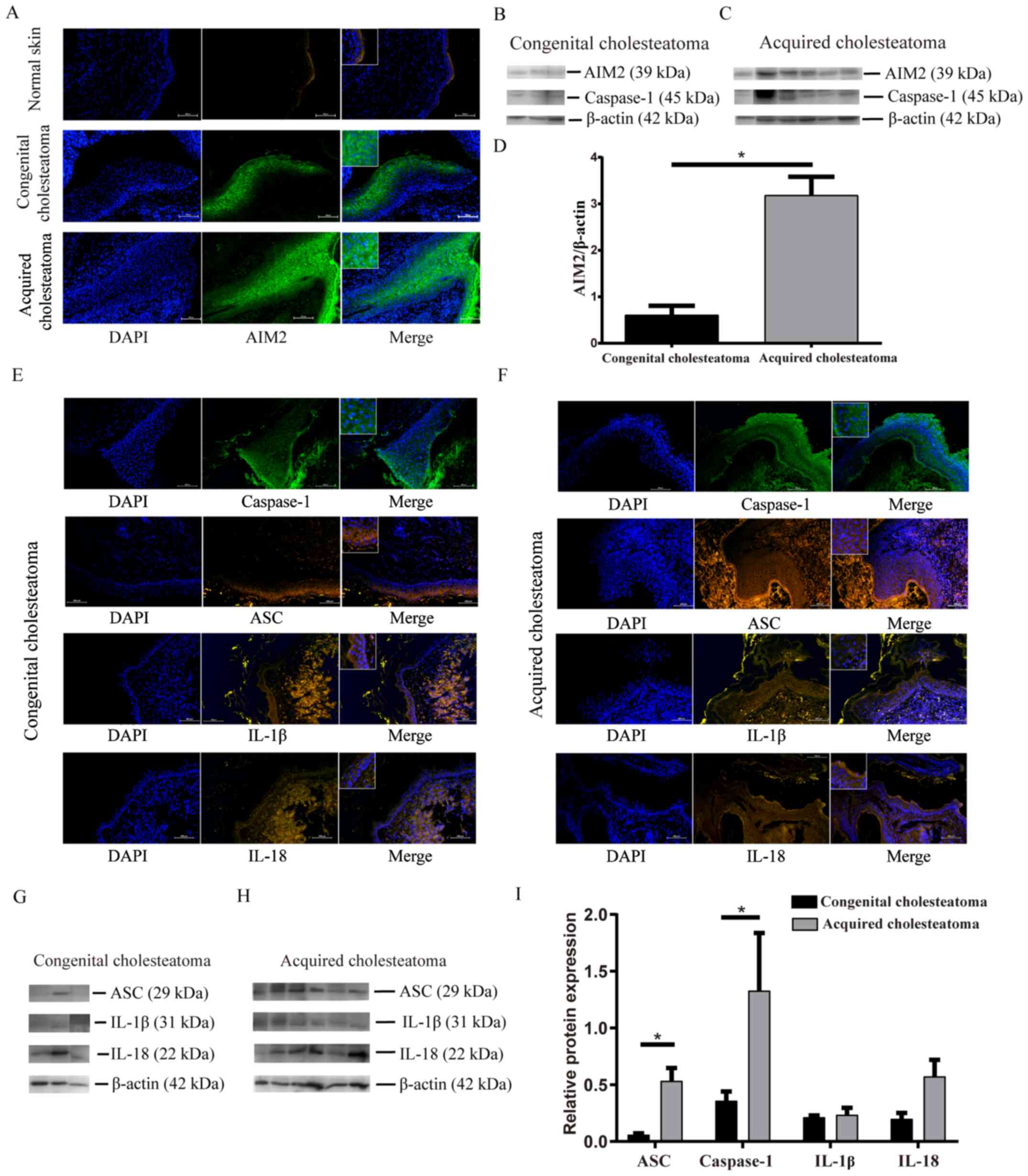 | Figure 1.Immunofluorescence and western
blotting analyses of inflammasome components in human congenital
and acquired cholesteatoma tissues. (A) Localization and expression
of AIM2 in cholesteatoma tissues and healthy skin tissues.
Immunofluorescence analysis revealed that AIM2 was localized in the
cytoplasm of cells, primarily in granular and prickle-cell sheets.
The nucleus was labelled using DAPI (magnification, ×200). (B and
C) Western blotting images of AIM2 and caspase-1. (D) Corresponding
semi-quantitative analysis of the AIM2 protein content in
congenital and acquired cholesteatoma tissues. Localization of ASC,
caspase-1, IL-1β and IL-18 in (E) congenital cholesteatoma tissues
and (F) acquired cholesteatoma tissues. The immunofluorescence
assay revealed that ASC and caspase-1 were mainly localized in the
cytoplasm of cells, primarily in granular and prickle-cell layers.
The nucleus was labelled using DAPI (magnification, ×200). (G and
H) Western blotting images and (I) corresponding semi-quantitative
analysis of the ASC, caspase-1, IL-1β and IL-18 protein expression
levels in congenital and acquired cholesteatoma tissues. The
relative protein expression content was standardized to β-actin.
The columns indicate the mean ± standard deviation values (n=3 per
group). *P<0.05. ASC, apoptosis-associated speck-like protein
containing a CARD; AIM2, absent in melanoma 2. |
Protein expression of inflammasome
constituents is increased in cholesteatoma tissues
As common components of the inflammasome, the
expression levels of ASC and caspase-1 proteins were similarly
higher in human cholesteatoma tissues. ASC and caspase-1 were
localized in the cytoplasm of cells, primarily in granular and
prickle-cell sheets in the cholesteatoma epithelium (Fig. 1E and F). The western blotting
results revealed that the ASC and caspase-1 protein expression
levels in acquired cholesteatoma (Fig.
1C and H) were markedly higher than those in congenital
cholesteatoma (Fig. 1B and G). As
shown in Fig. 1I, these differences
were statistically significant. The present study also observed
markedly higher IL-1β and IL-18 protein expression levels in
acquired cholesteatoma tissues (Fig. 1G
and H). However, the concentrations of IL-1β and IL-18 did not
differ significantly between the acquired and congenital
cholesteatoma (Fig. 1I). These data
suggested that there was enhanced inflammasome activation in the
cholesteatoma epithelium.
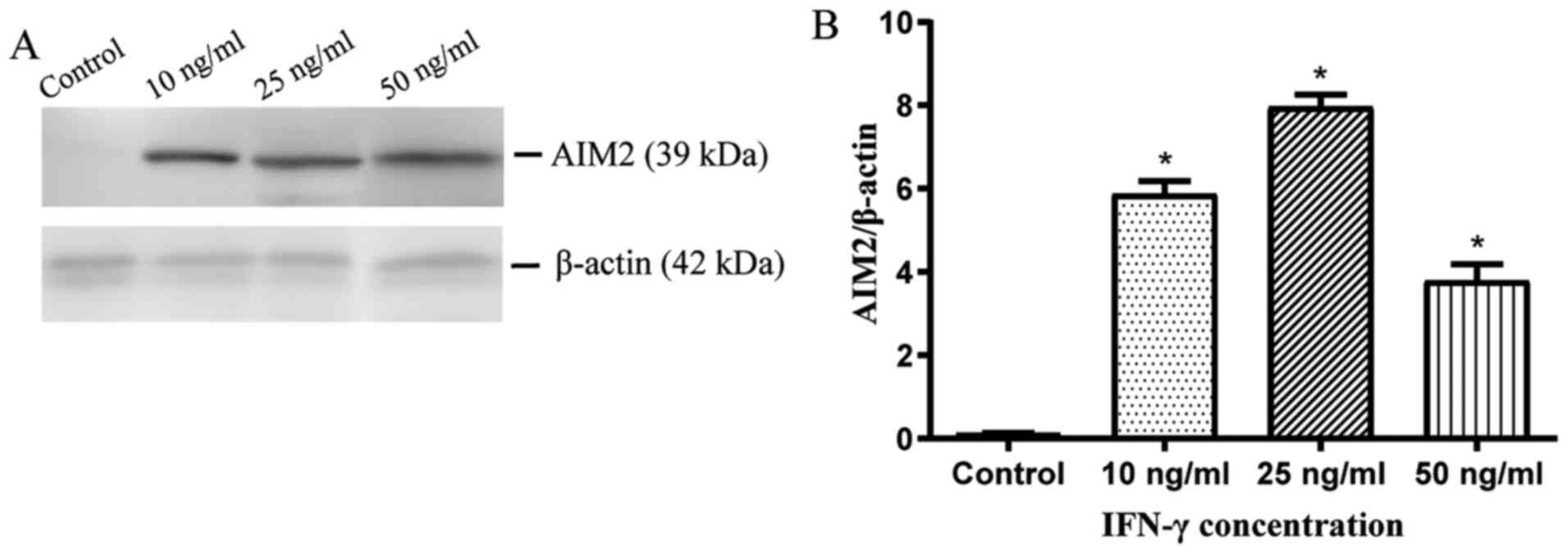
Expression of AIM2 in cholesteatoma
keratinocytes is strongly induced by IFN-γ
In order to further investigate the effect of IFN-γ
on the expression of AIM2 in cholesteatoma keratinocytes, varied
titers of IFN-γ were introduced to cholesteatoma keratinocytes for
24 h, and then the AIM2 protein expression levels were assessed via
western blotting. Untreated cholesteatoma keratinocytes expressed
low levels of AIM2 protein. At the same time, it was observed that
AIM2 protein expression was increased following IFN-γ
administration (Fig. 2A and B).
Based on these data, the treatment of cholesteatoma keratinocytes
with 25 ng/ml IFN-γ for 24 h was established as the optimum
concentration for the downstream assays. Collectively, these
findings implied that IFN-γ induced and upregulated AIM2 expression
in cholesteatoma keratinocytes.
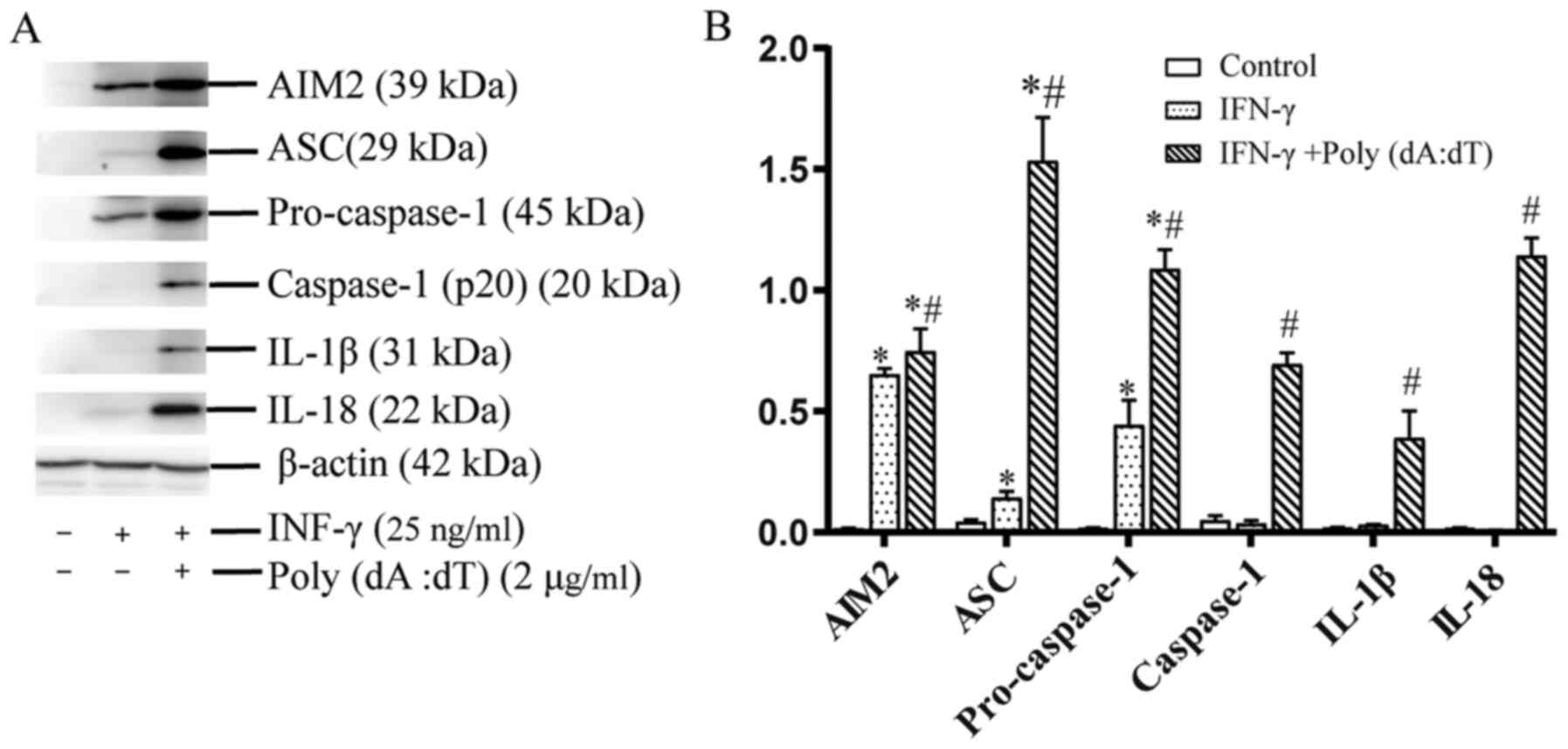
Effect of induction of IL-1β and IL-18
release on activation of the AIM2 inflammasome in cholesteatoma
keratinocytes in response to cytoplasmic DNA
A previous study documented a marked increase in
AIM2 expression following induction with IFN-γ (21). Similarly, induction with IFN-γ (25
ng/ml) or poly(dA:dT) after priming with IFN-γ (25 ng/ml) for 24 h
upregulated the protein expression levels of pro-caspase-1 and ASC,
which are both constituents of AIM2 inflammasomes (Fig. 3). However, the present study also
established that IFN-γ actually primed inflammasomes rather than
activated them, since the protein expression of the inflammasome
constituents was markedly raised in human cholesteatoma
keratinocytes, but without any alterations in cleaved-caspase-1,
IL-1β or IL-18 titers (Fig. 3A and
B). Hence, an AIM2 ligand, poly(dA:dT), was utilized to elevate
AIM2 inflammasome activation in cholesteatoma keratinocytes.
Treatment with poly(dA:dT) activated the protein contents of
cleaved-caspase-1, and the production of IL-1β and IL-18 in
IFN-γ-primed cholesteatoma keratinocytes, relative to the untreated
IFN-γ-primed cells (Fig. 3A and B).
These data demonstrated that poly(dA:dT) induced IL-1β and IL-18
production via the activation of caspase-1-dependent AIM2
inflammasomes in cholesteatoma keratinocytes.
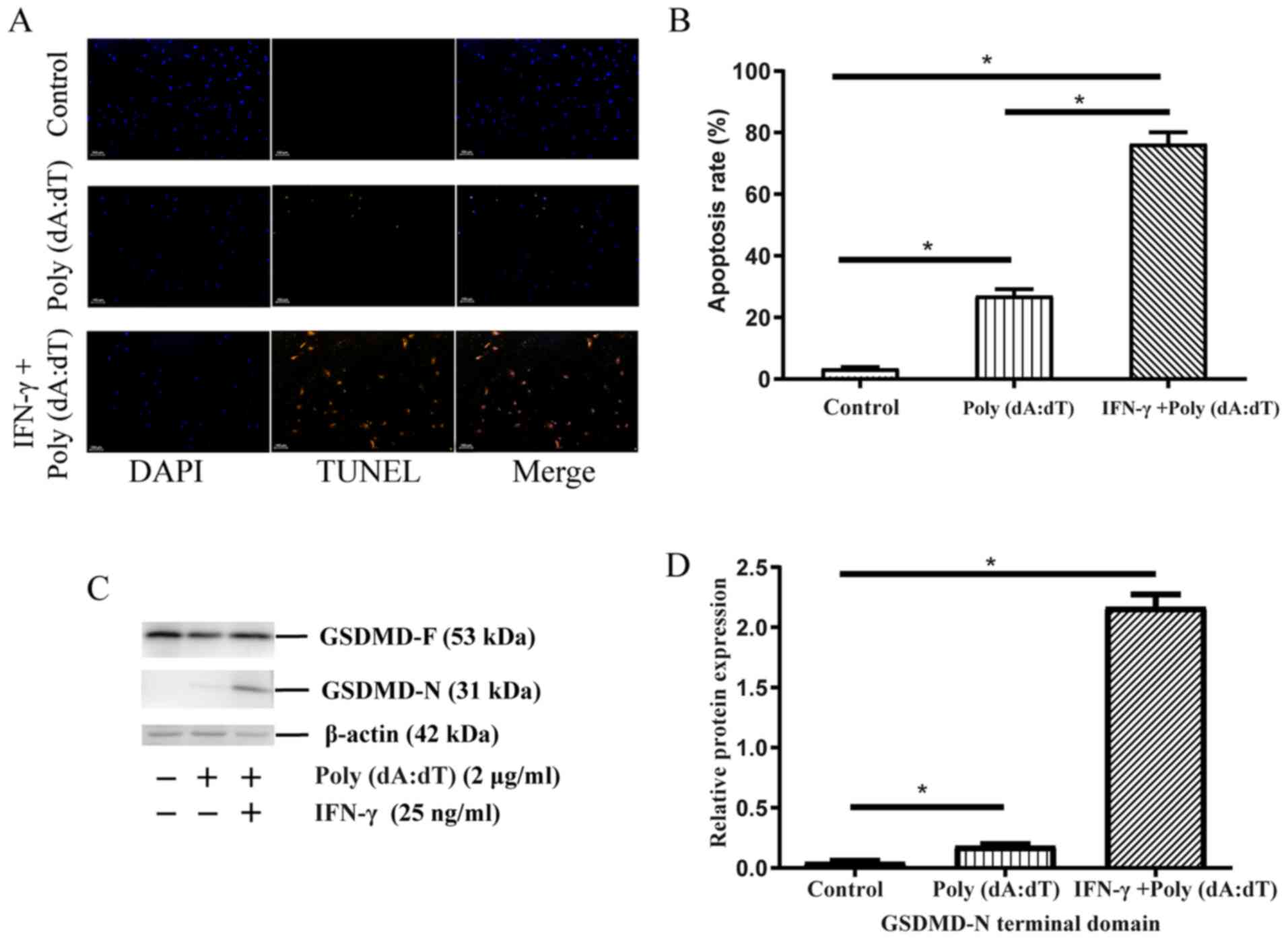
Pyroptosis of cholesteatoma
keratinocytes on the activation of the AIM2 inflammasome
Pyroptosis is regarded as a pivotal underlying
mechanism of active cytokine production caused by alterations in
membrane permeability (22,23). As indicated in Fig. 4A and B, the results of the TUNEL
assay demonstrated an increased percentage of apoptotic cells in
the poly(dA:dT) and IFN-γ + poly(dA:dT) groups relative to the
control. The results in Fig. 4C and
D also demonstrated that there was an increase in the
expression levels of GSDMD-N (31 kDa) in the poly(dA:dT) and IFN-γ
+ poly(dA:dT) groups when compared with the blank controls.
Discussion
To the best of our knowledge, the present study was
the first to demonstrate the expression patterns of AIM2
inflammasome in cholesteatoma tissues. As the cytoplasmic dsDNA
sensor (15,16) and the first non-NLR family
inflammasome, AIM2 has been reported to be present primarily in
peripheral monocytes, and is also a significant node in the innate
immune system (24). The AIM2
inflammasome can also be expressed and activated in certain tissue
cells, such as keratinocytes (18,25),
neurons (26) and islet-β cells
(27). It was revealed in the
present study that AIM2, ASC, caspase-1, IL-1β and IL-18 protein
expression levels were notably increased in cholesteatoma tissues
compared with healthy skin. These data suggested that the AIM2
inflammasome cascade may be activated in cholesteatoma tissues.
Collectively, the present study provided evidence
that AIM2 is expressed in human cholesteatoma tissues and cultured
cholesteatoma keratinocytes. Cholesteatomas are often categorized
as congenital or acquired. Congenital cholesteatoma is referred to
as the presence of an epithelial sac behind an intact tympanic
membrane without a history of otitis media (28). Acquired cholesteatoma is often
accompanied by chronic bacterial infection (1). The present study demonstrated that
AIM2 was expressed at higher levels in cholesteatoma compared with
that in external auditory canal skin. However, increased AIM2
expression levels were reported in acquired cholesteatoma tissues
compared with congenital cholesteatoma tissues. As previous studies
revealed that IFN-γ is a strong inducer of AIM2 (25), the present study investigated the
effect of IFN-γ on the expression of AIM2 in cholesteatoma
keratinocytes. Low basal titers of AIM2 protein were detected in
cultured cholesteatoma keratinocytes, which were increased
following IFN-γ stimulation. Thus, a potential reason for the
higher AIM2 expression levels in acquired cholesteatoma could be
that the pro-inflammatory cytokines secreted by the inflammatory
cells in the pre-matrix of acquired cholesteatoma tissues may
influence the expression of AIM2.
In the present study, the protein expression levels
of IL-1β and IL-18 were not statistically significantly different
between congenital and acquired cholesteatoma. One possible reason
is that IL-1β and IL-18 were secreted into the extracellular matrix
and were then degraded. Another possible reason is that the sample
size was not large enough to be significant. Of course, there are
other mechanisms that can induce the production of IL-1β and IL-18,
such as bacterial or viral infection. However, the purpose of
investigating cholesteatoma tissues was to confirm whether the
target proteins, IL-1β and IL-18, which are regulated by AIM2, were
expressed in cholesteatoma tissues.
Cytoplasmic DNA has been recognized as a danger
signal that can be identified using AIM2 as a DNA sensor. For
example, cytoplasmic dsDNA induces the initiation of the AIM2
inflammasome that leads to the production of IL-1β in human
keratinocytes (25). In the present
study, AIM2, ASC and caspase-1 expression levels increased
following induction with poly(dA:dT), implying that AIM2 serves as
a DNA sensor in the inflammatory response in cholesteatoma. A
previous study reported that numerous extracellular DNA fragments
were observed in the cholesteatoma epithelium (18). These findings suggested that
cholesteatoma keratinocytes inherently express AIM2 and facilitate
the activation of AIM2 inflammasomes in response to host-derived
cytoplasmic DNA. Therefore, despite the upregulation of AIM2,
caspase-1 and ASC, the expression levels of IL-18,
cleaved-caspase-1 and IL-1β were increased following treatment with
cytoplasmic DNA after priming with IFN-γ. These data demonstrated
that dsDNA-triggered IL-1β and IL-18 secretion in cholesteatoma
keratinocytes was mediated by the activated AIM2 inflammasome,
which could be a pivotal mediator of inflammation in
cholesteatoma.
The present study observed an increased percentage
of TUNEL-positive cells, increased expression of GSDMD-N, and
increased IL-1β and IL-18 production following stimulation of
cholesteatoma keratinocytes with poly(dA:dT). These data indicated
the loss of cell membrane integrity and increased cell pyroptosis
of cholesteatoma keratinocytes. Furthermore, IFN-γ was documented
to markedly enhance poly(dA:dT)-triggered pyroptosis. Pyroptosis is
a highly distinct type of inflammatory programmed cell death that
is different from necrosis or apoptosis (22). Pyroptosis results in plasma membrane
rupture and the secretion of damage-associated molecular pattern
(DAMPs) biomolecules into the extracellular space (23). Thus, cell pyroptosis has been
documented to lead to inflammation, chemotaxis of immune cells and
further perpetuate the inflammatory cascade through release of
various DAMPs, as well as pro-inflammatory mediators, consisting of
IL-1β, IL-18 and the alarmin high mobility group box 1 (22,23).
Also named IFN-γ-inducing factor, IL-18 induces IFN-γ secretion
(29). In addition, previous
studies demonstrated that IL-1 may serve a vital role in the
pathogenesis of acquired and congenital cholesteatomas by
modulating bone resorption and cell proliferation (1,10,17).
Thus, the present study hypothesized that AIM2
inflammasome-mediated maturation and release of IL-1β and IL-18
from cholesteatoma keratinocytes both contribute to the immune
response in cholesteatoma tissues.
In summary, the present study reported increased
expression levels of inflammasome constituents (caspase-1, AIM2 and
ASC) and their downstream cytokines (IL-18 and IL-1β) in
cholesteatoma tissues; these findings indicated the redundant
activation of inflammasomes in cholesteatoma tissue. In addition,
the results of the present study demonstrated that poly(dA:dT)
induced AIM2 inflammasome activation in cholesteatoma
keratinocytes, which increased cell pyroptosis and the production
of IL-1β and IL-18. Pro-inflammatory cytokines (such as IFN-γ)
could enhance the expression levels of AIM2 inflammasome
constituents and promote poly(dA:dT)-induced cell death and
bioactive cytokine production. These events demonstrated that the
persistent expression of extracellular DNA fragments and the
subsequent expression of AIM2 in cholesteatoma led to cell
pyroptosis, as well as the production of inflammatory cytokines,
which could cause bone resorption and perpetuate the inflammatory
cascade in cholesteatoma.
Acknowledgements
The authors would like to thank Professor Shufeng Li
(ENT Institute and Department of Otolaryngology, Eye and ENT
Hospital, Fudan University, China) for helpful instruction and
comments on the manuscript.
Funding
This work was funded by Science Project of Shanghai
Municipal Health Bureau (grant no. 20174Y0062).
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
CZ and MC performed the experiments. ZC and CZ wrote
and revised the manuscript. CZ and MC confirm the authenticity of
all the raw data. CZ and ZC performed the statistical analysis. MC
conducted the pathological diagnosis. All authors read and approved
the final manuscript.
Ethics approval and consent to
participate
Signed informed consent was obtained from all
participants, if a patient was <18 years old, informed consent
was obtained from a parent or guardian. Human studies conformed to
the principles of the Declaration of Helsinki and were approved by
the Research Ethics Committee of the Eye and ENT Hospital of Fudan
University (approval no. 2017024).
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Olszewska E, Wagner M, Bernal-Sprekelsen
M, Ebmeyer J, Dazert S, Hildmann H and Sudhoff H: Etiopathogenesis
of cholesteatoma. Eur Arch Otorhinolaryngol. 261:6–24. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Si Y, Chen YB, Chen SJ, Zheng YQ, Liu X,
Liu Y, Jiang HL, Xu G, Li ZH, Huang QH, et al: TLR4 drives the
pathogenesis of acquired cholesteatoma by promoting local
inflammation and bone destruction. Sci Rep. 5:166832015. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Inagaki T and Paparella MM: Chronic otitis
media with cholesteatoma: Middle ear/inner ear interaction. Otol
Neurotol. 30:430–431. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Grundfast KM, Ahuja GS, Parisier SC and
Culver SM: Delayed diagnosis and fate of congenital cholesteatoma
(keratoma). Arch Otolaryngol Head Neck Surg. 121:903–907. 1995.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Tlaskalová-Hogenová H, Tucková L,
Lodinová-Zádniková R, Stepánková R, Cukrowska B, Funda DP, Striz I,
Kozáková H, Trebichavský I, Sokol D, et al: Mucosal immunity: Its
role in defense and allergy. Int Arch Allergy Immunol. 128:77–89.
2002. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Chang JH, McCluskey PJ and Wakefield D:
Toll-like receptors in ocular immunity and the immunopathogenesis
of inflammatory eye disease. Br J Ophthalmol. 90:103–108. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Kumar A, Hazlett LD and Yu FS: Flagellin
suppresses the inflammatory response and enhances bacterial
clearance in a murine model of Pseudomonas aeruginosa keratitis.
Infect Immun. 76:89–96. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Kawai T and Akira S: TLR signaling. Cell
Death Differ. 13:816–825. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Lee MS and Kim YJ: Pattern-recognition
receptor signaling initiated from extracellular, membrane, and
cytoplasmic space. Mol Cells. 23:1–10. 2007.PubMed/NCBI
|
|
10
|
Jiang H, Si Y, Li Z, Huang X, Chen S,
Zheng Y, Xu G, Chen X, Chen Y, Liu Y, et al: TREM-2 promotes
acquired cholesteatoma-induced bone destruction by modulating TLR4
signaling pathway and osteoclasts activation. Sci Rep. 6:387612016.
View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Leichtle A, Klenke C, Ebmeyer J, Daerr M,
Bruchhage KL, Hoffmann AS, Ryan AF, Wollenberg B and Sudhoff H:
NOD-Like Receptor Signaling in Cholesteatoma. BioMed Res Int.
2015:4081692015. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Kariya S, Okano M, Zhao P, Kataoka Y,
Yoshinobu J, Maeda Y, Ishihara H, Higaki T and Nishizaki K:
Activation of NLRP3 inflammasome in human middle ear cholesteatoma
and chronic otitis media. Acta Otolaryngol. 136:136–140. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Strowig T, Henao-Mejia J, Elinav E and
Flavell R: Inflammasomes in health and disease. Nature.
481:278–286. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Lamkanfi M and Dixit VM: Inflammasomes and
their roles in health and disease. Annu Rev Cell Dev Biol.
28:137–161. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Fernandes-Alnemri T, Yu JW, Datta P, Wu J
and Alnemri ES: AIM2 activates the inflammasome and cell death in
response to cytoplasmic DNA. Nature. 458:509–513. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Hornung V, Ablasser A, Charrel-Dennis M,
Bauernfeind F, Horvath G, Caffrey DR, Latz E and Fitzgerald KA:
AIM2 recognizes cytosolic dsDNA and forms a caspase-1-activating
inflammasome with ASC. Nature. 458:514–518. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Chi Z, Wang Z, Liang Q, Zhu Y and du Q:
Induction of cytokine production in cholesteatoma keratinocytes by
extracellular high-mobility group box chromosomal protein 1
combined with DNA released by apoptotic cholesteatoma
keratinocytes. Mol Cell Biochem. 400:189–200. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Kopfnagel V, Wittmann M and Werfel T:
Human keratinocytes express AIM2 and respond to dsDNA with IL-1β
secretion. Exp Dermatol. 20:1027–1029. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
de Koning HD, Bergboer JG, van den Bogaard
EH, van Vlijmen-Willems IM, Rodijk-Olthuis D, Simon A, Zeeuwen PL
and Schalkwijk J: Strong induction of AIM2 expression in human
epidermis in acute and chronic inflammatory skin conditions. Exp
Dermatol. 21:961–964. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Derlacki EL and Clemis JD: Congenital
cholesteatoma of the middle ear and mastoid. Ann Otol Rhinol
Laryngol. 74:706–727. 1965. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Huang S, Song Z, Jiang L, Chen L, Wang R,
Qin W, Liu P and Lin Z: Absent in melanoma 2 (AIM2) expressed in
human dental pulp mediates IL-1β secretion in response to
cytoplasmic DNA. Inflammation. 38:566–575. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Xu YJ, Zheng L, Hu YW and Wang Q:
Pyroptosis and its relationship to atherosclerosis. Clin Chim Acta.
476:28–37. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Zhang Y, Chen X, Gueydan C and Han J:
Plasma membrane changes during programmed cell deaths. Cell Res.
28:9–21. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Guo Q, Wu Y, Hou Y, Liu Y, Liu T, Zhang H,
Fan C, Guan H, Li Y, Shan Z, et al: Cytokine secretion and
pyroptosis of thyroid follicular cells mediated by enhanced NLRP3,
NLRP1, NLRC4, and AIM2 inflammasomes are associated with autoimmune
thyroiditis. Front Immuno. 9:11972018. View Article : Google Scholar
|
|
25
|
Dombrowski Y, Peric M, Koglin S,
Kammerbauer C, Göss C, Anz D, Simanski M, Gläser R, Harder J,
Hornung V, et al: Cytosolic DNA triggers inflammasome activation in
keratinocytes in psoriatic lesions. Sci Transl Med. 3:82ra382011.
View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Kaushal V, Dye R, Pakavathkumar P, Foveau
B, Flores J, Hyman B, Ghetti B, Koller BH and LeBlanc AC: Neuronal
NLRP1 inflammasome activation of Caspase-1 coordinately regulates
inflammatory interleukin-1-beta production and axonal
degeneration-associated Caspase-6 activation. Cell Death Differ.
22:1676–1686. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Oslowski CM, Hara T, O'Sullivan-Murphy B,
Kanekura K, Lu S, Hara M, Ishigaki S, Zhu LJ, Hayashi E, Hui ST, et
al: Thioredoxin-interacting protein mediates ER stress-induced β
cell death through initiation of the inflammasome. Cell Metab.
16:265–273. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Bhutta MF, Williamson IG and Sudhoff HH:
Cholesteatoma. BMJ. 342:d10882011. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Novick D, Kim S, Kaplanski G and Dinarello
CA: Interleukin-18, more than a Th1 cytokine. Semin Immunol.
25:439–448. 2013. View Article : Google Scholar : PubMed/NCBI
|