Introduction
Asthma is one of the most common chronic respiratory
diseases, affecting ~300 million individuals worldwide, and the
prevalence is increasing (1).
Asthma is characterized by recurrent coughing, wheezing and chest
tightness, and affects the quality of life, bringing with it heavy
social and economic burdens to patients, their families and to
healthcare systems (2). Although
asthma is considered a heterogeneous disease, it has classically
been considered as a T helper (Th)2 cell allergic disease, with
increased infiltration of eosinophils and enhanced immunoglobulin
(Ig)E levels. Immune dysfunction, including an imbalance in
Th1/Th2, has been reported to be involved in the pathogenesis of
the allergic response observed in asthma for several decades
(3,4). The Th2-related cytokines interleukin
(IL)-4, IL-5 and IL-13 are essential in eosinophil accumulation,
airway hyper-responsiveness (AHR), mucus production and further
bronchial fibrosis (5,6). Th1 produces interferon (IFN)-γ, which
serves an important role in Th1 differentiation. Naïve
CD4+ T cells have the potential to differentiate into
different T subtypes, regulated by the type and strength of stimuli
and different transcription factors (7). Naïve CD4+ T cells are
activated by IL-4-STAT6 signaling, resulting in upregulation of the
transcription factor trans-acting T cell-specific
transcription factor GATA-3 (GATA3). GATA3 is crucial for Th2
differentiation and Th2 cytokine production (8); the Th2 locus control region has also
been reported to regulate Th2 cytokine genes (9). T cells possess the capacity to produce
IFN-γ, and this is regulated by several transcription factors,
including T-box transcription factor TBX21 (T-bet), eomesodermin
and runt-related transcription factor 3 (Runx3) (10).
Runx3 is a member of the Runt domain family of
transcription factors; it is expressed in the peripheral blood and
immune system and is involved in the development of T cell
differentiation (11,12). Previous studies have reported Runx3
is associated with the pathogenesis of asthma; the expression of
Runx3 in Th1 cells is higher than that in Th2 cells, and Runx3
knockout mice develop eosinophilic lung inflammation and AHR
spontaneously, suggesting that Runx3 is important in the repression
of Th2 responses (13,14). The human Runx3 gene is located on
chromosome 1p36 and contains two promoters P1 and P2. P2 is rich in
CpG dinucleotides, and hypermethylation of the CpG island adjacent
to Runx3 P2 promoter can result in the development of several
diseases, including leukemia and gastric cancer (15–17).
Men et al (18) reported
that methylation of the CpG islands in the Runx3 gene promoter
leads to silencing of Runx3 expression, and this underlies the
progression of bronchiolitis into asthma in Chinese children. It
has previously been suggested that Runx3 and GATA3 co-regulate each
other, and the relative expression of GATA3 and Runx3 may regulate
Th1/Th2 responses (19).
To date, >100 asthma-associated gene variants
have been reported to be involved in the occurrence and pathology
of asthma. However, hereditary factors explain only ~10% of the
risk of developing asthma (20–22).
Asthma is affected by both heredity and environmental exposure.
Owing to rapid industrialization, urbanization and population
growth, atmospheric pollution is now considered a major risk factor
for the development of asthma (1).
Particulate matter (PM) composition is the major component of air
pollution. PM is divided into three main groupings: PM10, coarse
(diameter, 2.5–10 µm); PM2.5, fine (diameter, <2.5 µm); and PM1,
ultrafine (diameter, <1 µm) (23). PM2.5 is small enough to penetrate
terminal bronchioles and alveoli, and previous studies have
indicated that PM2.5 is the most relevant with regards to
asthma-related environmental factors (24). PM2.5 has been reported to exacerbate
airway inflammation by upregulating the expression of transient
receptor potential cation channel subfamily A member 1 and
microRNAs (25), promoting reactive
oxygen species accumulation (26),
and activating toll-like receptor (TLR)2/TLR4/myeloid
differentiation primary response protein M2yD88 and NF-κB signaling
pathways in a murine model of asthma (27,28).
PM2.5 has also been reported to induce autophagy of human bronchial
epithelium cells via nitric oxide synthase inducible signaling
in vitro (29), as well as
disturb the balance between Th1/Th2 and Th17/regulatory T cells
(30,31). However, the precise mechanism by
which PM2.5 induces/aggravates asthma in patients remains
undetermined. Our previous study demonstrated that ambient PM2.5
exposure aggravated airway inflammation of asthmatic mice in a
dose-dependent manner, and a high dose of PM2.5 exposure induced
mixed eosinophilic/neutrophilic inflammation (32). However, to the best of our
knowledge, there are no studies that have assessed the expression
of related transcription factors, including GATA-3, T-bet and
Runx3, induced by PM2.5. Combined with our previous study, it was
hypothesized that PM2.5 aggravates airway inflammation and induces
an imbalance of Th1/Th2 cells by interfering with the expression of
the transcription factor GATA3, RUNX3 and T-bet. Therefore, the
present study aimed to examine the effects of 2.5 µm particulate
matter (PM2.5) on airway inflammation and to investigate the
possible underlying mechanism.
Materials and methods
Collection and measurement of
PM2.5
PM2.5 was collected using a 2033B high-volume air
sampler (Qingdao Laoying Environmental Technology Co., Ltd.) at a
flow rate of 100 l/min, 8 h/day for 12 months (between January and
December 2018) in Yantai, Shandong, China. After sampling, the
fiberglass filters were removed and cut into small pieces of 1×3
cm, the fibers were immersed in ultrapure water, ultrasonic
vibrations were used to elute the PM2.5, which was concentrated by
freeze-drying. The PM was mixed with saline to obtain a particle
suspension prior to experimentation. The concentration of
polycyclic aromatic hydrocarbons (PAHs) and metals in the samples
were determined using high-performance liquid chromatography (HPLC;
Hitachi Model 600 HPLC; Hitachi, Ltd.) and inductively coupled
plasma atomic emission spectrometry (ICP-AES; 61E Trace and
ICP-750; Thermo Jarrell-Ash) according to the manufacturer's
protocol. HPLC was conducted at room temperature. PM2.5 samples
(100 µl) were mixed with the same volume of
Na2SO4, then added into 10 ml normal
hexane/acetone (1:1). The samples were extracted by ultrasonication
(20 kHz) for 20 min at room temperature and moved into the
purification column (400×10 mm; cat. no. V0301050001; Shanghai
Chubo Laboratory Equipment Co., Ltd.) at 1.0 ml/min. Ultrasonic
extraction was performed for 5 min. PAHs were determined at 254 nm
using PAH standards (cat. no. PAH525-1JM; Chemservice). ICP-AES was
conducted at room temperature, the samples were added nitric acid
to limit the acidity to 5%, the wavelength range was 165–872 nm,
the power was 1100 W, and the analysis line and detection limits of
each metal was as follows: Ca (317.933 nm; 0.003 mg/l), Fe (328.204
nm; 0.000 mg/l), Mn (257.610 nm; 0.000 mg/l), Zn (213.867 nm; 0.015
mg/l), K (766.490 nm; 0.12 mg/l), Ba (455.403 nm; 0.000 mg/l), Na
(589.592 nm; 0.018 mg/l) and As (460.733 nm; 0.008 mg/l).
Animals and experimental design
A total of 48 6-week-old male BALB/c mice (8–22 g in
weight) were obtained from Ailingfei Animals Center (Nanjing,
China). The animal experiments were approved by the Institutional
Animal Care and Use Committee of Nanjing Medical University
(approval no. 20110217). Animals were anesthetized to minimize
suffering. The mice were maintained under a 12 h light/dark cycle
at a constant temperature (22±2°C) and a relative humidity of
55±10%; all animals had ad libitum access to food and water.
The mice were randomly divided into four groups (n=12/group) as
follows: i) Control group, ii) ovalbumin (OVA) group, iii) PM2.5
group and iv) OVA + PM2.5 group. A 26-day model was used based on
our previous study (33). On days
0, 7 and 14, mice in the OVA and OVA + PM2.5 groups were
intraperitoneally (i.p.) injected with 100 µg OVA (cat. no. A5503;
Sigma-Aldrich; Merck KGaA), which was combined with an alum
adjuvant (0.2 ml; cat. no. 77161; Thermo Fisher Scientific, Inc.);
mice in the control and PM2.5 groups were administered an
equivalent volume of PBS (i.p.). A total of 100 µg PM2.5 particles
in 50 µl physiological saline was administered intranasally to mice
in the PM2.5 and OVA + PM2.5 groups from days 16–20, an equivalent
volume of PBS was administered intranasally to mice in the control
and OVA group. Mice, except for those in the control group, were
challenged with aerosolized 1% OVA (total volume, 100 ml) for 30
min from days 21–25. The detailed protocols are shown in Fig. 1.
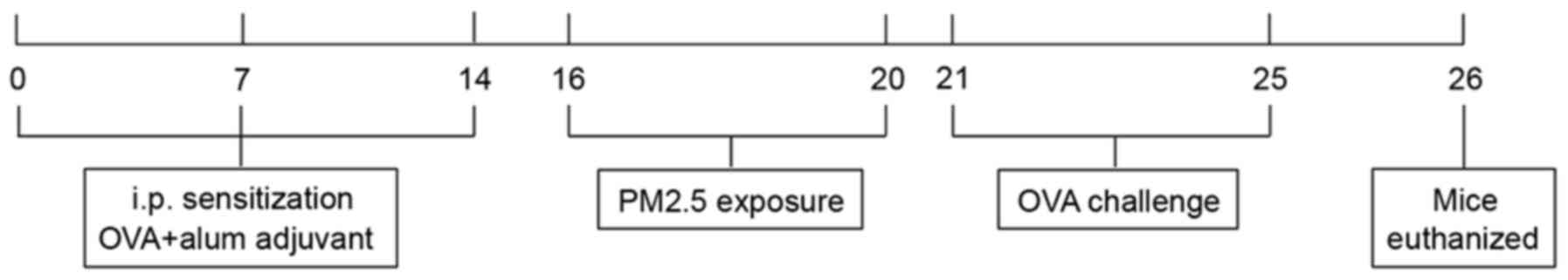 | Figure 1.Overview of the experimental design.
On days 0, 7 and 14 mice in the OVA and OVA + PM2.5 groups were
i.p. injected with 100 µg OVA and alum adjuvant, whereas mice in
the control and PM2.5 groups were administered an equivalent volume
of PBS (i.p.). A total of 100 µg PM2.5 particles in 50 µl
physiological saline was administered intranasally to mice in the
PM2.5 and OVA + PM2.5 groups from days 16–20, an equivalent volume
of PBS was administered intranasally to mice in the control and OVA
group. Mice, except for those in the control group, were challenged
with aerosolized 1% OVA (total volume, 100 ml) for 30 min from days
21–25. OVA, ovalbumin; PM2.5, 2.5 µm particulate matter; i.p.,
intraperitoneally; PBS, phosphate-buffer saline. |
Airway responsiveness
determination
Airway responsiveness to acetylcholine chloride
(ACH) was assessed 24 h after the final OVA challenge using an
AniRes 2005 animal lung function analysis system (Best Lab
Solutions). After anesthetizing mice with sodium pentobarbital (70
mg/kg) via intraperitoneal injection, mice were intubated through
the trachea for mechanical ventilation and the caudal vein for ACH
administration. The mice were ventilated to measure airway
resistance using progressively increasing doses of ACH (0, 10, 30,
90 and 180 µg/kg). All mice were ventilated at a breath rate set at
90 breaths/min and with the volume set as 6 ml/kg (34).
Determination of serum OVA-specific
IgE and cytokines in bronchoalveolar lavage fluid (BALF)
After sacrificing mice by cervical dislocation,
blood was collected from the heart. The blood samples were
centrifuged at 3,000 × g for 10 min at 4°C to obtain the serum. The
serum levels of OVA-specific IgE were assessed using an ELISA kit
(cat. no. ab157718; Abcam). The mice were subsequently intubated,
the left main bronchus was ligated and the airway lumina of the
right lungs were washed three times (0.4, 0.3 and 0.3 ml,
respectively) with sterile saline. The BALF was centrifuged (12,000
× g at 4°C for 10 min) and ELISA kits were used to detect the
levels of IL-4 (cat. no. 20186; Quanzhou Ruixin Biological
Technology Co., Ltd.), IL-5 (cat. no. ab204523; Abcam), IL-13 (cat.
no. ab219634; Abcam) and IFN-γ (cat. no. ab100689; Abcam),
according to the manufacturer's protocol.
Cell counting in BALF
After centrifugation (5,000 × g at 4°C for 10 min),
part of the BALF sediment was resuspended in 1 ml PBS, BALF
cytospins were stained at room temperature for 3 min using the
Wright-Giemsa method (cat. no. RBR00502; Shanghai Rongbai
Biotechnology Co., Ltd.), according to the manufacturer's protocol,
and cells were counted using a BX53 light microscope
(magnification, ×100; Olympus Corporation) and a hemocytometer. To
avoid errors, each sample was counted by two independent
researchers. The total number of cells and the number of four
different types of inflammatory cells (macrophages, lymphocytes,
eosinophils and neutrophils) were recorded.
Pulmonary histopathological
analysis
The left lungs of the mice were prepared for
histological analysis and fixed in 4% triformol for 48 h at room
temperature. Paraffin-embedded sections were sliced into 5-µm thick
sections for staining with the hematoxylin and eosin kit (Beijing
Solarbio Science & Technology Co., Ltd.) at room temperature
for 1 h to observe inflammatory changes using a BX53 light
microscope (magnification, ×100; Olympus Corporation). Carl Zeiss
Axiovision Viewer Image software (version 4.82.SP2 1CD; Carl Zeiss
AG) were used to assess the mean liner intercept (MLI), matrix
membrane layer and smooth muscle layer of the airways. Periodic
Acid Schiff (PAS) (Beijing Solarbio Science & Technology Co.,
Ltd.) staining was used to assess mucus secretion. PAS+
cells were analyzed using Axiovision Viewer Image software and the
ratio of PAS+ cells to bronchus was calculated.
Western blotting
The right lung was removed, and total protein was
extracted using Total Extraction Sample Kit (Sigma-Aldrich; Merck
KGaA) according to the manufacturer's protocols. Protein
concentrations were determined using the BCA method (cat. no.
23225; Thermo Fisher Scientific, Inc.). Proteins (30 µg) were
resolved by 10% SDS-PAGE, and then transferred to a PVDF membrane.
Following blocking with TBST supplemented with 5% skim milk powder
for 2 h at room temperature, the PVDF membranes were incubated with
antibodies against GATA3 (1:1,000; cat. no. ab106625; Abcam), STAT6
(1:2,000; cat. no. ab32520; Abcam), T-bet (1:500; cat. no. ab91109;
Abcam) or Runx3 (1:1,000; cat. no. ab135248; Abcam) at 4°C
overnight. The membranes were subsequently incubated with
HRP-conjugated goat anti-mouse secondary antibody (1:5,000; cat.
no. ab205719; Abcam) the following day for 1 h at room temperature.
Signals were visualized using ECL Chemiluminescence Detection kit
(cat. no. SW2010; Beijing Solarbio Science & Technology Co.,
Ltd.). Protein expression levels were semi-quantified using
Gel-Pro-Analyzer software (version 4.0; Media Cybernetics, Inc.)
with β-actin as the loading control.
Reverse transcription-quantitative PCR
(RT-qPCR)
Right lung was removed, and total cellular RNA was
extracted using TRIzol® reagent (cat. no. 15596026;
Invitrogen; Thermo Fisher Scientific, Inc.). A One-step RT-PCR kit
(cat. no. 639504; Takara Bio, Inc.) was used according to the
manufacturer's protocols to reverse transcribe RNA into cDNA.
Subsequently, mRNA expression levels of cytokines and transcription
factors were evaluated via RT-qPCR using BlazeTaq™ SYBR®
Green qPCR Mix (GeneCopoeia, Inc.) according to the manufacturer's
protocol. The following thermocycling conditions were used for
qPCR: 95°C for 30 sec; followed by 40 cycles at 95°C for 30 sec and
at 60°C for 30 sec; and then at 72°C for 30 sec. Lightcycler
software (version 480; Roche Molecular Diagnostics) was used to
measure the Cq values and the results were expressed relative to
GAPDH. Primers were selected from PrimerBank (35). The forward and reverse sequences of
the primers used for amplification are presented in Table I. mRNA expression levels were
quantified using the 2−∆∆Cq method (36).
 | Table I.Primer sequences used for reverse
transcription-quantitative PCR. |
Table I.
Primer sequences used for reverse
transcription-quantitative PCR.
| Gene | Primer sequences
(5′→3′) |
|---|
| IL-4 | F:
TGAACGAGGTCACAGGAGAA |
|
| R:
CGAGCTCACTCTCTGTGGTG |
| IL-5 | F:
CTCTGTTGACAAGCAATGAGACG |
|
| R:
TCTTCAGTATGTCTAGCCCCTG |
| IL-13 | F:
TGTGTCTCTCCCTCTGACCC |
|
| R:
CACACTCCATACCATGCTGC |
| IFN-γ | F:
ATGAACGCTACACACTGCATC |
|
| R:
CCATCCTTTTGCCAGTTCCTC |
| GATA3 | F:
TTATCAAGCCCAAGCG |
|
| R:
CCATTAGCGTTCCTCCTC |
| T-bet | F:
GTGACCCAGATGATTGTGCTC |
|
| R:
GTAGGCAGTCACGGCAATG |
| Runx3 | F:
TCCAACAGCATCTTGACTCCTT |
|
| R:
GGTGCTCGGGTCTCGTAT |
| GAPDH | F:
AGGTCGGTGTGAACGGATTTG |
|
| R:
TGTAGACCATGTAGTTGAGGTCA |
Flow cytometry
The spleen was taken to the ultra-clean table,
ground using a cell strainer (mesh size, 100), and the cell
suspension was obtained. After stimulation with phorbol myristate
acetate (25 µg/ml) + lonomycin (20 µg/ml) at 37°C for 4 h (Beijing
Solarbio Science & Technology Co., Ltd.), the cells were fixed
using 4% paraformaldehyde at room temperature for 10 min. Cells
were incubated with PerCP/cyanine5.5-conjugated anti-CD4 (cat. no.
45-0042-80; Thermo Fisher Scientific, Inc.), PE-conjugated
anti-IL-4 (cat. no. 12-7041-41; Thermo Fisher Scientific, Inc.) and
FITC-conjugated anti-IFN-γ (cat. no. 11-7311-41; Thermo Fisher
Scientific, Inc.) antibodies for 20 min at room temperature. Th1
and Th2 cells were sorted using a FACSCanto™ flow cytometer (BD
Biosciences). The corresponding percentage of Th1 cells, Th2 cells
and Th1/Th2 ratio were calculated and analyzed using FlowJo
software (version 10; BD Biosciences).
Statistical analysis
All experiments were performed at least three times.
Data are presented as the mean ± standard deviation, and
differences between four groups were analyzed using one-way ANOVA
followed by Tukey's post hoc test. P<0.05 was considered to
indicate a statistically significant difference.
Results
PM2.5 components
The chemical constituents identified in PM2.5
samples are listed in Table II.
Metals and PAHs accounted for the primary constituents of ambient
PM2.5 samples collected in Yantai, China. The results showed that
sodium, magnesium, aluminum, calcium and zinc were the primary
metals found. Naphthalene, acenaphthene, phenanthrene,
benzofluoranthene and benzopyrene were the primary PAHs in the
ambient PM2.5. The results suggested that the metals and PAHs may
account for the exacerbating effect of PM2.5 on the occurrence and
prevalence of asthma.
| Table II.Metals and PAH composition in
PM2.5. |
Table II.
Metals and PAH composition in
PM2.5.
| A, Metal |
|---|
|
|---|
| Type | PM2.5, µg/mg |
|---|
| Na | 60.51 |
| Mg | 40.93 |
| Al | 37.28 |
| Ca | 32.17 |
| Zn | 18.93 |
| Fe | 11.24 |
| K | 9.17 |
| Mn | 5.40 |
| Ba | 4.92 |
| As | 3.14 |
|
| B, PAHs |
|
| Type | PM2.5,
µg/mg |
|
| Naphthalene | 11.41 |
| Acenaphthene | 11.06 |
| Phenanthrene | 9.17 |
|
Benzofluoranthene | 4.69 |
| Benzo | 1.59 |
| Fluoranthene | 0.97 |
| Pyrene | 0.34 |
| Chrysene | 0.25 |
| Anthracene | 0.13 |
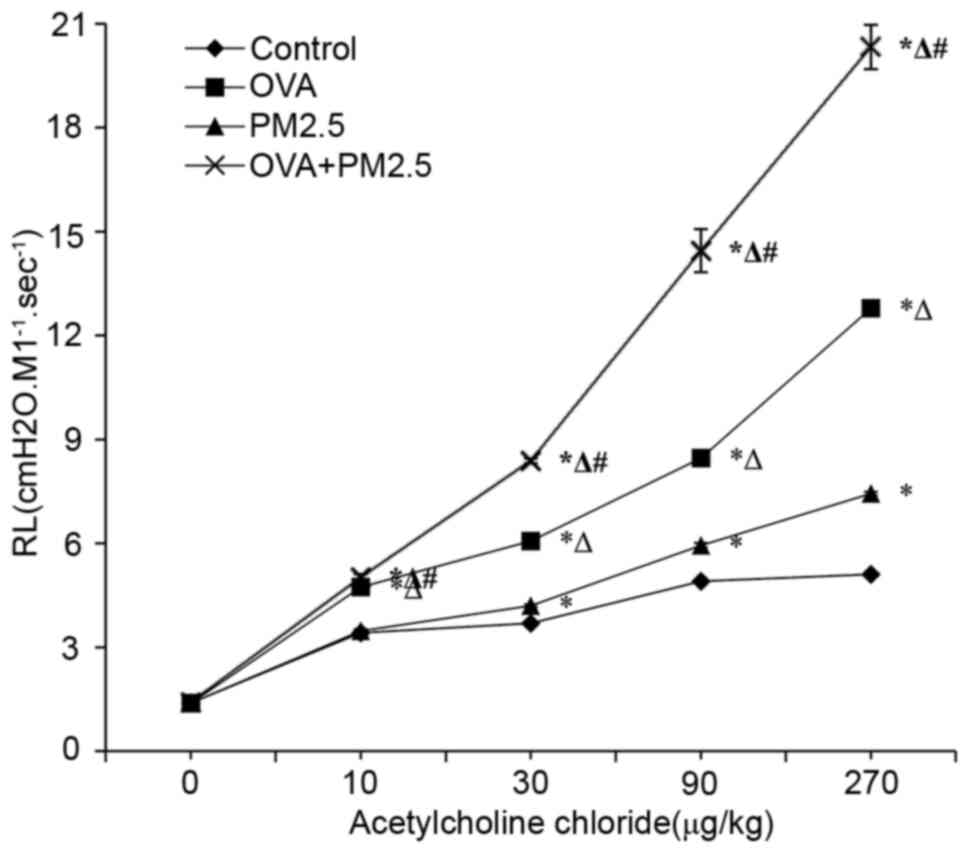
Effects of PM2.5 on AHR
The results of in vivo experiments showed
that increased administration of ACH, OVA and PM2.5 co-exposure
significantly increased the lung resistance compared with that
observed in the groups treated with PM2.5 and OVA alone (Fig. 2). The airway resistance of mice in
the PM2.5 group treated with ACH was significantly higher compared
with the control group, but lower than that in the OVA group.
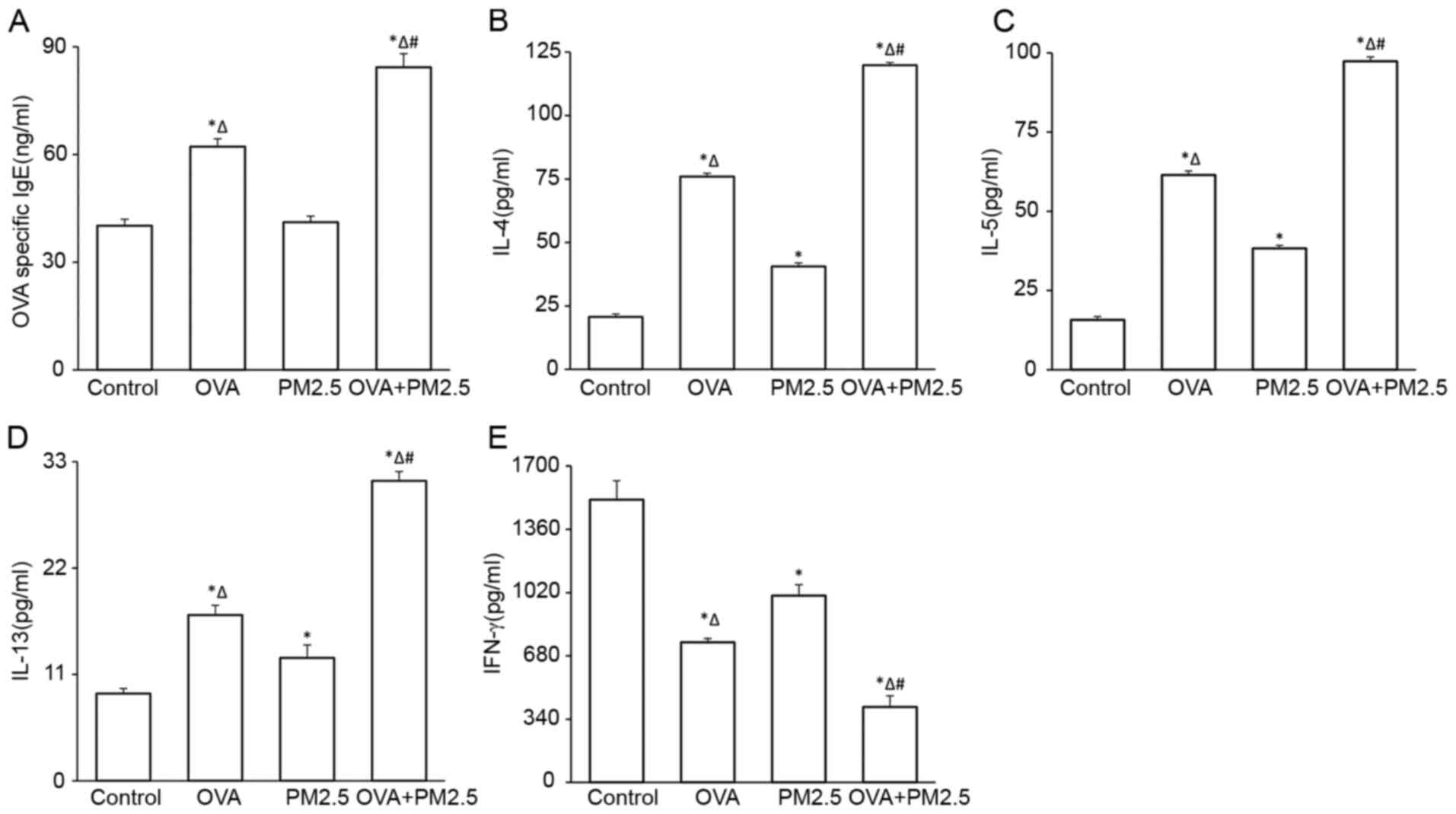
Effects of PM2.5 on the serum levels
of OVA-specific IgE and cytokines in BALF
The levels of serum OVA-specific IgE increased
significantly in the OVA and OVA + PM2.5 groups compared with the
PM2.5 and control groups (Fig. 3A).
Levels of OVA-specific IgE in the OVA + PM2.5 group were increased
notably compared with the OVA group. However, there were no
significant differences between the PM2.5 and control groups. OVA +
PM2.5 significantly increased IL-4, IL-5 and IL-13 levels in BALF
compared with the control, OVA and PM2.5 groups (Fig. 3B-D, respectively); OVA and PM2.5
alone significantly increased the levels of these cytokines in BALF
compared with the control group. PM2.5 alone resulted in less
secretion of these cytokines compared with the OVA mice. The
changes in expression of IFN-γ in these four groups were opposite
to that observed for IgE, IL-4, IL-5 and IL-13 (Fig. 3E).
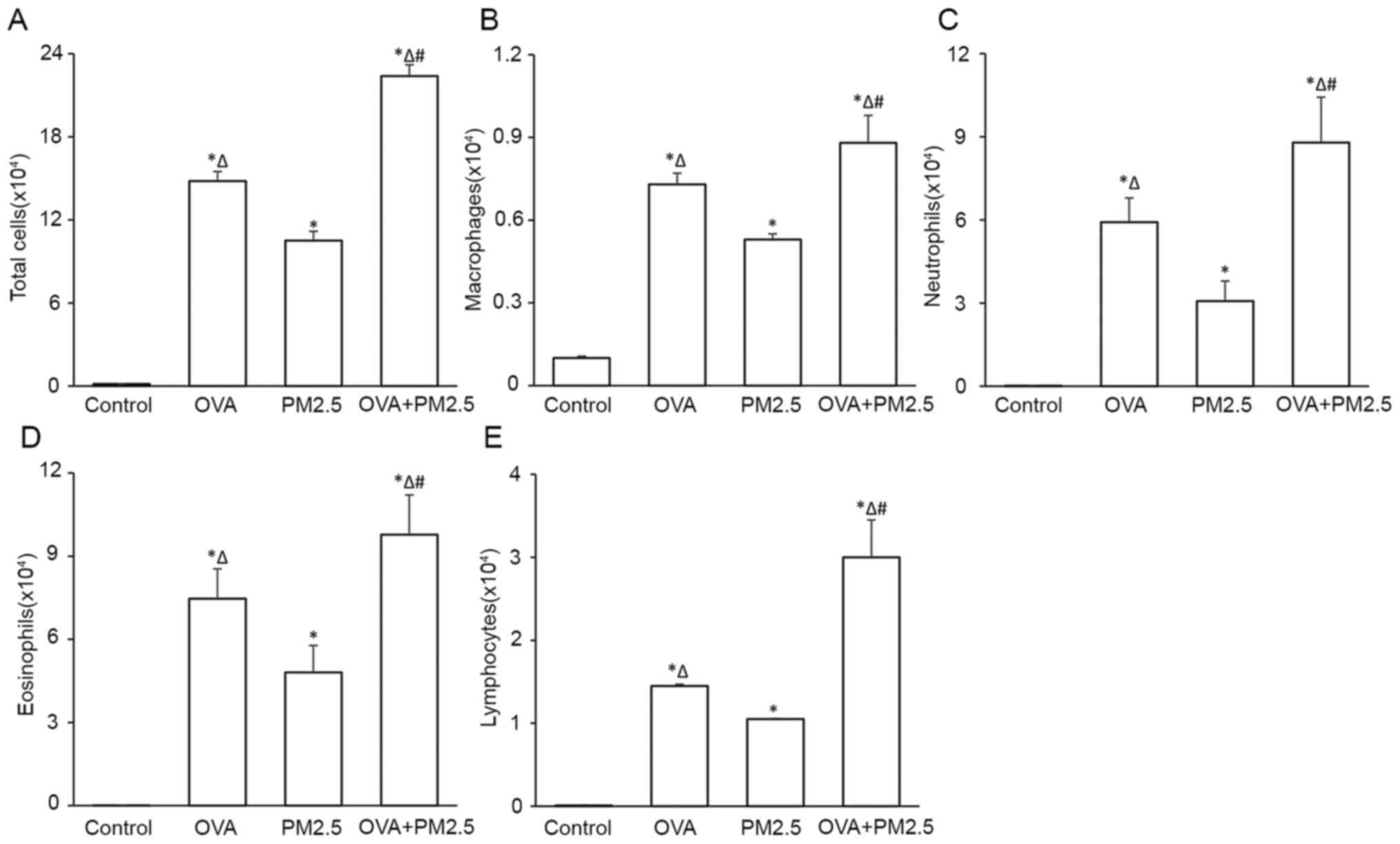
Effects of PM2.5 on inflammatory cell
count in BALF
PM2.5 exposure increased the total inflammatory cell
count and the count of each type of inflammatory cell assessed,
including macrophages, neutrophils, eosinophils and lymphocytes, in
BALF sediments, particularly in the OVA + PM2.5 group (Fig. 4). The increase observed in the OVA
group was significantly higher compared with the PM2.5 alone and
control groups.
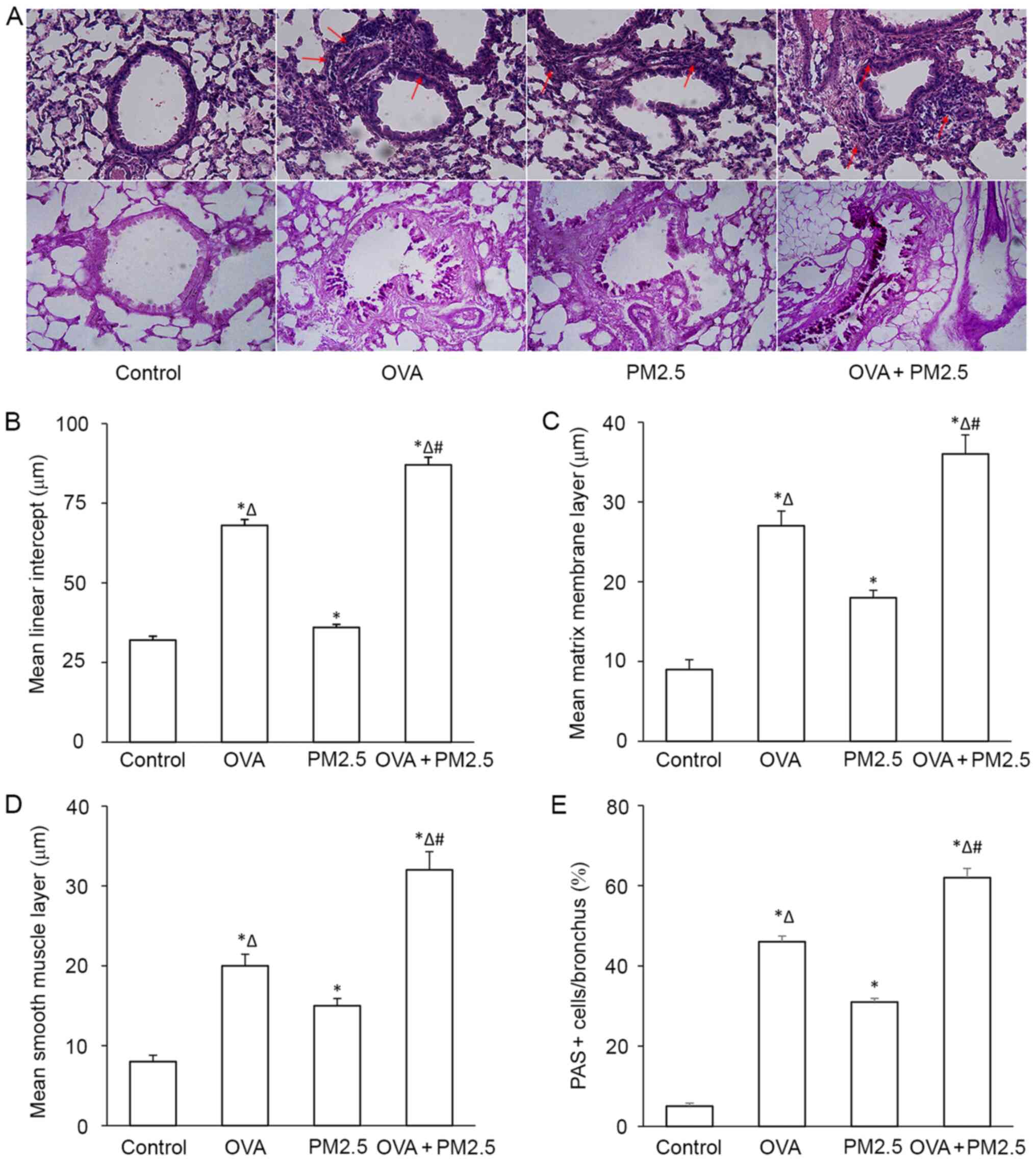
Effects of PM2.5 on pathological
changes in the airway
No pathological alterations were found in the lungs
of the control group. PM2.5 alone caused a small degree of airway
inflammation, whereas OVA exposure exacerbated airway inflammation,
particularly in the OVA + PM2.5 group, OVA + PM2.5 exposure
resulted in focal infiltration of inflammatory cells into the
airway (Fig. 5A). PM2.5 increased
mlI of mice, and a significant difference was observed between the
asthmatic mice compared with the OVA group (Fig. 5B). PM2.5 increased matrix membrane
layer thickness and smooth muscle layer thickness significantly
(Fig. 5C and D, respectively),
particularly in mice treated with both OVA and PM2.5. Furthermore,
mucus secretion was significantly increased in the mice of the OVA
+ PM2.5 group; OVA and PM2.5 alone increased mucus secretion
significantly compared with the control group, and the PM2.5 group
showed fewer PAS+ cells compared with the OVA group
(Fig. 5E).
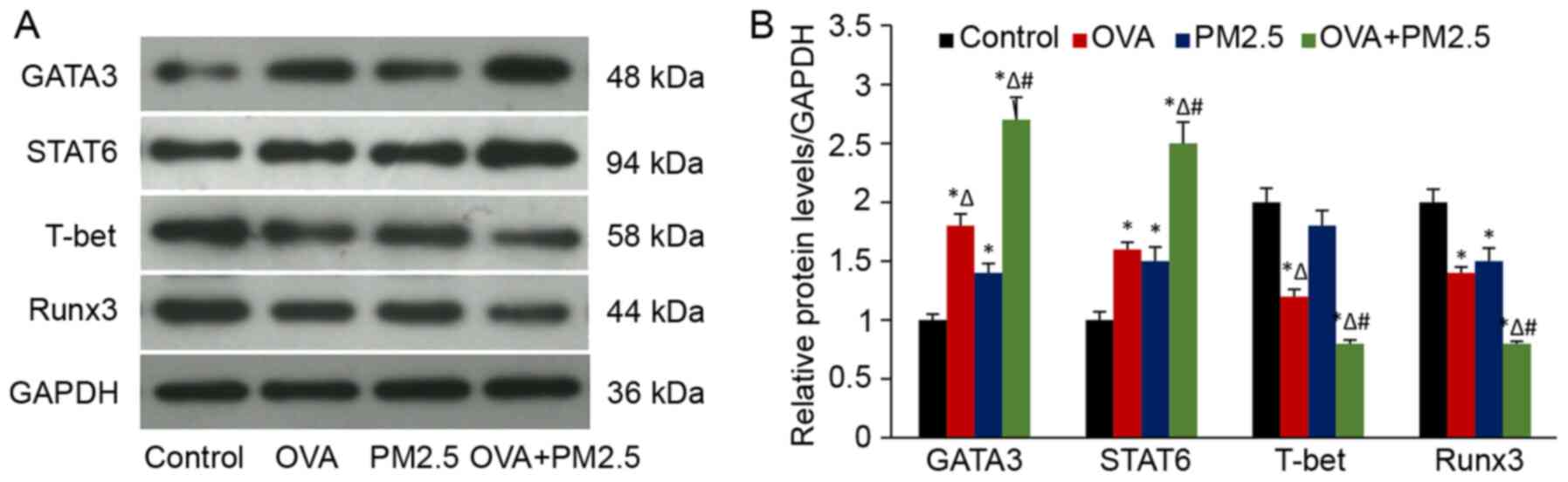
Effects of PM2.5 on the expression of
transcription factors
The results of western blotting showed that a
combination of OVA and PM2.5 significantly enhanced the protein
expression levels of the GATA3 and STAT6 compared with the other
groups (Fig. 6). The expression of
GATA3 and STAT6 in the mice treated with PM2.5 alone was higher
compared with the control group. Expression of the transcription
factors in the OVA group were significantly higher compared with
the PM2.5 group. Conversely, PM2.5 lowered the expression of Runx3,
and the decrease was more pronounced in the OVA + PM2.5 group.
Additionally, expression of Runx3 was significantly lower in the
OVA group compared with the control group. No significant
differences in Runx3 expression were observed between the OVA and
PM2.5 groups. However, expression of T-bet in OVA and OVA + PM2.5
groups were significantly lower compared with all the other groups.
PM2.5 alone decreased the expression of T-bet compared with the
control mice, but no significant differences were observed in the
expression levels of T-bet between the PM2.5 and control groups.
These results suggested that transcription factor Runx3, but not
T-bet served a vital role in PM2.5-induced asthmatic pathological
processes.
Effects of PM2.5 on the mRNA
expression of the cytokines
The mRNA expression levels of inflammatory cytokines
and transcription factors were measured by RT-qPCR. The variation
in trends and mRNA ratios were the same as those observed for the
changes in the protein expression levels (Fig. 7).
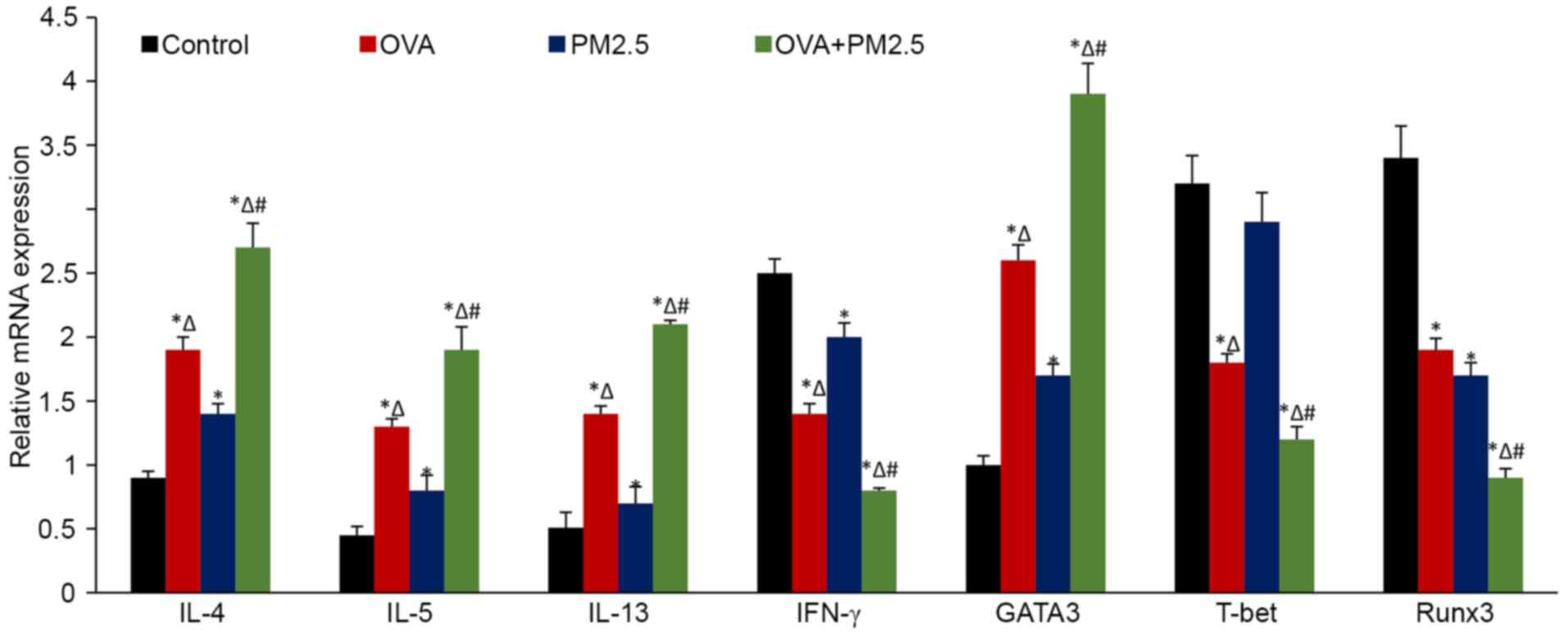 | Figure 7.mRNA expression levels of IL-4, IL-5,
IL-13, IFN-γ, GATA3, T-bet and Runx3 in the lung tissues of mice.
Data are expressed as the mean ± SD; n=6; *P<0.05 vs. Control;
∆P<0.05 vs. PM2.5; #P<0.05 vs. OVA.
OVA, ovalbumin; PM2.5, 2.5 µm particulate matter; GATA3,
trans-acting T cell-specific transcription factor GATA-3;
T-bet, T-box transcription factor TBX21; Runx3, runt-related
transcription factor 3; IFN-γ, interferon; IL-, interleukin. |
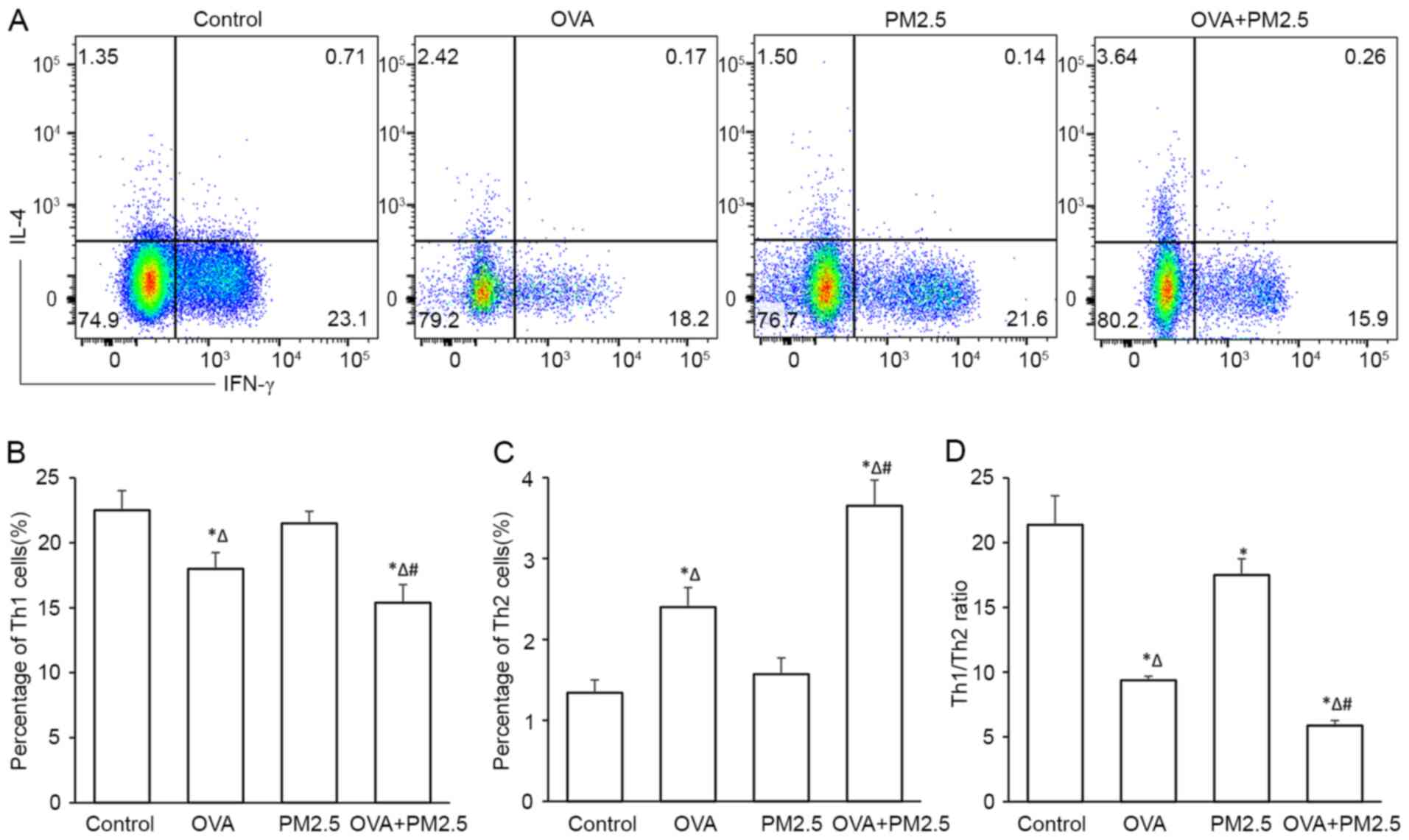
Effects of PM2.5 on the
differentiation of Th1 and Th2
The proportion (%) of Th2 was significantly
increased in the OVA group compared with the control group, as
measured by flow cytometry (Fig.
8). PM2.5 exposure slightly decreased the proportion of Th1
cells and increased the proportion of Th2 cells, but no significant
difference was observed compared with the control group.
Co-exposure of OVA and PM2.5 significantly decreased the population
of Th1 cells and increased the population of Th2 cells. In
addition, PM2.5 significantly disturbed the balance between Th1/Th2
cells compared with the control group, OVA + PM2.5 co-exposure
exhibited an enhanced effect, further disturbing the balance
significantly compared with the OVA group.
Discussion
Although asthma is recognized as a heterogeneous
disease, eosinophilic airway inflammation serves a classical role
in the development of asthma (37),
accompanied by infiltration of mast cells, T lymphocytes and
neutrophils, which is associated with increased release of
Th2-related cytokines (38). Th2
cytokines, mainly result from Th2 cells, serve a crucial role in
allergic inflammation. IL-4 promotes differentiation in Th2 cells,
IL-13 mediates AHR and mucus hyperproduction, whereas IL-5 is
highly specific to activation and recruitment of eosinophils.
IFN-γ, produced by Th1 cells, induces immunity against
intracellular pathogens. The cytokines form a complex inflammatory
network and regulate the immune response. In the present study,
PM2.5 alone significantly increased the levels of Th2 cytokines
IL-4, IL-5 and IL-13, and decreased the levels of IFN-γ compared
with the control mice, consistent with the aggravation of
inflammatory cells, particularly eosinophil infiltration into the
airway, airway mucus hypersecretion and AHR. Although the effect
was lower compared with the OVA group, airway inflammation and AHR
increased significantly in the OVA + PM2.5 group. Together, the
results of the present study showed that ambient PM2.5 collected in
Yantai evoked an allergic inflammatory response in the airway of
mice, and the aggravating effect was notably increased in the mouse
model of asthma. These results share some similarity with previous
studies (39–41). Meanwhile, an imbalance of Th1/Th2
was found in groups exposed to PM2.5, especially in the OVA + PM2.5
group. In the present study, it was found that although PM2.5
decreased the percentage of Th1 cell and increased the percentage
of Th2 cells slightly, PM2.5 did alter the Th1/Th2 cell ratio
significantly. The present study also demonstrated that co-exposure
of PM2.5 and OVA aggravated the imbalance of Th1/Th2.
An imbalance in Th1/Th2 cells was observed in the
groups exposed to PM2.5, particularly in the OVA + PM2.5 group. The
capacity of T cells to produce inflammatory cytokines and the
differentiation of T cells is programmed by transcription factors.
GATA3 is a master transcription factor involved in induction of Th2
differentiation, which is upregulated by IL-4-STAT6 signaling
(42). T-bet controls Th1
differentiation and programme cytokines patterns (43,44).
Additionally, Runx3 is another Th1 transcription factor.
Runt-related transcription factors serve a vital role in T
cell-mediated immunity, which is also responsible for IFN-γ
production. Knockdown of Runx3 results in a notable decrease in
IFN-γ levels in GATA3 and T-bet double-deficient mice (45). Runx3 transgenic mice exhibited a Th1
cell phenotype (45),
Runx3fl/fl mice exhibited spontaneous eosinophilic
airway inflammation and AHR (46).
Zhou et al (47)
demonstrated that mislocalization of Runx3 induces airway
inflammation and AHR in asthmatic mice. Furthermore, Runx3 has been
reported to suppress IL-4, and these inhibitory effects can be
observed without the involvement of T-bet (18,48).
Thus, blocking Runx3 results in diminished IFN-γ production and
Th2-related inflammation. In the present study, it was hypothesized
that PM2.5 evoked airway inflammation by regulating the expression
of transcription factors. It was first found that PM2.5 exposure
significantly increased the protein and mRNA levels of GATA3 and
STAT6, which demonstrated that PM2.5 may alter Th2 differentiation.
The protein and mRNA levels of T-bet in the PM2.5-treated group
(alone) did not differ significantly compared with the control
group, which was different from the OVA group. Whereas expression
of Runx3 decreased significantly compared with the control group.
OVA exposure decreased the expression of both T-bet and Runx3, and
enhanced the expression of both GATA-3 and STAT6. Thus, the results
demonstrated that the transcription factors GATA3, STAT6, T-bet and
Runx3 served a vital role in the pathology of asthma. The present
study also suggested that PM2.5 inhibited the Th1 response
primarily through downregulation of Runx3, but not T-bet. Combined
exposure to PM2.5 and OVA exhibited a enhanced suppressor effect on
Th1 differentiation.
The gene expression level of Runx3 is controlled by
promoters that are rich in CpG islands. Previous studies have
reported that methylation of promoter regions results in silencing
of Runx3 gene expression, which leads to the development of several
diseases (49–51). The present study showed that the
protein and mRNA levels of Runx3 were significantly decreased in
the PM2.5 and OVA exposure groups. However, whether methylation of
Runx3 influences its expression remains undetermined, and will be
assessed in future studies.
The primary ambient components collected in Yantai
were metals and PAHs. An increasing number of studies have
demonstrated that metals in the PM2.5 category contribute to
allergic airway inflammation (52,53).
Additionally, previous studies have reported that PM2.5-associated
PAHs are involved in the development of allergic inflammation
through interactions with the arylhydrocarbon receptor and PAHs
have been suggested to regulate the differentiation of T cells
(54–56). The components of PM2.5 may be
responsible for the initiation and exacerbation of asthma; however,
it remains unclear which specific metals or PAHs are responsible
for the exacerbation observed in the present study.
The present study has some limitations. Numerous
transcription factors are involved in the inflammatory network of
asthma, the master transcription factors GATA3, T-bet and Runx3
have been reported to interact with each other. Previous studies
have demonstrated that GATA3 regulates the expression of T-bet by
repressing the IL-12-STAT4-IFN-γ pathway (57), and T-bet can function with Runx3 to
repress IL-4 and GATA3 expression (19). However, whether these transcription
factors interact with each other in PM2.5-exposed mice remains
unknown, thus future studies will study the effect of gene knockout
mice to assess the roles of these transcription factors.
In conclusion, ambient PM2.5 exposure upregulated
the expression levels of the transcription factors GATA3 and STAT6
and downregulated the expression of Runx3. This resulted in an
imbalance in the ratio of Th1/Th2 cells, and evoked allergic
inflammation and AHR in the airways, and these effects were notably
exacerbated in the mouse model of asthma.
Acknowledgements
Not applicable.
Funding
The present study was supported by a grant from The
Science and Technology Planning Project of Yantai (grant nos.
2020YD004 and 2020YD082).
Availability of data and materials
All data generated or analyzed during the present
study are included in this published article.
Authors' contributions
LP and SZ participated in the design of the study.
YS and YF performed experimental procedures. PY and XL conducted
data analysis. LP, SZ and PY confirmed the authenticity of all the
raw data. All authors read and approved the final manuscript.
Ethics approval and consent to
participate
The experimental animal study was approved by the
Institutional Animal Care and Use Committee of Nanjing Medical
University (Nanjing, China; approval no. 20110217).
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Global Initiative for Asthma (GINA), .
GINA Global strategy for asthma management and prevention. GINA;
Fontana, WI: 2019, www.ginasthma.org
|
|
2
|
GBD 2016 Disease and Injury Incidence and
Prevalence Collaborators, . Global, regional, and national
incidence, prevalence, and years lived with disability for 328
diseases and injuries for 195 countries, 1990–2016: A systematic
analysis for the Global Burden of Disease Study 2016. Lancet.
390:1211–1259. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Holgate ST: Innate and adaptive immune
responses in asthma. Nat Med. 18:673–683. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Hogg JC: The pathology of asthma. APMIS.
105:735–745. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Walsh GM: Anti-IL-4/-13 based therapy in
asthma. Expert Opin Emerg Drug. 20:349–352. 2015. View Article : Google Scholar
|
|
6
|
Arima M and Fukuda T: Prostaglandin
D2 and T(H)2 inflammation in the pathogenesis
of bronchial asthma. Korean J Intern Med. 26:8–18. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Zhu J, Yamane H and Paul WE:
Differentiation of effector CD4 T cell populations. Annu Rev
Immunol. 28:445–489. 2012. View Article : Google Scholar
|
|
8
|
Ho IC, Tai TS and Pai SY: GATA3 and the
T-cell lineage: Essential functions before and after
T-helper-2-cell differentiation. Nat Rev Immunol. 9:125–135. 2010.
View Article : Google Scholar
|
|
9
|
Koh BH, Hwang SS, Kim JY, Lee W, Kang MJ,
Lee CG, Park JW, Flavell RA and Lee GR: Th2 LCR is essential for
regulation of Th2 cytokine genes and for pathogenesis of allergic
asthma. Proc Natl Acad Sci U S A. 23:10614–10619. 2010. View Article : Google Scholar
|
|
10
|
Lighvani AA, Frucht DM, Jankovic D, Yamane
H, Aliberti J, Hissong BD, Nguyen BV, Gadina M, Sher A, Paul WE and
O'Shea JJ: T-bet is rapidly induced by interferon-gamma in lymphoid
and myeloid cells. Proc Natl Acad Sci U S A. 98:15137–15142. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Voon DC, Hor YT and Ito Y: The RUNX
complex: Reaching beyond haematopoiesis into immunity. Immunology.
146:523–536. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Shan Q, Zeng Z, Xing S, Li F, Hartwig SM,
Gullicksrud JA, Kurup SP, Van Braeckel-Budimir N, Su Y, Martin MD,
et al: The transcription factor Runx3 guards cytotoxic
CD8+ effector T cells against deviation towards
follicular helper cell lineage. Nat immunol. 18:931–939. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Cruz-Guilloty F, Pipkin ME, Djuretic IM,
Levanon D, Lotem J, Lichtenheld MG, Groner Y and Rao A: Runx3 and
T-box proteins cooperate to establish the transcriptional program
of effector CTLs. J Exp Med. 206:51–59. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Naoe Y, Setoguchi R, Akiyama K, Muroi S,
Kuroda M, Hatam F, Littman DR and Taniuchi I: Repression of
interleukin-4 in T helper type 1 cells by Runx/Cbf beta binding to
the Il4 silencer. J Exp Med. 204:1749–1755. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Estécio MR, Maddipoti S, Bueso-Ramos C,
DiNardo CD, Yang H, Wei Y, Kondo K, Fang Z, Stevenson W, Chang KS,
et al: RUNX3 promoter hypermethylation is frequent in leukemia cell
lines and associated with acute myeloid leukemia inv(16) subtype.
Br J Haematol. 169:344–351. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Lin Z, Luo M, Chen X, He X, Qian Y, Lai S,
Si J and Chen S: Combined detection of plasma ZIC1, HOXD10 and
RUNX3 methylation is a promising strategy for early detection of
gastric cancer and precancerous lesions. J Cancer. 8:1038–1044.
2017. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Kang KA, Piao MJ, Ryu YS, Maeng YH and
Hyun JW: Cytoplasmic localization of RUNX3 via histone
deacetylase-mediated SRC expression in oxidative-stressed colon
cancer cells. Cell Physiol. 232:1914–1921. 2017. View Article : Google Scholar
|
|
18
|
Men S, Yu Y, Zhang Y, Wang Y, Qian Q, Li W
and Yin C: Methylation landscape of RUNX3 promoter region as a
predictive marker for Th1/Th2 imbalance in bronchiolitis. Med Sci
Monit. 25:7795–7807. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Yagi R, Junttila IS, Wei G, Urban JF Jr,
Zhan K, Paul WE and Zhu J: The transcription factor GATA3 actively
represses RUNX3 protein-regulated production of interferon-gamma.
Immunity. 32:507–517. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Vercelli D: Discovering susceptibility
genes for asthma and allergy. Nat Rev Immunol. 8:169–182. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Lockett GA and Holloway JW: Genome-wide
association studies in asthma; perhaps, the end of the beginning.
Ourr Opin Allergy Clin Immunol. 13:463–469. 2013. View Article : Google Scholar
|
|
22
|
Weiss ST and Silverman EK: Pro: Geome-wide
association studies (GWAS) in asthma. Am J Respir Crit Care Med.
184:631–633. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Orellano P, Quaranta N, Reynoso J, Balbi B
and Vasquez J: Effect of outdoor air pollution on asthma
exacerbations in children and adults: Systematic review and
multilevel meta-analysis. PLoS One. 12:e01740502017. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Vempilly J, Abejie B, Diep V, Gushiken M,
Rawat M and Tyner TR: The synergetic effect of ambient PM2.5
exposure and rhinovirus infection in airway dysfunction in asthma:
A pilot observational study from the central valley of California.
Exp Lung Res. 39:434–440. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Liu H, Fan X, Wang N, Zhang Y and Yu J:
Exacerbating effects of PM2.5 in OVA-sensitized and channenged mice
and the expression of TRPA1 and TRPV1 proteins in lungs. J Asthma.
54:807–817. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Wang L, Xu J, Liu H, Li J and Hao H: PM2.5
inhibits SOD1 expression by up-regulating microRNA-206 and promotes
ROS accumulation and disease progression in asthmatic mice. Int
Immunopharmacology. 76:1058712019. View Article : Google Scholar
|
|
27
|
He M, Ichinose T, Yoshida Y, Arashidani K,
Yoshida S, Takano H, Sun G and Shibamoto T: Urban PM2.5 exacerbates
allergic inflammation in the murine lung via a
TLR2/TLR4/MyD88-signaling pathway. Sci Rep. 7:110272017. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Pang L, Zou S, Shi Y, Mao Q and Chen Y:
Apigenin attenuates PM2.5-induced airway hyperresponsiveness and
inflammation by down-regulating NF-κB in murine model of asthma.
Int J Exp Pathol. 12:3700–3709. 2019.
|
|
29
|
Zhu XM, Wang Q, Xing WW, Long MH, Fu WL,
Xia W, Jin C, Guo N, Xu DQ and Xu DG: PM2.5 induces
autophagy-mediated cell death via NOS2 signaling in human bronchial
epithelium cells. Int J Biol Sci. 14:557–564. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Lu X, Fu H, Han F, Fang Y, Xu J, Zhang L
and Du Q: Lipoxin A4 regulates PM2.5-induced severe allergic asthma
in mice via the Th1/Th2 balance of group 2 innate lymphoid cells. J
Thorac Dis. 10:1449–1459. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Sun L, Fu J, Lin SH, Sun JL, Xia L, Lin
CH, Liu L, Zhang C, Yang L, Xue P, et al: Particulate matter of
2.5µm or less in diameter disturbs the balance of TH17/regulatory T
cells by targeting glutamate oxaloacetate transaminase 1 and
hypoxia-inducible factor 1α in an asthma model. J Allergy Clin
Immunol. 145:402–414. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Yu PF, Pang LL, Mao QS, Zou SC, Shi Y and
Lin DJ: Dose dependency PM2.5 aggravated airway inflammation in
asthmatic mice via down-regulating expression of ITGB4. Eur Rev Med
Pharmacol Sci. 23:1688–1697. 2019.PubMed/NCBI
|
|
33
|
Li RR, Pang LL, Du Q, Shi Y, Dai WJ and
Yin KS: Apigenin inhibits allergen-induced airway inflammation and
switches immune response in a murine model of asthma.
Immunopharmacol Immunotoxicol. 32:364–370. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Shi Y, Tan Y, Mao S and Gu W: Naringenin
inhibits allergen induced airway remodeling in a murine model of
asthma. Mol Med Rep. 9:1204–1208. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Wang X, Spandidos A, Wang H and Seed B:
Primerbank: A PCR primer database for quantiative gene expression
analysis, 2012 update. Nucleic Acids Res. 40:D1144–D1149. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Nakagome K, Matsushita S and Nagata M:
Neutrophilic inflammation in severe asthma. Int Arch Allergy
Immunol. 158 (Suppl 1):96–102. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Holgate ST: The epidemic of allergy and
asthma. J R Soc Med. 97:103–110. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Ogino K, Nagaoka K, Okuda T, Oka A, Kubo
M, Eguchi E and Fujikura Y: PM2.5-induced airway inflammation and
hyperresponsiveness in NC/Nga mice. Environ Toxicol. 32:1047–1054.
2017. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Ogino K, Nagaoka K, Ito T, Takemoto K,
Okuda T, Nakayama SF, Ogino N, Seki Y, Hanada H, Takashiba S and
Fujikura Y: Involvement of PM2.5-bound protein and metals in
PM2.5-induced allergic inflammation in mice. Inhal Toxicol.
30:498–508. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Shen Y, Zhang ZH, Hu D, Ke X, Gu Z, Zou
QY, Hu GH, Song SH, Kang HY and Hong SL: The airway inflammation
induced by nasal inoculation of PM2.5 and the treatment of
bacterial lysates in rats. Sci Rep. 8:98162018. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Ouyang W, Ranganath SH, Weindel K,
Bhattacharya D, Murphy TL, Sha WC and Murphy KM: Inhibition of Th1
development mediated by GATA-3 through an IL-4-independent
mechanism. Immunity. 9:745–755. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Robinson DS and Lloyd CM: Asthma: T-bet-a
master controller? Curr Biol. 12:R322–R324. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Robinson DS: The role of the T cell in
asthma. J Allergy Clin Immmunol. 126:1081–1091. 2010. View Article : Google Scholar
|
|
45
|
Kohu K, Ohmori H, Wong WF, Onda D, Wakoh
T, Kon S, Yamashita M, Nakayama T, Kubo M and Satake M: The Runx3
transcription factor augments Th1 and down-modulates Th2 phenotypes
by interacting with and attenuating GATA3. J Immunol.
183:7817–7824. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Fainaru O, Woolf E, Lotem J, Yarmus M,
Brenner O, Goldenberg D, Negreanu V, Bernstein Y, Levanon D, Jung S
and Groner Y: Runx3 regulates mouse TGF-beta-mediated dendritic
cell function and its absence results in airway in inflammation.
EMBO J. 23:969–979. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Zhou X, Zhu J, Bian T, Wang R and Gao F:
Mislocalization of Runt-related transcription factor 3 results in
airway inflammation and airway hyper-responsiveness in a murine
asthma model. Exp Ther Med. 14:2695–2701. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Djuretic IM, Levanon D, Negreanu V, Groner
Y, Rao A and Ansel KM: Transcription factors T-bet and Runx3
cooperate to activate Ifng and silence Il4 in T helper type 1
cells. Nat Immunol. 8:145–153. 2007. View
Article : Google Scholar : PubMed/NCBI
|
|
49
|
Saikia S, Rehman AU, Barooah P, Sarmah P,
Bhattacharyya M, Deka M, Deka M, Goswemi B, Husain SA and Medhi S:
Alteration in the expression of MGMT and RUNX3 due to non-CpG
promoter methylation and their correlation with different risk
factors in esophageal cancer patients. Tumour Biol.
39:10104283177016302017. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Wang S, He Z, Li D, Zhang B, Li M, Li W,
Zhu W, Xing X, Zeng X, Wang Q, et al: Aberrant methylation of RUNX3
is present in aflatoxin B1-induced transformation of the L02R cell
line. Toxocology. 385:1–9. 2017. View Article : Google Scholar
|
|
51
|
Liu ZH, Liu JJ, Li SS and Yang XM:
Association of RUNX3 methylation with clinical outcome and cell
migration/invasion in laryngeal squamous cell carcinoma. Cancer
Invest. 34:105–113. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Ogino K, Zhang R, Takahashi H, Takemoto K,
Kubo M, Murakami I, Wang DH and Fujikura Y: Allergic airway
inflammation by nasal inoculation of particulate matter(PM2.5) in
NC/Nga mice. PLoS One. 26:e927102014. View Article : Google Scholar
|
|
53
|
Ng CFS, Hashizume M, Obase Y, Doi M,
Tamura K, Tomari S, Kawano T, Fukushima C, Matsuse H, Chung Y, et
al: Associations of chemical composition and sources of PM with
lung function of severe asthmatic adults in a low air pollution
environment of urban Nagasaki, Japan. Environ Pollut. 252:599–606.
2019. View Article : Google Scholar : PubMed/NCBI
|
|
54
|
Quintana FJ, Basso AS, Iglesias AH, Korn
T, Farez FM, Bettelli E, Caccamo M, Oukka M and Weiner HL: Control
of T(reg) and T(H)17 cell differentiation by the ary1 hydrocarbon
receptor. Nature. 453:65–71. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
55
|
Xia M, Harb H, Saffari A, Sioutas C and
Chatila TA: A jagged 1-Notch 4 molecular switch mediates airway
inflammation induced by ultrafine particles. J Allergy Clin
Immunol. 142:1243–1256.e17. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
56
|
Kawasaki H, Chang HW, Tseng HC, Hsu SC,
Yang SJ, Hung CH, Zhou Y and Huang SK: A tryptophan metabolite,
kynurenine, promotes mast cell activation through aryl hydrocarbon
receptor. Allergy. 69:445–452. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
57
|
Usui T, Nishikomori R, Kitani A and
Strober W: GATA-3 suppresses Th1 development by downregulation of
Stat4 and not through effects on IL-12Rbeta2 chain or T-bet.
Immunity. 18:415–428. 2003. View Article : Google Scholar : PubMed/NCBI
|