Introduction
Age-related cataract (ARC) is the leading cause of
vision impairment and blindness worldwide (1). Surgery is the most effective treatment
strategy for cataracts (2,3). However, some postoperative
complications may occur, such as posterior capsule opacification
(3,4). Previous studies have demonstrated that
lens epithelial cell apoptosis is an early event in cataract
development (5,6). Thus, determining the underlying
molecular mechanism of lens epithelial cells in cataract remains
essential.
MicroRNAs (miRNAs/miRs) are a class of small
non-coding RNAs (~22 nt) that can modulate the transcription of
genes (7,8). Previous studies have reported that
abnormal expression of miRNAs is involved in cataracts development
(9,10). For example, miR-34 facilitates
apoptosis of lens epithelial cells via transforming growth factor
(TGF)-β/Smads signaling (11).
Furthermore, miR-378a modulates reactive oxygen species and the
PI3K/AKT pathway in cataract (12),
while miR-23b-3p regulates lens epithelial cell apoptosis and
autophagy by repressing sirtuin-1 in cataract (13). Recently, miR-124 was reported to
repress the viability and invasion of retinoblastoma cells by
regulating signal transducer and activator of transcription 3
(STAT3) (14). However, the
function of miR-124 in ARC progression remains unclear.
Several studies have suggested that dysregulation of
protein sprouty homolog 2 (SPRY2) participates in the progression
of different ocular diseases (15,16).
For example, Shin et al (17) reported that SPRY2 overexpression in
lens cells suppresses TGF-β-induced epithelial-to-mesenchymal
transition (EMT) transition and cataract formation. In addition,
Tan et al (18) demonstrated
that SPRY2 inhibits the development of lens epithelial cells in
anterior subcapsular cataract via the TGF-β signaling pathway.
Matrix metalloproteinase-2 (MMP-2) is crucial for
retinoblastoma cellular migration and angiogenesis (19). Awasthi et al (20) demonstrated that downregulation of
MMP-2 by proteasome suppression decreases the migration of lens
epithelial cells and prevents posterior capsular opacification.
However, the biological roles of SPRY2 and MMP-2 in ARC are yet to
be investigated.
Thus, the present study aimed to investigate miR-124
expression in ARC tissues and determine whether miR-124 modulates
lens epithelial cell apoptosis by regulating SPRY2 and MMP-2.
Materials and methods
Clinical samples
A total of 28 ARC anterior capsular tissues
(cataracts) and paired normal anterior capsule tissues (without
cataracts) were collected from the First People's Hospital of
Changzhou (Changzhou, China) between September 2016 and February
2018. The patients included 16 women and 12 men, with a mean age of
63 years (age range, 58–72 years). Based on Lens Opacities
Classification System III (21), 28
patients whose lenses with a score of C1-C3, N1-N3, or P1-P3 were
enrolled as ARC group, and 28 age-matched individuals who underwent
vitrectomy operation of epiretinal membranes were taken as the
control group. Patients with complex cataracts with high myopia,
ocular trauma, diabetes, and ocular inflammation were excluded from
the study. The tissue samples were rapidly frozen in liquid
nitrogen and then stored at −80°C prior to subsequent
experimentation. The present study was approved by the Ethics
Committee of the First People's Hospital of Changzhou and all
patients provided written informed consent prior to the study.
Cell culture
The human lens epithelial cell line, SRA01/04, was
purchased from the American Type Culture Collection and maintained
in Dulbecco's modified Eagle's medium (Gibco; Thermo Fisher
Scientific, Inc.) supplemented with 10% fetal bovine serum (Gibco;
Thermo Fisher Scientific, Inc.), 100 U/ml penicillin (Gibco; Thermo
Fisher Scientific, Inc.), and 100 U/ml streptomycin (Gibco; Thermo
Fisher Scientific, Inc.) at 37°C in 5% CO2.
Cell transfection
Short hairpin RNA (shRNA) targeting SPRY2 (shSPRY2;
10 nM; 5′-CCUUACCAUUCCUCCACUUTT-3′), shRNA targeting MMP-2
(shMMP-2; 10 nM; 5′-GCUGACCUGGAAGAGAACATT-3′) and their negative
control (shNC; 10 nM; 5′-UUCUCCGAACGUGUC-3′), miR-124 mimics (10
nM; 5′-AACAUUCAACGCUGUCGGUGAGU-3′), NC mimics (10 nM;
5′-UUCUCCGAACGUGUCACGUTT-3′), miR-124 inhibitor (10 nM;
5′-ACUCACCGACAGCGUUGAAUGUU-3′) and NC inhibitor (10 nM;
5′-CAGUACUUUUGUGUAGUACAA-3′) were purchased from Shanghai
GenePharma Co., Ltd. Cell transfection was performed in SRA01/04
(1×105 cells/well) using Lipofectamine® 2000
reagent (Invitrogen; Thermo Fisher Scientific, Inc.) for 48 h at
37°C according to the manufacturer's instructions. The full-length
of SPRY2 and MMP-2 were subcloned into pcDNA3.1 (10 nM; Shanghai
GenePharma Co., Ltd.) to overexpress SPRY2 and MMP-2 levels with
empty pcDNA3.1 (10 nM; Shanghai GenePharma Co., Ltd.) serving as
the control. The efficiency of transfection was determined in each
experiment using reverse transcription-quantitative (RT-q)PCR 48 h
post-transfection. Subsequent experiments were performed at 48 h
post-transfection.
RT-qPCR
Total RNA was extracted from ARC tissues and
SRA01/04 cells using TRIzol® reagent (Invitrogen; Thermo
Fisher Scientific, Inc.), according to the manufacturer's
instructions. Total RNA was reverse transcribed into cDNA using the
PrimeScript™ RT Reagent Kit (cat. no. RR047A; Takara Biotechnology
Co., Ltd.) according to the manufacturer's instructions. miR-124
expression was detected using the TaqMan™ MicroRNA kit (cat. no.
4440885; Applied Biosystems; Thermo Fisher Scientific, Inc.). qPCR
was subsequently performed using the TB Green® Premix Ex
Taq™ II detection kit (cat. no. RR820A; Takara Bio, Inc.) and
LightCycler® 480 Real-Time PCR System (Roche
Diagnostics). The following thermocycling conditions were used for
qPCR: Initial denaturation at 95°C for 3 min; 40 cycles of 95°C for
5 sec and 60°C for 30 sec. Relative expression levels were
calculated using the 2−∆∆Cq method (22) and normalized to the internal
reference genes GAPDH or U6. The sequences of the primers were as
follows: miR-124 forward, 5′-GTTGGTGGGAGCTACATTGTCTGC-3′ and
reverse, 5′- GTGTCGTGGACTCGGCAATTC-3′; SPRY2 forward,
5′-TCGGCAGGTCCCTTTGTCATCC-3′ and reverse,
5′-TGCAGGTCAACTGGTGTCGT-3′; MMP-2 forward,
5′-CACTTCCACCACCTCCTGTT-3′ and reverse, 5′-TGACCGCTTTCTCCTAGCTC-3′;
GAPDH forward, 5′-TGAGCGCGGCTACAGCTT-3′ and reverse,
5′-TCCTTAATGTCACGCACGATTT-3′; U6 forward,
5′-ATTGGAACGATACAGAGAAGATT-3′ and reverse,
5′-GGAACGCTTCACGAATTTG-3′.
Cell proliferation
The Cell Counting Kit-8 (CCK-8; Dojindo Molecular
Technologies, Inc.) assay was performed to assess cell viability.
Briefly, SRA01/04 cells were seeded into 96-well plates at
1×104 cells per well. Following incubation for 48 h at
37°C, 10 µl CCK-8 reagent was added to each well and cell viability
was subsequently analyzed at a wavelength of 450 nm, using a
microplate reader (Thermo Fisher Scientific, Inc.).
Cell apoptosis
The TUNEL assay was performed using the One-Step
TUNEL Apoptosis Assay kit (cat. no. C1089; Beyotime Institute of
Biotechnology) to assess cell apoptosis according to the
manufacturer's protocol. Briefly, SRA01/04 cells (1×104)
were seeded onto coverslips and fixed with 4% paraformaldehyde at
4°C for 25 min. Cells were subsequently treated with DAPI solution
(300 nM) at room temperature for 1 min and observed in five
randomly selected fields of view under a fluorescence microscope
(Olympus Corporation; magnification, ×200).
Dual-luciferase reporter assay
starbase (starbase.sysu.edu.cn/) was used to predict the
potential target sequences between miR-124 and SPRY2 and between
miR-124 and MMP-2. The mutant sequences of SPRY2 and MMP-2 were
generated using a Site-Directed Mutagenesis Kit (cat. no. E0554S;
New England Biolabs, Inc.). The wild-type (wt) or mutant (mut)
3′-untranlasted region of SPRY2 or MMP-2 was cloned into a pmirGLO
reporter vector (Promega Corporation). The pmirGLO-SPRY2-wt,
SPRY2-mut, pmirGLO-MMP-2-wt or MMP-2-mut reporter vector was
co-transfected with miR-124 mimics, NC mimics, miR-124 inhibitor or
NC inhibitor into SRA01/04 cells (1×105 cells/well)
using Lipofectamine 2000 reagent (Invitrogen; Thermo Fisher
Scientific, Inc.). Following incubation for 48 h at 37°C, the
luciferase activities were detected using a Dual-Luciferase
Reporter Assay System (Promega Corporation). Renilla
luciferase activity was used as the normalization.
Western blotting
Total protein was extracted from SRA01/04 cells
using RIPA buffer. (Thermo Fisher Scientific, Inc.). Total protein
was quantified using an Enhanced BCA Protein Assay kit (Beyotime
Institute of Biotechnology), following which, 50 µg protein/lane
was separated via SDS-PAGE on 10% gel (Sigma-Aldrich; Merck KGaA).
The separated proteins were transferred onto PVDF membranes (EMD
Millipore) and subsequently blocked with 5% skimmed milk for 1 h at
room temperature. The membranes were then incubated for 12 h at 4°C
with the primary antibodies (all Abcam): Anti-SPRY2 (1:1,000; cat.
no. ab180527), anti-MMP-2 (1:1,000; cat. no. ab92536) and
anti-GAPDH (1:1,000, cat. no. ab9485). Following the primary
incubation, membranes were incubated with HRP-conjugated secondary
antibodies goat anti-rabbit immunoglobin G (IgG; 1:20,000; cat. no.
ab205718; Abcam) for 2 h at 37°C. Protein bands were visualized
using an ECL detection system (EMD Millipore).
RNA immunoprecipitation (RIP)
Argonaute 2 (Ago2) RIP was used to determine the
interaction between miR-124 and SPRY2 or MMP-2, using the EZ-Magna
RIP™ RNA-Binding Protein Immunoprecipitation kit (cat. no. 17-701;
EMD Millipore). Briefly, SRA01/04 cells (1×105
cells/well) were transfected with miR-124 mimics or NC mimics for
48 h at 37°C. Cells were subsequently lysed using RIP Lysis Buffer
(EMD Millipore) for 5 min at 4°C. After centrifugation 10,000 × g
at 4°C for 5 min, cell lysates were conjugated to magnetic beads (2
µg; Thermo Fisher Scientific, Inc.) via rotation, using Ago2 and
control IgG (cat. no. ab109761; 1:50; Abcam). The magnetic beads
were treated with 0.5 mg/ml Proteinase K (EMD Millipore) to digest
the protein and then immunoprecipitated RNA was isolated. The
immunoprecipitated RNA was analyzed via RT-qPCR analysis.
Statistical analysis
Statistical analysis was performed using SPSS 22.0
(IBM Corp.). All experiments were performed in triplicate and data
are presented as the mean ± standard deviation. Both paired and
unpaired Student's t-test were used to compare differences between
two groups, while one-way ANOVA, followed by Tukey's post hoc test
was used to compare differences between multiple groups. P<0.05
was considered to indicate a statistically significant
difference.
Results
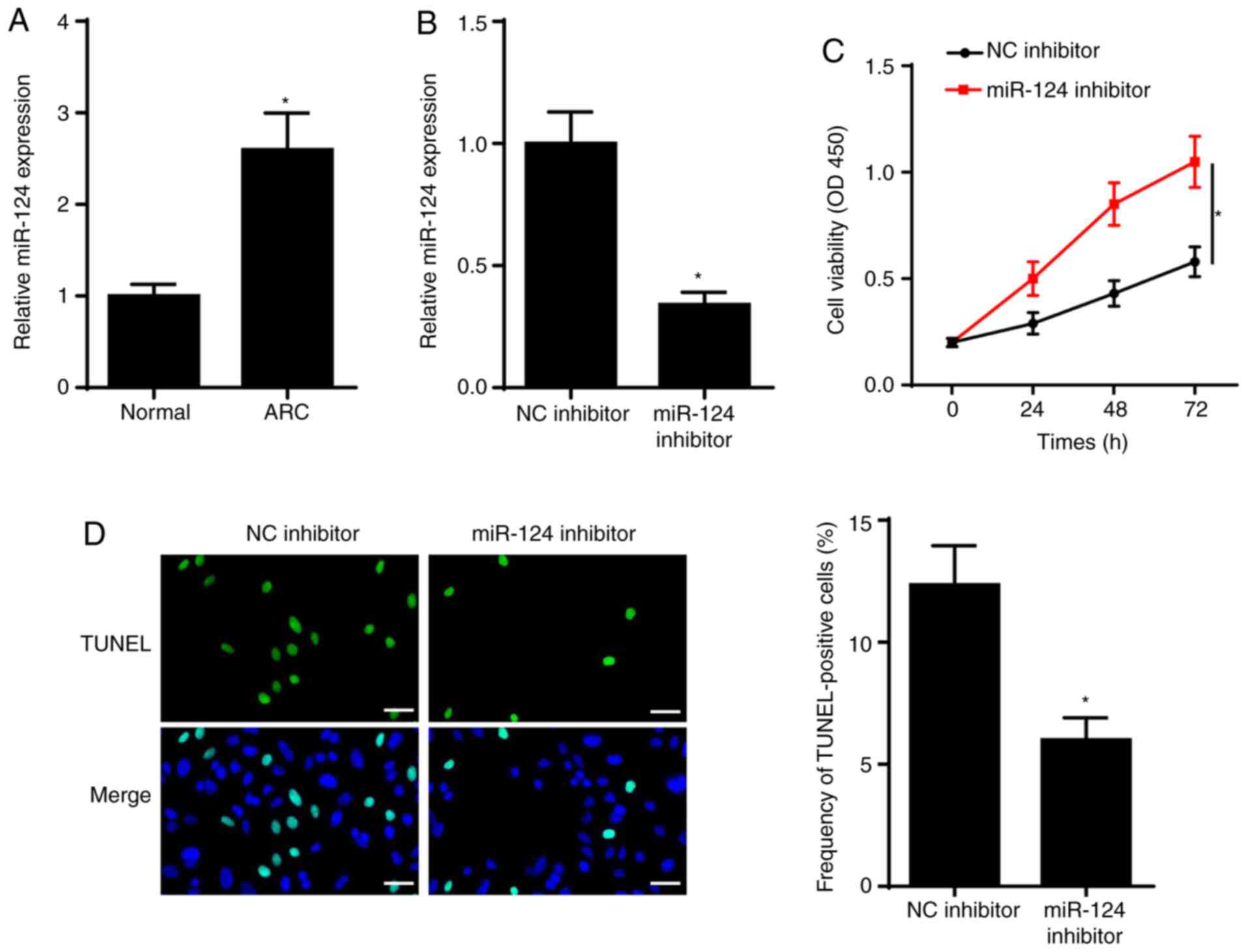
miR-124 expression is upregulated in
ARC and miR-124 inhibitor inhibits SRA01/04 cell apoptosis
To investigate the role of miR-124 in ARC, RT-qPCR
analysis was performed to detect miR-124 expression in ARC tissues.
The results demonstrated that miR-124 expression was significantly
increased in the anterior lens capsules of patients with ARC
(Fig. 1A). To determine the effect
of miR-124 on ARC, SRA01/04 cells were transfected with NC
inhibitor and miR-124 inhibitor. As presented in Fig. 1B, miR-124 expression significantly
decreased in SRA01/04 cells transfected with miR-124 inhibitor. The
results of the CCK-8 assay demonstrated that miR-124 knockdown
increased SRA01/04 cell viability (Fig.
1C). The results of the TUNEL assay indicated that SRA01/04
cell apoptosis significantly decreased following transfection with
miR-124 inhibitor (Fig. 1D). Taken
together, these results suggested that miR-124 promoted cell
viability and repressed apoptosis of SRA01/04 cells.
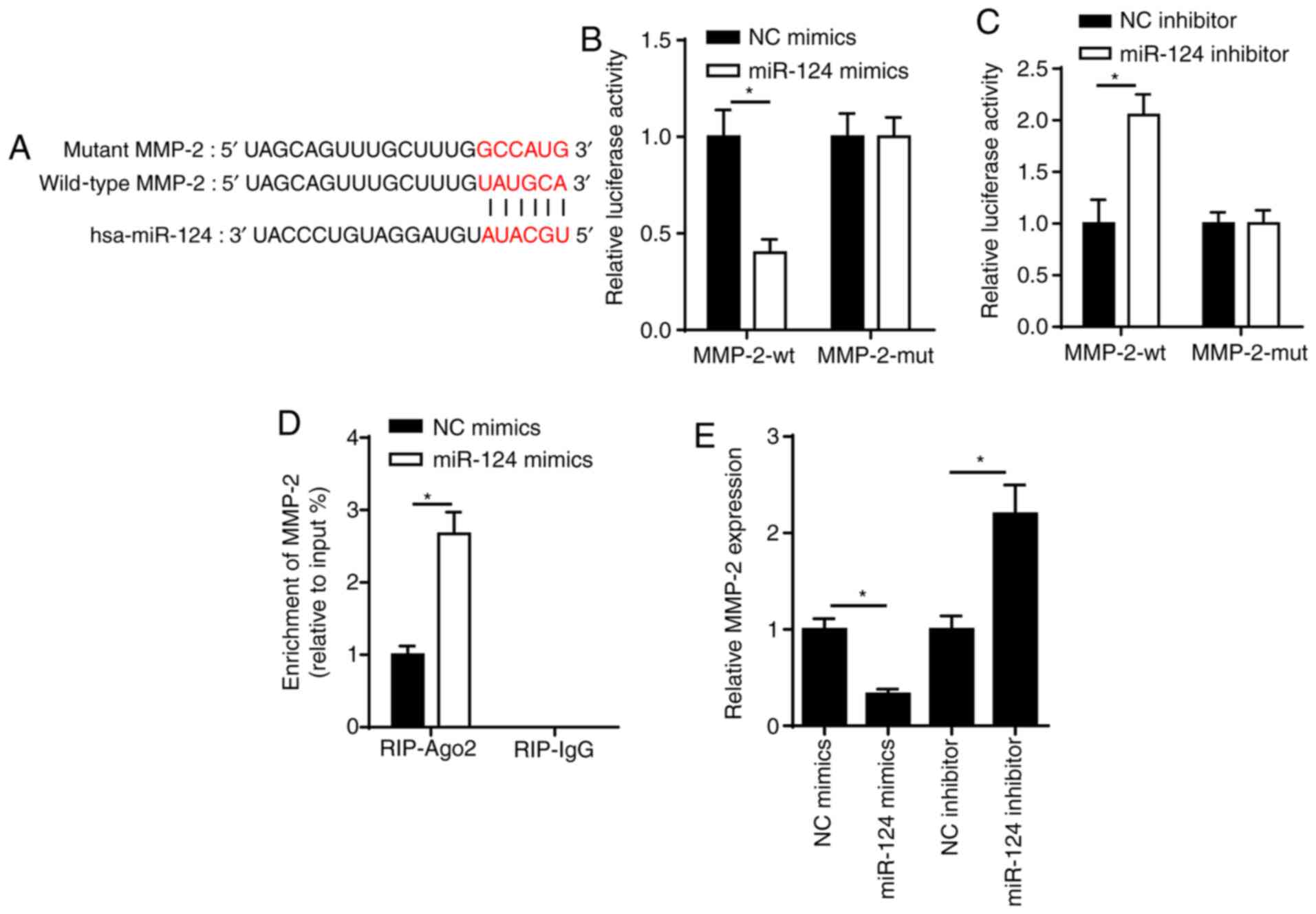
SPRY2 is a target of miR-124
To verify the potential molecular mechanism of
miR-124 in ARC progression, StarBase was used to predict the
putative binding sequences of miR-124 and SPRY2 (Fig. 2A). This interaction was further
validated via the dual-luciferase reporter and RIP assays. The
dual-luciferase reporter assay demonstrated that overexpression of
miR-124 significantly decreased the relative luciferase activity of
wt-SPRY2; however, no significant changes were observed in the
relative luciferase activity of mut-SPRY2 (Fig. 2B). In addition, miR-124 knockdown
increased the luciferase activity of wt-SPRY2; however, no
significant changes were observed in the luciferase activity of
mut-SPRY2 (Fig. 2C). The results of
the RIP assay demonstrated that SRA01/04 cells transfected with
miR-124 mimics enhanced the enrichment of SPRY2 in the Ago2 group
(Fig. 2D). Notably, the addition of
miR-124 elevated miR-124 expression in SRA01/04 cells (Fig. 2E). SPRY2 expression was decreased in
SRA01/04 cells transfected with miR-124 mimics; however, SPRY2
expression increased in SRA01/04 cells transfected with miR-124
inhibitor (Fig. 2F). Collectively,
these results confirmed that SPRY2 was a target of miR-124, and
SPRY2 expression was negatively regulated by miR-124.
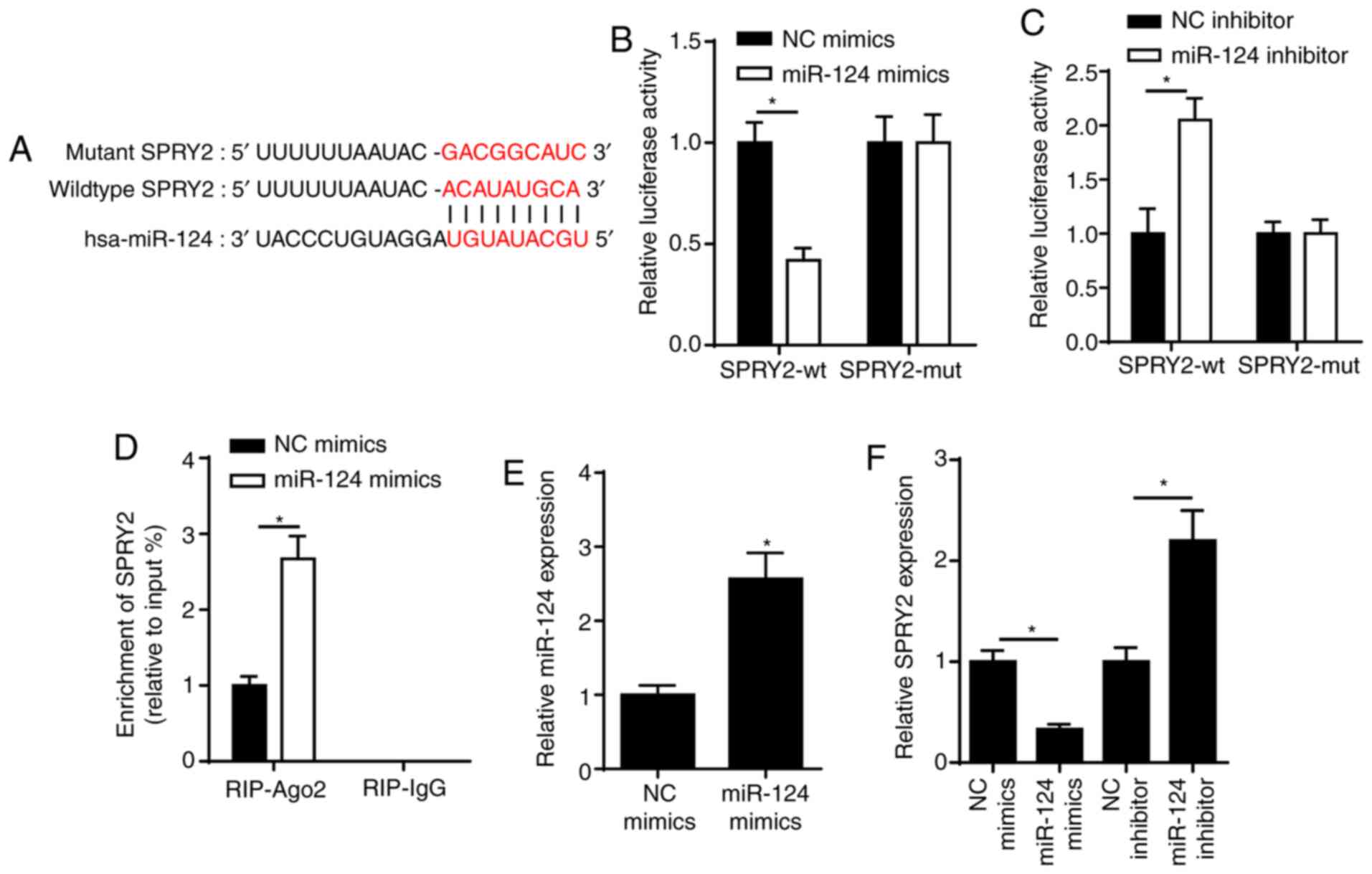 | Figure 2.SPRY2 is a target of miR-124. (A) The
binding sequence between miR-124 and SPRY2 was predicted using the
StarBase website. (B) Dual-luciferase reporter assay showed the
luciferase activity of SPRY2-wt or SPRY2-mut in SRA01/04 cells
transfected with NC mimics or miR-124 mimics. (C) Dual-luciferase
reporter assay showed the luciferase activity of SPRY2-wt or
SPRY2-mut in SRA01/04 cells transfected with NC inhibitor or
miR-124 inhibitor. (D) RIP assay showed the abundance of SPRY2
enriched by Ago2 or IgG in SRA01/04 cells transfected with NC
mimics or miR-124 mimics. (E) RT-qPCR was performed to determine
miR-124 expression in SRA01/04 cells transfected with miR-124
mimics or NC mimics. (F) RT-qPCR assay showed the expression of
SPRY2 in SRA01/04 cells transfected with NC mimics, miR-124 mimics,
NC inhibitor or miR-124 inhibitor. *P<0.05 vs. control group.
miR, microRNA; RT-qPCR, reverse transcription-quantitative PCR; NC,
negative control; SPRY2, protein sprouty homolog 2; wt, wild-type;
mut, mutant; RIP, RNA immunoprecipitation; Ago2, argonaute 2; IgG,
immunoglobin G. |
miR-124 facilitates SRA01/04 cell
apoptosis by targeting SPRY2
RT-qPCR demonstrated that the expression of SPRY2
was decreased in ARC (Fig. 3A). To
determine whether miR-124 exerts its function by regulating SPRY2,
SRA01/04 cells were transfected with shSPRY2, shNC, pcDNA3.1,
pcDNA3.1/SPRY2 and pcDNA3.1/SPRY2 + miR-124 mimics. Efficacy
analysis demonstrated that SPRY2 mRNA and protein expression levels
were significantly decreased in SRA01/04 cells transfected with
shSPRY2 (Fig. 3B). Silencing of
SPRY2 suppressed SRA01/04 cell viability (Fig. 3C). Moreover, the TUNEL assay
determined that knockdown of SPRY2 promoted ARC cell apoptosis
(Fig. 3D). Furthermore, SPRY2 mRNA
and protein expression levels were increased in SRA01/04 cells
transfected with pcDNA3.1/SPRY2 compared with the pcDNA3.1 group
(Fig. 3E). In addition,
overexpression of SPRY2 increased SRA01/04 cell viability, whereas
overexpression of miR-124 reversed its promoting effect on cell
viability (Fig. 3F). The results of
the TUNEL assay indicated that overexpression of SPRY2 decreased
ARC cell apoptosis, the effects of which were reversed following
overexpression of miR-124 (Fig.
3G). Taken together, these results suggested that miR-124
regulated SRA01/04 cell functions by targeting SPRY2.
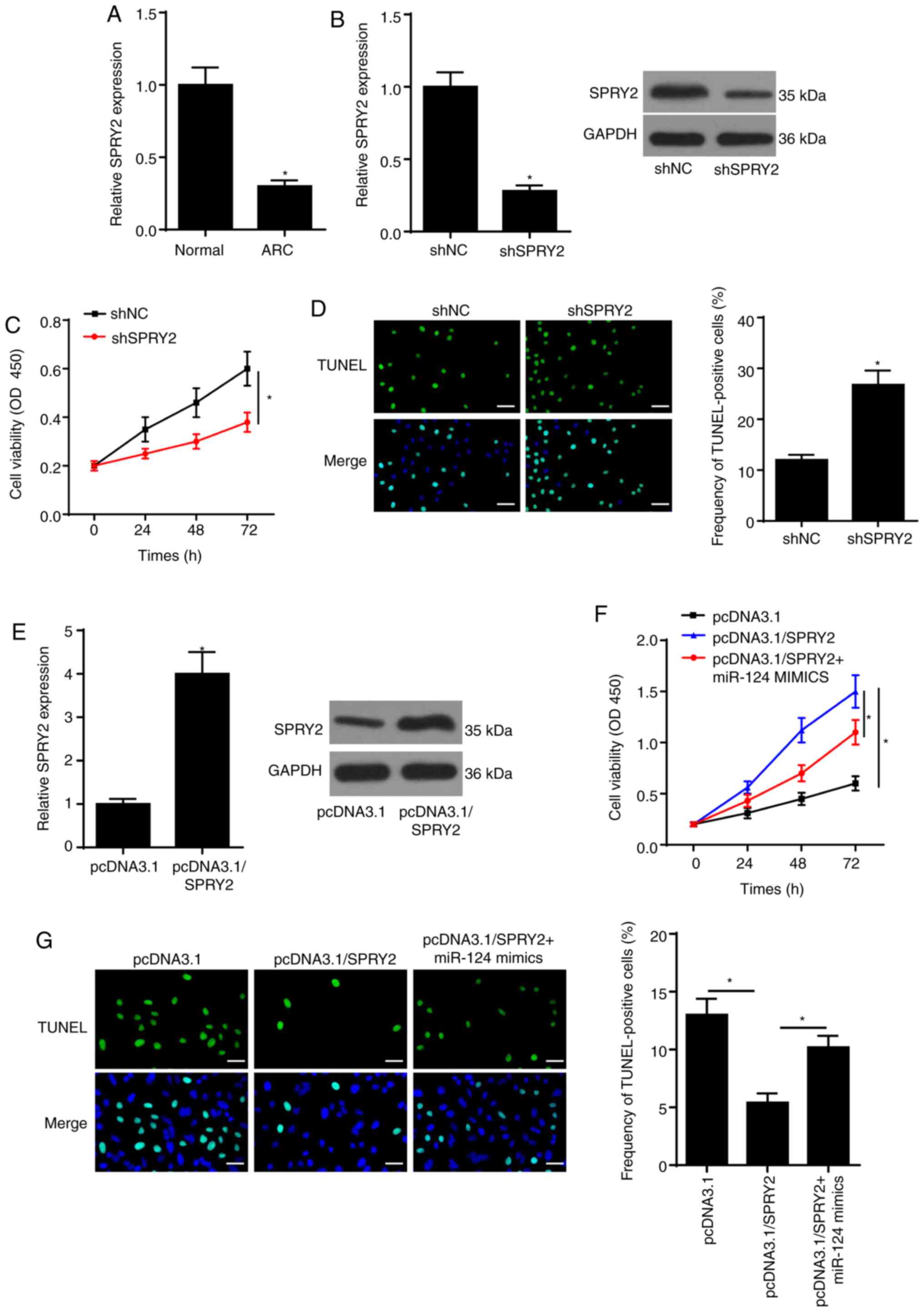 | Figure 3.miR-124 increases SRA01/04 cell
apoptosis by targeting SPRY2. (A) RT-qPCR assay was employed to
assess SPRY2 expression in ARC. (B) RT-qPCR and western blotting
assays showed SPRY2 expression in SRA01/04 cells transfected with
shSPRY2 or shNC. (C) CCK-8 assay showed the viability of SRA01/04
cells transfected with shSPRY2 or shNC. (D) TUNEL assay
(magnification, ×200; scale bar, 50 µm) showed the apoptosis of
SRA01/04 cells transfected with shSPRY2 or shNC. (E) RT-qPCR and
western blotting assays showed SPRY2 expression in SRA01/04 cells
transfected with pcDNA3.1/SPRY2 or pcDNA3.1. (F) CCK-8 assay showed
the proliferation of SRA01/04 cells transfected with pcDNA3.1,
pcDNA3.1/SPRY2, pcDNA3.1/SPRY2 + miR-124 mimics. (G) TUNEL assay
(magnification, ×200; scale bar, 50 µm) showed the apoptosis of
SRA01/04 cells transfected with pcDNA3.1, pcDNA3.1/SPRY2,
pcDNA3.1/SPRY2 + miR-124 mimics. *P<0.05 vs. control group. miR,
microRNA; RT-qPCR, reverse transcription-quantitative PCR; NC,
negative control; SPRY2, protein sprouty homolog 2; sh-, short
hairpin RNA; CCK-8, Cell Counting Kit-8; ARC, age-related
cataract. |
MMP-2 is also targeted by miR-124
StarBase was used to predict the binding site
between miR-124 and MMP-2 (Fig.
4A). The dual-luciferase reporter assay indicated that the
overexpression of miR-124 decreased the luciferase activity of
wt-MMP-2 in SRA01/04 cells; however, no significant differences
were observed in the luciferase activity of mut-MMP-2 (Fig. 4B and C). These effects were reversed
following miR-124 knockdown. In addition, MMP-2 was enriched by
Ago2 antibody in SRA01/04 cells transfected with miR-124 mimics
(Fig. 4D). RT-qPCR analysis
demonstrated that overexpression of miR-124 decreased MMP-2
expression, while transfection with miR-124 inhibitor elevated
MMP-2 expression in SRA01/04 cells (Fig. 4E). Collectively, these results
suggested that miR-124 targeted MMP-2 in SRA01/04 cells.
miR-124 depends on MMP-2 to regulate
SRA01/04 cell apoptosis
To determine the function of MMP-2 in ARC
development, RT-qPCR was performed to detect MMP-2 expression in
ARC tissues. As presented in Fig.
5A, MMP-2 expression was downregulated in ARC tissues. To
further determine whether MMP-2 is a vital effector in
miR-124-mediated ARC progression, SRA01/04 cells were transfected
with shMMP-2, shNC, pcDNA3.1, pcDNA3.1/MMP-2 and pcDNA3.1/MMP-2 +
miR-124 mimics. Transfection of shMMP-2 significantly decreased
MMP-2 expression in SRA01/04 cells, at both the mRNA and protein
levels (Fig. 5B). Then, the CCK-8
assay showed that MMP-2 knockdown inhibited SRA01/04 cell viability
(Fig. 5C). On the contrary,
transfection with shMMP-2 promoted ARC cell apoptosis (Fig. 5D). Moreover, overexpression of MMP-2
significantly increased MMP-2 expression in SRA01/04 cells, at both
the mRNA and protein levels (Fig.
5E). The results of the CCK-8 assay confirmed that miR-124 can
abolish the promotion of MMP-2 overexpression-induced SRA01/04 cell
viability (Fig. 5F). In addition,
transfection with pcDNA3.1/MMP-2 attenuated ARC cell apoptosis, the
effects of which were reversed following overexpression of miR-124
(Fig. 5G). Taken together, these
results suggested that MMP-2 was involved in miR-124-mediated
SRA01/04 cell apoptosis.
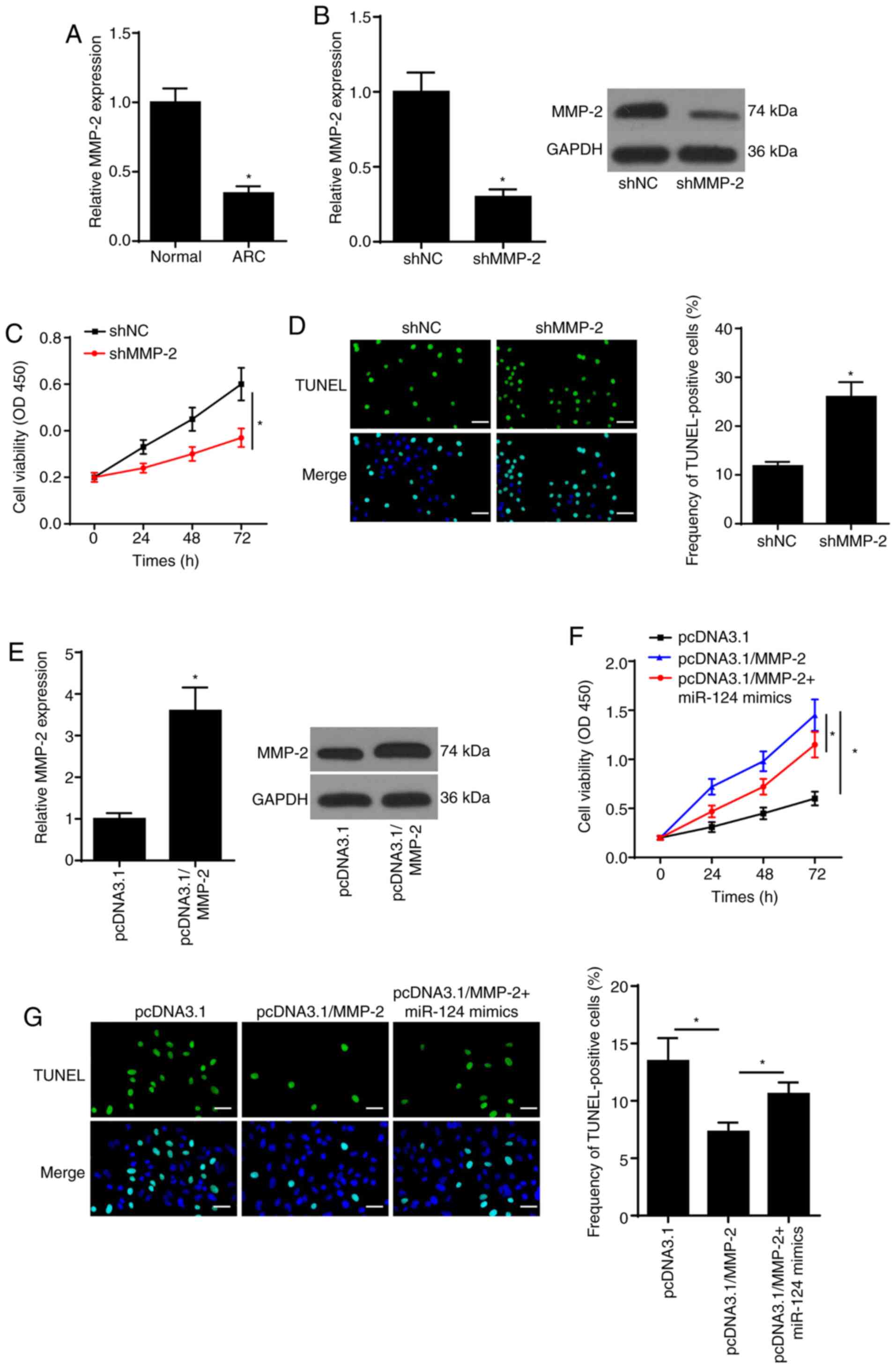 | Figure 5.miR-124 depends on MMP-2 to regulate
SRA01/04 cell apoptosis. (A) RT-qPCR was used to analyze MMP-2
expression in ARC tissues. (B) RT-qPCR and western blotting assays
were performed to determine MMP-2 expression in SRA01/04 cells
transfected with shMMP-2 or shNC. (C) CCK-8 assay showed the
proliferation of SRA01/04 cells transfected with shMMP-2 or shNC.
(D) TUNEL assay (magnification, ×200; scale bar, 50 µm) showed the
cell apoptosis of SRA01/04 cells transfected with shMMP-2 or shNC.
(E) RT-qPCR and western blotting assays were conducted to measure
MMP-2 expression in SRA01/04 cells transfected with pcDNA3.1/MMP-2
or pcDNA3.1. (F) CCK-8 assay showed the proliferation of SRA01/04
cells transfected with pcDNA3.1, pcDNA3.1/MMP-2, pcDNA3.1/MMP-2 +
miR-124 mimics. (G) TUNEL assay (magnification, ×200; scale bar, 50
µm) showed the apoptosis of SRA01/04 cells transfected with
pcDNA3.1, pcDNA3.1/MMP-2, pcDNA3.1/MMP-2 + miR-124 mimics.
*P<0.05 vs. control group. ARC, age-related cataract; miR,
microRNA; RT-qPCR, reverse transcription-quantitative PCR; NC,
negative control; MMP-2, matrix metalloproteinase-2; sh-, short
hairpin RNA; CCK-8, Cell Counting Kit-8. |
Discussion
ARC is one of the most common disorders of the lens
worldwide, which leads to reversible blindness (23,24).
To the best of our knowledge, the present study was the first to
investigate the role of miR-124 in lens epithelial cell apoptosis,
and the results demonstrated the interactions between miR-124 and
SPRY2 or MMP-2 in SRA01/04 cells. Taken together, these results
provided a novel insight into the process of ARC formation.
Increasing evidence suggests that miR-124
participates in several cellular processes, such as proliferation,
metastasis, autophagy and apoptosis (25–27).
For example, nuclear enriched abundant transcript 1 modulates the
viability and apoptosis of retinoblastoma cells by targeting
miR-124 (28). Furthermore,
silencing of lncRNA XIST suppresses cell proliferation and induces
cell apoptosis in retinoblastoma via the miR-124/STAT3 axis
(29). The results of the present
study demonstrated that miR-124 was upregulated in ARC tissues. The
effect of miR-124 on ARC cell viability and apoptosis was also
assessed, and the results indicated that miR-124 inhibition
promoted SRA01/04 cell viability, but suppressed cell
apoptosis.
SPRY2 is a member of the Sprouty family, which
exerts essential roles in cellular viability, differentiation and
apoptosis (30,31). A recent study reported that miR-23b
increases the viability, migration and EMT of lens epithelial cells
by targeting SPRY2 (32). The
results of the present study demonstrated that SPRY2 expression
decreased in ARC, and SPRY2 was confirmed to be a downstream target
of miR-124. The results of the CCK-8 and TUNEL assays indicated
that SPRY2 facilitated SRA01/04 cell viability and inhibited cell
apoptosis, the effects of which were reversed following
transfection with miR-124 mimics. Taken together, these results
suggested that miR-124 induced apoptosis of SRA01/04 cells by
targeting SPRY2.
MMP-2, a member of the MMP family, has been reported
to participate in cell processes, including cell proliferation and
metastasis (33,34). The results of the present study
demonstrated that MMP-2 was downregulated in ARC tissues, and
miR-124 inhibited MMP-2 expression by direct interaction. In
addition, rescue experiments demonstrated that overexpression of
MMP-2 promoted SRA01/04 cell viability, which was attenuated
following overexpression of miR-124. Conversely, overexpression of
miR-124 reversed the inhibitory effect of overexpressed MMP-2 on
SRA01/04 cell apoptosis.
In conclusion, the results of the present study
demonstrated that miR-124 expression was elevated in the lens
capsule of ARC tissues, and miR-124 regulated SRA01/04 cell
viability and apoptosis by targeting SPRY2 and MMP-2. Collectively,
these results provided a novel potential therapeutic target for
ARC. However, the present study primarily focused on in
vitro experiments and the number of clinical samples used was
limited. Further in vivo experiments should be performed
using an increased number of clinical samples.
Acknowledgements
Not applicable.
Funding
The present study was supported by the Youth Project
of Jiangsu Natural Science Foundation (grant no. BK20190162), the
Applied Basic Research Plan of Changzhou (grant no. CJ20190090),
the Applied Basic Research Plan of Changzhou (grant no. CJ20180067)
and Young Talent Development Plan of Changzhou Health Commission
(2020–233) (grant no. CZQM20200027).
Availability of data and materials
The datasets used and/or analyzed during the present
study are available from the corresponding author upon reasonable
request.
Authors' contributions
YaoL and XL conceived and designed the study. YanL,
SL and QZ performed the experiments. YaoL and YanL analyzed the
data and drafted the manuscript. SL and XL revised the manuscript.
All authors read and approved the final manuscript.
Ethics approval and consent to
participate
The present study was approved by the Ethics
Committee of the First People's Hospital of Changzhou (Changzhou,
China) and all patients provided written informed consent prior to
the study.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Nangia V, Jonas JB, George R, Lingam V,
Ellwein L, Cicinelli MV, Das A, Flaxman SR, Keeffe JE, Kempen JH,
et al Vision Loss Expert Group of the Global Burden of Disease
Study, : Prevalence and causes of blindness and vision impairment:
Magnitude, temporal trends and projections in South and Central
Asia. Br J Ophthalmol. 103:871–877. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Jin C, Chen X, Law A, Kang Y, Wang X, Xu W
and Yao K: Different-sized incisions for phacoemulsification in
age-related cataract. Cochrane Database Syst Rev.
9:CD0105102017.PubMed/NCBI
|
|
3
|
Tang Y, Wang X, Wang J, Huang W, Gao Y,
Luo Y, Yang J and Lu Y: Prevalence of age-related cataract and
cataract surgery in a Chinese adult population: The Taizhou Eye
Study. Invest Ophthalmol Vis Sci. 57:1193–1200. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Milazzo S, Grenot M and Benzerroug M:
Posterior capsule opacification. J Fr Ophtalmol. 37:825–830.
2014.(In French). View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Mathias RT, White TW and Gong X: Lens gap
junctions in growth, differentiation, and homeostasis. Physiol Rev.
90:179–206. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Belusko PB, Nakajima T, Azuma M and
Shearer TR: Expression changes in mRNAs and mitochondrial damage in
lens epithelial cells with selenite. Biochim Biophys Acta.
1623:135–142. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Sun T, Li MY, Li PF and Cao JM: MicroRNAs
in cardiac autophagy: Small molecules and big role. Cells.
7:E1042018. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Wang J, Haubrock M, Cao KM, Hua X, Zhang
CY, Wingender E and Li J: Regulatory coordination of clustered
microRNAs based on microRNA-transcription factor regulatory
network. BMC Syst Biol. 5:1992011. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Yu X, Zheng H, Chan MT and Wu WKK:
MicroRNAs: New players in cataract. Am J Transl Res. 9:3896–3903.
2017.PubMed/NCBI
|
|
10
|
Li ZN, Ge MX and Yuan ZF: MicroRNA-182-5p
protects human lens epithelial cells against oxidative
stress-induced apoptosis by inhibiting NOX4 and p38 MAPK
signalling. BMC Ophthalmol. 20:2332020. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Zhang GB, Liu ZG, Wang J and Fan W: MiR-34
promotes apoptosis of lens epithelial cells in cataract rats via
the TGF-β/Smads signaling pathway. Eur Rev Med Pharmacol Sci.
24:3485–3491. 2020.PubMed/NCBI
|
|
12
|
Liu Y, Li H and Liu Y: microRNA-378a
regulates the reactive oxygen species (ROS)/phosphatidylinositol
3-kinases (PI3K)/AKT signaling pathway in human lens epithelial
cells and cataract. Med Sci Monit. 25:4314–4321. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Zhou W, Xu J, Wang C, Shi D and Yan Q:
miR-23b-3p regulates apoptosis and autophagy via suppressing SIRT1
in lens epithelial cells. J Cell Biochem. 120:19635–19646. 2019.
View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Liu S, Hu C, Wang Y, Shi G, Li Y and Wu H:
miR-124 inhibits proliferation and invasion of human retinoblastoma
cells by targeting STAT3. Oncol Rep. 36:2398–2404. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Kuracha MR, Burgess D, Siefker E, Cooper
JT, Licht JD, Robinson ML and Govindarajan V: Spry1 and Spry2 are
necessary for lens vesicle separation and corneal differentiation.
Invest Ophthalmol Vis Sci. 52:6887–6897. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Zhang Y, Yuan F, Liu L, Chen Z, Ma X, Lin
Z and Zou J: The role of the miR-21/SPRY2 axis in modulating
proangiogenic factors, epithelial phenotypes, and wound healing in
corneal epithelial cells. Invest Ophthalmol Vis Sci. 60:3854–3862.
2019. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Shin EH, Basson MA, Robinson ML, McAvoy JW
and Lovicu FJ: Sprouty is a negative regulator of transforming
growth factor β-induced epithelial-to-mesenchymal transition and
cataract. Mol Med. 18:861–873. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Tan X, Zhu Y, Chen C, Chen X, Qin Y, Qu B,
Luo L, Lin H, Wu M, Chen W, et al: Sprouty2 suppresses
epithelial-mesenchymal transition of human lens epithelial cells
through blockade of Smad2 and ERK1/2 pathways. PLoS One.
11:e01592752016. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Webb AH, Gao BT, Goldsmith ZK, Irvine AS,
Saleh N, Lee RP, Lendermon JB, Bheemreddy R, Zhang Q, Brennan RC,
et al: Inhibition of MMP-2 and MMP-9 decreases cellular migration,
and angiogenesis in in vitro models of retinoblastoma. BMC Cancer.
17:4342017. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Awasthi N, Wang-Su ST and Wagner BJ:
Downregulation of MMP-2 and −9 by proteasome inhibition: A possible
mechanism to decrease LEC migration and prevent posterior capsular
opacification. Invest Ophthalmol Vis Sci. 49:1998–2003. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Chylack LT Jr, Wolfe JK, Singer DM, Leske
MC, Bullimore MA, Bailey IL, Friend J, McCarthy D and Wu SY; The
Longitudinal Study of Cataract Study Group, : The Lens Opacities
Classification System III. Arch Ophthalmol. 111:831–836. 1993.
View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) Method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Thrimawithana TR, Rupenthal ID, Räsch SS,
Lim JC, Morton JD and Bunt CR: Drug delivery to the lens for the
management of cataracts. Adv Drug Deliv Rev. 126:185–194. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Wei M, Xing KY, Fan YC, Libondi T and Lou
MF: Loss of thiol repair systems in human cataractous lenses.
Invest Ophthalmol Vis Sci. 56:598–605. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Fu W, Wu X, Yang Z and Mi H: The effect of
miR-124-3p on cell proliferation and apoptosis in bladder cancer by
targeting EDNRB. Arch Med Sci. 15:1154–1162. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Yan G, Li Y, Zhan L, Sun S, Yuan J, Wang
T, Yin Y, Dai Z, Zhu Y, Jiang Z, et al: Decreased miR-124-3p
promoted breast cancer proliferation and metastasis by targeting
MGAT5. Am J Cancer Res. 9:585–596. 2019.PubMed/NCBI
|
|
27
|
Huang J, Yang Y, Fang F and Liu K: MALAT1
modulates the autophagy of retinoblastoma cell through
miR-124-mediated stx17 regulation. J Cell Biochem. 119:3853–3863.
2018. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Wang L, Yang D, Tian R and Zhang H: NEAT1
promotes retinoblastoma progression via modulating miR-124. J Cell
Biochem. 120:15585–15593. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Hu C, Liu S, Han M, Wang Y and Xu C:
Knockdown of lncRNA XIST inhibits retinoblastoma progression by
modulating the miR-124/STAT3 axis. Biomed Pharmacother.
107:547–554. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Taketomi T, Onimura T, Yoshiga D, Muratsu
D, Sanui T, Fukuda T, Kusukawa J and Nakamura S: Sprouty2 is
involved in the control of osteoblast proliferation and
differentiation through the FGF and BMP signaling pathways. Cell
Biol Int. 42:1106–1114. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Zhao Q, Chen S, Zhu Z, Yu L, Ren Y, Jiang
M, Weng J and Li B: miR-21 promotes EGF-induced pancreatic cancer
cell proliferation by targeting Spry2. Cell Death Dis. 9:11572018.
View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Liu W, Yang Y, Yan J and Wang L:
MicroRNA-23b-3p promotes the proliferation, migration, and
epithelial-mesenchymal transition of lens epithelial cells by
targeting Sprouty2. Acta Histochem. 121:704–711. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Sun MX, Yu F, Gong ML, Fan GL and Liu CX:
Effects of curcumin on the role of MMP-2 in endometrial cancer cell
proliferation and invasion. Eur Rev Med Pharmacol Sci.
22:5033–5041. 2018.PubMed/NCBI
|
|
34
|
Wang X, Yang B, She Y and Ye Y: The lncRNA
TP73-AS1 promotes ovarian cancer cell proliferation and metastasis
via modulation of MMP2 and MMP9. J Cell Biochem. 119:7790–7799.
2018. View Article : Google Scholar : PubMed/NCBI
|