Introduction
Osteoarthritis (OA) is a common joint disease that
affects 50% of the population aged >65 years, and 12% of
individuals aged >25 years (1).
The primary pathological mechanisms of OA are progressive
inflammation of chondrocytes, extracellular matrix (ECM)
degradation, loss of articular cartilage, proliferation of
subchondral bone and formation of osteophytes, which result in the
loss of joint function (2,3). The risk factors for OA include aging,
history of joint injury, obesity, sex and genetic and anatomical
factors (4). However, the exact
molecular pathogenesis of OA remains unclear. Considering that
articular cartilage has no self-repair capability and its
homeostasis is precisely regulated by a series of factors, for
example signal transduction (5),
cytokines (6) and hormones
(7), understanding the homeostatic
mechanism of chondrocytes in the process of OA is of significance
for the development of effective therapies for OA.
OA was previously considered to be a
non-inflammatory joint disease, but more recent studies have
suggested that inflammatory mechanisms are involved in the
pathological process of OA chondrocytes (8,9).
Inflammatory factors, such as IL-1β, IL-6 and TNF-α, in the
microenvironment of the joint cavity may decrease the viability of
chondrocytes, and increase necrosis and apoptosis (6). Elevated expression levels of MMPs
degrade the ECM of chondrocytes, which is accompanied by an active
inflammatory response (10).
Additionally, a disintegrin and metalloproteinase with
thrombospondin motifs (Adamts) family of proteins, which are also
known as aggrecanases, exert a proteoglycan/aggrecan (Acan)
depletion effect that is associated with cartilage degradation and
inflammation during OA (11). The
effects of certain genetic molecules on OA have been determined.
For example, inhibiting the expression of m6A methyltransferase
complex including methyltransferase-like 3 decreases inflammation
and apoptosis of OA chondrocytes, which slows progression of OA
(12). Overexpression or activation
of silent mating type information regulation 2 homolog 1 decreases
loss of cartilage by exerting an inhibitory effect on MMP13
(13). In addition, Adamts-5 is
inhibited by microRNA-137, and this decreases levels of
inflammatory factors and ECM degradation in OA (14). Therefore, investigation of the
molecular function of OA chondrocytes would improve understanding
of the molecular pathology of OA, and may highlight potential novel
therapeutic targets.
3-Phosphoglycerate dehydrogenase (Phgdh) is a key
enzyme involved in serine biosynthesis and serves as a
rate-limiting enzyme in the conversion from 3-phosphoglycerate to
serine. The serine released during this process provides a large
amount of energy and metabolites for cell growth and metabolism
(15,16). Elevated levels of Phgdh have been
observed in several types of cancer, including colon (17), breast (18) and cervical cancer (19), which suggests that overexpression of
Phgdh is associated with a poor prognosis in these types of cancer.
Moreover, a previous study suggested that Phgdh-deficient mouse
embryonic fibroblasts are more vulnerable to oxidative damage,
accompanied by an increase in levels of inflammatory factors
(16). These findings suggest that
upregulation of Phgdh expression may increase cell viability and
proliferation, whereas downregulation of Phgdh may result in
oxidative damage and the promotion of an inflammatory response. To
the best of our knowledge, however, the role of Phgdh in OA
chondrocytes has not yet been studied.
In the present study, the expression levels of Phgdh
in OA chondrocytes were assessed and its biological effects on
chondrocytes were determined. Phgdh levels were assessed using both
an in vitro and in vivo model. Next, the effect of
Phgdh on ECM synthesis, inflammation, apoptosis and oxidative
stress levels were determined. The present study aimed to improve
understanding of the pathogenesis of OA and highlight potentially
novel therapeutic targets for the management of OA.
Materials and methods
Affymetrix microarray analysis
In order to determine the levels of Phgdh in the
chondrocytes in an OA rat model, the microarray dataset GSE42295
[(Rat230_2) Affymetrix Rat Genome 230 2.0 Array] from Gene
Expression Omnibus was used. There were a total of 12 samples,
including 3 cases of surgically induced 2 or 8 week rat OA models,
and corresponding sham controls, which underwent surgical incision
without structural modification. The differentially expressed genes
(DEG) were identified between the sham- and OA-2 and 8 week groups
using the GEO2R tool (20).
Criteria for classification as a DEG were P<0.05 and a |log2Fold
Change (FC)|>1.
Establishment of the in vitro OA
model, and isolation and culture of chondrocytes
A total of 36 male newborn (weight, 5–6 g)
Sprague-Dawley rats, which were all reared at room temperature
under 12/12 h day/night cycles, were purchased from the
Experimental Animal Centre of the Kunming Medical University
(Kunming, China) for chondrocyte extraction. Briefly, after rats
were sacrificed by cervical dislocation without anesthesia,
articular cartilage was harvested from the knee joints. The
cartilage tissue was cut into 1–3 mm3 pieces followed by
digestion with 2 mg/ml collagenase II (Sigma-Aldrich; Merck KGaA)
for 3 h at 37°C. Finally, the digested chondrocytes were suspended
in DMEM (Gibco; Thermo Fisher Scientific, Inc.) supplemented with
1% penicillin/streptomycin (Beijing Solarbio Science &
Technology Co., Ltd.) and 10% FBS (Zhejiang Tianhang Biotechnology
Co., Ltd.) at 37°C with 5% CO2 in a humidified
incubator. The chondrocytes adhered to the plate after 2–3 days of
culture, at which point the tissue pieces were discarded and the
medium replaced. The cells were cultured for three passages for
chondrocyte identification and use in subsequent experiments. IL-1β
(10 ng/ml; Beijing Solarbio Science & Technology Co., Ltd.) was
used to stimulate chondrocytes to establish the in vitro OA
model as previously described (21). The present study was approved by the
Ethics Committee of Qujing First People's Hospital (approval no.
19-025).
Chondrocyte identification and
immunofluorescence analysis
Immunofluorescence analysis was performed for
chondrocyte identification and Phgdh detection in IL-1β-induced
chondrocytes. Collagen type II α 1 chain (Col2a1), a specific
marker for chondrocytes, was utilized for chondrocyte
identification via immunofluorescence assay. Briefly, third
generation chondrocytes and chondrocytes from the Control and IL-1β
groups were harvested, washed using PBS and fixed using 4%
paraformaldehyde for 30 min at room temperature. After blocking
with 5% BSA (Boster Biological Technology) for 30 min at room
temperature, primary antibodies against Col2a1 (1:900; cat. no.
28459-1-AP; ProteinTech Group, Inc.) and Phgdh (1:1,000; cat. no.
14719-1-AP; ProteinTech Group, Inc.) were used to incubate the
cells at 4°C overnight. The following day, the primary antibody was
removed and the cells were incubated with the FITC-conjugated mouse
anti-rabbit IgG (1:5,000; cat. no. BM2012; Boster Biological
Technology) for 1 h at room temperature. After staining the cell
nuclei with DAPI (1:10,000; Beijing Solarbio Science &
Technology Co., Ltd.) for 5 min at room temperature, images were
obtained using a fluorescence microscope (magnification, ×100;
Olympus Corporation).
Cell transfection
The expression plasmid encoding the full-length open
reading frame of rat Phgdh with EGFP tags (pIRES2-EGFP-Phgdh) and
the corresponding negative control plasmid were synthesized and
purchased from Shanghai GenePharma Co., Ltd. For cell transfection,
chondrocytes were seeded into six-well plates at a density of
5×106 cells per well and cultured overnight at 37°C with
5% CO2 in a humidified incubator. When confluence
reached 70–80% density, Lipofectamine® 3000 (Invitrogen;
Thermo Fisher Scientific, Inc.) reagent was used for transfection
according to the manufacturer's protocol. The mixture contained
plasmids and transfection reagents (1 µg: 3 µl) and cell
transfection with the plasmid was performed at a final
concentration of 50 nM; cells were incubated with the transfection
mixture and Opti-MEM (Gibco; Thermo Fisher Scientific, Inc.) for
6–7 h at 37°C with 5% CO2 in a humidified incubator.
Subsequently, DMEM supplemented with 10% FBS was used to culture
cells for 48 h. The experiments were grouped as follows: Control
(chondrocytes without any treatment); IL-1β (chondrocytes treated
with IL-1β); IL-1β + pcDNA (chondrocytes treated with IL-1β and
transfected with negative control plasmid) and Il-1β + Phgdh
(chondrocytes treated with IL-1β and transfected with Phgdh cDNA
plasmid).
Cell viability assay
Chondrocytes were seeded into 96-well plates at a
density of 8×103 cells per well and cultured for 24, 48
or 72 h. Cell Counting Kit-8 (CCK-8; Dojindo Molecular
Technologies, Inc.) assay was used to measure cell viability.
Briefly, 10 µl CCK-8 solution was added to the wells and cultured
for 2 h. The absorbance was then measured at 450 nm using a
microplate reader (Thermo Fisher Scientific, Inc.).
Safranin O staining
In order to detect the deposition of
glycosaminoglycans (GAGs) from chondrocytes, safranin O staining
kit (Beijing Solarbio Science & Technology Co., Ltd.) was used
to perform the safranin O staining assay according to the
manufacturer's protocol. Cells were harvested and fixed using 95%
ethanol for 30 min at room temperature, followed by washing three
times with PBS. Next, 0.1% safranin O (Beijing Solarbio Science
& Technology Co., Ltd.) solution was used to stain the cells
for 10–15 min at room temperature. Staining was observed using a
light microscope (magnification, ×100; Olympus Corporation) and
images were captured.
GAGs detection
Treated chondrocytes were incubated with 60 µg/ml
proteinase K (Sigma-Aldrich; Merck KGaA) for 10 h at 56°C and the
digested aliquot was used to detect GAGs and DNA content.
Dimethylmethylene blue (DMMB) dye binding experiment was used to
measure the GAGs, as previously described (22), and the absorbance was measured at
525 nm using a microplate reader (Thermo Fisher Scientific, Inc.).
Hoechst 33258 (1 µg/ml; Sigma-Aldrich; Merck KGaA) was used to
incubate samples at room temperature for 5 min and measured at 460
nm on a microplate fluorescence reader (FLx800; BioTek Instruments,
Inc.) to detect DNA content. After the levels of GAGs and DNA
content were measured separately, production of GAGs was expressed
as the GAG/DNA ratio.
Reverse transcription-quantitative
(RT-q)PCR
RT-qPCR was performed to quantify gene expression
levels of Col2a1, Acan, sex determining region Y-box 9 (Sox9),
Col1a1, catalase (Cat) and superoxide dismutase 1 (Sod1). The
sequences of the primers used are listed in Table I. Total RNA from differently treated
cells was extracted using a total RNA isolation kit (Megentec),
according to the manufacturer's protocol. A RT kit (Takara Bio,
Inc.) was used to reverse transcribe RNA into cDNA, according to
the manufacturer's protocols. qPCR was performed with a Fast Start
Universal SYBR Green Master Mix (Roche Diagnostics) using a light
Cycler 96 system (Roche Diagnostics). The thermocycling conditions
were as follows: 10 min at 95°C; followed by 40 cycles of 95°C for
10 sec and 60°C for sec. Relative gene expression levels were
normalized to GAPDH and calculated using the 2−ΔΔCq
method (23).
 | Table I.Primer sequences used for reverse
transcription-quantitative PCR. |
Table I.
Primer sequences used for reverse
transcription-quantitative PCR.
| Gene | Forward primer,
5′→3′ | Reverse primer,
5′→3′ |
|---|
| Phgdh |
GAACCCTGCCTAGTCACTGGA |
CCTTAGTAGCTGACCGGACG |
| Sox9 |
TCCAGCAAGAACAAGCCACA |
CGAAGGGTCTCTTCTCGCTC |
| Acan |
GAATGGGAGCCAGCCTACAC |
GAGAGGCAGAGGGACTTTCG |
| Col2a1 |
ATTGCCTACCTGGACGAAGC |
GACAGGCCCTATGTCCACAC |
| Col1a1 |
GCTTCACCTACAGCGTCACT |
AAGCCGAATTCCTGGTCTGG |
| Catalase |
AGAGGAAACGCCTGTGTGAG |
TAGTCAGGGTGGACGTCAGT |
| Sod1 |
ATTCACTTCGAGCAGAAGGCA |
ATTGCCCAGGTCTCCAACAT |
| GAPDH |
CTATAAATTGAGCCCGCAGC |
ACCAAATCCGTTGACTCCG |
ELISA
The medium of chondrocytes following treatment was
collected to investigate the levels of pro-inflammatory cytokines
(IL-6 and TNF-α) using specific ELISA kits (cat. nos. PR6000B and
PRTA00; R&D Systems, Inc.), according to the manufacturer's
protocol. The absorbance at 450 nm was measured using a microplate
reader (Thermo Fisher Scientific, Inc.).
Apoptosis assay
Apoptosis of chondrocytes following treatment was
investigated using an apoptosis kit (Beijing Solarbio Science &
Technology Co., Ltd.). Chondrocytes were stained with 10 µl Annexin
V-FITC and 10 µl PI solution in the dark for 10 min at room
temperature. Next, cells were rinsed, resuspended in PBS and
analyzed using a flow cytometer (BD FACSCalibur™; BD Biosciences)
and FlowJo software (version 7; FlowJo LLC).
Measurement of production of reactive
oxygen species (ROS)
The levels of ROS in chondrocytes were determined
using a 2′-7′-dichlorodihydrofluorescein-diacetate (DCFH-DA) kit
(Beyotime Institute of Biotechnology). Briefly, serum-free medium
containing DCFH-DA (10 µM) was used to incubate the cells at 37°C
for 30 min in the dark. Next, the cells were digested, resuspended
in PBS and analyzed using a flow cytometer (BD Biosciences) and
FlowJo software (version 7).
Western blot assay
Western blot analysis was used to assess the protein
expression levels of Phgdh, Sox9, Acan, Col2a1, Col1a1, MMP13,
Adamts-5, Bcl2, Bax, cleaved caspase-3, Kelch like ECH associated
protein 1 (Keap1) and Nuclear factor erythroid 2-related factor 2
(Nrf2). Total protein from differently treated cells was extracted
using RIPA Lysis Buffer (Boster Biological Technology) supplemented
with 1 mM phenylmethylsulfonyl fluoride (Boster Biological
Technology). The concentrations of protein were detected using a
BCA kit (Nanjing Jiancheng Bioengineering Institute). A total of 50
µg protein from each group was loaded on a 10% SDS gel, resolved
using SDS-PAGE and transferred to PVDF membranes (Biosharp Life
Sciences). Membranes were blocked using 5% non-fat milk for 1 h at
room temperature. Subsequently, membranes were incubated with
primary antibodies against Phgdh (1:1,000; cat. no. 14719-1-AP;
ProteinTech Group, Inc.), Sox9 (1:1,000; cat. no. 67439-1-Ig;
ProteinTech Group, Inc.), Acan (1:1,000; cat. no. 13880-1-AP;
ProteinTech Group, Inc.), Col2a1 (1:900; cat. no. 28459-1-AP;
ProteinTech Group, Inc.), Col1a1 (1:5,000; cat. no. 67288-1-Ig;
ProteinTech Group, Inc.), MMP13 (1:1,000; cat. no. 18165-1-AP;
ProteinTech Group, Inc.), Adamts-5 (1:1,000; cat. no. ab41037;
Abcam), cleaved caspase-3 (1:1,000; cat. no. 9664S; CST Biological
Reagents Co., Ltd.), Bcl2 (1:1,000; cat. no. 3498; CST Biological
Reagents Co., Ltd.), Bax (1:1,000; cat. no. 5023; CST Biological
Reagents Co., Ltd.), Keap1 (1:1,000; cat. no. 8047; CST Biological
Reagents Co., Ltd.), Nrf2 (1:1,000; cat. no. 12721; CST Biological
Reagents Co., Ltd.) and β-actin (1:5,000; cat. no. 66009-1-Ig;
ProteinTech Group, Inc.). Membranes were incubated with the primary
antibodies overnight at 4°C followed by incubation with
HRP-conjugated secondary antibody (1:10,000; cat. no. 7074; CST
Biological Reagents Co., Ltd.) for 1 h at room temperature. ECL kit
(Beijing Solarbio Science & Technology Co., Ltd.) was used to
visualize the protein membranes. Signals were visualized using an
Odyssey Infrared Imaging System (LI-COR Biosciences). Densitometry
was performed with ImageJ software (version 1.8.0.112; National
Institutes of Health).
Statistical analysis
Data were analyzed using SPSS version 22.0 (IBM
Corp.) and are presented as the mean ± SD (n=3). All experiments
were repeated three times. An unpaired Student's t-test was used to
compare differences between two groups and one-way ANOVA followed
by Tukey's multiple comparisons post hoc test was used to compare
differences between ≥3 groups. P<0.05 was considered to indicate
a statistically significant difference.
Results
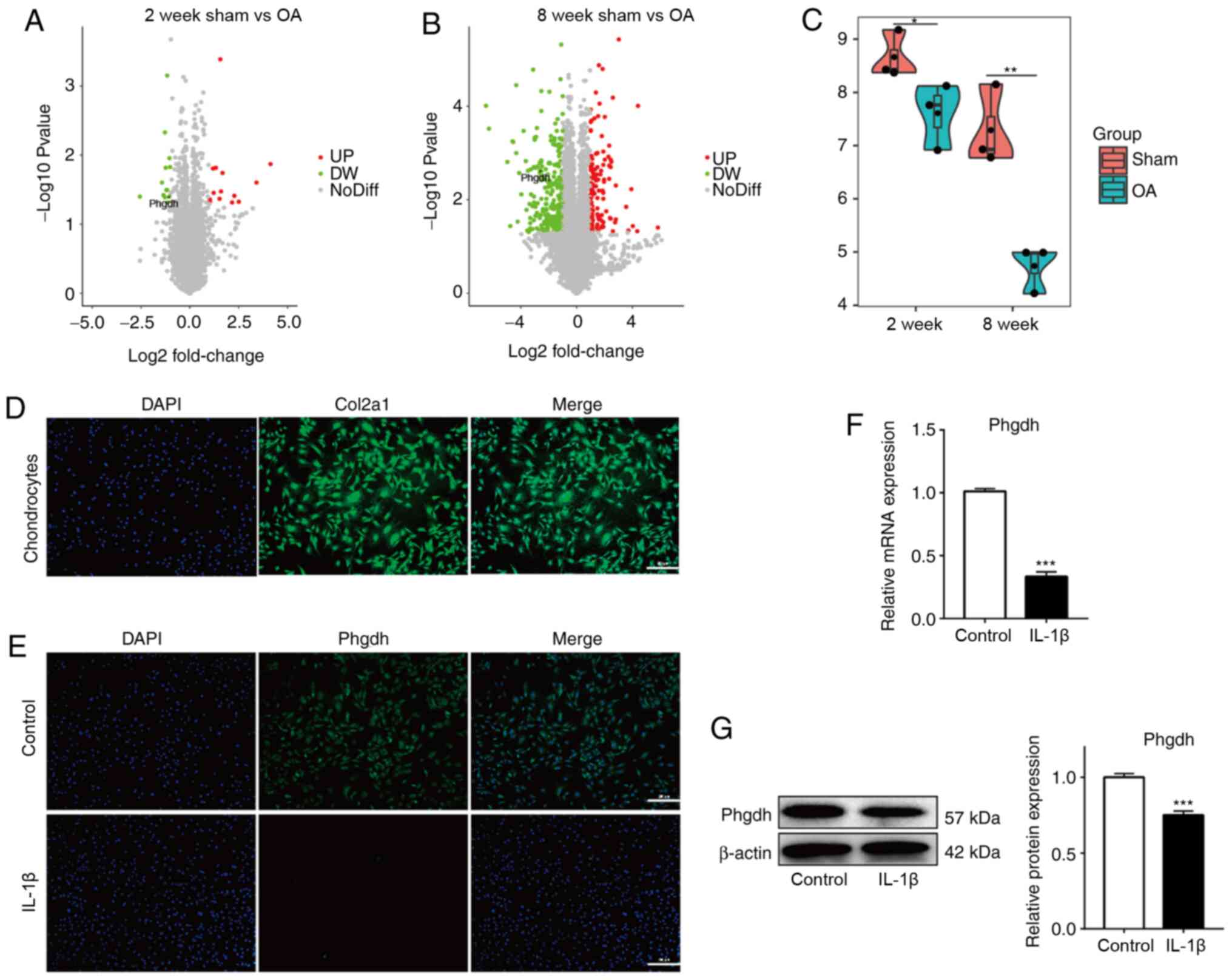
Phgdh expression is decreased in
chondrocytes both in vivo and in an in vitro OA model
First, the expression of Phgdh was investigated in
chondrocytes in an in vivo model of OA using microarray
analysis. In both the 2 and 8 week OA in vivo model, Phgdh
expression was significantly decreased compared with the
corresponding sham group (Fig. 1A and
B). Downregulation of Phgdh was greatest in the 8 week OA model
(Fig. 1C), suggesting that
downregulation of Phgdh may be associated with the severity of
OA.
Next, chondrocytes from newborn rats were extracted.
Successful harvesting was demonstrated by immunofluorescence
analysis to detect expression of Col2a1 (Fig. 1D). IL-1β treatment was used to
establish the in vitro OA model. Phgdh expression was
detected using immunofluorescence staining (Fig. 1E), RT-qPCR (Fig. 1F) and western blotting (Fig. 1G) assays. Phgdh expression in
chondrocytes was significantly decreased following IL-1β treatment.
These results suggest that Phgdh expression in chondrocytes was
decreased in OA.
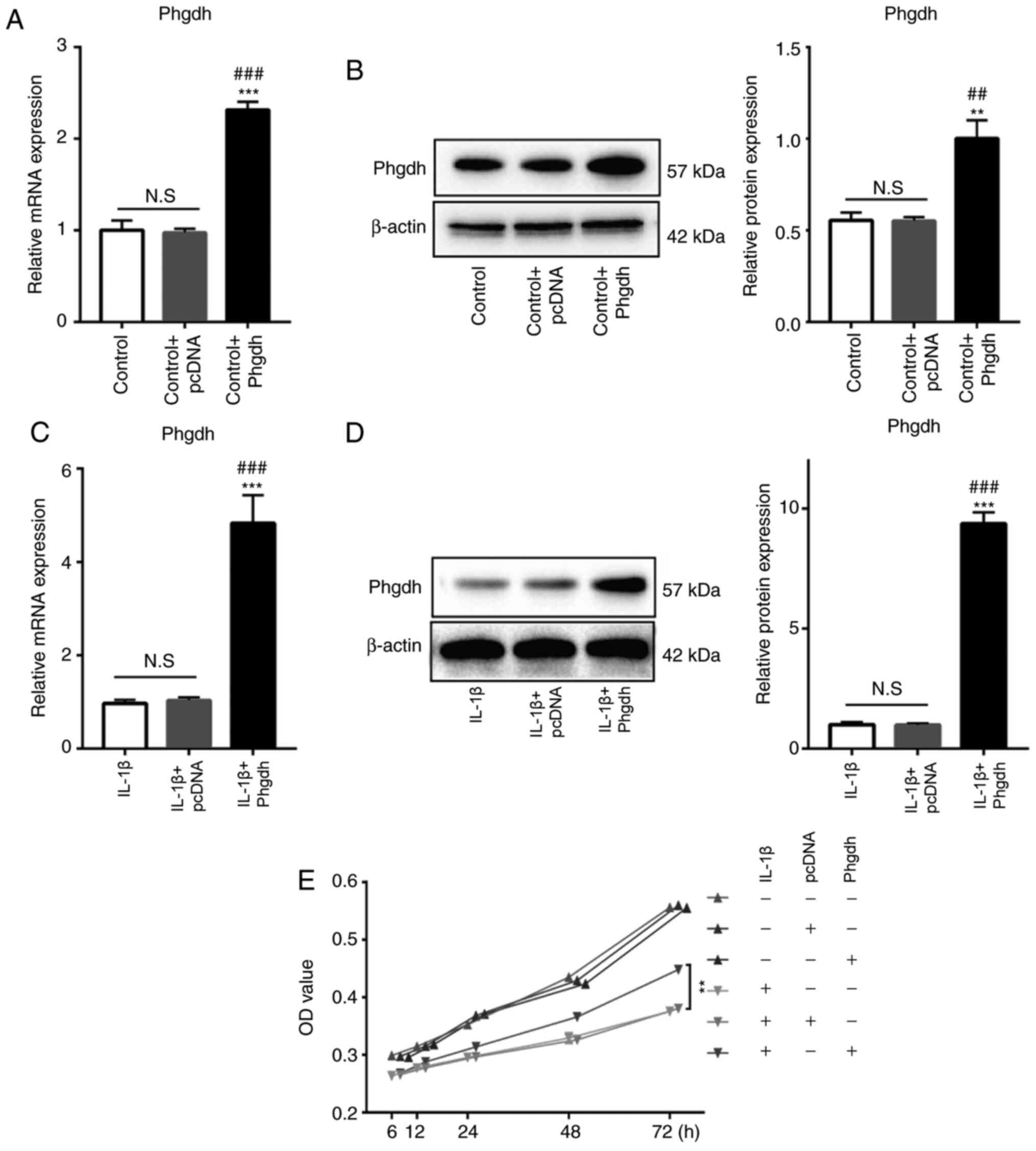
Phgdh increases the viability of
chondrocytes treated with IL-1β
In order to assess the biological effect of Phgdh on
chondrocytes, a Phgdh cDNA overexpression plasmid was used to
transfect the chondrocytes. Phgdh mRNA and protein expression
levels in chondrocytes in the presence or absence of IL-1β
treatment were significantly increased following Phgdh cDNA
transfection, whereas the negative control plasmid exhibited no
significant effect on Phgdh expression (Fig. 2A-D). CCK-8 assay was used to assess
cell viability; the results suggested that Phgdh cDNA and negative
control plasmid had no significant effect on the viability of
chondrocytes in untreated cells. However, overexpression of Phgdh
increased cell viability of chondrocytes treated with IL-1β
(Fig. 2E).
Phgdh alleviates IL-1β-induced
chondrocyte degeneration
The degeneration of chondrocytes primarily manifests
as decreased levels of GAGs and decreased expression of
chondrogenic-specific genes, including Col2a1, Sox9 and Acan
(24). Safranin-O staining showed
that chondrocytes in the IL-1β + Phgdh group exhibited increased
GAG staining compared with the IL-1β group (Fig. 3A). Moreover, the DMMB assay also
confirmed that GAG levels were increased in the IL-1β + Phgdh group
compared with the IL-1β group (Fig.
3B). The expression levels of cartilage-specific genes were
consistent with the aforementioned results; Sox9 (Fig. 3C), Acan (Fig. 3D) and Col2a1 (Fig. 3E) expression levels were increased
in the IL-1β + Phgdh group compared with the IL-1β group, whereas
Col1a1 expression levels were decreased (Fig. 3F). The protein expression levels of
Sox9, Acan, Col2a1 and Col1a1 were also assessed. Sox9, Acan,
Col2a1 protein expression levels were upregulated in the IL-1β +
Phgdh group compared with the IL-1β group, whereas Col1a1
expression levels were decreased (Fig.
3G). There results suggested that overexpression of Phgdh
alleviated IL-1β-induced chondrocyte degeneration.
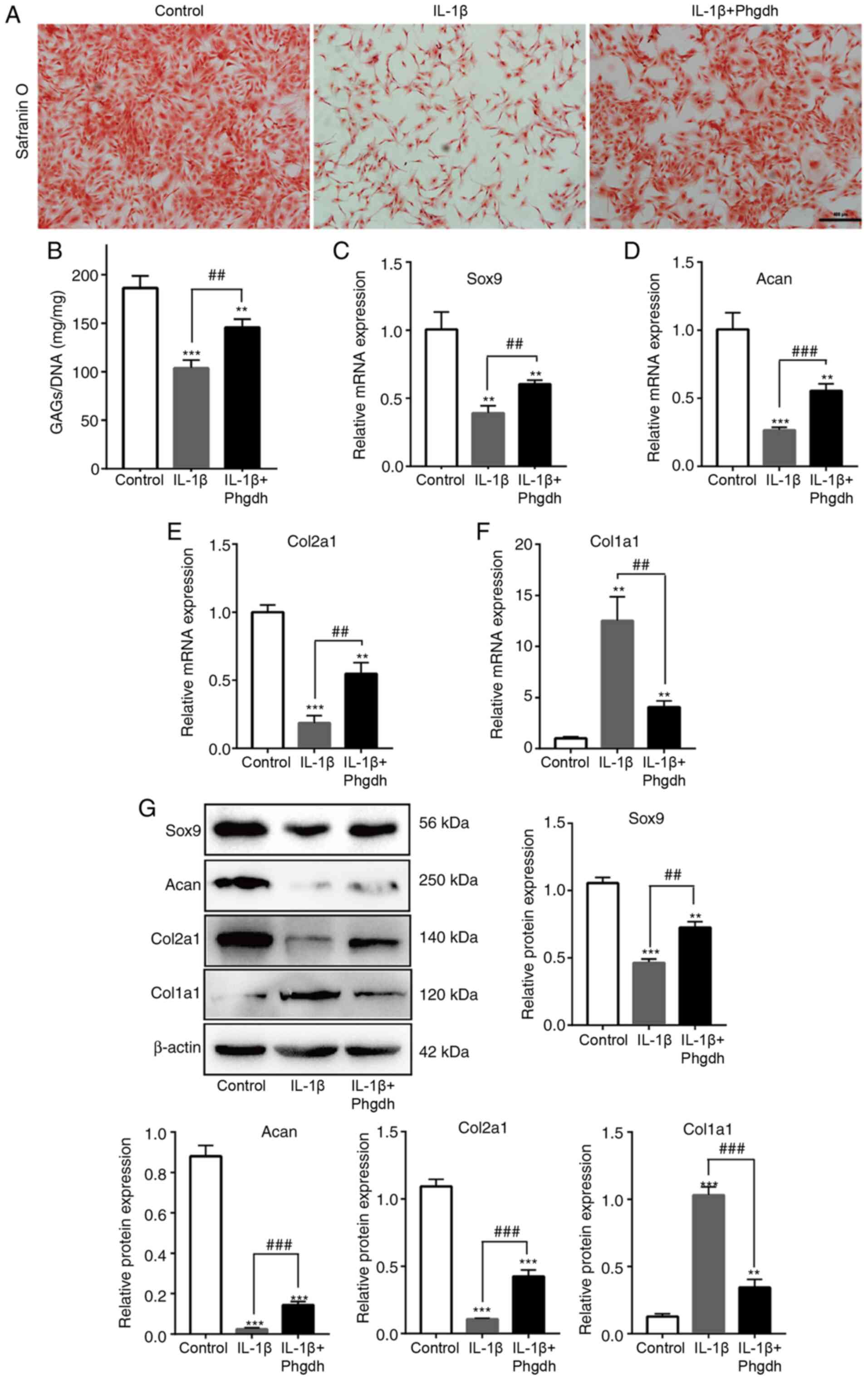 | Figure 3.Phgdh alleviates IL-1β-induced
chondrocyte degeneration. (A) GAG secretion in the different groups
was determined using Safranin O staining. Scale bar, 200 µm. (B)
Quantitative analysis of GAGs, normalized to DNA (in mg). mRNA
expression levels of (C) Sox9, (D) Acan, (E) Col2a1 and (F) Col1a1.
(G) Protein expression levels of Sox9, Acan, Col2a1 and Col1a1.
Data are presented as the mean ± SD (n=3). **P<0.01,
***P<0.001 vs. Control; ##P<0.01,
###P<0.001 vs. IL-1β. Phgdh, 3-phosphoglycerate
dehydrogenase; GAG, glycosaminoglycan; Sox9, sex determining region
Y-box 9; Acan, aggrecan; Col2a1, collagen type II α 1 chain;
Col1a1, collagen type I α 1 chain. |
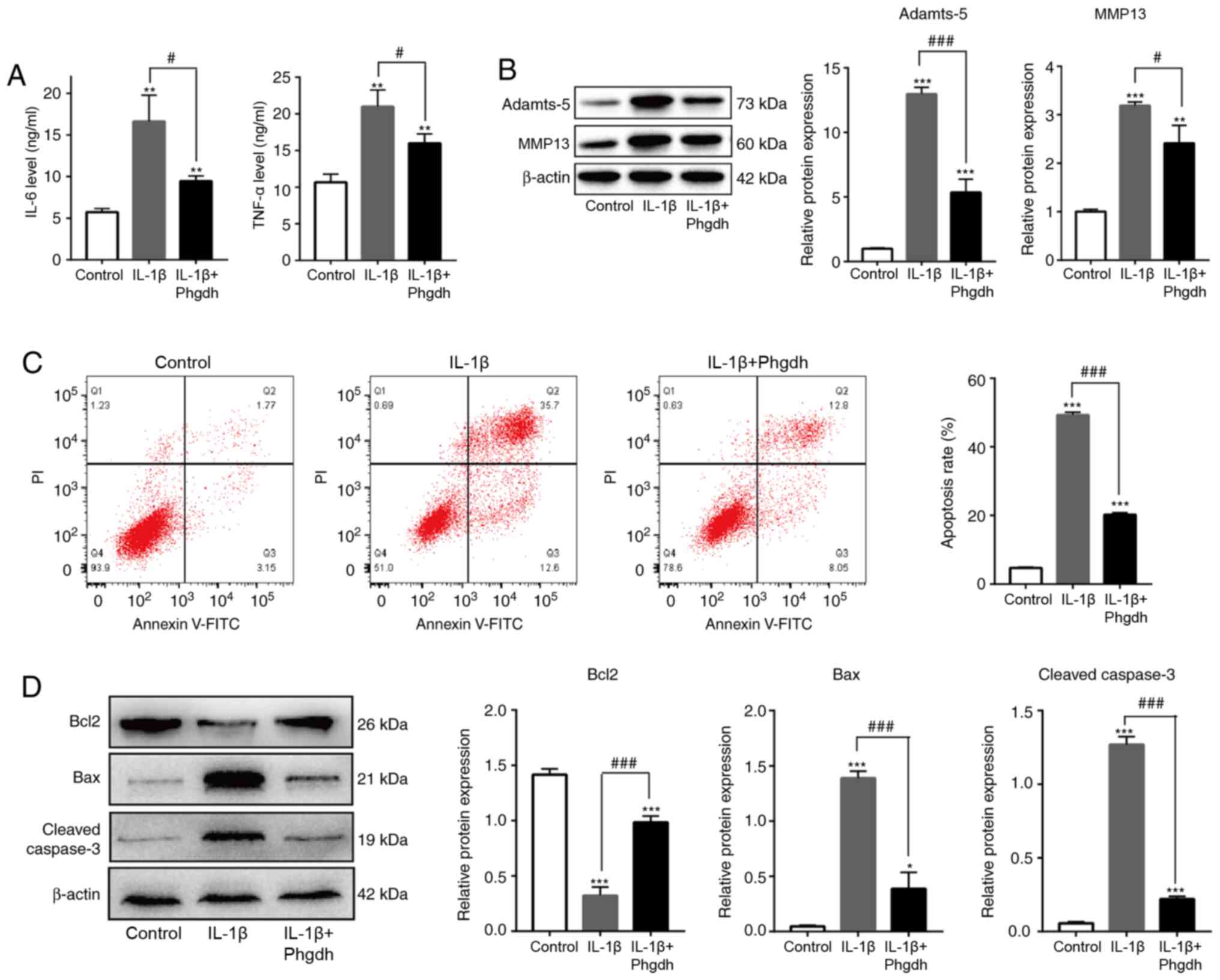
Phgdh decreases IL-1β-induced
chondrocyte inflammation and apoptosis
Production of inflammatory factors and expression of
matrix-degrading enzymes results in chondrocyte degeneration
(25). The effect of Phgdh on
chondrocyte inflammation and apoptosis was investigated. Using
ELISA, it was shown that the levels of key inflammatory factors in
OA, such as Il-6 and TNF-α, were decreased in the IL-1β + Phgdh
group compared with the IL-1β group (Fig. 4A). The protein expression levels of
Adamts-5 and MMP13 were also significantly decreased in the Il-1β +
Phgdh group (Fig. 4B). Moreover,
the apoptotic rate of cells in the IL-1β + Phgdh group was lower
than that in the IL-1β group (Fig.
4C). Expression of apoptosis-associated proteins, such as
cleaved caspase-3, Bax and Bcl2, were detected. Expression of the
pro-apoptotic proteins cleaved caspase-3 and Bax was decreased and
that of the anti-apoptotic protein Bcl2 was elevated in the IL-1β +
Phgdh group compared with the IL-1β group (Fig. 4D). These results suggested that
overexpression of Phgdh alleviated inflammation and apoptosis of
chondrocytes treated with IL-1β.
Phgdh decreases oxidative stress
damage of chondrocytes treated with IL-1β
Evidence has suggested that an imbalance in
oxidative stress is associated with inflammation (26). Studies have also shown that
inhibiting oxidative stress levels and improving the antioxidant
capacity of cells may decrease inflammation and degeneration of
chondrocytes (26,27). In the present study, it was shown
that overexpression of Phgdh decreased ROS levels of chondrocytes
treated with IL-1β (Fig. 5A). The
mRNA expression levels of the anti-oxidant enzymes Cat and Sod1
were measured. The results suggested that Phgdh increased the
levels of Cat and Sod1 (Fig. 5B).
Key proteins associated with oxidative stress include Nrf2 and
Keap1, and their expression was next determined. Overexpression of
Phgdh increased expression levels of Nrf2 and decreased those of
Keap1 (Fig. 5C). These results
suggested that Phgdh may exhibit a regulatory effect on the
oxidative stress levels of chondrocytes via the Nrf2/Keap1
axis.
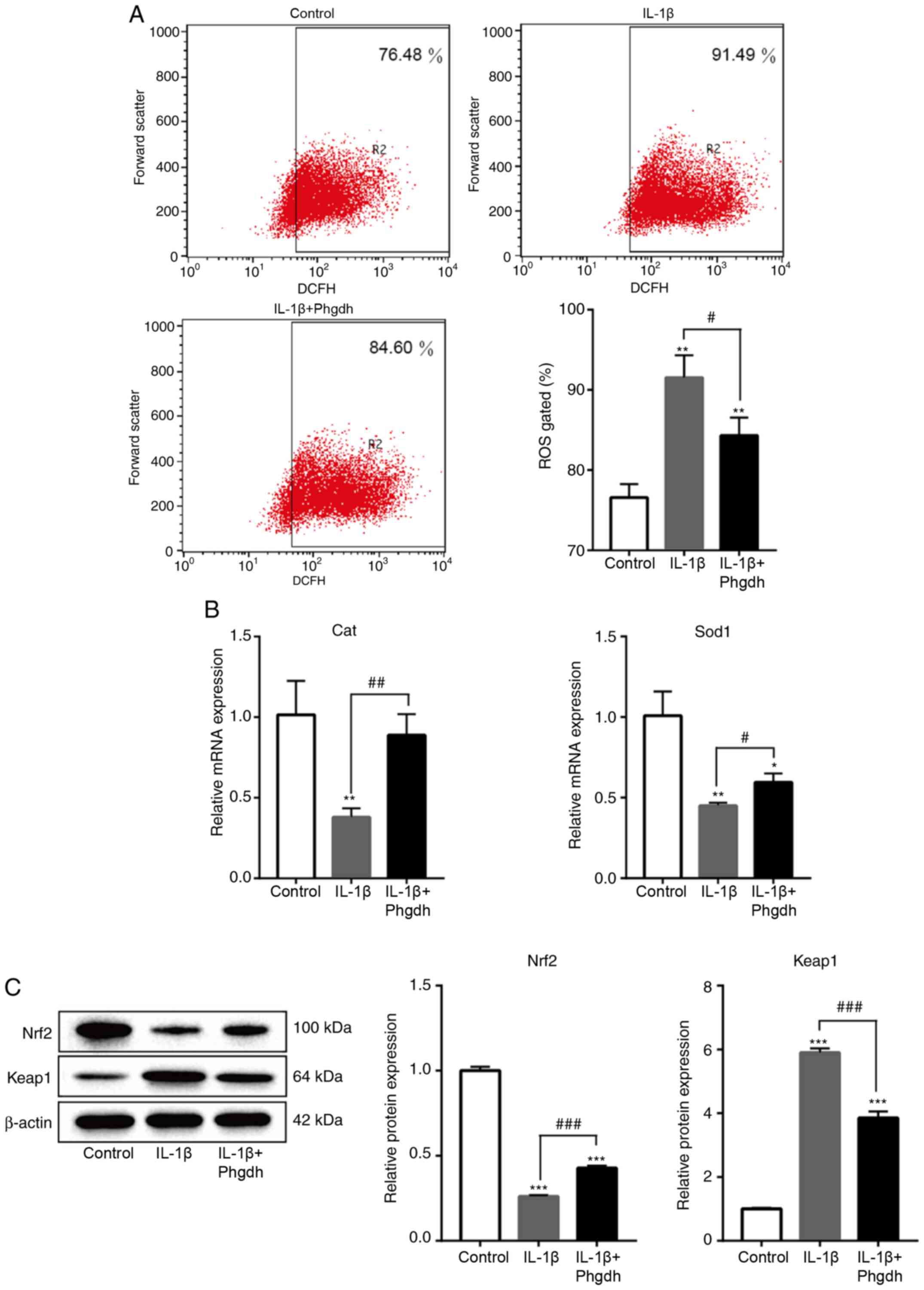 | Figure 5.Phgdh decreases oxidative stress
damage of chondrocytes treated with IL-1β. (A) Flow cytometry
analysis for ROS levels. (B) mRNA expression levels of Cat and
Sod1. (C) Protein expression levels of Nrf2 and Keap1 protein. Data
are presented as the mean ± SD (n=3). *P<0.05, **P<0.01,
***P<0.001 vs. Control; #P<0.05,
##P<0.01, ###P<0.001 vs. IL-1β. Phgdh,
3-phosphoglycerate dehydrogenase; ROS, reactive oxygen species;
Cat, catalase; Sod1, superoxide dismutase; Keap1, Kelch like ECH
associated protein 1; Nrf2, nuclear factor erythroid 2-related
factor 2. |
Discussion
OA is the most common disease of joints and is
characterized by damaged articular cartilage, which is primarily
composed of chondrocytes and cartilage matrix (1). The primary function of chondrocytes is
to secrete ECM proteins, such as Acan and Col2a1, to maintain
homeostasis of the articular cartilage (28). Alterations to chondrocyte function
are accompanied by degeneration of cartilage matrix, which
eventually results in the initiation and progression of OA.
Protecting or rescuing the function of chondrocytes may thus assist
in alleviating the progress of OA (29). Although a number of potential
targets and mechanisms of OA chondrocytes have been reported, these
studies have not translated into clinically useful therapeutic
options due to a lack of sensitivity and specificity (30–33).
Therefore, investigating the novel pathogenesis of OA chondrocytes
may highlight potential molecules for targeted therapy of OA. In
the present study, Phgdh was found to be downregulated in OA
chondrocytes and decreased with the progression of OA.
Overexpression of Phgdh prevented IL-1β-induced chondrocyte
degeneration, inflammation, apoptosis and oxidative damage. Thus,
the potential effect of Phgdh on OA chondrocytes was highlighted
and the results suggested that Phgdh may be a promising therapeutic
target for management of OA.
Phgdh is a key enzyme involved in serine
biosynthesis, where it synthesizes serine to provide large amounts
of energy and metabolites for cell growth and metabolism (34). In tumors, upregulated expression of
Phgdh has been detected in lung, breast, pancreatic and colorectal
cancer, where it is positively associated with cell proliferation,
migration, invasion and poor prognosis (17,18,35,36).
However, downregulation of Phgdh is also associated with certain
non-tumor diseases. For example, decreased Phgdh expression,
accompanied by low serine levels, is associated with the
development of fatty liver disease (37). Moreover, Phgdh deficiency is also a
risk factor for the development of Macular Telangiectasia type 2
development, an uncommon bilateral retinal disease (38). These aforementioned studies suggest
that the expression of Phgdh is carefully regulated, and its
abnormal expression can result in pathophysiological changes or
disease. In the present study, Phgdh expression was shown to be
decreased in the chondrocytes of OA and with the progression of OA
over time, suggesting that Phgdh was also involved in OA
pathogenesis.
Under normal physiological conditions, the activity
of synthesis and decomposition of cartilage matrix is in a dynamic
balance. Transcription factor Sox9 is a key molecule in maintaining
the phenotype of the chondrocyte (39). Sox9 activates a series of downstream
signaling molecules to promote the deposition of Col2a1, Acan and
GAGs (40). In patients with OA,
numerous inflammatory factors, such as IL-1β, IL-6 and TNF-α,
inhibit the expression of Sox9 and degrade cartilage ECM by
activating a series of proteases, such as MMPs and Adamts (41). Decreasing the level of inflammation
and restoring the homeostasis of chondrocytes are key to preventing
the progression of OA (42,43). In the present study, in chondrocytes
treated with IL-1β, it was observed that inflammatory stimulation
decreased the levels of GAGs, and overexpression of Phgdh restored
the deposition of GAGs. Furthermore, overexpression of Phgdh offset
the inhibitory effect of IL-1β on Sox9, Acan, and Col2a1 expression
in chondrocytes and decreased expression levels of the
osteogenesis-specific gene Col1a1. In addition, upregulation of
Phgdh decreased expression levels of IL-6, TNF-α, MMP13 and
Adamts-5 and inhibited apoptosis of chondrocytes treated with
IL-1β. These results suggested that Phgdh exerted a protective
effect on chondrocyte homeostasis, inflammation and apoptosis.
Oxidative stress is a negative outcome that results
from production of free radicals in tissue, and is a key factor for
mediating inflammation (44).
Oxidative stress damage is primarily the result of increased ROS
production and decreased levels of antioxidant enzymes, such as Sod
and Cat (45). Oxidative stress is
a key factor is associated with the progression of OA. Excessive
ROS production activates the inflammatory response via the NF-κB
signaling pathway and inhibits synthesis of proteoglycans and ECM
in chondrocytes (46). In addition,
a lack of Phgdh increases the vulnerability of fibroblasts to
oxidative stress damage (16). In
the present study, Phgdh exerted a regulatory effect on oxidative
stress levels of chondrocytes in OA. Phgdh decreased the levels of
ROS and increased expression levels of Cat and Sod1 in chondrocytes
treated with IL-1β. Nrf2 is a major antioxidant factor, which
normally binds to Keap1 (47).
Under oxidative damage, Nrf2 dissociates from Keap1 and
translocates to the nucleus to initiate the transcription of
antioxidant enzymes (48). It has
previously been shown that promoting the expression of Nrf2
decreases the inflammatory response and progression of OA (43). In the present study, overexpression
of Phgdh promoted the expression levels of Nrf2 and decreased those
of Keap1 in chondrocytes treated with IL-1β, suggesting that Phgdh
regulated oxidative stress levels of chondrocytes in OA via
Nrf2.
In conclusion, Phgdh was decreased in OA and was
associated with OA progression. Moreover, Phgdh was found to be
involved in chondrocyte homeostasis: Overexpression of Phgdh
decreased inflammation and apoptosis and restored the phenotype of
chondrocytes in OA. Additionally, overexpression of Phgdh
alleviated oxidative stress damage; this may have involved a
Keap1-Nrf2 axis which is involved in the pathological mechanism of
OA. Therefore, Phgdh may be a potentially significant target for OA
research and treatment.
Acknowledgements
The authors would like to thank Dr Li Hongwei
(Biological Laboratory of Qujing First People's Hospital; Qujing,
China) for their guidance on the experimental design and
technology.
Funding
No funding was received.
Availability of data and materials
The microarray dataset is available in the GSE42295
from Gene Expression Omnibus Database. The data collected and
analyzed during the current study are available from the
corresponding author upon reasonable request.
Authors' contributions
JW and XQ conceptualized and designed the study. HH,
KL and HO collected and analyzed data. HH and KL wrote the
manuscript. HH, KL, HO, HQ and JW confirm the authenticity of all
the raw data. All authors read and approved the final
manuscript.
Ethics approval and consent to
participate
The present study was approved by the Ethics
Committee of Qujing First People's Hospital (approval no. 19-025;
Qujing, China).
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
Glossary
Abbreviations
Abbreviations:
|
OA
|
osteoarthritis
|
|
Phgdh
|
3-phosphoglycerate dehydrogenase
|
|
ECM
|
extracellular matrix
|
|
Adamts-5
|
a disintegrin and metalloproteinase
with thrombospondin motifs 5
|
|
Cat
|
catalase
|
|
Sod1
|
superoxide dismutase 1
|
|
ROS
|
reactive oxygen species
|
|
Keap1
|
Kelch like ECH associated protein
1
|
|
Nrf2
|
nuclear factor erythroid 2-related
factor 2
|
|
Sox9
|
sex determining region Y-box 9
|
|
Acan
|
aggrecan
|
|
Col2a1
|
collagen type II α 1 chain
|
|
Col1a1
|
collagen type I α 1 chain
|
References
|
1
|
Hayami T, Pickarski M, Zhuo Y, Wesolowski
GA, Rodan GA and Duong LT: Characterization of articular cartilage
and subchondral bone changes in the rat anterior cruciate ligament
transection and meniscectomized models of osteoarthritis. Bone.
38:234–243. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Sellam J and Berenbaum F: The role of
synovitis in pathophysiology and clinical symptoms of
osteoarthritis. Nat Rev Rheumatol. 6:625–635. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Hawker GA: Osteoarthritis is a serious
disease. Clin Exp Rheumatol. 37 (Suppl 120):S3–S6. 2019.
|
|
4
|
Johnson VL and Hunter DJ: The epidemiology
of osteoarthritis. Best Pract Res Clin Rheumatol. 28:5–15. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Xie Y, Zinkle A, Chen L and Mohammadi M:
Fibroblast growth factor signalling in osteoarthritis and cartilage
repair. Nat Rev Rheumatol. 16:547–564. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Wojdasiewicz P, Poniatowski Ł A and
Szukiewicz D: The role of inflammatory and anti-inflammatory
cytokines in the pathogenesis of osteoarthritis. Mediators Inflamm.
2014:5614592014. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Savvidou O, Milonaki M, Goumenos S, Flevas
D, Papagelopoulos P and Moutsatsou P: Glucocorticoid signaling and
osteoarthritis. Mol Cell Endocrinol. 480:153–166. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Shen J, Abu-Amer Y, O'Keefe RJ and
McAlinden A: Inflammation and epigenetic regulation in
osteoarthritis. Connect Tissue Res. 58:49–63. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Houard X, Goldring MB and Berenbaum F:
Homeostatic mechanisms in articular cartilage and role of
inflammation in osteoarthritis. Curr Rheumatol Rep. 15:3752013.
View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Xu K, Ma C, Xu L, Ran J, Jiang L, He Y,
Adel Abdo Moqbel S, Wang Z and Wu L: Polygalacic acid inhibits MMPs
expression and osteoarthritis via Wnt/β-catenin and MAPK signal
pathways suppression. Int Immunopharmacol. 63:246–252. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Verma P and Dalal K: ADAMTS-4 and
ADAMTS-5: Key enzymes in osteoarthritis. J Cell Biochem.
112:3507–3514. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Liu Q, Li M, Jiang L, Jiang R and Fu B:
METTL3 promotes experimental osteoarthritis development by
regulating inflammatory response and apoptosis in chondrocyte.
Biochem Biophys Res Commun. 516:22–27. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Elayyan J, Lee EJ, Gabay O, Smith CA, Qiq
O, Reich E, Mobasheri A, Henrotin Y, Kimber SJ and Dvir-Ginzberg M:
LEF1-mediated MMP13 gene expression is repressed by SIRT1 in human
chondrocytes. FASEB J. 31:3116–3125. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Zhang Y, Wang G, Ma L, Wang C, Wang L, Guo
Y and Zhao X: miR-137 suppresses cell growth and extracellular
matrixdegradation through regulating ADAMTS-5 in chondrocytes. Am J
Transl Res. 11:7027–7034. 2019.PubMed/NCBI
|
|
15
|
Sayano T, Kawakami Y, Kusada W, Suzuki T,
Kawano Y, Watanabe A, Takashima K, Arimoto Y, Esaki K, Wada A, et
al: L-serine deficiency caused by genetic Phgdh deletion leads to
robust induction of 4E-BP1 and subsequent repression of translation
initiation in the developing central nervous system. FEBS J.
280:1502–1517. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Hamano M, Haraguchi Y, Sayano T, Zyao C,
Arimoto Y, Kawano Y, Moriyasu K, Udono M, Katakura Y, Ogawa T, et
al: Enhanced vulnerability to oxidative stress and induction of
inflammatory gene expression in 3-phosphoglycerate
dehydrogenase-deficient fibroblasts. FEBS Open Bio. 8:914–922.
2018. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Jia XQ, Zhang S, Zhu HJ, Wang W, Zhu JH,
Wang XD and Qiang JF: Increased expression of PHGDH and prognostic
significance in colorectal cancer. Transl Oncol. 9:191–196. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Samanta D, Park Y, Andrabi SA, Shelton LM,
Gilkes DM and Semenza GL: PHGDH expression is required for
mitochondrial redox homeostasis, breast cancer stem cell
maintenance, and lung metastasis. Cancer Res. 76:4430–4442. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Jing Z, Heng W, Aiping D, Yafei Q and
Shulan Z: Expression and clinical significance of phosphoglycerate
dehydrogenase and squamous cell carcinoma antigen in cervical
cancer. Int J Gynecol Cancer. 23:1465–1469. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Barrett T, Wilhite SE, Ledoux P,
Evangelista C, Kim IF, Tomashevsky M, Marshall KA, Phillippy KH,
Sherman PM, Holko M, et al: NCBI GEO: Archive for functional
genomics data sets-update. Nucleic Acids Res. 41:D991–D995. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Cheng F, Hu H, Sun K, Yan F and Geng Y:
miR-455-3p enhances chondrocytes apoptosis and inflammation by
targeting COL2A1 in the in vitro osteoarthritis model. Biosci
Biotechnol Biochem. 84:695–702. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Guingamp C, Gegout-Pottie P, Philippe L,
Terlain B, Netter P and Gillet P: Mono-iodoacetate-induced
experimental osteoarthritis: A dose-response study of loss of
mobility, morphology, and biochemistry. Arthritis Rheum.
40:1670–1679. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Shen S, Wu Y, Chen J, Xie Z, Huang K, Wang
G, Yang Y, Ni W, Chen Z, Shi P, et al: CircSERPINE2 protects
against osteoarthritis by targeting miR-1271 and ETS-related gene.
Ann Rheum Dis. 78:826–836. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Son YO, Park S, Kwak JS, Won Y, Choi WS,
Rhee J, Chun CH, Ryu JH, Kim DK, Choi HS and Chun JS:
Estrogen-related receptor γ causes osteoarthritis by upregulating
extracellular matrix-degrading enzymes. Nat Commun. 8:21332017.
View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Li B, Jiang T, Liu H, Miao Z, Fang D,
Zheng L and Zhao J: Andrographolide protects chondrocytes from
oxidative stress injury by activation of the Keap1-Nrf2-Are
signaling pathway. J Cell Physiol. 234:561–571. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Tang Q, Zheng G, Feng Z, Chen Y, Lou Y,
Wang C, Zhang X, Zhang Y, Xu H, Shang P and Liu H: Trehalose
ameliorates oxidative stress-mediated mitochondrial dysfunction and
ER stress via selective autophagy stimulation and autophagic flux
restoration in osteoarthritis development. Cell Death Dis.
8:e30812017. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Salinas D, Mumey BM and June RK:
Physiological dynamic compression regulates central energy
metabolism in primary human chondrocytes. Biomech Model
Mechanobiol. 18:69–77. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Charlier E, Deroyer C, Ciregia F, Malaise
O, Neuville S, Plener Z, Malaise M and de Seny D: Chondrocyte
dedifferentiation and osteoarthritis (OA). Biochem Pharmacol.
165:49–65. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Deng Z, Jia Y, Liu H, He M, Yang Y, Xiao W
and Li Y: RhoA/ROCK pathway: Implication in osteoarthritis and
therapeutic targets. Am J Transl Res. 11:5324–5331. 2019.PubMed/NCBI
|
|
31
|
Fernandes JC, Martel-Pelletier J and
Pelletier JP: The role of cytokines in osteoarthritis
pathophysiology. Biorheology. 39:237–246. 2002.PubMed/NCBI
|
|
32
|
Fernandes TL, Gomoll AH, Lattermann C,
Hernandez AJ, Bueno DF and Amano MT: Macrophage: A potential target
on cartilage regeneration. Front Immunol. 11:1112020. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Xie C and Chen Q: Adipokines: New
therapeutic target for osteoarthritis? Curr Rheumatol Rep.
21:712019. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Yoshida K, Furuya S, Osuka S, Mitoma J,
Shinoda Y, Watanabe M, Azuma N, Tanaka H, Hashikawa T, Itohara S
and Hirabayashi Y: Targeted disruption of the mouse
3-phosphoglycerate dehydrogenase gene causes severe
neurodevelopmental defects and results in embryonic lethality. J
Biol Chem. 279:3573–3577. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Song Z, Feng C, Lu Y, Lin Y and Dong C:
PHGDH is an independent prognosis marker and contributes cell
proliferation, migration and invasion in human pancreatic cancer.
Gene. 642:43–50. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Unterlass JE, Baslé A, Blackburn TJ,
Tucker J, Cano C, Noble MEM and Curtin NJ: Validating and enabling
phosphoglycerate dehydrogenase (PHGDH) as a target for
fragment-based drug discovery in PHGDH-amplified breast cancer.
Oncotarget. 9:13139–13153. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Sim WC, Lee W, Sim H, Lee KY, Jung SH,
Choi YJ, Kim HY, Kang KW, Lee JY, Choi YJ, et al: Downregulation of
PHGDH expression and hepatic serine level contribute to the
development of fatty liver disease. Metabolism. 102:1540002020.
View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Bonelli R, Woods SM, Ansell BRE, Heeren
TFC, Egan CA, Khan KN, Guymer R, Trombley J, Friedlander M, Bahlo M
and Fruttiger M: Systemic lipid dysregulation is a risk factor for
macular neurodegenerative disease. Sci Rep. 10:121652020.
View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Kozhemyakina E, Lassar AB and Zelzer E: A
pathway to bone: Signaling molecules and transcription factors
involved in chondrocyte development and maturation. Development.
142:817–831. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Lefebvre V and Dvir-Ginzberg M: SOX9 and
the many facets of its regulation in the chondrocyte lineage.
Connect Tissue Res. 58:2–14. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Lefebvre V, Angelozzi M and Haseeb A: SOX9
in cartilage development and disease. Curr Opin Cell Biol.
61:39–47. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Si HB, Zeng Y, Liu SY, Zhou ZK, Chen YN,
Cheng JQ, Lu YR and Shen B: Intra-articular injection of
microRNA-140 (miRNA-140) alleviates osteoarthritis (OA) progression
by modulating extracellular matrix (ECM) homeostasis in rats.
Osteoarthritis Cartilage. 25:1698–1707. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Sun K, Luo J, Jing X, Guo J, Yao X, Hao X,
Ye Y, Liang S, Lin J, Wang G and Guo F: Astaxanthin protects
against osteoarthritis via Nrf2: A guardian of cartilage
homeostasis. Aging. 11:10513–10531. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Ansari MY, Ahmad N and Haqqi TM: Oxidative
stress and inflammation in osteoarthritis pathogenesis: Role of
polyphenols. Biomed Pharmacother. 129:1104522020. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Dandekar A, Mendez R and Zhang K: Cross
talk between ER stress, oxidative stress, and inflammation in
health and disease. Methods Mol Biol. 1292:205–214. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Brandl A, Hartmann A, Bechmann V, Graf B,
Nerlich M and Angele P: Oxidative stress induces senescence in
chondrocytes. J Orthop Res. 29:1114–1120. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Sangokoya C, Telen MJ and Chi JT: MicroRNA
miR-144 modulates oxidative stress tolerance and associates with
anemia severity in sickle cell disease. Blood. 116:4338–4348. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Jiang HK, Miao Y, Wang YH, Zhao M, Feng
ZH, Yu XJ, Liu JK and Zang WJ: Aerobic interval training protects
against myocardial infarction-induced oxidative injury by enhancing
antioxidase system and mitochondrial biosynthesis. Clin Exp
Pharmacol Physiol. 41:192–201. 2014. View Article : Google Scholar : PubMed/NCBI
|