Introduction
Glucocorticoids (GCs) are anti-inflammatory and
immunosuppressant agents that have been studied as potential
treatments for hematological cancer (1). GCs are also used as drug adjuvants for
the prevention and amelioration of non-hematological cancer
(2,3). Mechanistically, GCs exert antitumor
effects by inducing apoptosis, inhibiting cell proliferation and
modulating the inflammatory pathway (4,5).
Prednisolone is a commonly used GC for treating liver disease and
cancer (6,7). Previous studies have demonstrated that
prednisolone increases apoptosis in CCRF-CEM cells (8,9).
Prednisolone has also been reported to enhance the metabolism of
sorafenib, which can improve the outcome of therapy in the
treatment of hepatocellular carcinoma (10). However, GC has anti-cancer effects
in haematologic malignancies, but it has limited effects in solid
tumors (11,12). Therefore, the development of novel
drug derivatives may have notable implications for improving the
efficacy for GC-based cancer treatment.
Ionizing radiation is used to modify the structure
of numerous drugs, and has been demonstrated to enhance their
biological properties in vitro (13,14).
Recent reports have demonstrated that γ-irradiation can create
novel GC derivatives, such as γ-irradiated (IR)-dexamethasone
(Dex-IR) and Kenalog-IR, the anticancer effects of which have been
confirmed in vitro (15,16).
To the best of our knowledge, however, the effects of γ-irradiation
on prednisolone structure, and the mechanism by which it induces
anticancer effects, have not yet been explained.
In order to determine the biological activities of
new prednisolone derivatives created using γ-irradiation, the
present study evaluated the anticancer activities of
IR-prednisolone in liver cancer cells. The present study
demonstrated that IR-prednisolone decreased the growth of Huh7 and
HepG2 cells. Thus, it was hypothesized that IR-prednisolone may
have anticancer properties in liver cancer cells.
Materials and methods
Cell lines, chemicals and
reagents
Huh7 and HepG2 cells were acquired from the American
Type Culture Collection. The cells were maintained in DMEM
containing 10% fetal bovine serum and 1% streptomycin/penicillin
(all from Thermo Fisher Scientific, Inc.) at 37°C in a humidified
5% CO2 incubator. Prednisolone, MTT and doxorubicin were
obtained from Sigma-Aldrich (Merck KGaA). The Muse®
Annexin V Dead cell kit and cell cycle arrest package for flow
cytometry analysis, and the PVDF membranes were purchased from EMD
Millipore. For nick-end labeling, the DeadEnd™ Fluorometric TUNEL
System was purchased from Promega Corporation. The human apoptosis
proteome array kit was obtained from Bio-Rad Laboratories, Inc. The
ECL detection reagent was bought from GE Healthcare. RIPA buffer
was procured from Rockland Immunochemicals, Inc. Bicinchoninic acid
(BCA) reagent was obtained from Thermo Fisher Scientific, Inc. The
cytochrome c release kit (cat. no. ab65311) was from Abcam.
Anti-GAPDH, anti-cleaved caspase 3, anti-cleaved caspase 9,
anti-caspase 9, anti-cleaved poly-ADP ribose polymerase (PARP),
anti-PARP, anti-caspase 3, anti-Bcl-2, anti-Bax, and horseradish
peroxidase (HRP)-conjugated anti-rabbit IgG and anti-mouse (cat.
nos. 2118, 9664, 9505, 9502, 9541, 9532, 9662, 2870, 5023, 7074 and
7076, respectively) were obtained from Cell Signaling Technology,
Inc.
Prednisolone modification using
γ-irradiation
Prednisolone was modified using γ-irradiation under
the conditions described by Lee et al (15). Liquid chromatography-mass
spectrometry (LC-MS) analysis was used to determine the structural
changes in IR-prednisolone. Briefly, a sample solution of
prednisolone (1 g) in MeOH (1 liter) in chapped vials was exposed
to 50 kGy (absorbed dose) of radiation. Irradiation was carried out
at ambient temperature using a Cobalt-60 irradiator [Nordion
(Canada), Inc.] at the Advanced Radiation Technology Institute,
Korea Atomic Energy Research Institute (Jeongup, Korea). The source
strength was ~320 kCi, with a dose rate at the location of the 10
kGy/h sample. The prednisolone and IR-prednisolone samples were
assessed using the HPLC 1260 system (Agilent Technologies, Inc.) on
a YMC-Pack ODS-A 302 column (4.6×150 mm; YMC Co., Ltd.). It was
developed at 40°C with 1% HCOOH/MeCN (1:1, flow rate, 1.0 per min;
detection, 360 nm). LC-MS was performed using an Agilent 1100
series LC-MSD system (Agilent Technologies, Inc.) in both positive
and negative ion modes. Separation was performed with a YMC-Pack
Pro C18 reverse-phase column (250×4.6 mm2 I.D. S-5 µm,
12 nm; YMC Co., Ltd.) at a flow rate of 1 ml/min. The mobile phase
consisted of 0.1% (v/v) trifluoracetic acid (TFA; solution A) and
50% (v/v) acetonitrile in 0.1% (v/v) TFA (solution B). Elution was
performed by a step gradient from a ratio of 80:20 (solutions A:B)
to 67:33 in 18 min; from 67:33 to 60:40 in 12 min; and then from
60:40 to 80:20 in 10 min. The eluted compounds were detected at a
wavelength of 520 nm. Electrospray ionization was performed with an
ES-APCI mass spectrometer (Agilent Technologies, Inc.). Hydrogen
was used as the nebulizing and drying gas. The ESI conditions used
were as follows: Nebulizer pressure, 35 psig; drying gas, 12 l/min
at 350 1C; ion spray voltage, 4,000 V; and charging voltage, 2,000
V.
MTT cell viability assay
Huh7 and HepG2 cells were grown for 24 h and treated
with prednisolone or IR-prednisolone (5, 10, 50 and 100 µg/ml), or
DMSO as negative control for 48 h at 37°C. Following incubation, 20
µl MTT mixture was added for 2 h at 37°C. The medium was replaced
by 100 µl DMSO to break down formazan precipitates. The absorbances
were measured at 562 nm using an Infinite M200 multimode microplate
reader (Tecan Group, Ltd.).
Propidium iodide (PI) staining for
cell cycle detection
Huh7 cells were grown for 24 h before being
stimulated with 100 µg/ml prednisolone or IR-prednisolone, or 0.5
µM doxorubicin as a positive control, for a further 24 h. Cells
were fixed in 70% ethanol at −20°C for 3 h, then stained with 150
µl PI at 25°C for 30 min in the absence of light. The Muse Cell
Analyzer (EMD Millipore) estimated the number of stained
nuclei.
Annexin V-FITC/PI staining for
apoptosis assay
Annexin V-FITC/PI staining determined the number of
cells in each population. Huh7 cells were cultured for 24 h. The
cells were treated as described in the cell cycle assay. Then cells
were harvested, and stained with 100 µl Annexin V-FITC/PI solution
at room temperature for 20 min in the dark, as per the
manufacturer's directions. The number of apoptotic cells was
determined using the Muse Cell Analyzer (EMD Millipore). The Muse
software (version 1.1.2; EMD Millipore) evaluated the numbers of
apoptotic and non-apoptotic cells in each of the four populations:
Live cell; Annexin V (−) and PI (−); early apoptotic cell; Annexin
V (+) and PI (−), late apoptotic or dead cell; Annexin V (+) and PI
(+); and dead non-apoptotic cell; Annexin V (−) and PI (+).
TUNEL assay
Huh7 cells were grown on microscope slides for 24 h.
The treatment conditions used for apoptosis analysis were applied
for 12 h, then cells were analysed using the TUNEL assay kit
(Promega Corporation) and counterstained using DAPI according to
the supplier's instructions. In brief, the cells were washed with
cold PBS, fixed with 4% paraformaldehyde for 20 min at room
temperature. Subsequently, the cells were permeabilized by
immersing the slide in 0.2% Triton X-100 solution in PBS for 5 min.
The cells were washed again in cold PBS twice for 2 min. The
equilibration buffer was added for 10 min at room temperature,
followed by 100 µl terminal deoxynucleotidyl transferase (TdT)
reaction mix, covered and incubated at 25°C for 60 min. The slides
were then immersed in 2X saline sodium citrate for 15 min to
terminate the reaction and washed three times for 5 min each in PBS
to remove unbound biotinylated nucleotides. The nuclei were stained
with DAPI and incubated at room temperature for 5 min. After
mounting, the apoptotic cells were captured on an Olympus IX71
fluorescence microscope (magnification, ×40; Olympus Corporation).
To quantify the immune-probed cells, the fluorescence intensity was
taken in 10 randomly selected images.
Human apoptosis array assessment
Huh7 cells were grown on a 100-mm plate for 24 h,
following treatment with 100 µg/ml IR-prednisolone or DMSO for 24
h. Briefly, harvested cells were lysed with 1X RIPA buffer
(Rockland Immunochemicals, Inc.), and concentrations were
calculated using the BCA protein assay. Then, 250 µg total proteins
were incubated in the nitrocellulose membrane (NC) overnight at
4°C. The assay was performed using the apoptosis proteome array kit
following the manufacturer's instructions; subsequently, NC
membranes were developed on X-ray film for 5 min. Each protein was
estimated using Multi Gauge software (version 3.0; FujiFilm).
Western blotting analysis
Huh7 cells (9×105 cells/plate) were grown
in a 100-mm plate for 24 h. The treatment conditions (for 12 and 24
h) were the same as for cell cycle analysis. Protein extraction was
performed using RIPA buffer and, the cytochrome c release kit was
used to isolate cytosol and mitochondria fractions. Protein
concentrations were determined using the BCA protein assay. Equal
volumes of 40 µg proteins were separated via SDS-PAGE on 10% gels,
and then transferred onto PVDF membranes and blocked in non-fat
milk in tris-buffered saline with 0.1% Tween-20 (TBST) buffer at
room temperature. The membranes were incubated overnight with
primary antibodies (1:1,000) and cytochrome c antibody (1:500) at
4°C, then washed with three changes of TBST buffer. The membranes
were incubated with anti-rabbit conjugated-HRP (1:3,000) and
anti-mouse conjugated-HRP (1:3,000) at room temperature for 1 h,
followed by three washes in TBST. The proteins were then visualised
using an enhanced chemiluminescence reagent (ECL; EMD Millipore)
and exposure to an X-ray film. The blots were scanned and
densitometric analysis was performed using ImageJ software (version
1.51k; National Institutes of Health).
JC-1 staining for determination of
mitochondrial membrane potential (ΔΨm)
The double-emission fluorescence dye JC-1 was used
to analyse the status of ΔΨm. Huh7 cells (5×104) were
grown for 24 h then treated with the aforementioned conditions for
48 h or with 50 µM carbonyl cyanide 3-chlorophenyl hydrazine (CCCP)
for 30 min. Cells were stained with 1 ml DMEM containing JC-1 dye
at 37°C for 20 min, then washed with two changes of PBS and
reconstituted in PBS. For positive control cells, CCCP at 50 µM for
5 min was used to alter the ΔΨm. JC-1-positive cells were detected
using a Cytomics FC500 flow cytometer (Beckman Coulter, Inc.),
whereas the red to the green fluorescent cell population was
determined using CPX software version 2.2 (Beckman Coulter,
Inc.).
Statistical analysis
The data are presented as the mean ± standard
deviation of ≥3 independent experiments. Statistical analysis was
performed using Student's t-test and PASW Statistics software
version 18 (SPSS, Inc.). Multiple comparisons were performed using
one-way ANOVA then Tukey's post hoc analysis. P<0.05 was
considered to indicate a statistically significant difference.
Results
Effects of γ-irradiation on
prednisolone structure
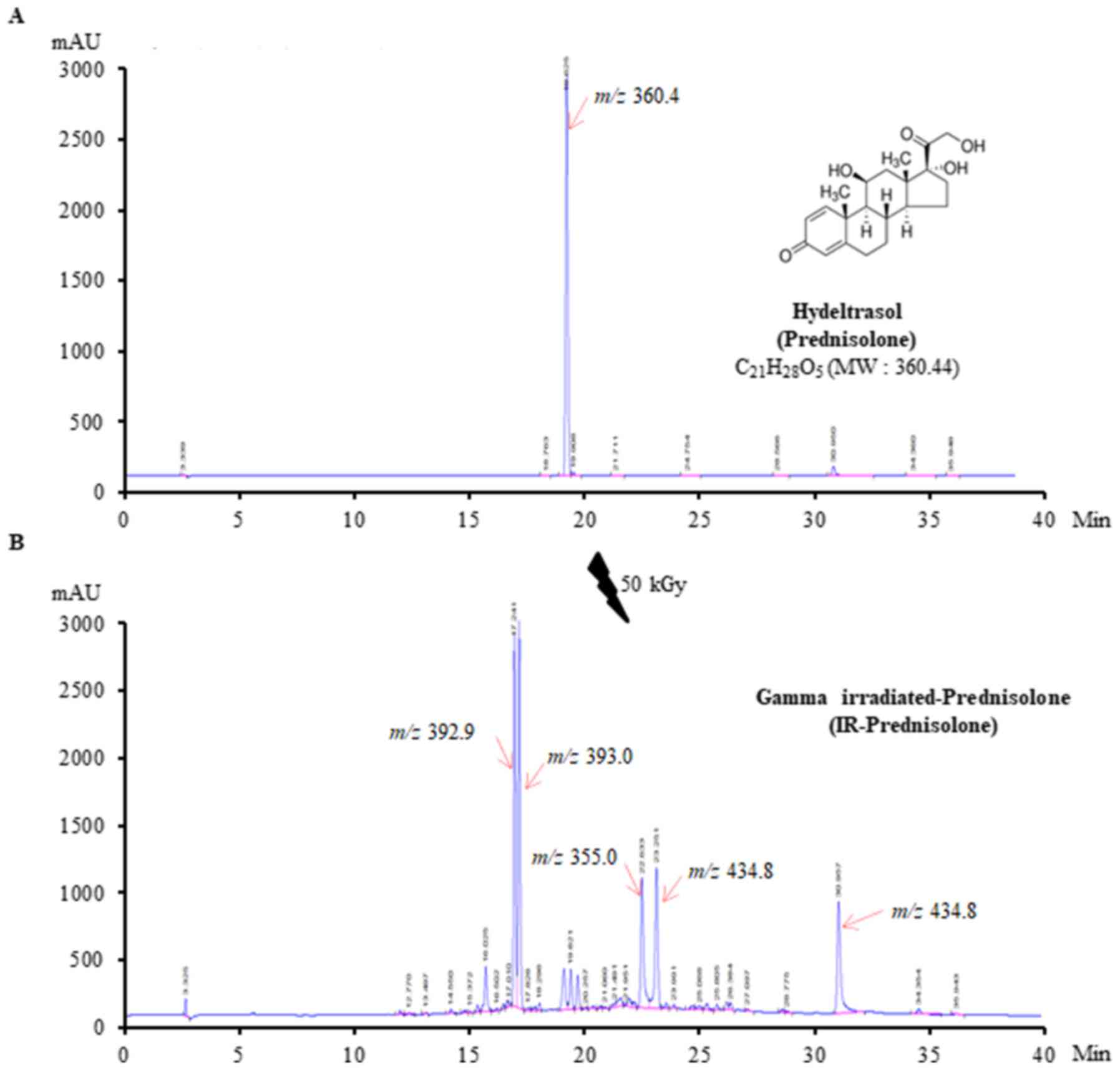
The results of the present study demonstrated that
the prednisolone chromatogram had a single peak at the retention
time of 19.625 min with a mass of 361.4 m/z (Fig. 1A). The IR-prednisolone chromatogram
showed five new daughter peaks detected at retention times of
17.241, 17.397, 22.251, 23.633 and 30.957 min with masses of 392.9,
393.0, 355.0, 434.8 and 816.6 m/z respectively, whereas the mother
peak of prednisolone had completely disappeared (Fig. 1B) indicating that 50 kGy
γ-irradiation was able to generate new prednisolone
derivatives.
IR-prednisolone prevents the growth of
liver cancer cells
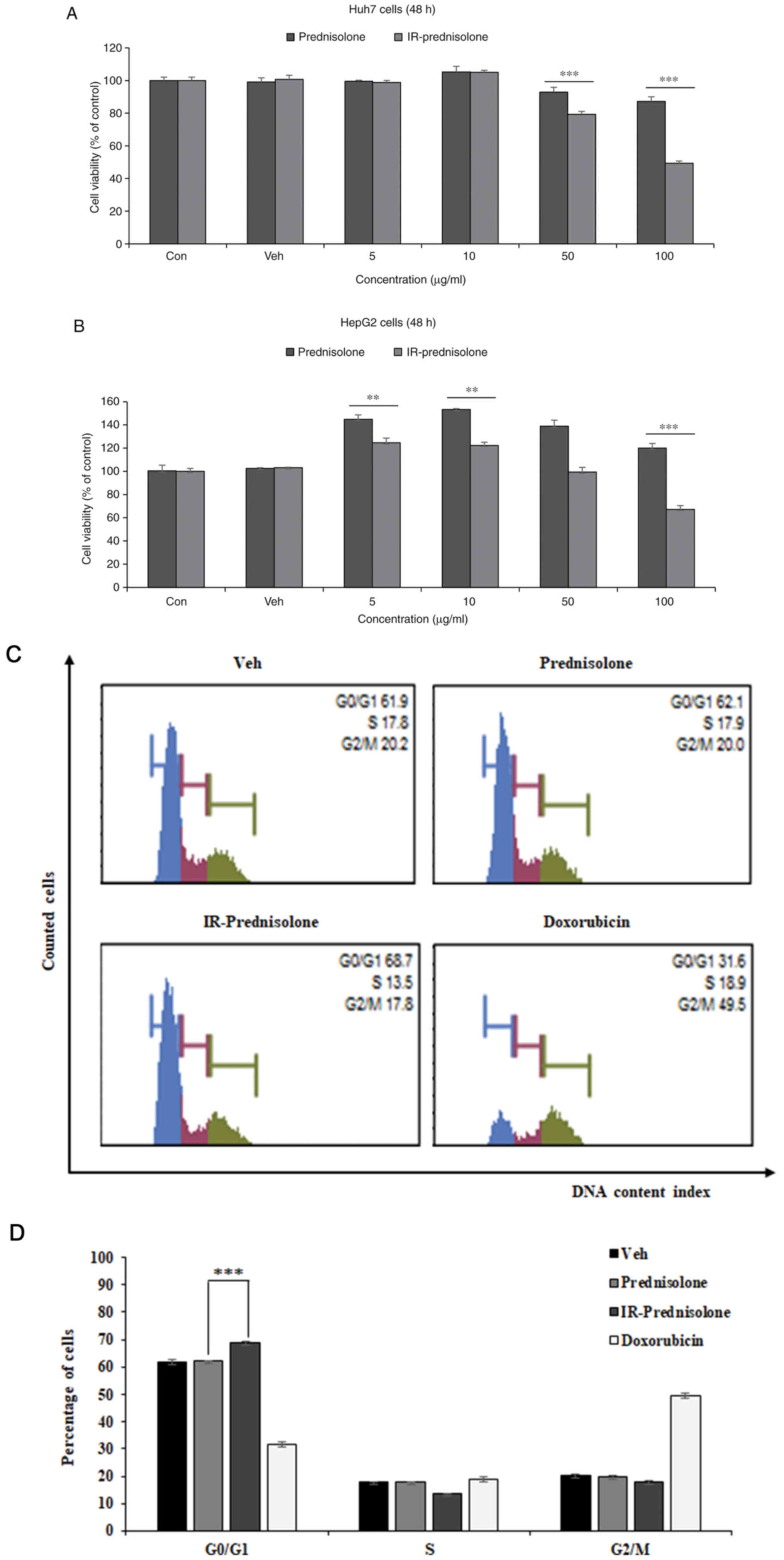
MTT assays were conducted to determine the
anticancer effects of IR-prednisolone on liver cancer cell lines.
IR-prednisolone significantly decreased the viability of Huh7 cells
from 83.9% with prednisolone to 47.4% with IR-prednisolone at 100
µg/ml (Fig. 2A). A significant
decrease in cell viability was observed in HepG2 cells from 120%
with prednisolone to 67% with IR-prednisolone at 100 µg/ml
(Fig. 2B). In contrast with the
control group, IR-prednisolone decreased the cell viability of Huh7
cells by 52.6%, whereas the viability of HepG2 cells decreased by
33%. This indicated that IR-prednisolone was more sensitive to Huh7
cells than to HepG2 cells. Therefore, this cell line was used to
investigate the anticancer effects of IR-prednisolone. In order to
determine whether IR-prednisolone decreases cell viability by
preventing cell cycle progression in Huh7 cells, the present study
investigated the DNA content of cells in different phases of the
cell cycle. The results of cell DNA content profiles demonstrated a
significant increase in the number of cells at G0/G1 from 62.1%
with prednisolone to 68.7% with IR-prednisolone. The slightly
increased number of cells in the G0/G1-phase indicates that
IR-prednisolone may induce cell cycle arrest in Huh7 cells
(Fig. 2C and D). These results
indicated that IR-prednisolone may prevent the growth of Huh7
cells.
IR-prednisolone increases the
apoptosis of Huh7 cells
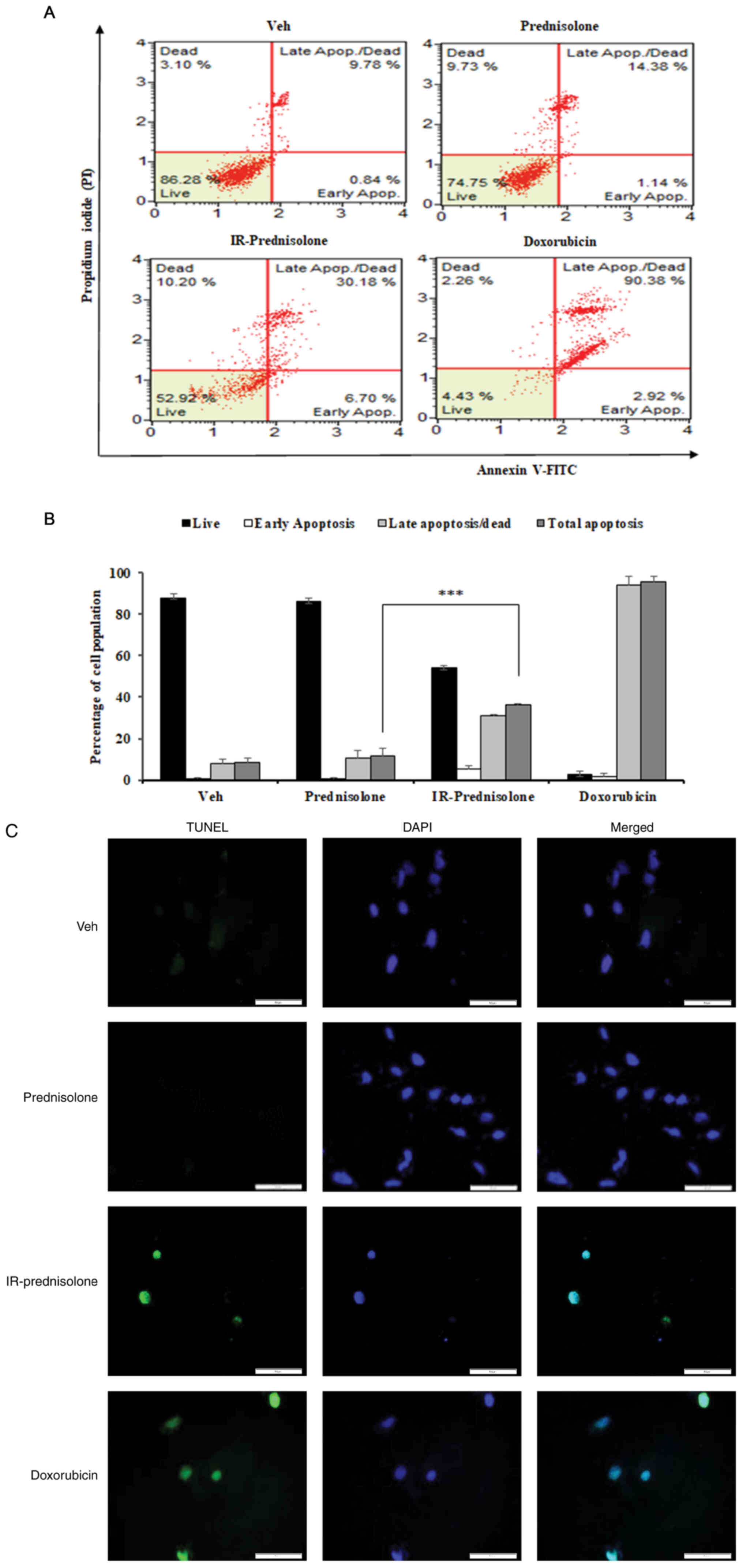
In order to determine the role of IR-prednisolone in
inhibition of cell viability, programmed cell death was evaluated
in Huh7 cells. The results of the present study demonstrated that
IR-prednisolone significantly increased the population of total
apoptotic cells to 36.3% compared with 11.3% in
prednisolone-treated cells (Fig. 3A and
B). In order to further confirm whether IR-prednisolone induced
apoptosis, a TUNEL assay was performed. The results demonstrated
that IR-prednisolone-treated cells displayed colocalization of
TUNEL staining, which stains small fragments of DNA (green), and
DAPI (blue), a nucleus and DNA staining dye, compared with
prednisolone-treated cells, where no colocalization was observed
(Fig. 3C). These results showed
that prednisolone transformed through ionizing radiation
effectively inhibit the viability of Huh7 cells by inducing certain
physiological changes.
IR-prednisolone induces intrinsic
apoptosis in Huh7 cells
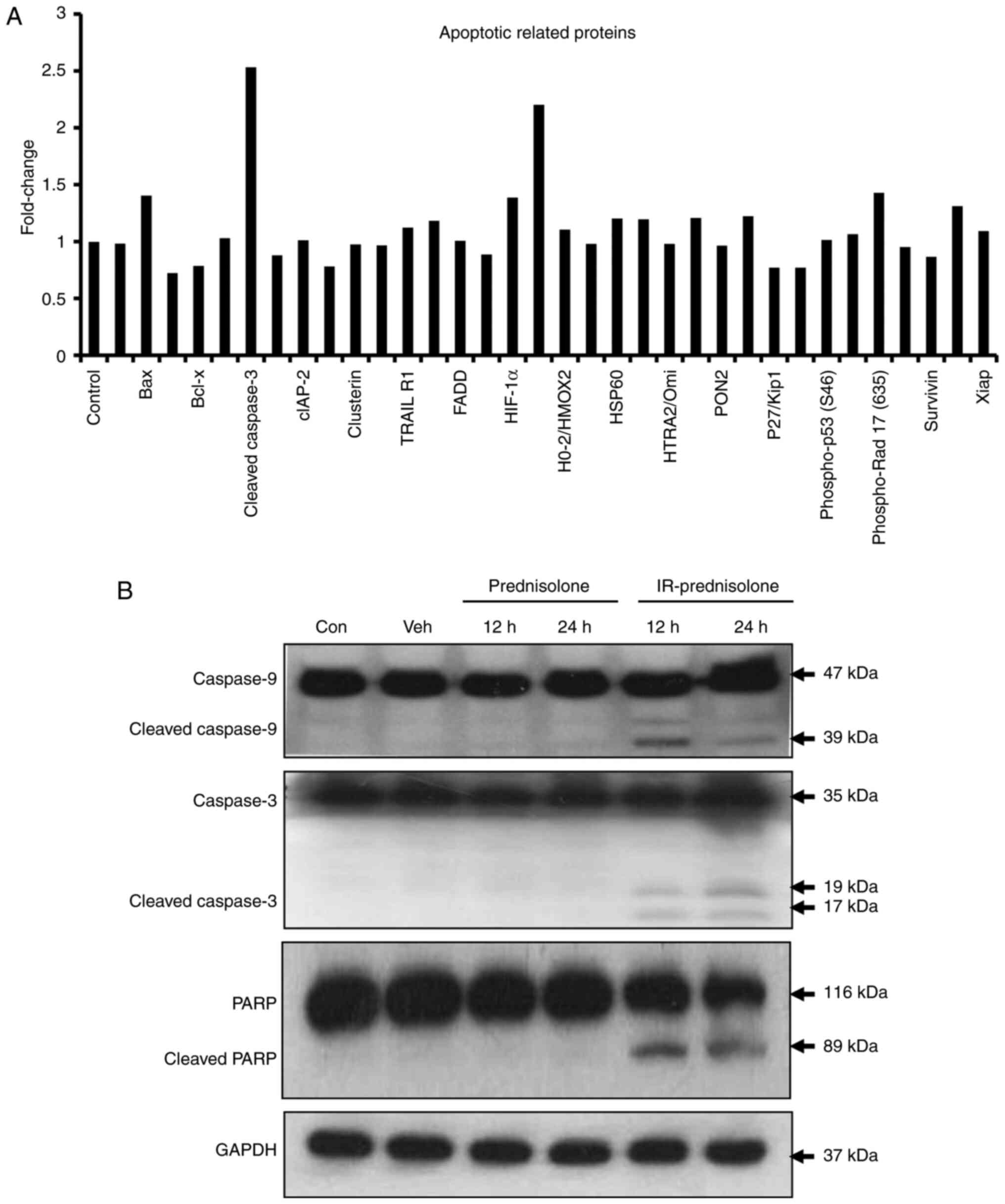
In order to identify the mechanism of
IR-prednisolone-induced apoptosis, apoptosis-associated proteins
were screened using western blotting. The apoptosis array results
demonstrated that IR-prednisolone increased levels of cleaved
caspase-3, Bax, tumor necrosis factor receptor 1, heme oxygenase-1,
p21, phosphorylated-Rad17 and hypoxia inducible factor-1α, but
decreased the expression levels of cellular inhibitor of
apoptosis-1, Claspin, Survinin, Bcl-2, Bcl-x and p27 (Fig. 4A). Furthermore, investigation of
different proteins connected to apoptosis pathways revealed that
IR-prednisolone treatment activated cleaved caspases 9 and 3, and
increased cleaved PARP levels compared with those in
prednisolone-treated cells in a time-dependent manner (Fig. 4B). These data demonstrated that
IR-prednisolone activates the intrinsic apoptosis pathway.
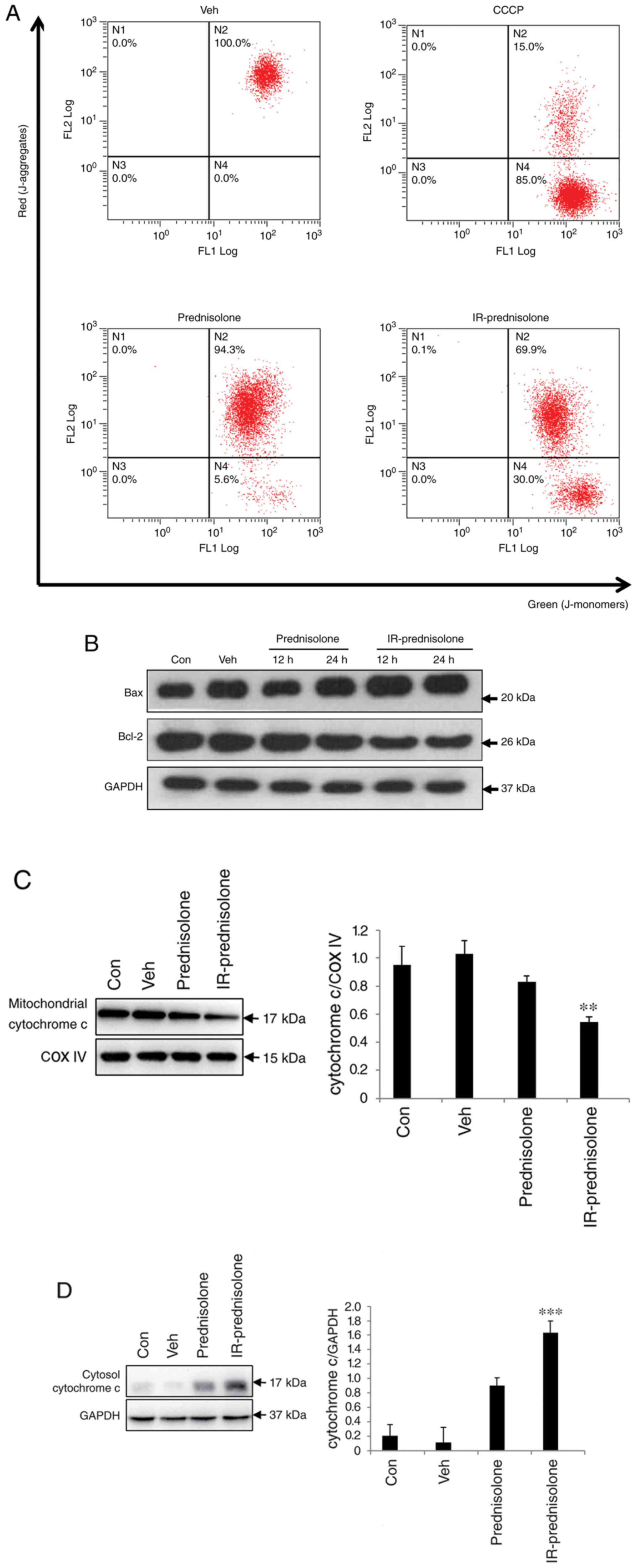
Effect of IR-prednisolone on ΔΨm
Alteration of ΔΨm is a critical process in the
intrinsic apoptosis pathway (17).
Therefore, involvement of intrinsic apoptosis was investigated by
JC-1 staining. The results of depolarization status of ΔΨm
demonstrated that IR-prednisolone-treated cells increased JC-1
monomers (green fluorescence) up to 30.0% compared with 5.6%
observed in prednisolone-treated cells (Fig. 5A). The Bcl-2 family of proteins is
known to be a regulator of mitochondrial membrane dysfunction
(18). Investigation of
mitochondrial membrane dysfunction regulator proteins demonstrated
that IR-prednisolone upregulated Bax and decreased levels of Bcl-2
compared with those in prednisolone-treated cells (Fig. 5B). The loss of ΔΨm has been
demonstrated to increase the total amount of cytochrome c movement
towards the cytosol and mediate apoptosis signaling (19). The present study demonstrated that
IR-prednisolone significantly increased cytosolic cytochrome c
levels and decreased the level of mitochondrial cytochrome c in
Huh7 cells (Fig. 5C and D),
indicating that IR-prednisolone induced the intrinsic apoptosis
signaling cascade.
Discussion
Prednisolone has been reported to exert anticancer
actions in a number of cancer cells, including leukemia, and
prostate and oesophageal cancers (20–22).
Previous studies have demonstrated that prednisolone increases the
level of cytochrome P450 3A4, induces metabolism of sorafenib and
enhances the efficacy of chemotherapy during treatment of patients
with liver cancer (10). Although
prednisolone shows therapeutic roles in numerous types of cancer,
the extent of its efficacy remains debatable. In the present study,
the structure of prednisolone was improved using γ-irradiation.
Higher levels of cytotoxic effects were observed following
treatment with IR-prednisolone compared with those in
prednisolone-treated liver cancer cells. Therefore, it was
hypothesized that IR-prednisolone may have anticancer activity in
liver cancer cells.
The present study demonstrated that the use of
ionizing radiation was a potential promising technique for
structural modification. A number of studies have demonstrated that
γ-irradiation modifies the structures of existing compounds in
order to develop novel compounds (14,23,24).
Lee et al (15) reported
that GCs exposed to IR exhibit improved physiological properties.
These studies have demonstrated that ionizing radiation can be used
to improve the anticancer activity of drugs. The results presented
in Fig. 1 are consistent with
previous reports that IR-prednisolone drugs produce several peaks
compared to the single peak of prednisolone (15,25).
These observations indicate that γ-irradiation may be used to
develop new prednisolone derivatives. The chemical structure for
IR-prednisolone was not determined in the present study; however,
based on previous observations, it was hypothesized that
γ-irradiation changes the arrangement of functional groups to
produce new prednisolone derivatives (14,25).
The potential biological activity of IR-prednisolone
was investigated using cancer cells. Among the tested cells
(SK-MEL-5 as the skin cancer cell line, MCF7 as the breast cancer
cell line and A549 as the lung cancer cell line), IR-prednisolone
reduced cell viability only in the liver cancer cell lines, Huh7
and HepG2 (data not shown). Previous reports have demonstrated that
γ-irradiated drugs promote anticancer activity primarily by
decreasing cell proliferation and apoptosis induction (15,25).
In the present study, IR-prednisolone significantly decreased the
viability of liver cancer cells. As the percentage of apoptotic
cells has been demonstrated to increase in numerous types of cancer
cell following treatment with γ-irradiated drugs, it was
hypothesized that IR-prednisolone may decrease the viability of
liver cancer cells by promoting apoptosis and cell arrest (16). Evading apoptosis is a hallmark of
cancer; potential anticancer drugs should therefore promote
apoptosis (17). In the present
study, apoptosis was accelerated in IR-prednisolone-treated Huh7
cells. DNA fragmentation is considered to be a hallmark of
apoptosis, and is controlled by both endonucleases and caspases to
form small fragments of DNA (17).
DNA fragmentation demonstrated that IR-prednisolone induced
apoptosis by a mechanism similar to that reported in other studies
(26–28). IR-prednisolone-treated cells were
shown to be arrested at G0/G1, and similar
results have been observed following treatment with Dex-IR
(15).
The activation of caspases (aspartate-specific
cysteine proteases) controls apoptosis (29). Also, Bcl-2/Bax play an important
role in regulating caspase-dependent and caspase-independent
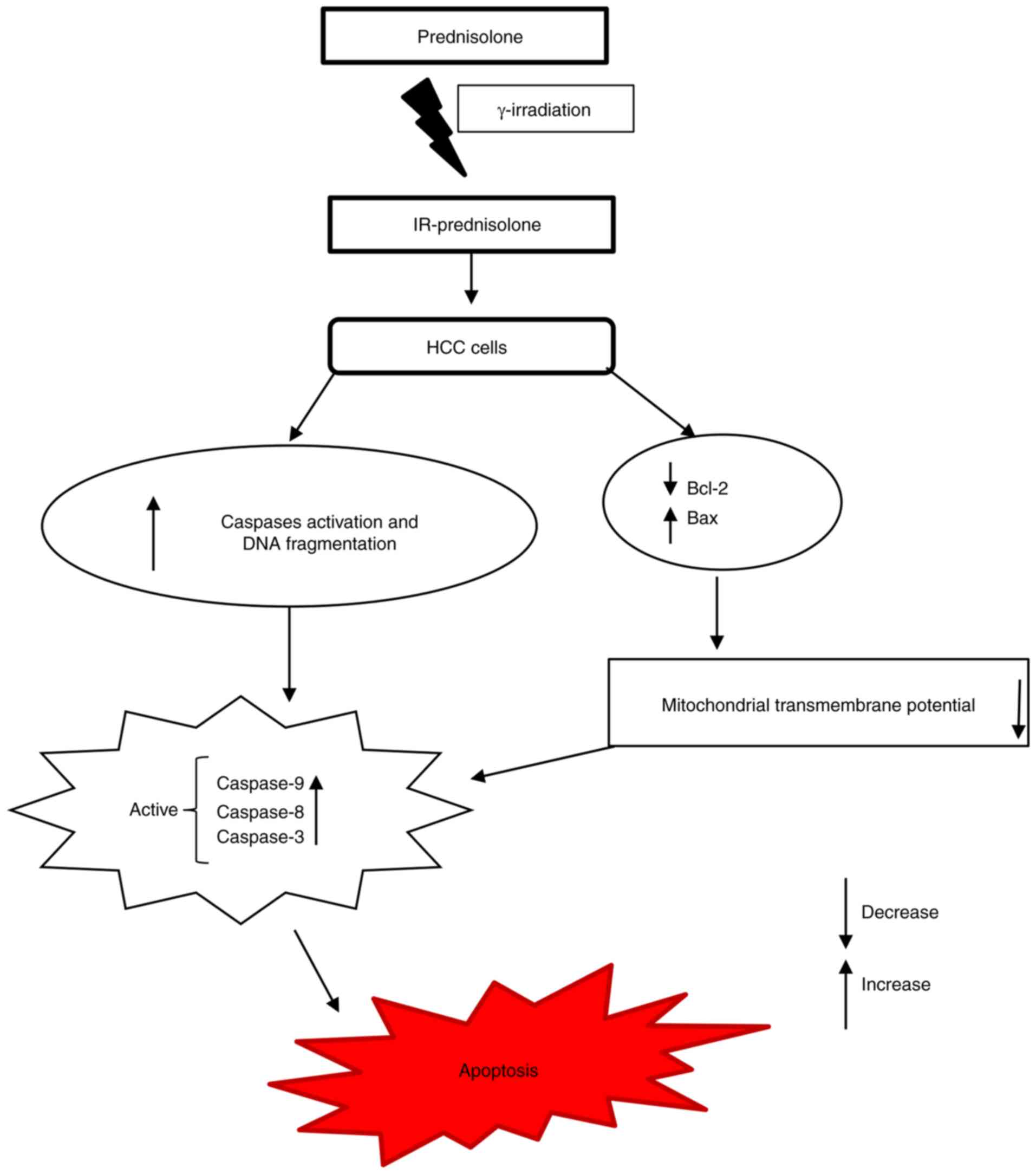
apoptosis mediated by the mitochondrial pathway (19). The intrinsic apoptosis pathway is
activated by Bax attached to mitochondria, following cytochrome c
movement towards the cytosol, and activation of cleavage of caspase
9, caspase 3 and PARP (30). These
events were observed in IR-prednisolone-treated cells, indicating
that the intrinsic pathway was involved in IR-prednisolone-induced
apoptosis (Fig. 6). Collectively,
these findings indicate that IR-prednisolone increased apoptosis by
activating the intrinsic apoptosis pathway.
Although the present study demonstrated the
anticancer effects of IR-prednisolone, further analysis is required
to identify the chemical structure and compounds within the
IR-prednisolone mixture. In addition, separating individual
fractions to analyse key single compounds and confirm their
effectiveness will have to be carried out. Afterward, further
research is required to investigate the molecular mechanism
underlying these effects.
In conclusion, the present study demonstrated that
γ-irradiation created a new prednisolone derivative. The results of
the present study demonstrated that IR-prednisolone prevented the
growth of liver cancer cells and arrested cells at the G0/G1 stage
of the cell cycle. IR-prednisolone promoted apoptosis of Huh7 cells
via alteration of ΔΨm, DNA fragmentation and decreased Bcl-2
expression levels. Mechanistically, IR-prednisolone activates the
intrinsic apoptotic signaling pathway via caspase-dependent
activation. However, further research is necessary to identify the
specific molecules and mechanisms involved in this effect. The
results of the present study indicate that IR-prednisolone obtained
through radiolytic transformation technology is a potential
candidate for the development of novel therapies for liver
cancer.
Acknowledgements
Not applicable.
Funding
This research was supported by the Nuclear R & D
Program from the Ministry of Science and ICT, Republic of
Korea.
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
FJR and HWB conceptualized and designed the study,
and wrote the manuscript. FJR, SHK and RAK conducted the
experiments, and interpreted and analyzed data. BYC and BSK made
substantial contributions to conception and design, and interpreted
the data. FJR and HWB confirm the authenticity of all the raw data.
All authors read and approved the final manuscript.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Coleman RE: Glucocorticoids in cancer
therapy. Biotherapy. 4:37–44. 1992. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Twycross R: Corticosteroids in advanced
cancer. BMJ. 305:969–970. 1992. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Kofler R: The molecular basis of
glucocorticoid-induced apoptosis of lymphoblastic leukemia cells.
Histochem Cell Biol. 114:1–7. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Schlossmacher G, Stevens A and White A:
Glucocorticoid receptor-mediated apoptosis: Mechanisms of
resistance in cancer cells. J Endocrinol. 211:17–25. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Smith LK and Cidlowski JA:
Glucocorticoid-induced apoptosis of healthy and malignant
lymphocytes. Prog Brain Res. 182:1–30. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Powell LW and Axelsen E: Corticosteroids
in liver disease: Studies on the biological conversion of
prednisone to prednisolone and plasma protein binding. Gut.
13:690–696. 1972. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Manabe A, Ohara A, Hasegawa D, Koh K,
Saito T, Kiyokawa N, Kikuchi A, Takahashi H, Ikuta K, Hayashi Y, et
al: Significance of the complete clearance of peripheral blasts
after 7 days of prednisolone treatment in children with acute
lymphoblastic leukemia: The Tokyo Children's cancer study group
study L99-15. Haematologica. 93:1155–1160. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Khanzadeh T, Hagh MF, Talebi M, Yousefi B,
Azimi A, Pour Feizi AAH and Baradaran B: Investigation of BAX and
BCL2 expression and apoptosis in a resveratrol-and
prednisolone-treated human T-ALL cell line, CCRF-CEM. Blood Res.
53:53–60. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Ganbarjeddi S, Azimi A, Zadi Heydarabad M,
Hemmatzadeh M, Mohammadi S, Ardehaie RM, Zamani M, Baharaghdam S,
Esmaeili S and Ghasemi A: Apoptosis induced by prednisolone occurs
without altering the promoter methylation of BAX and BCL-2 genes in
acute lymphoblastic leukemia cells CCRF-CEM. Asian Pac J Cancer
Prev. 21:523–529. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Noda S, Shioya M, Hira D, Fujiyama Y,
Morita SY and Terada T: Pharmacokinetic interaction between
sorafenib and prednisolone in a patient with hepatocellular
carcinoma. Cancer Chemother Pharmacol. 72:269–272. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Sundahl N, Clarisse D, Bracke M, Offner F,
Berghe WV and Beck IM: Selective glucocorticoid receptor-activating
adjuvant therapy in cancer treatments. Oncoscience. 3:188–202.
2016. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Volden PA and Conzen SD: The influence of
glucocorticoid signaling on tumor progression. Brain Behav Immun.
30 (Suppl):S26–S31. 2013. View Article : Google Scholar
|
|
13
|
Kim WS, Song HY, Mushtaq S, Kim JM, Byun
EH, Yuk JM and Byun EB: Therapeutic potential of gamma-irradiated
resveratrol in ulcerative colitis via the anti-inflammatory
activity and differentiation of tolerogenic dendritic cells. Cell
Physiol Biochem. 52:1117–1138. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Badaboina S, Bai HW, Na YH, Park CH, Kim
TH, Lee TH and Chung BY: Novel radiolytic rotenone derivative,
rotenoisin B with potent anti-carcinogenic activity in hepatic
cancer cells. Int J Mol Sci. 16:16806–16815. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Lee EH, Park CH, Choi HJ, Kawala RA, Bai
HW and Chung BY: Dexamethasone modified by gamma-irradiation as a
novel anticancer drug in human non-small cell lung cancer. PLoS
One. 13:e01943412018. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Kawala RA, Ramadhani FJ, Choi HJ, Lee EH,
Park CH, Chung BY and Bai HW: Kenalog modified by ionizing
radiation induces intrinsic apoptosis mediated by elevated levels
of reactive oxygen species in melanoma cancer. Oncol Rep.
41:1837–1850. 2019.PubMed/NCBI
|
|
17
|
Adams JM and Cory S: The Bcl-2 apoptotic
switch in cancer development and therapy. Oncogene. 26:1324–1337.
2007. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Martinou JC and Youle RJ: Mitochondria in
apoptosis: Bcl-2 family members and mitochondrial dynamics. Dev
Cell. 21:92–101. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Wolter KG, Hsu YT, Smith CL, Nechushtan A,
Xi XG and Youle RJ: Movement of Bax from the cytosol to
mitochondria during apoptosis. J Cell Biol. 139:1281–1292. 1997.
View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Kitazumi I and Tsukahara M: Regulation of
DNA fragmentation: The role of caspases and phosphorylation. FEBS
J. 278:427–441. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Inaba H and Pui CH: Glucocorticoid use in
acute lymphoblastic leukaemia. Lancet Oncol. 11:1096–1106. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Venkitaraman R, Lorente D, Murthy V,
Thomas K, Parker L, Ahiabor R, Dearnaley D, Huddart R, De Bono J
and Parker C: A randomised phase 2 trial of dexamethasone versus
prednisolone in castration-resistant prostate cancer. Eur Urol.
67:673–679. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Kataoka M, Anzai S, Shirasaki T, Ikemiyagi
H, Fujii T, Mabuchi K, Suzuki S, Yoshida M, Kawai T and Kitajima M:
Efficacy of short period, low dose oral prednisolone for the
prevention of stricture after circumferential endoscopic submucosal
dissection (ESD) for esophageal cancer. Endosc Int Open.
3:E113–E117. 2015.
|
|
24
|
Alshammari FM, Reda SM and Ghannam MM:
Potential effects of gamma irradiation on the stability and
therapeutic activity of anticancer drug, doxorubicin. J Radiat Res
Appl Sci. 10:103–109. 2017. View Article : Google Scholar
|
|
25
|
Bak DH, Kang SH, Park CH, Chung BY and Bai
HW: A novel radiolytic rotenone derivative, rotenoisin A, displays
potent anticarcinogenic activity in breast cancer cells. J Radiat
Res. 62:249–258. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Tharmarajah L, Samarakoon SR, Ediriweera
MK, Piyathilaka P, Tennekoon KH, Senathilake KS, Rajagopalan U,
Galhena PB and Thabrew I: In vitro anticancer effect of gedunin on
human teratocarcinomal (NTERA-2) cancer stem-like cells. Biomed Res
Int. 2017:24131972017. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Zheng SY, Li Y, Jiang D, Zhao J and Ge JF:
Anticancer effect and apoptosis induction by quercetin in the human
lung cancer cell line A-549. Mol Med Rep. 5:822–826.
2012.PubMed/NCBI
|
|
28
|
Samarghandian S and Shabestari MM: DNA
fragmentation and apoptosis induced by safranal in human prostate
cancer cell line. Indian J Urol. 29:177–183. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Varshney L and Dodke P: Radiation effect
studies on anticancer drugs, cyclophosphamide and doxorubicin for
radiation sterilization. Radiat Phys Chem. 71:1103–1111. 2004.
View Article : Google Scholar
|
|
30
|
Hengartner MO: The biochemistry of
apoptosis. Nature. 407:770–776. 2000. View
Article : Google Scholar : PubMed/NCBI
|