Introduction
Diabetic retinopathy (DR) is a common complication
of diabetes (1). With the
increasing prevalence of diabetes, it is estimated that the number
of patients with DR will reach 191 million worldwide by 2030
(2). In diabetes,
hyperglycemia-mediated injury of blood vessels and capillaries
causes retina ischemia and hypoxia, and vitreous hemorrhage, which
leads to proliferative diabetic retinopathy, and ultimately
blindness (1). At present, the
local therapeutic interventions for the treatment of DR include
intravitreal injection of anti-vascular endothelial growth factor
(VEGF) drugs, glucocorticoids and laser photocoagulation (3). However, due to the complicated
pathogenesis of DR, the current therapeutic interventions cannot
completely prevent complications, including vitreous hemorrhage,
traction retinal detachment and neovascular glaucoma (4). The primary pathological features of DR
are retinal vascular occlusive circulation disorder, as well as
retinal endothelial cell proliferation, migration and angiogenesis
(5,6). Therefore, inhibiting retinal
neovascularization is important for the treatment of DR.
Mangiferin
(1,3,6,7-tetrahydroxyxanthone-C2-β-D-glucoside) is a xanthone
glucoside. Mangiferin can be extracted from plants belonging to the
Liliaceae, Anacardiaceae and Gentianaceae families
(7,8). Mangiferin exerts a wide range of
pharmacological effects, including antitumor, anti-infectious,
hypoglycemic, antioxidation, and immune regulatory effects
(9). A previous study demonstrated
that mangiferin attenuates dexamethasone-induced osteoblastic
MC3T3-E1 cell apoptosis and oxidative stress via activating the
bone morphogenetic protein 2/Smad-1 signaling pathway (10). In atherosclerosis, mangiferin
promoted macrophage cholesterol efflux via enhancing ATP binding
cassette subfamily (ABC)A1 and ABCG1 expression (8). Moreover, it has been reported that the
PI3K/AKT and Rac1/Wiskott-Aldrich syndrome protein family member 2
(WAVE2) signaling pathways are blocked by mangiferin in cancer
(11). In particular, mangiferin
exerted beneficial effects on biochemical and hematological
parameters in rats with streptozotocin-induced diabetes (12). However, whether mangiferin protects
against high glucose (HG)- and hypoxia-induced injury in rat
retinal capillary endothelial cells (RRCECs) is not completely
understood. Thus, the present study aimed to investigate the
effects of mangiferin on RRCEC viability, migration and
angiogenesis following exposure to HG and hypoxia.
Materials and methods
Drug
Mangiferin (C19H18O11; molecular weight, 422.34;
purity, ≥98%; cat. no. 4773-96-0) was purchased from Sigma-Aldrich
(Merck KGaA).
Cell culture
RRCECs (Procell Life Science & Technology Co.,
Ltd.) were cultured in HG DMEM (cat. no. PM150220; Procell Life
Science & Technology Co., Ltd.) supplemented with 10% FBS (cat.
no. FB15015; Clark Bioscience) and 1% penicillin/streptomycin (cat.
no. sv30010; HyClone; Cytiva) with 5% CO2 at 37°C.
Cell treatment
To select the optimum concentration (A0), RRCECs
were treated with different concentrations of mangiferin (0.01,
0.1, 1 or 10 mM) for 24, 48 and 72 h, as previously described
(10,13). Subsequently, RRCECs were pretreated
with D-(+)-glucose (30 mM; cat. no. G7528; Sigma-Aldrich; Merck
KGaA)/hypoxia (1% O2, 94% N2 and 5% CO2) and then were treated with
0.1 mM (A0) mangiferin for 24 or 48 h. RRCECs were also treated
with D-(+)-glucose/hypoxia and followed by different doses of
mangiferin [0.05 mM (1/2 A0), 0.1 mM (A0) and 0.2 mM (2A0)] for 24
h at 37°C. Finally, RRCECs were treated with a PI3K/AKT signaling
pathway inhibitor (LY294002; Sigma-Aldrich; Merck KGaA 40 µmol/l)
or a PI3K/AKT signaling pathway activator (IGF-1; Sigma-Aldrich;
Merck KGaA 100 ng/ml) in the presence of D-(+)-glucose/hypoxia and
mangiferin (A0) for 24 h at 37°C. The control cells were treated
the same amount of DMSO for 24 h at 37°C. Cells were divided into
the following nine groups: i) Control; ii) HG/hypoxia; iii)
HG/hypoxia + 1/2 A0 mangiferin; iv) HG/hypoxia + A0 mangiferin; v)
HG/hypoxia + 2A0 mangiferin; vi) HG/hypoxia + 0.1 mM (A0)
mangiferin + LY294002; vii) and HG/hypoxia + 0.1 mM (A0) mangiferin
+ IGF-1; viii) HG/hypoxia + LY294002; ix) HG/hypoxia + IGF-1.
Western blotting
RRCECs were plated (1×106 cells) into
100-mm culture plates. The following day, cells were treated with
mangiferin as aforementioned. Following washing three times with
PBS, total protein was extracted from cells using RIPA buffer (cat.
no. P0013C; Beyotime Institute of Biotechnology) supplemented with
1% phenylmethylsulfonyl fluoride (Beyotime Institute of
Biotechnology). Protein concentrations were determined using a BCA
assay kit (CoWin Biosciences). Proteins (40 µg) were separated
alongside molecular weight standards (Bio-Rad Laboratories, Inc.)
via 10% SDS-PAGE. Proteins were transferred onto PVDF membranes
(Amersham; Cytiva). Following blocking with 5% non-fat milk at room
temperature for 1 h, the membranes were incubated overnight at 37°C
with anti-matrix metallopeptidase (MMP)2 (1:1,000; cat. no. AF5330;
Affinity Biosciences), anti-MMP9 (1:500; cat. no. ab119906; Abcam),
anti-VEGF (1:1,000; cat. no. ab46154; Abcam),
anti-hypoxia-inducible factor-1α (HIF-1α; 1:500; cat. no. ab1;
Abcam), anti-PI3K (1:1,000; cat. no. ab32089; Abcam), anti-AKT
(1:10,000; cat. no. ab179463; Abcam), anti-mTOR (1:1,000; cat. no.
ab32028; Abcam), anti-phosphorylated (p)-PI3K (1:500; cat. no.
ab182651; Abcam), anti-p-AKT (1:5,000; cat. no. ab81283; Abcam),
anti-p-mTOR (1:1,000; cat. no. ab109268; Abcam) and anti-β-actin
(1:1,000; cat. no. AC026; ABclonal Biotech Co., Ltd.).
Subsequently, the membranes were incubated with a HRP-conjugated
Goat Anti-Rabbit IgG secondary antibody (1:2,000; cat. no. ab6721;
Abcam) or a HRP-conjugated Goat Anti-Mouse IgG secondary antibody
(1:2,000; cat. no. ab6728; Abcam) at room temperature for 3 h.
Protein bands were visualized using an ECL system (cat. no. KF001;
Affinity Biosciences). Protein expression levels were
semi-quantified using Image-Pro Plus software (version 6.0; Media
Cybernetics, Inc.) with β-actin as the loading control.
Cell viability assay
RRCEC viability was assessed by performing Cell
Counting Kit-8 (CCK-8) assays (cat. no. BS350B; Biosharp Life
Sciences) according to the manufacturer's protocol. CCK-8 solution
was added to each well and incubated at 37°C for 1.5 h. Cell
viability was measured at a wavelength of 450 nm using a microplate
reader.
Immunofluorescence
RRCECs were seeded (5×104 cells/well)
into 96-well plates and cultured at 37°C with 5% CO2. At 90%
confluence, the culture medium was removed and cells were washed
with PBS. The cells were fixed with 4% paraformaldehyde at room
temperature for 30 min and washed 3 times with 0.02 M PBS at room
temperature for 3 min. Following blocking with 10% serum blocking
solution (cat. no. 22012-8612; Zhejiang Tianhang Biotechnology Co.,
Ltd.) at room temperature for 30 min, cells were incubated with an
anti-Ki67 primary antibody (1:200; cat. no. GB13030-2; Wuhan
Servicebio Technology Co., Ltd.) at 4°C overnight. Subsequently,
cells were incubated with rhodamine-labeled goat anti-rabbit IgG
antibody (1:100; cat. no. ZF-0316; OriGene Technologies, Inc.) at
37°C for 30 min. Finally, cell nuclei were labeled with DAPI (cat.
no. ZLI-9557; OriGene Technologies, Inc.) at room temperature for
10 min and the cells were incubated with anti-fluorescence decay
blocking solution at room temperature (cat. no. AR1109; Wuhan
Boster Biological Technology, Ltd.). Stained cells were observed
using a fluorescence microscope (magnification, ×400; Olympus
Corporation).
Tube formation assay
First, 60 µl dissolved Matrigel adhesive matrix was
added into each well of a pre-cooled 96-well plate and allowed to
solidify at 37°C for 1 h. RRCECs (5×104 cells/ml) were
added into each well, cultured under hypoxic conditions (1% O2, 94%
N2 and 5% CO2) for 6 h and then transferred to normoxic conditions
(95% air and 5% CO2) for 12 h. Subsequently, cells were treated
with mangiferin (0.05, 0.1 or 0.2 mM) at 37°C with 5% CO2 for 24 h.
Tube formation was observed under an inverted phase-contrast light
microscope in five randomly selected fields of view.
Wound healing assay
RRCEC migration was assessed by performing a wound
healing assay. Briefly, cells were grown in DMEM medium containing
10% FBS and then seeded (5×105 cells) into a 6-well
plate. At 100% confluence, a wound was created in each cell
monolayer using a sterile micropipette tip, followed by washing
with PBS to remove cell debris. Subsequently, cells were cultured
in serum-free DMEM under hypoxic conditions for 6 h, and then
transferred to normoxic conditions for 12 h. Finally, cells were
treated with different concentrations of mangiferin (0.05, 0.1 or
0.2 mM) for 24 h at 37°C with 5% CO2. An inverted phase-contrast
light microscope (Olympus Corporation) was used to obtain images of
the wound at different time points. The width of the wound was
measured at three different positions at 0 and 24 h. The relative
wound recovery distance was calculated as 0–24 h.
Transwell assay
To evaluate cell migration, RRCECs were resuspended
(1×106 cells/ml) in serum-free DMEM. Transwell chambers
were placed into a 24-well plate. Subsequently, cell suspension (20
µl/well) was added into the upper chamber. Cells were cultured
under hypoxic conditions for 6 h and then normoxic conditions for
12 h. Subsequently, 600 µl DMEM supplemented with 10% FBS (cat. no.
FB15015; Clark Bioscience) was added into the lower chamber. Cells
in the upper chamber were treated with mangiferin (0.05, 0.1 or 0.2
mM) for 24 h and cultured at 37°C with 5% CO2. Following fixation
with 4% paraformaldehyde for 20 min at room temperature, cells were
stained with 0.1% crystal violet for 15 min at room temperature.
Migratory cells were visualized using an inverted phase-contrast
light microscope (Olympus Corporation).
Statistical analysis
Statistical analysis was performed on the means of
triplicates that resulted from three independent replications of
each experiment. Data are presented as the mean ± SD. Statistical
analyses were performed using SPSS software (version 20.0; IBM
Corp.). Comparisons among groups were analyzed using one-way ANOVA
followed by Tukey's post hoc test. P<0.05 was considered to
indicate a statistically significant difference.
Results
Effect of mangiferin on
HG/hypoxia-induced RRCEC viability
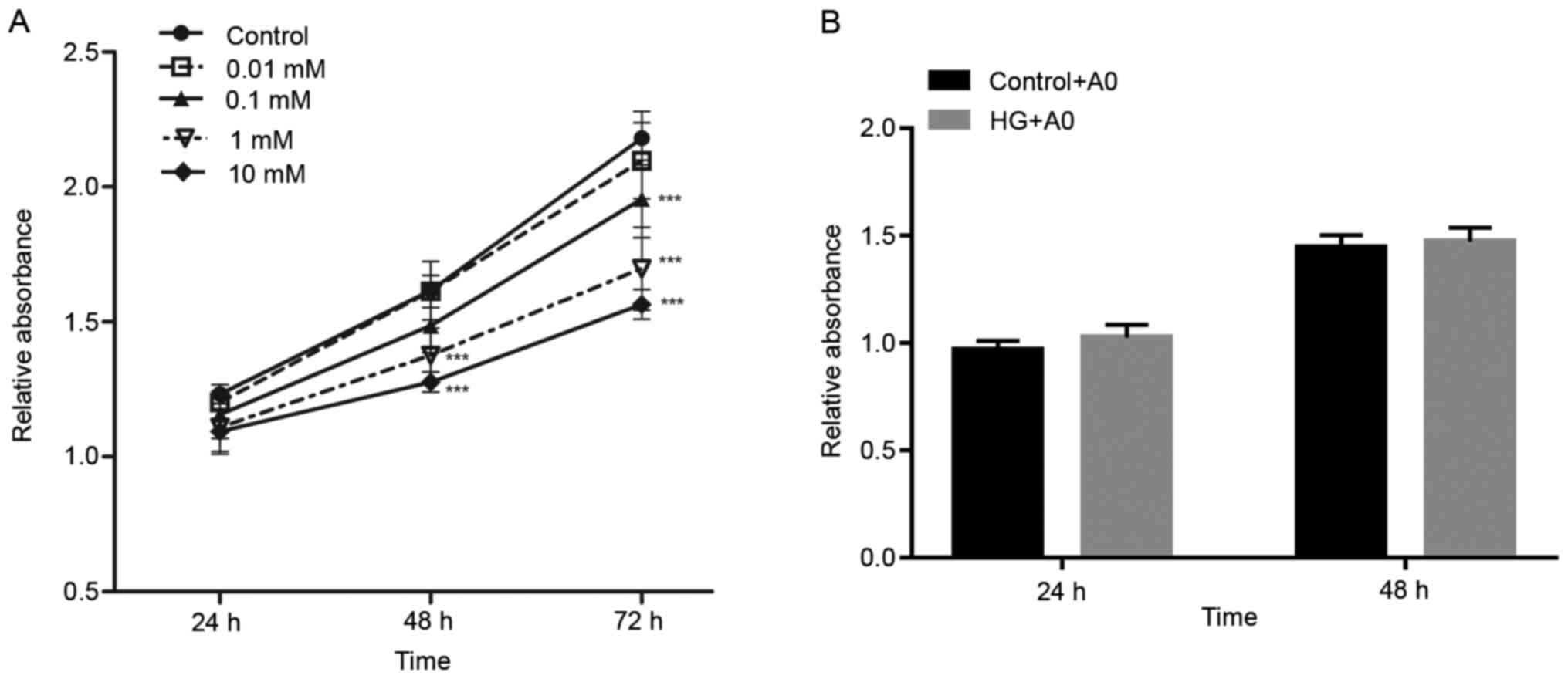
Cell viability was assessed by performing CCK-8
assays. The results demonstrated that mangiferin (0.01–10 mM)
decreased RRCEC viability in a dose-dependent manner (Fig. 1). Furthermore, treatment of RRCECs
with mangiferin for 24, 48 and 72 h increased cell viability in a
time-dependent manner (Fig. 1).
Following treatment with A0 mangiferin for 48 h, no significant
alteration in cell viability was observed compared with the control
group (Fig. 1A). In addition,
RRCECs were treated with A0 mangiferin and induced with HG/hypoxia
for 24 and 48 h. Co-treatment of RRCECs with HG and A0 mangiferin
for 24 and 48 h did not significantly alter cell viability compared
with the control + A0 group (Fig.
1B). Therefore, the results indicated that A0 mangiferin did
not display cytotoxicity.
Mangiferin attenuates
HG/hypoxia-induced RRCEC migration and angiogenesis
Subsequently, the effects of mangiferin on
HG/hypoxia-induced viability, migration and angiogenesis were
investigated. Compared with the control group, HG/hypoxia exposure
significantly increased RRCEC viability, whereas mangiferin did not
display significant inhibitory or promotive effects on RRCEC
viability (Fig. 2A). Furthermore,
the immunofluorescence results demonstrated that compared with the
control group, the number of Ki67+ cells was clearly
increased following treatment with HG/hypoxia, an effect that was
not notably reversed by mangiferin (Fig. 2B). In addition, HG/hypoxia exposure
significantly increased RRCEC migration compared with the control
group (Fig. 2C-F). Treatment with
1/2 A0, A0 or 2A0 mangiferin markedly attenuated
HG/hypoxia-mediated effects on cell migration (Fig. 2C-F). Compared with the control
group, RRCEC angiogenesis was markedly enhanced following treatment
with HG/hypoxia, which was notably attenuated by treatment with 1/2
A0, A0 or 2A0 mangiferin (Fig. 2G).
Furthermore, following exposure to HG/hypoxia, the expression
levels of HIF-1α, VEGF, MMP2 and MMP9 in RRCECs were significantly
upregulated compared with the control group. However, treatment
with A0 or 2A0 mangiferin significantly reversed HG/hypoxia
exposure-induced upregulation of HIF-1α, VEGF, MMP2 and MMP9
expression levels (Fig. 2H and
I).
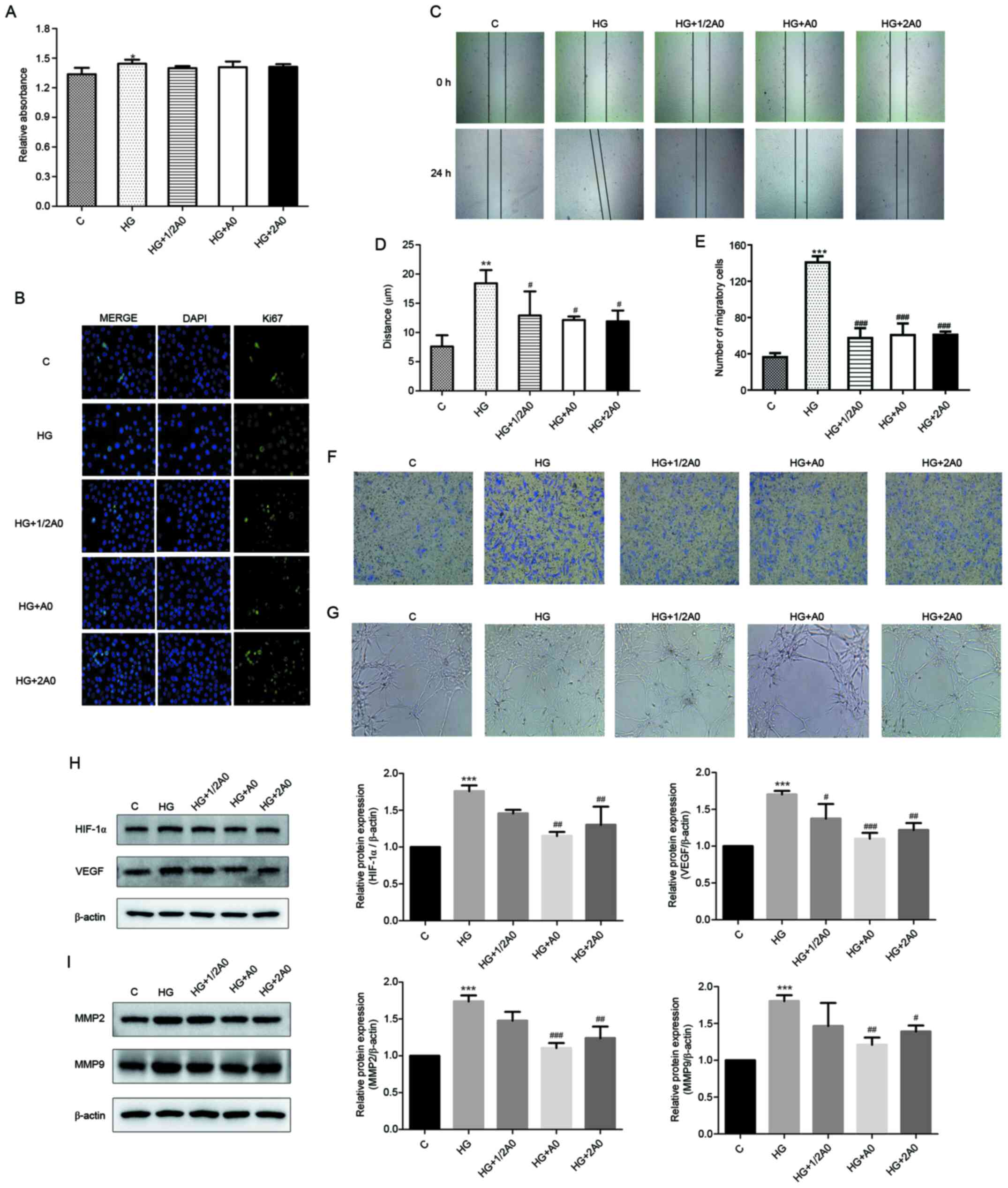 | Figure 2.Mangiferin attenuates
HG/hypoxia-induced RRCEC migration and angiogenesis. (A) RRCEC
viability was evaluated by performing Cell Counting Kit-8 assays.
(B) Ki67 expression was determined via immunofluorescence
(magnification, ×400). RRCEC migration was (C) determined by
performing wound healing assay (magnification, ×50) and (D)
quantified. (E) Quantification of RRCEC migration as determined by
(F) performing Transwell assays (magnification, ×100). (G) Tube
formation and branching points were determined (magnification,
×100). Protein expression levels of (H) HIF-1α, VEGF, (I) MMP2 and
MMP9 were determined via western blotting. **P<0.01 and
***P<0.001 vs. control; #P<0.05,
##P<0.01 and ###P<0.001 vs. HG. HG,
high glucose; RRCEC, rat retinal capillary endothelial cell;
HIF-1α, hypoxia-inducible factor-1α; VEGF, vascular endothelial
growth factor; MMP2, matrix metallopeptidase 2; C, control; 1/2 A0,
0.05 mM mangiferin; A0, 0.1 mM mangiferin; 2A0, 2 mM
mangiferin. |
Mangiferin suppresses the activity of
the PI3K/AKT and mTOR signaling pathways in HG/hypoxia-treated
RRCECs
The PI3K/AKT and mTOR signaling pathways serve
important roles in various intracellular events, including cell
migration, invasion and angiogenesis (14,15).
Therefore, the present study explored whether these signaling
pathways were implicated in mangiferin-mediated cell migration and
angiogenesis. The western blotting results demonstrated that the
protein expression levels of PI3K, p-PI3K, p-AKT, mTOR and p-mTOR
were significantly upregulated in HG/hypoxia-exposed cells compared
with control cells (Fig. 3).
However, the levels of PI3K, AKT and mTOR phosphorylation were
significantly decreased following treatment with A0 mangiferin in
HG/hypoxia-exposed cells (Fig. 3).
Similar results were observed for the p-AKT/AKT, p-PI3K/PI3K and
p-mTOR/mTOR ratios (Fig. 3D, G and
J).
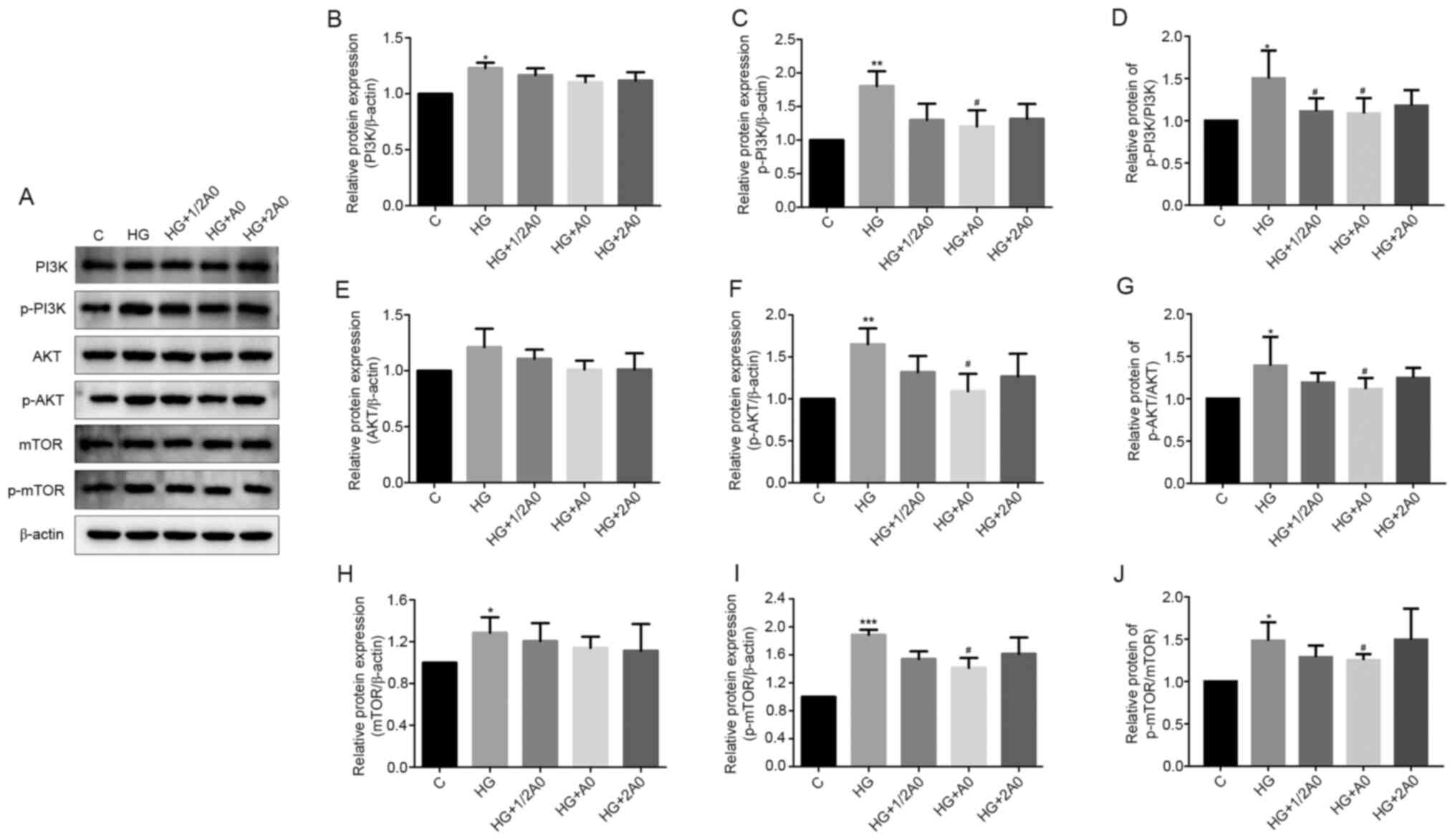 | Figure 3.Mangiferin suppresses PI3K/AKT/mTOR
signaling pathway activation in HG/hypoxia-treated RRCECs. Protein
expression levels were (A) determined via western blotting and
semi-quantified for (B PI3K, (C) p-PI3K, (D) p-PI3K/PI3K, (E) AKT,
(F) p-AKT, (G) p-AKT/AKT, (H) mTOR, (I) p-mTOR and (J) p-mTOR/mTOR.
*P<0.05, **P<0.01 and ***P<0.001 vs. control;
#P<0.05 vs. HG. HG, high glucose; RRCEC, rat retinal
capillary endothelial cell; p, phosphorylated; C, control; 1/2 A0,
0.05 mM mangiferin; A0, 0.1 mM mangiferin; 2A0, 2 mM
mangiferin. |
Mangiferin inhibits HG/hypoxia-induced
RRCEC migration and angiogenesis via the PI3K/AKT and mTOR
signaling pathways
The present study investigated whether the PI3K/AKT
and mTOR signaling pathways were involved in cell migration and
angiogenesis following exposure to HG/hypoxia. The western blotting
results demonstrated that the PI3K/AKT signaling pathway inhibitor
(LY294002) and the PI3K/AKT signaling pathway activator (IGF-1)
significantly downregulated and upregulated the expression levels
of p-PI3K, p-AKT and p-mTOR in HG/hypoxia-exposed and
mangiferin-treated RRCECs, respectively (Fig. 4A-J). Mangiferin-mediated
downregulation of p-AKT/AKT, p-PI3K/PI3K and p-mTOR/mTOR ratios was
significantly reversed by IGF-1 and significantly enhanced by
LY294002 (Fig. 4A-J). However,
RRCEC viability was not significantly altered among the groups
(Fig. 4K). Additionally,
mangiferin-induced inhibition of RRCEC migration was significantly
enhanced and inhibited following treatment with LY294002 and IGF-1,
respectively (Fig. 4L-O).
Furthermore, the tube formation assay results demonstrated that A0
mangiferin markedly reduced the number of branching points in
HG/hypoxia-treated cells, an effect that was notably enhanced and
inhibited following treatment with LY294002 and IGF-1, respectively
(Fig. 4P). Overall, the results of
the present study suggested that mangiferin inhibited RRCEC
migration and angiogenesis partially via deactivation of the
PI3K/AKT and mTOR signaling pathways.
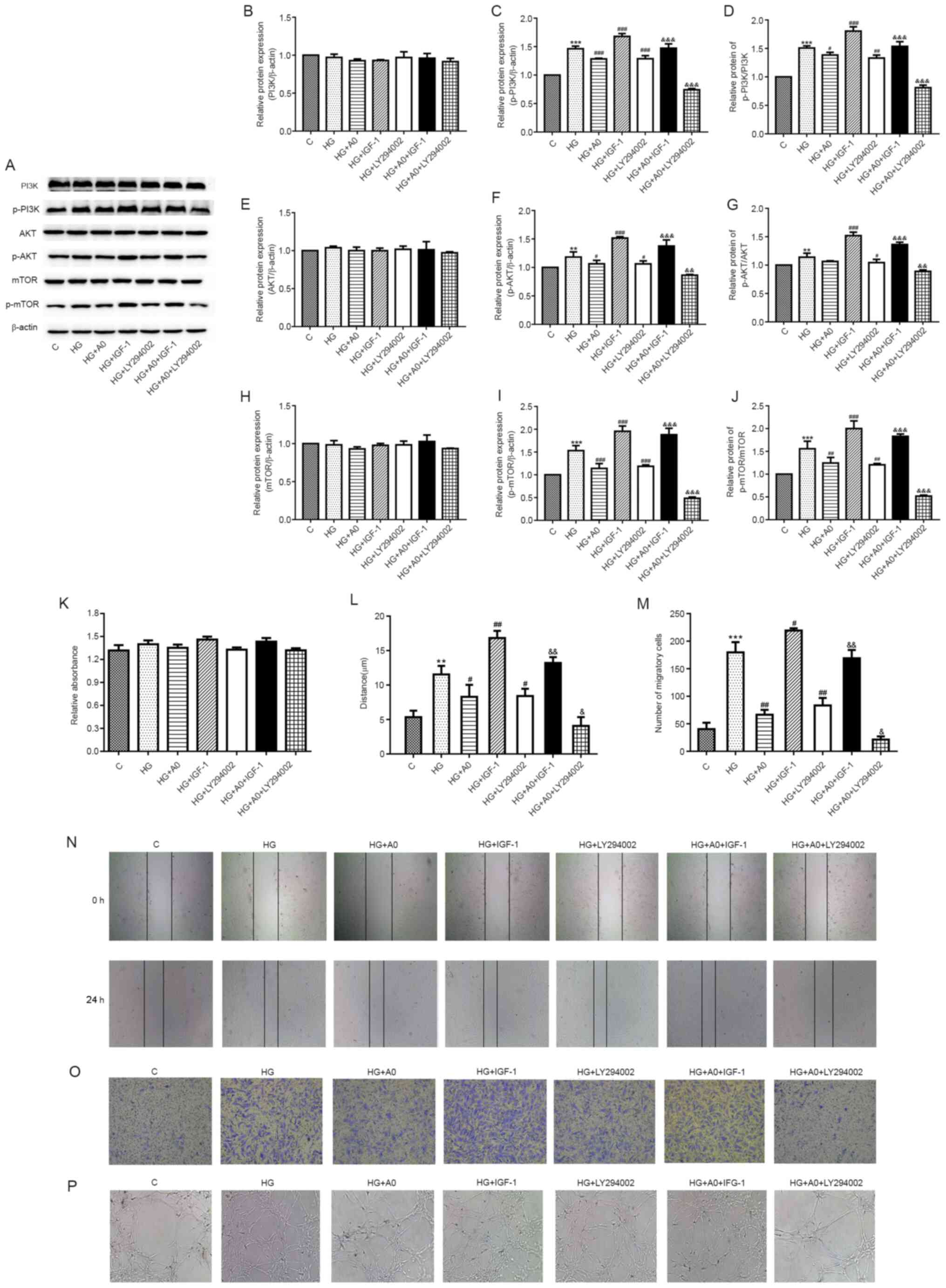 | Figure 4.Mangiferin inhibits
HG/hypoxia-induced cell migration and angiogenesis via the
PI3K/AKT/mTOR signaling pathways in RRCECs. Protein expression
levels were (A) determined via western blotting and semi-quantified
for (B) PI3K, (C) p-PI3K, (D) p-PI3K/PI3K, (E) AKT, (F) p-AKT, (G)
p-AKT/AKT, (H) mTOR, (I) p-mTOR and (J) p-mTOR/mTOR. (K) RRCEC
viability was evaluated by performing Cell Counting Kit-8 assays.
RRCEC migration was evaluated by performing (L) wound healing and
(M) Transwell assay. Representative images of the (N) wound healing
(×50) and (O) Transwell assays (magnification, ×100). (P) Tube
formation and branching points were determined magnification,
×100). **P<0.01 and ***P<0.001 vs. control;
#P<0.05, ##P<0.01 and
###P<0.001 vs. HG; &P<0.05,
&&P<0.01 and
&&&P<0.001 vs. HG+A0. HG, high glucose;
RRCEC, rat retinal capillary endothelial cell; p, phosphorylated;
C, control; A0, 0.1 mM mangiferin. |
Discussion
Long-term exposure to hyperglycemia and hypoxia is
highly associated with the development of DR pathological states,
and is considered the primary cause of blood-retinal barrier and
microvascular injury (16).
Mangiferin, a natural bioactive ingredient, is commonly found in
mango trees, and has been reported to display antitumor,
antidiabetic, lipometabolism regulating, anti-inflammatory and
antibacterial properties (17).
Previous studies have reported that mangiferin exerts
pharmacological effects on hypoglycemia and diabetes. In
streptozotocin-induced type 2 diabetic rats, mangiferin ameliorates
insulin sensitivity and dyslipidemia, depletes serum TNF-α and
elevates adiponectin (18).
Mangiferin suppresses increased plasma insulin and fasted plasma
non-esterified fatty acid concentrations at the baseline during an
oral glucose tolerance test in a rat model of fructose-induced
metabolic syndrome (19).
Furthermore, at 3 weeks post-oral administration, mangiferin
decreases blood glucose levels in KK-Ay mice, but displays no
effect on blood glucose levels in healthy mice (20). Moreover, mangiferin clearly improves
reactive oxygen species (ROS) generation, inflammation and
endoplasmic reticulum stress in gestational diabetes mellitus model
mice (21). Mangiferin increases
bovine aortic cell migration, but displays no effect on cell
proliferation in vitro (22). The results of the present study
demonstrated that mangiferin displayed no significant effect on
cell viability in HG/hypoxia-induced RRCECs. Additionally,
treatment with mangiferin markedly attenuated HG/hypoxia-induced
RRCEC migration and angiogenesis, which suggested that mangiferin
may be useful for the treatment of DR. Mechanistically, mangiferin
inhibits glioma cell proliferation and promotes apoptosis via
downregulating MMP9 expression (23). Furthermore, mangiferin reduces MMP7
and MMP9 expression, and inhibits breast cancer cell proliferation
and epithelial-mesenchymal transition (EMT) in vitro and
in vivo via activation of the β-catenin signaling pathway
(24). Increasing evidence has
demonstrated that mangiferin attenuates inflammation via inhibiting
the Toll-like receptor 4/p65 signaling pathway, and suppresses the
EMT process via blocking the Smad2/3 signaling pathway and
downregulating MMP9 expression in pulmonary fibrosis model mice
(25). Similarly, the results of
the present study demonstrated that mangiferin significantly
decreased MMP2 and MMP9 expression levels in HG/hypoxia-exposed
RRCECs.
HIF-α/VEGF expression serves a major role in
mediating active retinal angiogenesis and alleviating the
progression of DR. In DR, HIF-1α regulates the transcription of
VEGF, which ultimately leads to intracellular accumulation of VEGF
(26). VEGF is an important
stimulating factor involved in retinal and choroidal
neovascularization (27,28). Clinical data indicates that the
concentration of VEGF is enhanced in vitreous fluid of patients
with DR (29). It has been reported
that anti-VEGF agents may be used to treat angiogenesis in eye
diseases, including neovascular glaucoma, persistent vitreous
hemorrhage and DR (30). In the
present study, mangiferin treatment significantly downregulated
HG/hypoxia-induced HIF-α and VEGF expression levels.
In addition, several signaling pathways, including
the PI3K/AKT, STAT3 and mTOR signaling pathways, are associated
with HIF-1α/VEGF signaling in DR (31–33).
It has been reported that the PI3K/AKT/mTOR signaling pathway
promotes DR via inducing retinal cell viability, apoptosis,
migration and angiogenesis. The use of a combination of
PI3K/AKT/mTOR drugs is safe and effective for the treatment of
ocular neovascularization (34). In
retinal endothelial RF/6A and microglia BV-2 cells, erianin
significantly suppresses HG-induced increases in retinal vessels
and microglia activation via deactivating the VEGF/PI3K/AKT
signaling pathway (35).
Furthermore, microRNA-183 overexpression activates the PI3K/AKT
signaling pathway, increases VEGF expression and downregulates
B-cell translocation gene 1 expression in DR (36). In addition, Nogo-B induced VEGF
secretion and activation of the PI3K/AKT axis in HG-exposes human
retinal microvascular endothelial cells (37). A previous study reported that
Parkinsonism associated deglycase overexpression significantly
reduces ROS production, p-AKT and p-mTOR expression, and cell
apoptosis in HG-induced retinal capillary pericytes (38). Similarly, the present study
demonstrated that A0 mangiferin significantly inhibited the
PI3K/AKT/mTOR signaling pathway in HG/hypoxia-induced RRCECs.
Activation of the PI3K/AKT signaling pathway by IGF-1 clearly
reversed mangiferin-mediated effects on RRCEC migration and
angiogenesis. Moreover, A20 mangiferin did not significantly reduce
the phosphorylation levels of PI3K, AKT and mTOR in
HG/hypoxia-induced RRCECs, which suggested that high concentration
mangiferin was not conducive to the inhibition of protein
phosphorylation. It was hypothesized that high concentrations of
mangiferin might activate or inhibit additional upstream signaling
pathways to impair the phosphorylation levels of PI3K, AKT and
mTOR; however, further investigations are required to confirm the
effects of high concentration mangiferin on the upstream components
of the PI3K/AKT signaling pathway.
In summary, to the best of our knowledge, the
present study demonstrated for the first time that mangiferin
displayed beneficial effects against DR by suppressing
HG/hypoxia-induced RRCEC migration and angiogenesis via inhibition
of the PI3K/AKT/mTOR signaling pathway, suggesting the potential
use of mangiferin for the treatment of DR.
Acknowledgements
Not applicable.
Funding
This study was supported by the Sichuan Province
Science and Technology Support Program (grant no. 2015SZ0086).
Availability of data and materials
The datasets used or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
JS, CT, YL and HL conceptualized and designed the
study. JS, CT, YL, JH and HZ performed the experiments. JS, CT and
YL analyzed the data. JS, CT, YL and HL drafted the manuscript. JS
and HL confirmed the authenticity of all the raw data. All authors
read and approved the final version of the manuscript.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Cheung N, Mitchell P and Wong TY: Diabetic
retinopathy. Lancet. 376:124–136. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Ting DS, Cheung GC and Wong TY: Diabetic
retinopathy: global prevalence, major risk factors, screening
practices and public health challenges: a review. Clin Exp
Ophthalmol. 44:260–277. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Wang W and Lo AC: Diabetic retinopathy:
Pathophysiology and treatments. Int J Mol Sci. 19:18162018.
View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Whitehead M, Wickremasinghe S, Osborne A,
Van Wijngaarden P and Martin KR: Diabetic retinopathy: A complex
pathophysiology requiring novel therapeutic strategies. Expert Opin
Biol Ther. 18:1257–1270. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Kim D, Lee D, Trackman PC and Roy S:
Effects of high glucose-induced lysyl oxidase propeptide on retinal
endothelial cell survival: Implications for diabetic retinopathy.
Am J Pathol. 189:1945–1952. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Mei X, Zhou L, Zhang T, Lu B, Sheng Y and
Ji L: Chlorogenic acid attenuates diabetic retinopathy by reducing
VEGF expression and inhibiting VEGF-mediated retinal
neoangiogenesis. Vascul Pharmacol. 101:29–37. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Imran M, Arshad MS, Butt MS, Kwon JH,
Arshad MU and Sultan MT: Mangiferin: A natural miracle bioactive
compound against lifestyle related disorders. Lipids Health Dis.
16:842017. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Ren K, Li H, Zhou HF, Liang Y, Tong M,
Chen L, Zheng XL and Zhao GJ: Mangiferin promotes macrophage
cholesterol efflux and protects against atherosclerosis by
augmenting the expression of ABCA1 and ABCG1. Aging (Albany NY).
11:10992–11009. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Núñez Selles AJ, Daglia M and Rastrelli L:
The potential role of mangiferin in cancer treatment through its
immunomodulatory, anti-angiogenic, apoptopic, and gene regulatory
effects. Biofactors. 42:475–491. 2016. View Article : Google Scholar
|
|
10
|
Ding LZ, Teng X, Zhang ZB, Zheng CJ and
Chen SH: Mangiferin inhibits apoptosis and oxidative stress via
BMP2/Smad-1 signaling in dexamethasone-induced MC3T3-E1 cells. Int
J Mol Med. 41:2517–2526. 2018.PubMed/NCBI
|
|
11
|
Deng Q, Tian YX and Liang J: Mangiferin
inhibits cell migration and invasion through Rac1/WAVE2 signalling
in breast cancer. Cytotechnology. 70:593–601. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Sellamuthu PS, Arulselvan P, Fakurazi S
and Kandasamy M: Beneficial effects of mangiferin isolated from
Salacia chinensis on biochemical and hematological
parameters in rats with streptozotocin-induced diabetes. Pak J
Pharm Sci. 27:161–167. 2014.PubMed/NCBI
|
|
13
|
Zhang Q, Kong X, Yuan H, Guan H, Li Y and
Niu Y: Mangiferin improved palmitate-induced-insulin resistance by
promoting free fatty acid metabolism in HepG2 and C2C12 cells via
PPARα: mangiferin improved insulin resistance. J Diabetes Res.
2019:20526752019. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Karar J and Maity A: PI3K/AKT/mTOR pathway
in angiogenesis. Front Mol Neurosci. 4:512011. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Huang W, Ding X, Ye H, Wang J, Shao J and
Huang T: Hypoxia enhances the migration and invasion of human
glioblastoma U87 cells through PI3K/Akt/mTOR/HIF-1α pathway.
Neuroreport. 29:1578–1585. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Nyengaard JR, Ido Y, Kilo C and Williamson
JR: Interactions between hyperglycemia and hypoxia: Implications
for diabetic retinopathy. Diabetes. 53:2931–2938. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Du S, Liu H, Lei T, Xie X, Wang H, He X,
Tong R and Wang Y: Mangiferin: An effective therapeutic agent
against several disorders (Review). Mol Med Rep. 18:4775–4786.
2018.PubMed/NCBI
|
|
18
|
Saleh S, El-Maraghy N, Reda E and Barakat
W: Modulation of diabetes and dyslipidemia in diabetic
insulin-resistant rats by mangiferin: Role of adiponectin and
TNF-α. An Acad Bras Cienc. 86:1935–1948. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Zhou L, Pan Y, Chonan R, Batey R, Rong X,
Yamahara J, Wang J and Li Y: Mitigation of insulin resistance by
mangiferin in a rat model of fructose-induced metabolic syndrome is
associated with modulation of CD36 redistribution in the skeletal
muscle. J Pharmacol Exp Ther. 356:74–84. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Miura T, Ichiki H, Hashimoto I, Iwamoto N,
Kato M, Kubo M, Ishihara E, Komatsu Y, Okada M, Ishida T, et al:
Antidiabetic activity of a xanthone compound, mangiferin.
Phytomedicine. 8:85–87. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Sha H, Zeng H, Zhao J and Jin H:
Mangiferin ameliorates gestational diabetes mellitus-induced
placental oxidative stress, inflammation and endoplasmic reticulum
stress and improves fetal outcomes in mice. Eur J Pharmacol.
859:1725222019. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Daud NH, Aung CS, Hewavitharana AK,
Wilkinson AS, Pierson JT, Roberts-Thomson SJ, Shaw PN, Monteith GR,
Gidley MJ and Parat MO: Mango extracts and the mango component
mangiferin promote endothelial cell migration. J Agric Food Chem.
58:5181–5186. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Xiao J, Liu L, Zhong Z, Xiao C and Zhang
J: Mangiferin regulates proliferation and apoptosis in glioma cells
by induction of microRNA-15b and inhibition of MMP-9 expression.
Oncol Rep. 33:2815–2820. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Li H, Huang J, Yang B, Xiang T, Yin X,
Peng W, Cheng W, Wan J, Luo F, Li H, et al: Mangiferin exerts
antitumor activity in breast cancer cells by regulating matrix
metalloproteinases, epithelial to mesenchymal transition, and
β-catenin signaling pathway. Toxicol Appl Pharmacol. 272:180–190.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Jia L, Sun P, Gao H, Shen J, Gao Y, Meng
C, Fu S, Yao H and Zhang G: Mangiferin attenuates bleomycin-induced
pulmonary fibrosis in mice through inhibiting TLR4/p65 and
TGF-β1/Smad2/3 pathway. J Pharm Pharmacol. 71:1017–1028. 2019.
View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Wang N, Zhang C, Tan H, et al: Insulin
accelerates progression of diabetic retinopathy through activating
HIF-1α/VEGF pathway in retinal endothelial. cells. 2019.
|
|
27
|
Gao X, Li Y, Wang H, Li C and Ding J:
Inhibition of HIF-1α decreases expression of pro-inflammatory IL-6
and TNF-α in diabetic retinopathy. Acta Ophthalmol. 95:e746–e750.
2017. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Zhang D, Lv FL and Wang GH: Effects of
HIF-1α on diabetic retinopathy angiogenesis and VEGF expression.
Eur Rev Med Pharmacol Sci. 22:5071–5076. 2018.PubMed/NCBI
|
|
29
|
Aiello LP, Avery RL, Arrigg PG, Keyt BA,
Jampel HD, Shah ST, Pasquale LR, Thieme H, Iwamoto MA, Park JE, et
al: Vascular endothelial growth factor in ocular fluid of patients
with diabetic retinopathy and other retinal disorders. N Engl J
Med. 331:1480–1487. 1994. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Osaadon P, Fagan XJ, Lifshitz T and Levy
J: A review of anti-VEGF agents for proliferative diabetic
retinopathy. Eye (Lond). 28:510–520. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Cui J, Gong R, Hu S, Cai L and Chen L:
Gambogic acid ameliorates diabetes-induced proliferative
retinopathy through inhibition of the HIF-1α/VEGF expression via
targeting PI3K/AKT pathway. Life Sci. 192:293–303. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Yang X, Cao J, Du Y, Gong Q, Cheng Y and
Su G: Angiopoietin-like protein 4 (ANGPTL4) induces retinal pigment
epithelial barrier breakdown by activating signal transducer and
activator of transcription 3 (STAT3): Evidence from ARPE-19 cells
under hypoxic condition and diabetic rats. Med Sci Monit.
25:6742–6754. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Wei J, Jiang H, Gao H and Wang G: Blocking
mammalian target of rapamycin (mTOR) attenuates HIF-1α pathways
engaged-vascular endothelial growth factor (VEGF) in diabetic
retinopathy. Cell Physiol Biochem. 40:1570–1577. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Sasore T and Kennedy B: Deciphering
combinations of PI3K/AKT/mTOR pathway drugs augmenting
anti-angiogenic efficacy in vivo. PLoS One. 9:e1052802014.
View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Yu Z, Zhang T, Gong C, Sheng Y, Lu B, Zhou
L, Ji L and Wang Z: Erianin inhibits high glucose-induced retinal
angiogenesis via blocking ERK1/2-regulated HIF-1α-VEGF/VEGFR2
signaling pathway. Sci Rep. 6:343062016. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Zhang ZZ, Qin XH and Zhang J: MicroRNA-183
inhibition exerts suppressive effects on diabetic retinopathy by
inactivating BTG1-mediated PI3K/Akt/VEGF signaling pathway. Am J
Physiol Endocrinol Metab. 316:E1050–E1060. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Zhang Y, Wang L, Zhang Y, Wang M, Sun Q,
Xia F, Wang R and Liu L: Nogo-B Promotes angiogenesis in
proliferative diabetic retinopathy via VEGF/PI3K/Akt pathway in an
autocrine manner. Cell Physiol Biochem. 43:1742–1754. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Zeng J, Zhao H and Chen B: DJ-1/PARK7
inhibits high glucose-induced oxidative stress to prevent retinal
pericyte apoptosis via the PI3K/AKT/mTOR signaling pathway. Exp Eye
Res. 189:1078302019. View Article : Google Scholar : PubMed/NCBI
|