Introduction
Cl− and HCO3− are
important anions in the body and they are involved in cellular
processes associated with various physiological functions and
disease development, including cell cycle, proliferation,
differentiation, membrane potential, reactive oxygen species (ROS)
levels, and intracellular pH (pHi) and extracellular pH (pHo)
regulation (1,2). HCO3− is the
second most abundant anion in bodily fluids after Cl−
and has an important role in human physiology (3,4).
Cl− regulates the functions of different organelles,
including endosomes, phagosomes, lysosomes, endoplasmic reticulum,
and mitochondria (1). In addition
to its main role as a pHi regulator, HCO3−
controls the activity and stability of dissolved proteins in bodily
fluids, such as saliva, pancreatic juice, intestinal juice and
airway surface fluid. HCO3− exists in balance
with CO2 produced by mammalian cell metabolism, and they
are interconverted in a pH-dependent manner to compose the main
buffer system that regulates human pH (4). The Cl− and
HCO3− levels in the cells are regulated by
ion channels and exchangers, including cystic fibrosis
transmembrane conductance regulator (CFTR), bicarbonate transport
proteins, and members of the solute carrier (SLC) 4 and SLC26
families. The SLC4 family consists of 10 transmembrane proteins,
including two electrogenic
Na+-HCO3− cotransporters [SLC4A4
(NBCe1) and SLC4A5 (NBCe2)], two electroneutral [SLC4A7 (NBCn1) and
SLC4A10 (NBCn2)], three electroneutral anion exchangers [AEs;
SLC4A1 (AE1), SLC4A2 (AE2), SLC4A3 (AE3)], the
Na+-dependent Cl−/HCO3−
exchanger SLC4A8 (NDCBE), and two other uncommon members [SLC4A9
(AE4) and SLC4A11 (BTR1] (5). Of
these, AEs include AE1-3, which are 53–56% identical at the amino
acid level. AE1 is expressed in the kidney and the erythrocyte
membrane (6), AE2 is widely
distributed in tissues and organs, and AE3 is expressed in the
central nervous system (7) and
heart specimens (8). The expression
distribution of AE members is listed in Table I. The majority of AEs are located
basolaterally in polarized cells. Under physiological conditions,
all three AEs undergo Na+-independent,
electron-neutralized Cl−/HCO3−
exchange (5,9,10).
Numerous studies have shown that AE2 is involved in a variety of
physiological and pathophysiological processes. For example, in
ameloblasts, AE2 mediates the efflux of HCO3−
across the basement membrane, supports H+ secretion
across the apical membrane, and contributes to tooth remodeling
(11). In B lymphoid neoplasms, AE2
can be used as a target for specific peptide-targeted therapy
(12). In human nucleated cells,
AE2 mediates the transport of abscisic acid, which is a hormone
involved in inflammatory response and glycemic control (13). In osteoclasts, AE2-mediated efflux
of HCO3− across the basement membrane serves
a key role in bone resorption activity, suggesting that the loss of
AE2 function shifts bone balance toward ossification or
pathological high bone density (14). Loss of AE2 function in AE2 (−/-)
mice is lethal to embryos, as it leads to emaciation, hypodontia or
edentulism and severe growth retardation, and most of them die
during weaning (15). Recent
studies have found that AE2 is expressed in most organs of the
digestive system and has an important role in the physiological
processes of the digestive system and digestive system diseases,
such as gastric cancer (16) and
primary biliary cholangitis (PBC) (17). However, the mechanism and function
of AE2 in the digestive system, particularly in certain digestive
system diseases and tumors, have not been fully elucidated. The aim
of the present review was to describe the role of AE2 in the
physiology and pathophysiology of the digestive system and the
current use of AE2-based treatments.
 | Table I.Organizational characteristics and
expression of human AEs. |
Table I.
Organizational characteristics and
expression of human AEs.
| Protein name | Alias | Chromosome
localization | Gene length
(kb) | Tissue
distribution |
|---|
| AE1 | SLC4A1 | 17q21-q22 | ~20 | Erythrocyte
membrane, kidney |
| AE2 | SLC4A2 | 7q36.1 | ~17 | Salivary gland,
stomach, duodenal enterocytes, proximal colon, epididymal
epithelium, ameloblasts, bile duct, hepatocyte, pancreas, airway,
kidney |
| AE3 | SLC4A3 | 2q36 | ~14 | Central nervous
system, heart |
Structural features of AE2
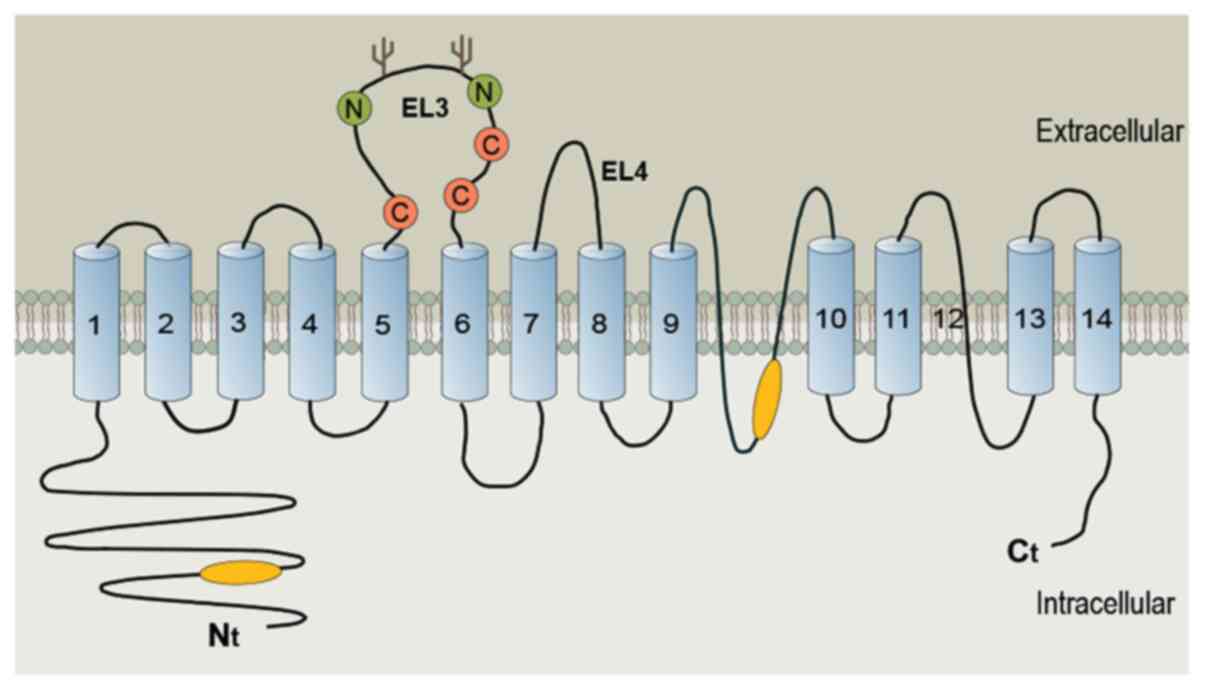
AE2 is a transmembrane bidirectional transporter
that forms a homodimer in the membrane. AE2 has a two-domain
structure, including an N-terminal cytosolic domain and a
C-terminal membrane domain, consisting of 14 transmembrane segments
(18,19). The human AE2 gene contains three
alternative promoters, resulting in at least three different
transcripts (AE2a, b and c); the human AE2a protein contains 1,240
residues with a molecular weight of 137 kDa, while the mouse AE2
gene encodes five N-terminal variant polypeptides (20). The N-terminal cytosolic domain and
putative re-entrant loop 1 of the transmembrane domain of AE2 also
contain structural determinants of pH sensitivity (Fig. 1) (21–23).
AE2 has a carbonic anhydrase (CA) II-binding site in its C-terminal
tail that can localize the enzyme to the basolateral membrane
(BLM). The activity of AE2 is stimulated by both alkaline pH and
hyperosmosis through a conserved motif in the N-terminal
cytoplasmic domain, although the membrane domain also plays a role
in AE2 anion permeability (20).
AE2 and the esophagus
In the esophagus, AE2 localizes to the BLM and
exports one HCO3− for every Cl−
that is imported. Therefore, Cl− is transported across
cells from the basal side into the lumen and is involved in
esophageal fluid secretion (24,25).
The esophagus is a muscular tube that connects the
hypopharynx to the gastric cardia; its main function is delivering
food to the stomach. The ingested food enters the esophagus after a
short stay in the oral cavity, so the upper part of the esophagus
is exposed to the temperature, pH and osmotic pressure of the
ingested food, while the lower end of the esophagus can be corroded
by reflux of gastric contents (26). The most important defense mechanism
of esophageal epithelial cells against reflux-induced injury is
esophageal epithelial resistance consisting of functional and
structural components including the following: i) A surface mucus
and an unstirred aqueous layer containing
HCO3−, which provides an alkaline
environment; ii) cellular junctions (tight junctions) of the apical
surface and BLM that prevent diffusion of H+ into the
intercellular space and cell, respectively; and iii) intracellular
buffer systems, such as HCO3− or phosphate
buffer systems (26). Among them,
the secretion of HCO3− in the lumen is
considered to protect the esophagus by neutralizing acidic reflux.
Secretion of esophageal HCO3− is associated
with the esophageal submucosal glands (SMG) (27). In the SMG, CFTR and SLC26A6 on the
apical membrane and BLM, respectively, are mainly involved in
HCO3− secretion, and apical Cl−
transport in the SMG can be linked to the CFTR Cl−
channel (Fig. 2) (27). Transcellular Cl−
transport, on the other hand, is particularly important in SMG, as
fluid secretion from SMG is more significant. Transcellular
Cl− transport is mediated by AE2 localized at the
basolateral side of the cell and, at the same time, AE2,
independently of Na+, mediates the entry of one
Cl− into the cell with exocytosis of one
HCO3−, which acidifies the cell and prevents
excessive alkalinization (24,25).
In combination, these findings suggest that the role of AE2 in the
esophagus may be to protect the esophageal mucosa by mediating
Cl− transcellular secretion into the lumen in the basal
layer, further causing fluid secretion. Therefore, AE2 may serve as
a therapeutic target for reflux esophagitis.
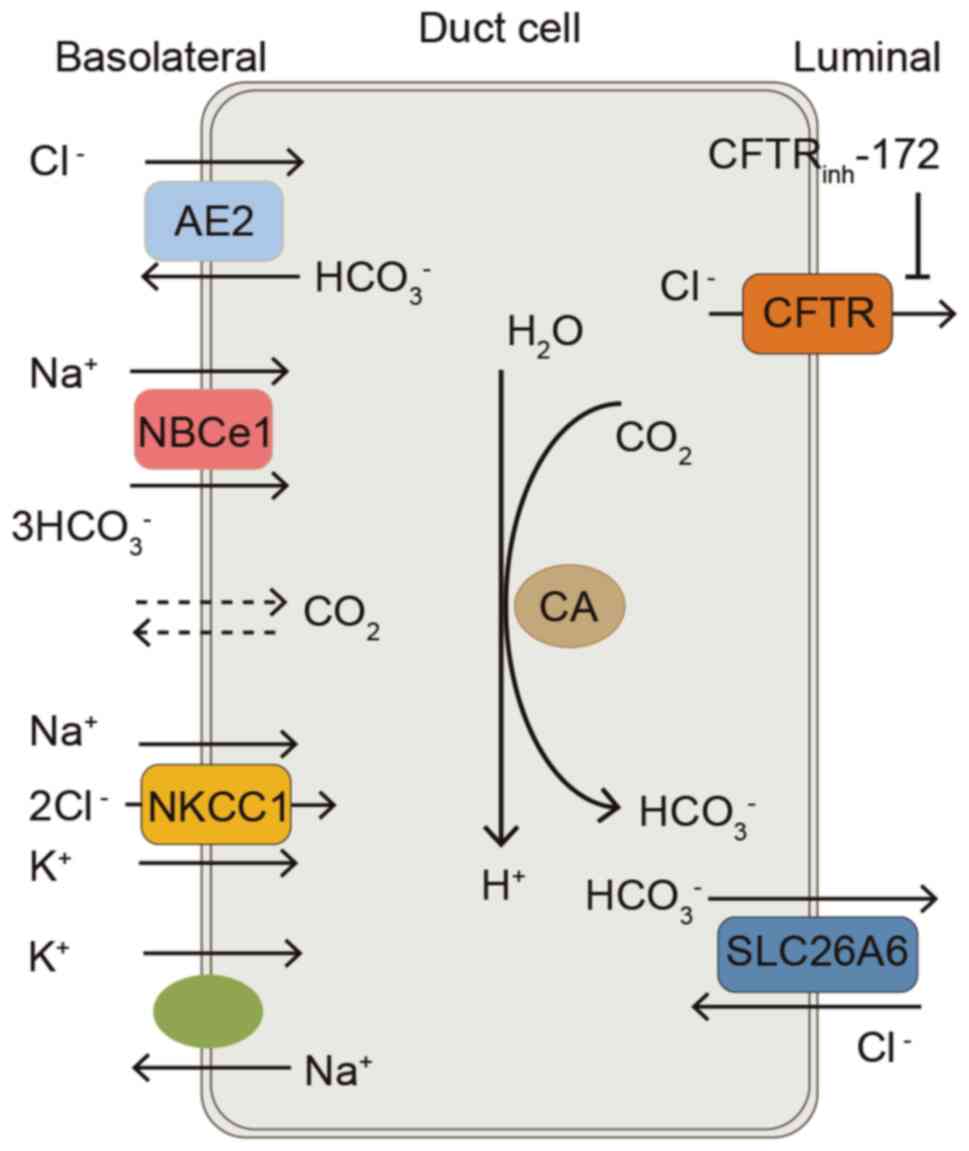 | Figure 2.Model of a duct cell illustrating ion
transport mechanisms involved in Cl− and
HCO3− transport in the esophageal submucosal
gland. At the basolateral membrane, HCO3−
entry is mediated by the NBCe1. AE2 mediates Cl− uptake
by the cell. NKCC1 mediates Na+, K+ and
Cl− entry into the cell. At the apical membrane,
HCO3− efflux is mediated by
Cl−/HCO3− exchanger SLC26A6.
Apical CFTR may be permeable to HCO3− or
Cl− efflux. Cl− efflux drives apical SLC26A6,
leading to HCO3− secretion. AE2, anion
exchanger 2; NBCe1, Na+-HCO3−
cotransporter; NKCC1, Na+-K+−2Cl−
cotransporter 1; CA, carbonic anhydrase; CFTR, cystic fibrosis
transmembrane conductance regulator; CFTRinh-172, glibenclamide,
cystic fibrosis transmembrane conductance regulator inhibitor;
SLC26A6, solute carrier family 26 member 6. |
AE2 also has a role in the pathogenesis of
esophageal cancer (28). In
esophageal squamous cell carcinoma (ESCC), AE2 has been
demonstrated to be upregulated and mainly located in the cell
membrane or the cytoplasm of esophageal cancer cells. AE2 knockdown
decreased apoptosis, and increased migration and invasion of ESCC
cells through regulation of matrix metalloproteinase expression
(28). The altered AE2 expression
may affect tumorigenesis by altering the pHi. Decreased AE2
expression facilitates intracellular alkalinization, which promotes
cancer cell metabolism (29).
Clinically, by analyzing the clinicopathological characteristics of
61 patients with ESCC, it was found that the expression levels of
AE2 in the whole tumor were not significantly associated with the
overall 5-year survival rate, but the reduced expression of AE2 at
the invasive front was correlated with poor prognosis (28); this finding requires further
study.
AE2 and the stomach
The stomach serves as a center for the mechanical
distribution of food and contains digestive glands; some of the
chemical processes of digestion also occur in the stomach. Gastric
mucosa secretes HCO3− with mucus, which
creates a pH gradient in the mucus of surface cells and, although
the lumen is acidic, the pH value near the surface cells is almost
neutral. This ‘mucus-bicarbonate diffusion barrier’ is an important
part of protection against gastric acid damage (30). AE2 serves an important role as a
Cl−/HCO3− exchanger. First, AE2 is
localized at the basal side (31).
Second, studies have shown that rabbit parietal cells and mucous
cells derived from the same gastric stem cell population exhibited
completely different AE2 isoform expression patterns (32). The isoform expression pattern in
parietal cells is AE2b>>AE2c>or=AE2a, and the isoform
expression pattern in mucous cells is AE2a>AE2b>>AE2c
(32). Parietal cells are activated
by secreting H+ into the lumen, inducing pHi increase
and intracellular alkalinization (31). AE2, which is localized in the BLM
and acts as an output of HCO3−, is activated
and pumps HCO3− to the basolateral side,
which can promote cell recovery from alkaline pH, reabsorb
HCO3− into the blood, avoid excessive
alkalinization of cells and further provide Cl− into the
lumen by pumping Cl− into cells. The activity of AE2 is
stimulated by both alkaline pH and hyperosmosis, with the
hypertonic activation of AE2 being secondary to the hypertonic
activation of Na+/H+ exchange (31). AE2-mediated anion exchange is also
stimulated by ammonium and hyperosmolarity, and the mechanism
involves intracellular Ca2+ chelation and
calmidazolium-sensitive inhibition of anion exchange (33). Both gastric acid secretion and
osmolality in the gastric cavity are increased following the
ingestion of food, which indicates that AE2 serves a vital role in
assisting parietal cells to excrete HCO3− and
supply the gastric lumen with Cl−. AE2 is essential for
the secretion of acid in acid-secreting parietal cells.
Accordingly, a previous study identified gastric acid deficiency,
moderate dilatation of the gastric glandular lumen and a reduced
number of parietal cells in AE2 (−/-) mice (15). Ultrastructural analysis revealed
abnormal parietal cell architecture, severely impaired secretory
tubule development and few tubular vesicles, but normal apical
microvilli, indicating that normal AE2 function is required in
mouse parietal cells, particularly for normal gastric acid
secretion and normal development of secretory tubules and
canalicular vesicle membranes (15). In humans, the complete loss of AE2
function may be lethal to embryos (15). Of note, in AE2 (a and b) (−/-) mice,
basal acid secretion was normal and no parietal cells were
hypoplastic, whereas carbachol/histamine-stimulated acid secretion
was impaired by 70%. These results indicated the key role of the
AE2 a and b isoforms in gastric acid secretion. Gastric expression
of the residual AE2c isoform was low and mice deficient in AE2c did
not exhibit impaired acid-stimulated secretion (34). Further studies are required to
explain the differences between the physiological and
pathophysiological functions.
During the stimulation of gastric acid secretion,
parietal cells alkalinize the mucosal interstitial fluid. The
expression of AE2 variants with overlapping pHo sensitivities in
the parietal cell BLM serves to extend the range of pHo. Under this
range, pHo regulates parietal cell basolateral
Cl−/HCO3− exchange, while allowing
other mechanisms of pHi homeostasis (35). Maintaining the pHi in the optimal
range is essential for cell metabolism and survival. In addition,
secretion leads to the decrease in cell volume which, in turn,
leads to further changes in volume regulation to stabilize the
shape of the cell, subcellular structure and concentration of
cytoplasmic components, involving Na+/H+
exchange and Cl−/HCO3− exchange,
Na+/HCO3− cotransport and
Na+/K+/2Cl− cotransport (NKCC)
(30).
AE2 may have a role in cell proliferation and
apoptosis. AE2 increases intracellular chloride and mediates high
glucose-induced apoptosis of umbilical vein endothelial cells in a
time- and concentration-dependent manner through the mitochondrial
permeability transition pore/ROS/caspase-3-dependent pathway
(36). Studies on wound repair in
rat gastric epithelial cells have shown that
4.4′-diisothiocyano-2,2′-stilbene disulfonic acid (DIDS), or the
removal of Na+, Cl− and/or
HCO3−, inhibit the key processes involved in
wound healing, such as cell migration. The transport processes
responsible for these phenomena remain unclear. However, NBCe1 and
AE2 transcripts are detected in these cells.
In conclusion, AE2 has an important physiological
function in the stomach, and the abnormality of AE2 may also lead
to the occurrence of tumors. AE2 was found to be downregulated in
gastric cancer cells (16), and its
downregulation was correlated with poor differentiation and
prognosis of gastric cancer (37).
The lower expression of AE2 in gastric cancer was partly due to the
promotion of ubiquitin-mediated rapid degradation in the presence
of p16, whereas gastrin inhibited the growth of gastric cancer
cells, at least partly by upregulating AE2 (37). The potential underlying mechanism is
the stimulation of AE2 expression in gastric cancer cells through
the transcription factor early growth response 1 (EGR1) in a
cholecystokinin B receptor-dependent manner (38). It was previously reported that the
combination of trastuzumab and gastrin exerted a synergistic
inhibitory effect on human epidermal growth factor receptor
2-negative gastric cancer cells by targeting cytoplasmic AE1 and
p16 inhibition, and that AE2 expression, which was originally
decreased in gastric cancer tissues, was upregulated during
combination therapy (39). It can
be concluded that AE2 participates in the occurrence and
development of gastric cancer, suggesting that AE2 may be a
potential target for the treatment of gastric cancer.
AE2 and the intestine
AE2 and the small intestine
AE2 protein is expressed in the small intestine and
colon, and is mainly localized in the BLM, exchanging
HCO3− and Cl− in an electroneutral
manner. It is involved in physiological regulatory mechanisms, such
as HCO3− secretion and Cl−
excretion. Ion secretion in the duodenum contributes to the
formation of a liquid, buffered intraluminal environment with an
optimized pH, protects the intestinal epithelium from gastric acid
and promotes digestion in the proximal intestine (40). In the intestine, the osmotic
gradient that pulls water into the intestine is mainly produced by
Cl− and, to a lesser extent by
HCO3− secretion, with Na+
passively following from the paracellular space (41). On the apical membrane,
Cl− is secreted into the intestinal lumen through three
pathways, CFTR, calcium-activated Cl− channel and the
voltage-gated chloride channel protein 2, which further produces a
driving osmotic gradient for fluid secretion (41). In the basolateral membrane, the
ability of the intestine to secrete fluid depends on the NKCC
cotransporter which mediates Cl− uptake at the
basolateral side of enterocytes, thus providing a substrate for
apical Cl− secretion (42). However, no significant
manifestations of constipation were observed in patients treated
with ring diuretics compared with the placebo group in a previous
study (43), indicating that an
alternative pathway for Cl− uptake by enterocytes
exists. AE2 may be a protein that assists Cl− uptake in
the mouse duodenum (44).
Similarly, the phenomenon of AE2 acting as an assistant
Cl− uptake channel has already been observed in
submandibular acinar cells involved in salivary gland fluid
secretion (45). The duodenal
mucosa senses luminal acidity through epithelial ion transporters
and neuronal acid receptors, further increasing the absorption of
luminal acid and HCO3− secretion, thereby
maintaining the acid-base balance between the stomach and the
duodenum (46). Chloride uptake via
AE2 is coupled to basolateral NaHCO3 cotransport to
support CFTR-mediated Cl− and
HCO3− secretion (44). Both endogenous prostaglandin and NO
are involved in the local regulation of acid-induced duodenal
HCO3− secretion; NO stimulates
HCO3− secretion by increasing prostaglandin
production, and prostaglandin E2 stimulates
HCO3− secretion by activating prostaglandin
receptor subtypes EP3 and EP4 (47). In addition, estrogen can regulate
HCO3− secretion in the duodenal mucosa of
mice (48), but whether these
regulatory factors regulate AE2 expression and function requires
further study.
AE2 and the colon
The colon is responsible for regulating the
electrolyte and water content of stool. In the human intestine, AE2
and bAE3 (the brain subtype) are expressed throughout the
intestine, and both localize basolaterally in epithelial cells.
Their expression is higher in the colon compared with the ileum and
jejunum (49). The expression of
AE2 and bAE3 in the mouse intestine is the same as that in the
human intestine (50). AE2
polypeptides were found to be more abundant in colonic surface
cells compared with crypt cells, whereas ileal crypts and villi
exhibited a similar abundance of AE2 (50). AE2 has also been observed in the
mural and vascular smooth muscle of the murine intestine (50). As previously reported, the osmotic
gradient for the secretion of fluid into the intestine is mainly
produced by Cl− on the apical surface of epithelial
cells and to a lesser extent by HCO3−
secretion, with Na+ passively following through the
paracellular space (41). In the
apical membrane, the three routes of HCO3−
extrusion into the lumen are the
Cl−/HCO3− exchanger SLC26A3, CFTR
and the short chain fatty acid/HCO3−
exchanger (51). On the basement
membrane, HCO3− is transported into cells
through Na+/H+ exchange (SLC family 9 member
A1), as well as carbonic anhydrase and
Na+/HCO3− cotransporter (51). NKCC1 takes up Cl− at the
BLM (51). There is a
carrier-mediated electroneutral
Cl−/HCO3− exchange in the BLM,
which can function as a Cl−/OH− exchanger in
the absence of HCO3− and modulate pHi in the
BLM of the rat distal colon (52).
A study on intestinal organoids showed that the CFTR knockout crypt
epithelium maintains an alkaline pHi due to loss of Cl−
and HCO3− efflux, which impairs pHi
regulation by AE2 (44). In the AE2
(−/-) colon, basolateral NKCC1-supported Cl− secretion
was increased, whereas HCO3− secretion was
reduced (53). Spinophilin and CA
XII enhanced the Cl−/HCO3−
exchange activity of AE2. Spinophilin bound to AE2 and
significantly increased its anion exchange activity; in particular,
the spinophilin 1–480 domain was required to enhance AE2 activity
(54). The BLM-associated CA
isoform CAXII colocalized with AE2 to the plasma membrane and
significantly increased the activity of AE2 (54). Activators of Cl−
secretion include growth hormone, neuropeptides, opioids,
norepinephrine and autocrine survival factors (55).
AE2 expression is increased in colon tumor tissue
compared with adjacent non-cancerous tissue. In addition, the
inhibition of AE2 expression in colorectal cancer cells has been
found to decrease cell proliferation, indicating that AE2 may have
a role in promoting colorectal cancer growth (56). Increased metabolism accelerates
local acid production in cancer tissue. In one study, it was found
that acid uptake was inhibited in colorectal cancer cells, whereas
cellular acid uptake increased together with AE2 expression upon
TGF-β1 stimulation (57). However,
no acid transmission was observed between colorectal cancer cells
with or without TGF-β1 treatment (57). Whether AE2 regulates pH in tumor
tissue by assisting in the expulsion of
HCO3−, and thus affecting the biological
behavior of tumors, requires further study. Clinically, in a
survival analysis of 57 patients with colon malignancies, AE2
expression was associated with poor prognosis (56). In colon cancer, AE2 expression
promoted the proliferation of colon cancer cells, AE2
overexpression was correlated with Ki67 expression (a nuclear
proliferation marker) and gastrin inhibited the proliferation of
colon cancer cells by inhibiting the expression of EGR1 and AE2 and
blocking extracellular signal-regulated kinase phosphorylation
(56). The mechanism of AE2
function in colon cancer and its clinical significance require
further study.
AE2 and the pancreas
The pancreas is a complex endocrine and exocrine
organ; the exocrine pancreas is composed of acinar and ductal
cells. AE2 is expressed in the BLM of acinar cells in the pancreas
(58). It is also expressed in the
pancreatic duct cells (59,60), but in low quantities (33). Acinar cells initially secrete an
isotonic Cl−-rich fluid. Subsequently, pancreatic ductal
cells alter the ionic composition of pancreatic juice and secrete
large amounts of pancreatic juice and HCO3−.
In humans, under conditions of pH 7.4 and 5% CO2, the
HCO3− equilibrium concentration is ~25 mM,
and the HCO3− concentration in stimulated
pancreatic juice can reach 140 mM (4), which is necessary for normal
digestion. Since AE2 is minimally expressed in the duct, it is
reasonable to hypothesize that the function of AE2 in the duct is
very limited. Of note, previous data suggest that the chloride
channel CFTR in pancreatic ductal cells may switch to the
HCO3−-conducting channel by activating
with-no-lysine kinase 1 and the downstream kinases oxidative
stress-responsive kinase1 and sterile 20/SPS1-related
proline/alanine-rich kinase (61).
The increase of CFTR HCO3−/Cl−
permeability from 0.4 to 1.0 has little effect on the secreted
HCO3− concentration or volume flow. However,
the model showed that the ~80% reduction in basolateral AE2
activity was essential for minimizing the intracellular
Cl− concentration following cyclic adenosine
monophosphate (cAMP) stimulation, and thus maximized the secreted
HCO3− concentration (62). Conversely, in the pancreas, whether
AE2 is involved in HCO3− secretion and
whether its regulation occurs in a similar manner has not yet been
demonstrated. However, it has been experimentally shown that the
knockdown of AE2 and CAXII inhibits AE2 activity and fluid
secretion in the pancreatic and salivary ducts (63). Furthermore, the exchange activity of
AE2 increased significantly following binding to spinophilin. In
particular, the spinophilin1-480 domain was required to enhance AE2
activity. The BLM-associated CA isoform, CAXII, was found to
colocalize with AE2 in the plasma membrane and to significantly
increase the activity of AE2 (54).
In conclusion, studies on the AE family's role in the process of
Cl−/HCO3− exchange in the pancreas
are limited, and further research is required.
AE2 and the liver
AE2 is located in the extrahepatic bile duct and
cholangiocytes (64), and its
distribution is different from that of the aforementioned digestive
organs. Subapical (65) or apical
distribution (66) of AE2 has been
observed in hepatobiliary epithelial cells. In addition, both AE2
and CFTR coexisted with water channel protein aquaporin 1 (AQP1) in
the vesicles of cholangiocytes, which were concentrated in the
apical membrane of cholangiocytes stimulated by cAMP and secretin
(65). AE2 mRNA signals were
detectable in the cytoplasm of some hepatocytes (mainly periportal)
(67). Even at the apical membrane,
the direction of AE2-driven ion transport was found to remain
unchanged, with AE2 still pumping in one Cl− and pumping
out one HCO3−, demonstrating that AE2 can
participate in the secretion of HCO3−.
Collectively, these results demonstrated that AE2 is involved in
the regulation of pHi homeostasis and secretin-stimulated biliary
HCO3− secretion. Biliary
HCO3− is largely dependent on apical
HCO3− release. The main exchangers involved
in Cl−/HCO3− exchange in apical
bile duct membranes are AE2, cAMP-regulated Cl− channels
and Ca2+-regulated Cl− channels (68). AE2 contributes to the creation of an
alkaline protective environment in the bile duct lumen through
Cl−/HCO3− exchange at the apical
membrane and HCO3− secretion (69), thus protecting the liver from bile
acid attack (5). An intact
glycocalyx is present on the apical membrane of cholangiocytes, and
the intracellular HCO3− sensor soluble
adenylyl cyclase (sAC) and functional AE2 are crucial for
maintaining biliary HCO3− concentration and
protecting cholangiocytes from toxic bile acid-induced apoptosis
(70–72). The bile duct requires biliary
cellular HCO3− secretion to maintain the
ionization of toxic bile acids and, thus, make them
membrane-impermeable (68,70). Therefore, the expression of AE2
affects bile acid uptake in immortalized non-malignant human
intrahepatic cholangial and human cholangiocarcinoma cells. In
addition, the bile acid uptake rate and toxicity are high in the
presence of decreased AE2 expression in those cells (73). AE2 internalization in hepatocytes
may result in decreased tubular HCO3− output
and decreased bile flow (73). In
addition, AE2 is responsible for maintaining pHi in cholangiocytes
through Cl−/HCO3− exchange
(69). Mouse cholangiocytes express
NBCe1, which may compensate for the lack of AE2 (74). As illustrated in Fig. 3, AE2 is known to be regulated by
glucagon, secretin, and the microRNA (miR)-506 which is located on
the X chromosome (75,76). The target organs of glucagon and
secretin are hepatocytes and cholangiocytes, respectively. These
two hormones appear to be quite similar and to interact with their
specific G protein-coupled receptors, causing an increase in the
intracellular cAMP level and activating cAMP-dependent
Cl− and HCO3− secretion. Both
hepatocytes and cholangiocytes appear to have cAMP-responsive
intracellular vesicles (76).
Glucagon may induce pericanalicular vesicles which contain AE2 and
the water channel AQP8 and the glutathione carrier ATP binding
cassette subfamily C member 2 to the canalicular membrane in
hepatocytes, leading to canalicular hypersecretion of
HCO3−-rich bile; secretin may also stimulate
ductal HCO3− secretion by interacting with
secretin receptors that activate cAMP/CFTR/AE2 signaling, which is
increased by biliary hyperplasia (77). The proinflammatory cytokines
interleukin (IL)-8, IL-12, IL-17, IL-18 and tumor necrosis factor
(TNF)-α enhance the expression of miR-506 in the biliary
epithelium, which inhibits AE2 expression by directly targeting AE2
mRNA, and sensitizes cholangiocytes to bile salt-induced apoptosis
(BSIA) (78). In addition, miR-506
is located on the X chromosome (79), which explains the fact that PBC
mainly affects women and emphasizes the role of the X chromosome in
PBC. Increased AE2 protein expression and activity are the main
reasons for the increase in rifampicin-induced bile flow (80). AE2 knockdown sensitized immortalized
H69 human cholangial cells to not only etoposide-induced apoptosis,
but also BSIA. Mechanistically, AE2 downregulation mediates
apoptosis through the activation of sAC and then through the
intrinsic apoptotic pathway, since sAC is an evolutionarily
conserved HCO3− sensor that regulates
apoptosis, barrier function and TNF signaling. In addition, sAC
inhibition prevents Bax phosphorylation at Thr167, as well as Bax
translocation into mitochondria and cytochrome c release; these
molecules are involved in endogenous apoptosis during BSIA in
cholangiocytes (71). This process
depends on intracellular Ca2+ storage (71). Insufficient AE2 function in
lymphocytes may interfere with pHi regulation in cells and alter
immune homeostasis, leading to autoimmunity (81). On the other hand, as aforementioned
(71), AE2 downregulation
sensitized immortalized human cholangiocytes to BSIA which,
together with alterations in immune homeostasis, may favor the
production of antimitochondrial antibodies and autoimmune attack in
the bile duct (71).
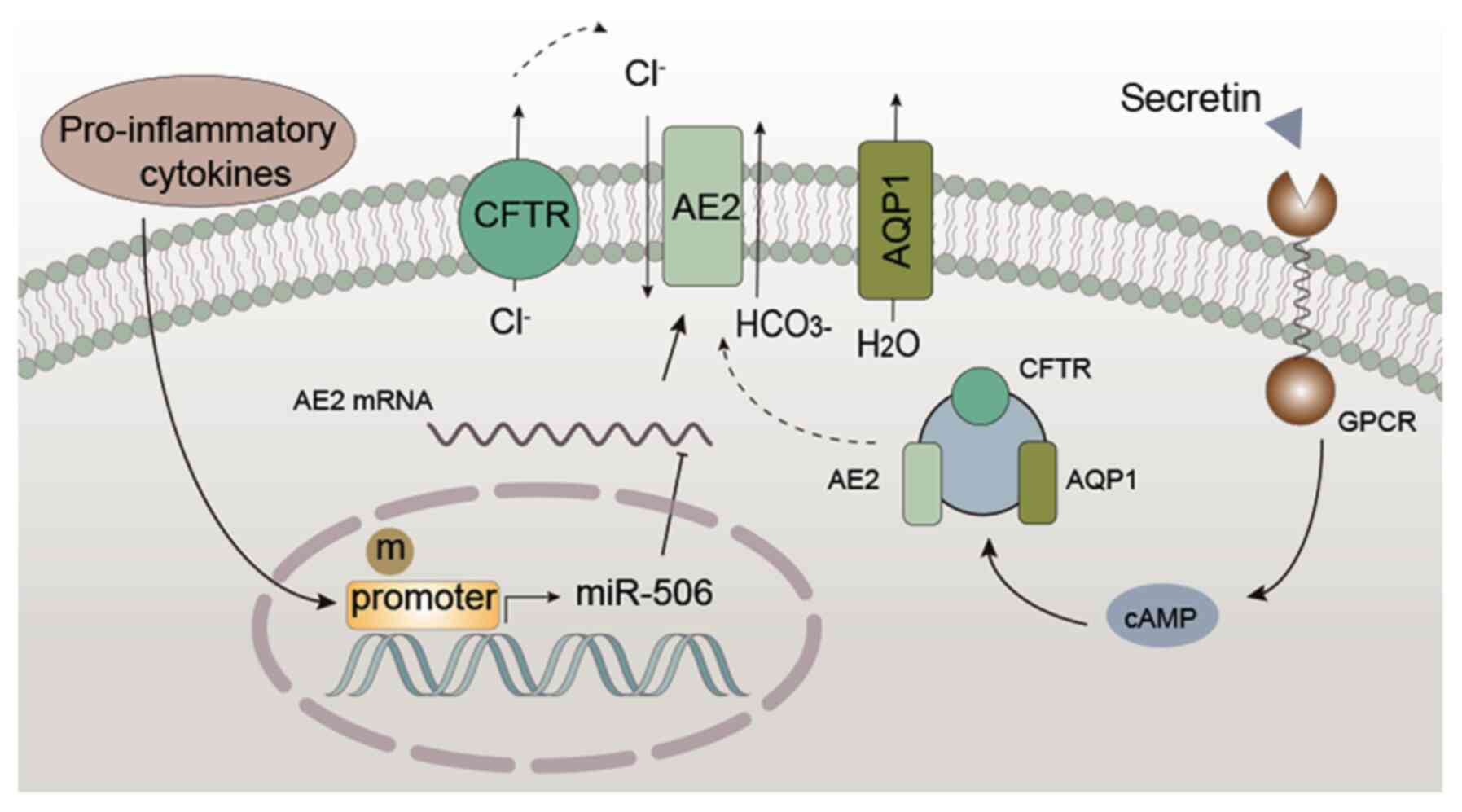 | Figure 3.Regulatory mechanism of AE2 in
cholangiocytes. Pro-inflammatory cytokines (IL-8, IL-12, IL-17,
IL-18 and TNF-α) enhance the expression of miR-506, which directly
targets AE2, thereby inhibiting the expression of AE2. In addition,
the hypermethylation of the AE2 promoter region inhibits AE2
expression. Under physiological conditions, secretin binds to
receptors and promotes cAMP/protein kinase C-dependent exocytosis
of vesicles containing CFTR, AE2 and AQP1 in the apical membrane of
cells. This results in the secretion of Cl− via CFTR and
further exchange with HCO3− via AE2, creating
a luminal osmotic gradient for AQP1 to move water, leading to
biliary secretion. AE2, anion exchanger 2; miR, microRNA; cAMP,
cyclic adenosine monophosphate; CFTR, cystic fibrosis transmembrane
conductance regulator; AQP1, aquaporin 1; GPCR, G protein-coupled
receptor. |
AE2 and primary biliary
cholangitis
PBC and liver cancer are the main liver diseases
associated with AE2. Biliary atresia-specific induced pluripotent
stem cells were found to exhibit reduced AE2 expression (82). Similarly, in patients with PBC, the
expression of AE2 in liver biopsies was significantly reduced
compared with healthy controls (83), and AE2 activity was also decreased
(84). Decreased AE2 expression is
associated with dysregulated autophagy, abnormal pyruvate
dehydrogenase complex, E2 component expression, cellular
senescence, as well as hypermethylation of the AE2 promoter region
in PBC bile duct lesions, followed by the insufficiency of
secretin-stimulated bile bicarbonate secretion (17,83).
In addition, secondary inflammatory and pro-inflammatory
cytokine-mediated injury could lead to a decrease in AE2-mediated
bile secretion (85). AE2
deficiency of PBC immunocytes may have an essential role in
autoimmune phenomena (86,87). Furthermore, AE2 downregulation was
found to sensitize cholangiocytes to apoptotic damage (71). Consistently, AE2 (a and b) (−/-)
mice displayed PBC-like features, including extensive portal
inflammation, bile duct injury and infiltration by surrounding
CD4+ and CD8+ T lymphocytes, increased
oxidative stress in cholangiocytes, liver fibrosis, increased
production of interferon-c and IL-12, increased serum IgM, IgG and
liver alkaline phosphatase levels, and serum detectable
antimitochondrial antibodies (88).
Among them, CD8+ T cells were found to rely on AE2 to
maintain pHi at physiological levels (89). Therefore, AE2 (a and b) (−/-) mice
exhibited a higher number of intrahepatic cytotoxic CD8+
T cells, which inhibited programmed cell death-1 (PD-1) expression,
accompanied by a reduction in apoptosis, thus favoring chronic
immune-mediated cholangitis (87).
Early in the life of mice, AE2 deficiency leads to intrahepatic
T-cell activation and PD-1/programmed cell death ligand 1-mediated
deletion. With age, intrahepatic CD8+ T cells
epigenetically inhibit PD-1, and their corresponding expansion and
further activation favor autoimmune cholangitis (87). In conclusion, the possible mechanism
through which AE2 mediates PBC is that, in PBC, various factors,
such as glucagon, insufficient secretion of secretin and
pro-inflammatory cytokines (IL-8, IL-12, IL-17, IL-18 and TNF-α),
lead to a reduced expression and/or function of AE2. This leads to
impaired bile HCO3− secretion, followed by
bile duct cell injury and cholestasis, as well as destruction of
cholangiocyte autoimmune response and increased sensitivity to
apoptotic injury, finally resulting in bile duct injury (Fig. 4). In a clinical study of 258
patients with PBC, and two independent groups of 286 and 269
healthy controls, AE2 variants were shown to be an independent
prognostic factor for PBC by multivariate Cox regression analysis
that included clinical and biochemical parameters (90); the progression of liver disease
under ursodeoxycholic acid (UDCA) treatment was found to be
significantly associated with single-nucleotide polymorphisms in
the TNF-α and AE2 genes (90). UDCA
can upregulate the expression of bile salt export pump, but not
that of AE2 (91). UDCA is
conjugated to promote secretin-stimulated hydrocholeresis through
AE2, intracellular Ca2+, microtubules, protein kinase
C-α, PI3K, protein kinase A and mitogen-activated protein kinase
kinase (92). Similarly, the
secretion of carbonate-rich bile was significantly enhanced in rats
treated with UDCA (92). These data
strongly suggest that AE2 may serve as a novel target for the
diagnosis and treatment of PBC.
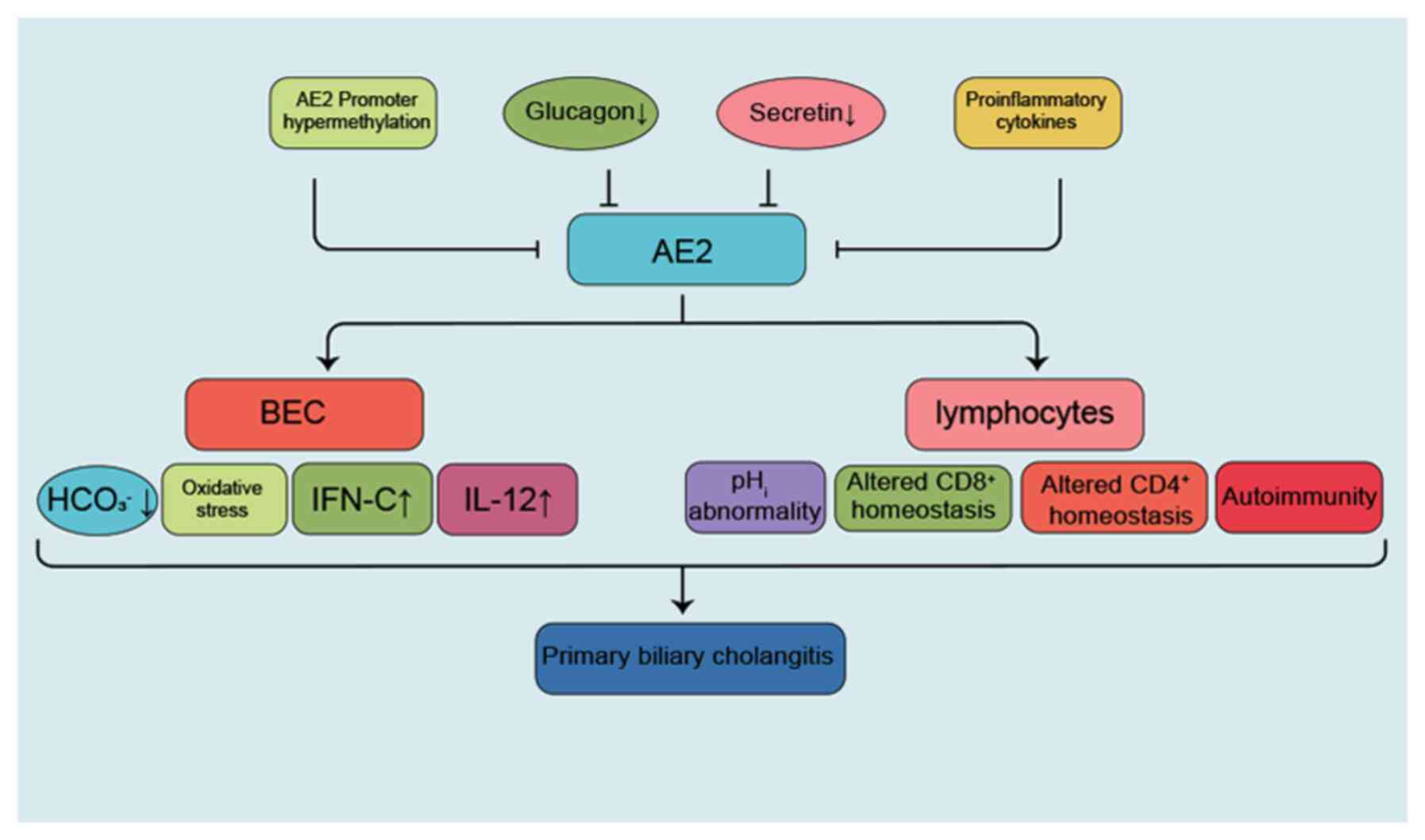 | Figure 4.AE2-associated pathogenesis of PBC.
In PBC, various factors, such as insufficient secretion of glucagon
or secretin and pro-inflammatory cytokines (IL-8, IL-12, IL-17,
IL-18 and TNF-α), lead to reduced expression and/or function of
AE2, resulting in impaired bile HCO3−
secretion, followed by bile duct injury and cholestasis. Finally,
the destruction of cholangiocyte autoimmune response and its
sensitivity to apoptotic injury result in PBC. AE2, anion exchanger
2; PBC, primary biliary cholangitis; BEC, bile duct cell; IFN,
interferon. |
AE2 and liver cancer
AE2 was found to be overexpressed in hepatocellular
carcinoma (93,94). AE2 knockdown significantly reduced
the viability of poorly differentiated HA22T/VGH cells and arrested
the cell cycle in the sub-G1 phase. In addition, treatment with
inhibitor DIDS significantly inhibited cell proliferation and
induced apoptosis of the poorly differentiated HA22T/VGH cells
(94,95). It has been found that CA IX
interacts with AE2 and Na+-HCO3−
cotransporter (NBCe1) in lamellipodia and increases cell migration
through its ability to facilitate ion transport and pH control at
the protruding front of moving cells (96). Whether AE2 is involved in other
biological behaviors of liver cancer requires further study.
Conclusion
In conclusion, AE2 is involved in the development
and progression of digestive system diseases by regulating
intracellular and extracellular
Cl−/HCO3− exchange and further
regulating cellular function. The physiology and pathophysiology of
AE2 are summarized in Table II.
The role of AE2 in digestive system diseases, such as PBC, and
digestive tract tumors, is set to become a new research hotspot.
AE2 may become a novel molecular marker for the diagnosis and
treatment of digestive system diseases, and drug development for
AE2 may open up a new direction for the diagnosis and treatment of
digestive system diseases in the future.
| Table II.Physiology and pathophysiology
characteristics of AE2. |
Table II.
Physiology and pathophysiology
characteristics of AE2.
| Digestive
organ | Distribution | Physiological
function |
Pathophysiology |
|---|
| Esophagus | Basolateral
membrane | AE2 may further
cause fluid secretion | 1. In esophageal
squamous cell carcinoma, |
|
|
| by mediating
Cl− secretion into the | AE2 is
upregulated. |
|
|
| lumen across cells
in the basal layer. | 2. Low-grade
expression of AE2 at the |
|
|
|
|
invasive front is associated
with shorter |
|
|
|
|
postoperative survival (sample
size, 61). |
| Stomach | Basolateral
membrane | AE2 assists
parietal cells to expel | 1. In gastric
cancer, AE2 is downregulated. |
|
|
|
HCO3− and import
Cl−. | 2. AE2 expression
is associated with poor |
|
|
| Maintains pHi
within optimum range. |
prognosis. |
| Small | Basolateral
membrane | Bicarbonate
secretion and | Unknown |
| intestine |
| Cl−
excretion provide an appropriate |
|
|
|
| pH environment,
protect the intestinal |
|
|
|
| epithelium from
gastric acid, and |
|
|
|
| promote digestion
in the intestine. |
|
| Colon | Basolateral
membrane | 1. AE2 binding to
SLC9A1 constitutes | 1. In colorectal
cancer, AE2 is upregulated. |
|
|
| a minor
component of colonic Cl− | 2. AE2 expression
is associated with poor |
|
|
|
uptake. |
prognosis (sample size,
57). |
|
|
| 2. AE2 is involved
in regulating pHi in |
|
|
|
| the
basolateral membrane of the rat |
|
|
|
| distal
colon. |
|
| Pancreas | Basolateral
membrane | An ~80% reduction
in basolateral AE2 | Unknown |
|
| of acinar
cells. | activity is
obtained by maximizing the |
|
|
| Pancreatic duct
cells, | secreted
HCO3− concentration. |
|
|
| but in limited
quantities. |
|
|
| Liver | Subapical or
apical | AE2 is involved in
the regulation | 1. Decreased
expression of AE2 leads to |
|
| membrane of | of pHi homeostasis
and secretin- |
insufficient biliary
bicarbonate secretion |
|
| hepatobiliary
epithelial | stimulated biliary
bicarbonate | under
secretin stimulation. |
|
| cells. | secretion. | 2. In primary
biliary cholangitis, AE2 |
|
| Apical membrane
of |
|
expression and/or function are
reduced, |
|
|
cholangiocytes. |
| leading
to bile duct injury. |
|
|
|
| 3. In
hepatocellular carcinoma, AE2 is |
|
|
|
|
upregulated. |
Acknowledgements
Not applicable.
Funding
The present study was supported by grants from the
National Natural Science Foundation of China (grant no. 82073087),
National Natural Science Foundation of China (grant no. 81960507),
the Guizhou Provincial Department of Education Youth Science and
Technology Talents Growth Project [grant no. QIAN-JIAO-HE KY ZI
(2018)236], the Zunyi Medical University 2017 New Academic
Cultivation and Innovation Exploration Special Project [grant no.
Qian-Ke-He-Ping-Tai-Ren-Cai (2017)5733-072].
Availability of data and materials
Not applicable.
Authors' contributions
HW wrote the manuscript; JA, HJ and CL collected,
analyzed and organized the relevant literature; SH and JW revised
the grammar of the manuscript; BT revised the manuscript for
clarity and style. All authors read and approved the final
manuscript. Data authentication is not applicable.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Valdivieso ÁG and Santa-Coloma TA: The
chloride anion as a signalling effector. Biol Rev Camb Philos Soc.
94:1839–1856. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Trampert DC, van de Graaf SFJ, Jongejan A,
Oude Elferink RPJ and Beuers U: Hepatobiliary acid-base
homeostasis: Insights from analogous secretory epithelia. J
Hepatol. 74:428–441. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Jun I, Cheng MH, Sim E, Jung J, Suh BL,
Kim Y, Son H, Park K, Kim CH, Yoon JH, et al: Pore dilatation
increases the bicarbonate permeability of CFTR, ANO1 and glycine
receptor anion channels. J Physiol. 594:2929–2955. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Shin DH, Kim M, Kim Y, Jun I, Jung J, Nam
JH, Cheng MH and Lee MG: Bicarbonate permeation through anion
channels: Its role in health and disease. Pflugers Arch.
472:1003–1018. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Parker MD and Boron WF: The divergence,
actions, roles, and relatives of sodium-coupled bicarbonate
transporters. Physiol Rev. 93:803–959. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Bertocchio JP, Genetet S, Da Costa L,
Walsh SB, Knebelmann B, Galimand J, Bessenay L, Guitton C, De
Lafaille R, Vargas-Poussou R, et al: Red blood cell AE1/band 3
transports in dominant distal renal tubular acidosis patients.
Kidney Int Rep. 5:348–357. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Casey JR, Sly WS, Shah GN and Alvarez BV:
Bicarbonate homeostasis in excitable tissues: Role of AE3
Cl−/HCO3− exchanger and carbonic
anhydrase XIV interaction. Am J Physiol Cell Physiol.
297:C1091–C1102. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Sowah D, Brown BF, Quon A, Alvarez BV and
Casey JR: Resistance to cardiomyocyte hypertrophy in
ae3−/− mice, deficient in the AE3
Cl−/HCO3− exchanger. BMC
Cardiovasc Disord. 14:892014. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Kopito RR, Lee BS, Simmons DM, Lindsey AE,
Morgans CW and Schneider K: Regulation of intracellular pH by a
neuronal homolog of the erythrocyte anion exchanger. Cell.
59:927–937. 1989. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Linn SC, Kudrycki KE and Shull GE: The
predicted translation product of a cardiac AE3 mRNA contains an N
terminus distinct from that of the brain AE3
Cl−/HCO3− exchanger. Cloning of a
cardiac AE3 cDNA, organization of the AE3 gene, and identification
of an alternative transcription initiation site. J Biol Chem.
267:7927–7935. 1992. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Bronckers AL, Jalali R and Lytton J:
Reduced protein expression of the
Na+/Ca2++K+-exchanger (SLC24A4) in
apical plasma membranes of maturation ameloblasts of fluorotic
mice. Calcif Tissue Int. 100:80–86. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Celay J, Lozano T, Concepcion AR, Beltrán
E, Rudilla F, García-Barchino MJ, Robles EF, Rabal O, de Miguel I,
Panizo C, et al: Targeting the anion exchanger 2 with specific
peptides as a new therapeutic approach in B lymphoid neoplasms.
Haematologica. 103:1065–1072. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Vigliarolo T, Zocchi E, Fresia C, Booz V
and Guida L: Abscisic acid influx into human nucleated cells occurs
through the anion exchanger AE2. Int J Biochem Cell Biol.
75:99–103. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Wu C, Liu X, Sun R, Sun R, Qin Y, Liu Z,
Yang S, Tang T, Zhu Z, Yu D and Liu F: Targeting anion exchange of
osteoclast, a new strategy for preventing wear particles
induced-osteolysis. Front Pharmacol. 9:12912018. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Gawenis LR, Ledoussal C, Judd LM, Prasad
V, Alper SL, Stuart-Tilley A, Woo AL, Grisham C, Sanford LP,
Doetschman T, et al: Mice with a targeted disruption of the AE2
Cl−/HCO3− exchanger are
achlorhydric. J Biol Chem. 279:30531–30539. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Yang Y, Wu PP, Wu J, Shen WW, Wu YL, Fu
AF, Zheng L, Jin XL and Fu GH: Expression of anion exchanger 2 in
human gastric cancer. Exp Oncol. 30:81–87. 2008.PubMed/NCBI
|
|
17
|
Sasaki M, Sato Y and Nakanuma Y: An
impaired biliary bicarbonate umbrella may be involved in
dysregulated autophagy in primary biliary cholangitis. Lab Invest.
98:745–754. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Alper SL: Molecular physiology and
genetics of Na+-independent SLC4 anion exchangers. J Exp
Biol. 212:1672–1683. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Romero MF: Molecular pathophysiology of
SLC4 bicarbonate transporters. Curr Opin Nephrol Hypertens.
14:495–501. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Cordat E and Reithmeier RA: Structure,
function, and trafficking of SLC4 and SLC26 anion transporters.
Curr Top Membr. 73:1–67. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Liu Y, Yang J and Chen LM: Structure and
function of SLC4 Family [Formula: See text] Transporters. Front
Physiol. 6:3552015. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Stewart AK, Kurschat CE, Vaughan-Jones RD
and Alper SL: Putative re-entrant loop 1 of AE2 transmembrane
domain has a major role in acute regulation of anion exchange by
pH. J Biol Chem. 284:6126–6139. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Stewart AK, Kerr N, Chernova MN, Alper SL
and Vaughan-Jones RD: Acute pH-dependent regulation of AE2-mediated
anion exchange involves discrete local surfaces of the NH2-terminal
cytoplasmic domain. J Biol Chem. 279:52664–52676. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Layden TJ, Schmidt L, Agnone L, Lisitza P,
Brewer J and Goldstein JL: Rabbit esophageal cell cytoplasmic pH
regulation: Role of Na+-H+ antiport and
H+-dependent HCO3− transport
systems. Am J Physiol. 263:G407–G413. 1992.PubMed/NCBI
|
|
25
|
Tobey NA, Reddy SP, Khalbuss WE, Silvers
SM, Cragoe EJ Jr and Orlando RC: Na(+)-dependent and -independent
Cl−/HCO3− exchangers in cultured
rabbit esophageal epithelial cells. Gastroenterology. 104:185–195.
1993. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Laczkó D, Rosztóczy A, Birkás K, Katona M,
Rakonczay Z Jr, Tiszlavicz L, Róka R, Wittmann T, Hegyi P and
Venglovecz V: Role of ion transporters in the bile acid-induced
esophageal injury. Am J Physiol Gastrointest Liver Physiol.
311:G16–G31. 2016. View Article : Google Scholar
|
|
27
|
Abdulnour-Nakhoul S, Nakhoul HN, Kalliny
MI, Kalliny MI, Gyftopoulos A, Rabon E, Doetjes R, Brown K and
Nakhoul NL: Ion transport mechanisms linked to bicarbonate
secretion in the esophageal submucosal glands. Am J Physiol Regul
Integr Comp Physiol. 301:R83–R96. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Shiozaki A, Hikami S, Ichikawa D, Kosuga
T, Shimizu H, Kudou M, Yamazato Y, Kobayashi T, Shoda K, Arita T,
et al: Anion exchanger 2 suppresses cellular movement and has
prognostic significance in esophageal squamous cell carcinoma.
Oncotarget. 9:25993–26006. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Persi E, Duran-Frigola M, Damaghi M, Roush
WR, Aloy P, Cleveland JL, Gillies RJ and Ruppin E: Systems analysis
of intracellular pH vulnerabilities for cancer therapy. Nat Commun.
9:29972018. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Seidler U, Song P, Xiao F, Riederer B,
Bachmann O and Chen M: Recent advances in the molecular and
functional characterization of acid/base and electrolyte
transporters in the basolateral membranes of gastric and duodenal
epithelial cells. Acta Physiol (Oxf). 201:3–20. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Humphreys BD, Jiang L, Chernova MN and
Alper SL: Hypertonic activation of AE2 anion exchanger in Xenopus
oocytes via NHE-mediated intracellular alkalinization. Am J
Physiol. 268:C201–C209. 1995. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Rossmann H, Bachmann O, Wang Z, Shull GE,
Obermaier B, Stuart-Tilley A, Alper SL and Seidler U: Differential
expression and regulation of AE2 anion exchanger subtypes in rabbit
parietal and mucous cells. J Physiol. 534:837–848. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Alper SL: Molecular physiology of SLC4
anion exchangers. Exp Physiol. 91:153–161. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Recalde S, Muruzábal F, Looije N, Kunne C,
Burrell MA, Sáez E, Martínez-Ansó E, Salas JT, Mardones P, Prieto
J, et al: Inefficient chronic activation of parietal cells in
Ae2a,b(−/-) mice. Am J Pathol. 169:165–176. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Kurschat CE, Shmukler BE, Jiang L, Wihelm
S, Kim EH, Chernova MN, Kinne RK, Stewart AK and Alper SL:
Alkaline-shifted pHo sensitivity of AE2c1-mediated anion exchange
reveals novel regulatory determinants in the AE2 N-terminal
cytoplasmic domain. J Biol Chem. 281:1885–1896. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Huang QR, Li Q, Chen YH, Li L, Liu LL, Lei
SH, Chen HP, Peng WJ and He M: Involvement of anion exchanger-2 in
apoptosis of endothelial cells induced by high glucose through an
mPTP-ROS-Caspase-3 dependent pathway. Apoptosis. 15:693–704. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Wang T, Fei HJ, Yang Y, Jiang XS, Yan M,
Zeng Z, Wu J, Song LJ, Tian H and Fu GH: Expression of AE1/p16
promoted degradation of AE2 in gastric cancer cells. BMC Cancer.
16:7162016. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Wang T, Zhao L, Yang Y, Tian H, Suo WH,
Yan M and Fu GH: EGR1 is critical for gastrin-dependent
upregulation of anion exchanger 2 in gastric cancer cells. FEBS J.
280:174–183. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Cui Y, Li SB, Peng XC, Wu J and Fu GH:
Trastuzumab inhibits growth of HER2-negative gastric cancer cells
through gastrin-initialized CCKBR signaling. Dig Dis Sci.
60:3631–3641. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Walker NM, Flagella M, Gawenis LR, Shull
GE and Clarke LL: An alternate pathway of cAMP-stimulated Cl
secretion across the NKCC1-null murine duodenum. Gastroenterology.
123:531–541. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Murek M, Kopic S and Geibel J: Evidence
for intestinal chloride secretion. Exp Physiol. 95:471–478. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Zhu JX, Xue H, Ji T and Xing Y: Cellular
localization of NKCC2 and its possible role in the Cl- absorption
in the rat and human distal colonic epithelia. Transl Res.
158:146–154. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Hashemian F, Ghorbanian MA, Hashemian F,
Mortazavi SA, Sheikhi M, Jahanshahi J and Poorolajal J: Effect of
topical furosemide on rhinosinusal polyposis relapse after
endoscopic sinus surgery: A randomized clinical trial. JAMA
Otolaryngol Head Neck Surg. 142:1045–1049. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Walker NM, Liu J, Stein SR, Stefanski CD,
Strubberg AM and Clarke LL: Cellular chloride and bicarbonate
retention alters intracellular pH regulation in Cftr KO crypt
epithelium. Am J Physiol Gastrointest Liver Physiol. 310:G70–G80.
2016. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Peña-Münzenmayer G, Catalán MA, Kondo Y,
Jaramillo Y, Liu F, Shull GE and Melvin JE: Ae4 (Slc4a9) anion
exchanger drives Cl- uptake-dependent fluid secretion by mouse
submandibular gland acinar cells. J Biol Chem. 290:10677–10688.
2015. View Article : Google Scholar
|
|
46
|
Kaji I and Kaunitz JD: Luminal
chemosensing in the gastroduodenal mucosa. Curr Opin Gastroenterol.
33:439–445. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Takeuchi K, Kita K, Hayashi S and Aihara
E: Regulatory mechanism of duodenal bicarbonate secretion: Roles of
endogenous prostaglandins and nitric oxide. Pharmacol Ther.
130:59–70. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Tuo B, Wen G, Wei J, Liu X, Wang X, Zhang
Y, Wu H, Dong X, Chow JY, Vallon V and Dong H: Estrogen regulation
of duodenal bicarbonate secretion and sex-specific protection of
human duodenum. Gastroenterology. 141:854–863. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Alrefai WA, Tyagi S, Nazir TM, Barakat J,
Anwar SS, Hadjiagapiou C, Bavishi D, Sahi J, Malik P, Goldstein J,
et al: Human intestinal anion exchanger isoforms: Expression,
distribution, and membrane localization. Biochim Biophys Acta.
1511:17–27. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Alper SL, Rossmann H, Wilhelm S,
Stuart-Tilley AK, Shmukler BE and Seidler U: Expression of AE2
anion exchanger in mouse intestine. Am J Physiol. 277:G321–G332.
1999.PubMed/NCBI
|
|
51
|
Bachmann O and Seidler U: News from the
end of the gut-how the highly segmental pattern of colonic
HCO3− transport relates to absorptive
function and mucosal integrity. Biol Pharm Bull. 34:794–802. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Ikuma M, Geibel J, Binder HJ and Rajendran
VM: Characterization of Cl-HCO3 exchange in basolateral
membrane of rat distal colon. Am J Physiol Cell Physiol.
285:C912–C921. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
53
|
Gawenis LR, Bradford EM, Alper SL, Prasad
V and Shull GE: AE2 Cl−/HCO3−
exchanger is required for normal cAMP-stimulated anion secretion in
murine proximal colon. Am J Physiol Gastrointest Liver Physiol.
298:G493–G503. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
54
|
Jeong YS and Hong JH: Governing effect of
regulatory proteins for Cl(−)/HCO3(−) exchanger 2
activity. Channels (Austin). 10:214–224. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
55
|
O'Mahony F and Harvey BJ: Sex and estrous
cycle-dependent rapid protein kinase signaling actions of estrogen
in distal colonic cells. Steroids. 73:889–894. 2008. View Article : Google Scholar
|
|
56
|
Song LJ, Liu RJ, Zeng Z, Alper SL, Cui HJ,
Lu Y, Zheng L, Yan ZW and Fu GH: Gastrin inhibits a novel,
pathological colon cancer signaling pathway involving EGR1, AE2,
and P-ERK. J Mol Med (Berl). 90:707–718. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
57
|
Hulikova A, Black N, Hsia LT, Wilding J,
Bodmer WF and Swietach P: Stromal uptake and transmission of acid
is a pathway for venting cancer cell-generated acid. Proc Natl Acad
Sci USA. 113:E5344–E5353. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
58
|
Roussa E, Alper SL and Thévenod F:
Immunolocalization of anion exchanger AE2, Na(+)/H(+) exchangers
NHE1 and NHE4, and vacuolar type H(+)-ATPase in rat pancreas. J
Histochem Cytochem. 49:463–474. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
59
|
Rakonczay Z Jr, Fearn A, Hegyi P, Boros I,
Gray MA and Argent BE: Characterization of H+ and
HCO3− transporters in CFPAC-1 human
pancreatic duct cells. World J Gastroenterol. 12:885–895. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
60
|
Rakonczay Z Jr, Hegyi P, Hasegawa M, Inoue
M, You J, Lida A, Lgnáth I, Alton EWFWA, Griesenbach U, Ovári G, et
al: CFTR gene transfer to human cystic fibrosis pancreatic duct
cells using a Sendai virus vector. J Cell Physiol. 214:442–455.
2008. View Article : Google Scholar : PubMed/NCBI
|
|
61
|
Park HW, Nam JH, Kim JY, Namkung W, Yoon
JS, Lee JS, Kim KS, Venglovecz V, Gray MA, Kim KH and Lee MG:
Dynamic regulation of CFTR bicarbonate permeability by [Cl-]i and
its role in pancreatic bicarbonate secretion. Gastroenterology.
139:620–631. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
62
|
Yamaguchi M, Steward MC, Smallbone K,
Sohma Y, Yamamoto A, Ko SB, Kondo T and Ishiguro H:
Bicarbonate-rich fluid secretion predicted by a computational model
of guinea-pig pancreatic duct epithelium. J Physiol. 595:1947–1972.
2017. View Article : Google Scholar : PubMed/NCBI
|
|
63
|
Hong JH, Muhammad E, Zheng C, Hershkovitz
E, Alkrinawi S, Loewenthal N, Parvari R and Muallem S: Essential
role of carbonic anhydrase XII in secretory gland fluid and
HCO3(−) secretion revealed by disease causing human
mutation. J Physiol. 593:5299–5312. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
64
|
Venter J, Francis H, Meng F, DeMorrow S,
Kennedy L, Standeford H, Hargrove L, Wu N, Wan Y, Frampton G, et
al: Development and functional characterization of extrahepatic
cholangiocyte lines from normal rats. Dig Liver Dis. 47:964–972.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
65
|
Tietz PS, Marinelli RA, Chen XM, Huang B,
Cohn J, Kole J, McNiven MA, Alper S and LaRusso NF: Agonist-induced
coordinated trafficking of functionally related transport proteins
for water and ions in cholangiocytes. J Biol Chem. 278:20413–20419.
2003. View Article : Google Scholar : PubMed/NCBI
|
|
66
|
Aranda V, Martínez I, Melero S, Lecanda J,
Banales JM, Prieto J and Medina JF: Shared apical sorting of anion
exchanger isoforms AE2a, AE2b1, and AE2b2 in primary hepatocytes.
Biochem Biophys Res Commun. 319:1040–1046. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
67
|
García C, Montuenga LM, Medina JF and
Prieto J: In situ detection of AE2 anion-exchanger mRNA in the
human liver. Cell Tissue Res. 291:481–488. 1998. View Article : Google Scholar
|
|
68
|
Beuers U, Maroni L and Elferink RO: The
biliary HCO(3)(−) umbrella: Experimental evidence revisited. Curr
Opin Gastroenterol. 28:253–257. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
69
|
Rodrigues PM, Perugorria MJ, Santos-Laso
A, Bujanda L, Beuers U and Banales JM: Primary biliary cholangitis:
A tale of epigenetically-induced secretory failure? J Hepatol.
69:1371–1383. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
70
|
Hohenester S, Wenniger LM, Paulusma CC,
van Vliet SJ, Jefferson DM, Elferink RP and Beuers U: A biliary
HCO3- umbrella constitutes a protective mechanism
against bile acid-induced injury in human cholangiocytes.
Hepatology. 55:173–183. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
71
|
Chang JC, Go S, de Waart DR, Munoz-Garrido
P, Beuers U, Paulusma CC and Oude Elferink R: Soluble adenylyl
cyclase regulates bile salt-induced apoptosis in human
cholangiocytes. Hepatology. 64:522–534. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
72
|
Maillette de Buy Wenniger LJ, Hohenester
S, Maroni L, van Vliet SJ, Oude Elferink RP and Beuers U: The
cholangiocyte glycocalyx stabilizes the ‘Biliary HCO3
umbrella’: An integrated line of defense against toxic bile acids.
Dig Dis. 33:397–407. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
73
|
Miszczuk GS, Banales JM, Zucchetti AE,
Pisani GB, Boaglio AC, Saez E, Medina JF, Roma MG and Crocenzi FA:
Adaptive downregulation of
Cl−/HCO3− exchange activity in rat
hepatocytes under experimental obstructive cholestasis. PLoS One.
14:e02122152019. View Article : Google Scholar : PubMed/NCBI
|
|
74
|
Uriarte I, Banales JM, Sáez E, Arenas F,
Oude Elferink RP, Prieto J and Medina JF: Bicarbonate secretion of
mouse cholangiocytes involves Na(+)-HCO(3)(−) cotransport in
addition to Na(+)-independent Cl(−)/HCO(3)(−) exchange. Hepatology.
51:891–902. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
75
|
Erice O, Munoz-Garrido P, Vaquero J,
Perugorria MJ, Fernandez-Barrena MG, Saez E, Santos-Laso A,
Arbelaiz A, Jimenez-Agüero R, Fernandez-Irigoyen J, et al:
MicroRNA-506 promotes primary biliary cholangitis-like features in
cholangiocytes and immune activation. Hepatology. 67:1420–1440.
2018. View Article : Google Scholar : PubMed/NCBI
|
|
76
|
Banales JM, Prieto J and Medina JF:
Cholangiocyte anion exchange and biliary bicarbonate excretion.
World J Gastroenterol. 12:3496–3511. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
77
|
Renzi A, DeMorrow S, Onori P, Carpino G,
Mancinelli R, Meng F, Venter J, White M, Franchitto A, Francis H,
et al: Modulation of the biliary expression of arylalkylamine
N-acetyltransferase alters the autocrine proliferative responses of
cholangiocytes in rats. Hepatology. 57:1130–1141. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
78
|
Banales JM, Sáez E, Uriz M, Sarvide S,
Urribarri AD, Splinter P, Tietz Bogert PS, Bujanda L, Prieto J,
Medina JF and LaRusso NF: Up-regulation of microRNA 506 leads to
decreased Cl−/HCO3− anion
exchanger 2 expression in biliary epithelium of patients with
primary biliary cirrhosis. Hepatology. 56:687–697. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
79
|
Brooks WH and Renaudineau Y: Epigenetics
and autoimmune diseases: The X chromosome-nucleolus nexus. Front
Genet. 6:222015. View Article : Google Scholar : PubMed/NCBI
|
|
80
|
Wang W, Ren X, Cai Y, Chen L, Zhang W and
Xu J: Rifampicin induces bicarbonate-rich choleresis in rats:
Involvement of anion exchanger 2. Dig Dis Sci. 61:126–136. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
81
|
Medina JF: Role of the anion exchanger 2
in the pathogenesis and treatment of primary biliary cirrhosis. Dig
Dis. 29:103–112. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
82
|
Tian L, Ye Z, Kafka K, Stewart D, Anders
R, Schwarz KB and Jang YY: Biliary atresia relevant human induced
pluripotent stem cells recapitulate key disease features in a dish.
J Pediatr Gastroenterol Nutr. 68:56–63. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
83
|
Arenas F, Hervías I, Sáez E, Melero S,
Prieto J, Parés A and Medina JF: Promoter hypermethylation of the
AE2/SLC4A2 gene in PBC. JHEP Rep. 1:145–153. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
84
|
Melero S, Spirlì C, Zsembery A, Medina JF,
Joplin RE, Duner E, Zuin M, Neuberger JM, Prieto J and Strazzabosco
M: Defective regulation of cholangiocyte
Cl−/HCO3(−) and Na+/H+
exchanger activities in primary biliary cirrhosis. Hepatology.
35:1513–1521. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
85
|
Hisamoto S, Shimoda S, Harada K, Iwasaka
S, Onohara S, Chong Y, Nakamura M, Bekki Y, Yoshizumi T, Ikegami T,
et al: Hydrophobic bile acids suppress expression of AE2 in biliary
epithelial cells and induce bile duct inflammation in primary
biliary cholangitis. J Autoimmun. 75:150–160. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
86
|
Prieto J, Qian C, García N, Díez J and
Medina JF: Abnormal expression of anion exchanger genes in primary
biliary cirrhosis. Gastroenterology. 105:572–578. 1993. View Article : Google Scholar : PubMed/NCBI
|
|
87
|
Concepcion AR, Salas JT, Sáez E, Sarvide
S, Ferrer A, Portu A, Uriarte I, Hervás-Stubbs S, Oude Elferink RP,
et al: CD8+ T cells undergo activation and programmed death-1
repression in the liver of aged Ae2a,b−/− mice favoring
autoimmune cholangitis. Oncotarget. 6:28588–28606. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
88
|
Salas JT, Banales JM, Sarvide S, Recalde
S, Ferrer A, Uriarte I, Oude Elferink RP, Prieto J and Medina JF:
Ae2a,b-deficient mice develop antimitochondrial antibodies and
other features resembling primary biliary cirrhosis.
Gastroenterology. 134:1482–1493. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
89
|
Concepcion AR, Salas JT, Sarvide S, Sáez
E, Ferrer A, López M, Portu A, Banales JM, Hervás-Stubbs S, Oude
Elferink RP, et al: Anion exchanger 2 is critical for CD8(+) T
cells to maintain pHi homeostasis and modulate immune responses.
Eur J Immunol. 44:1341–1351. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
90
|
Poupon R, Ping C, Chrétien Y, Corpechot C,
Chazoillères O, Simon T, Heath SC, Matsuda F, Poupon RE, Housset C
and Barbu V: Genetic factors of susceptibility and of severity in
primary biliary cirrhosis. J Hepatol. 49:1038–1045. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
91
|
Zhang L, Su H, Li Y, Fan YJ, Wang Q, Jiang
J, Hu Y, Chen G, Tan B and Qiu F: Different effects of
ursodeoxycholic acid on intrahepatic cholestasis in acute and
recovery stages induced by alpha-naphthylisothiocyanate in mice.
Toxicol Appl Pharmacol. 342:69–78. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
92
|
Úriz M, Sáez E, Prieto J, Medina JF and
Banales JM: Ursodeoxycholic acid is conjugated with taurine to
promote secretin-stimulated biliary hydrocholeresis in the normal
rat. PLoS One. 6:e287172011. View Article : Google Scholar
|
|
93
|
Wu TT, Hsieh YH, Wu CC, Tsai JH, Hsieh YS,
Huang CY and Liu JY: Overexpression of anion exchanger 2 in human
hepatocellular carcinoma. Chin J Physiol. 49:192–198.
2006.PubMed/NCBI
|
|
94
|
Liu CJ, Hwang JM, Wu TT, Hsieh YH, Wu CC,
Hsieh YS, Tsai CH, Wu HC, Huang CY and Liu JY: Anion exchanger
inhibitor DIDS induces human poorly-differentiated malignant
hepatocellular carcinoma HA22T cell apoptosis. Mol Cell Biochem.
308:117–125. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
95
|
Hwang JM, Kao SH, Hsieh YH, Li KL, Wang
PH, Hsu LS and Liu JY: Reduction of anion exchanger 2 expression
induces apoptosis of human hepatocellular carcinoma cells. Mol Cell
Biochem. 327:135–144. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
96
|
Svastova E, Witarski W, Csaderova L, Kosik
I, Skvarkova L, Hulikova A, Zatovicova M, Barathova M, Kopacek J,
Pastorek J and Pastorekova S: Carbonic anhydrase IX interacts with
bicarbonate transporters in lamellipodia and increases cell
migration via its catalytic domain. J Biol Chem. 287:3392–3402.
2012. View Article : Google Scholar : PubMed/NCBI
|