Introduction
Liver cancer is a malignant tumor with high
morbidity and mortality rate. Liver cancer is the sixth commonest
cancer; its 2018 global mortality rate was ~8.2%, ranking fourth
among all types of cancer mortality (1–3). Most
liver cancer cases reach middle or late stages before they are
diagnosed, and the best time for surgical treatment is lost
(4,5). Therefore, chemotherapy is the main
treatment for liver cancer. However, chemotherapy is prone to drug
resistance, which has become a major problem faced by chemotherapy
for liver cancer. Oxaliplatin has been approved for systemic
chemotherapy in patients with liver cancer, but its efficacy is
often reduced because of drug resistance (6,7).
Therefore, it is necessary to explore the resistance mechanisms and
find a more effective treatment strategy.
Resistance of liver cancer to chemotherapy is
related to abnormal activation of the PI3K/AKT/hypoxia inducible
factor (HIF)-1α signaling pathway (8–10).
PI3K catalyzes the production of phosphatidylinositol-
3,4,5-triphosphate through phosphorylation of
phosphatidy-linositol, phosphatidylinositol-4-phosphate and
phosphatidylinositol- 4,5-bisphosphate (11,12).
Chemotherapy drugs trigger this phosphorylation event, which in
turn regulates cell proliferation, cell-cycle progression, cell
migration and cell survival (13).
In addition, several chemotherapy drugs can activate the
serine/threonine kinase AKT (also known as protein kinase B). As a
proto-oncogene, AKT plays an important role in regulating various
cell functions, including metabolism, proliferation, survival,
transcription and protein synthesis (14–17).
HIF-1α, a functional subunit of HIF-1, can regulate angiogenesis,
red blood cell production, cell cycle, metabolism and apoptosis.
More importantly, HIF-1α can be induced by a variety of cytokines
(including PTEN, SRC and P53) in an oxygen-independent manner
through the PI3K/AKT pathway (18,19).
However, whether the combined inhibition of the PI3K/AKT pathway
and HIF-1α expression can increase the sensitivity of liver cancer
cells to chemotherapy is not known.
LY294002 is a protein kinase inhibitor that can
block the cell signaling pathway of phosphotidylinsitol-3-kinase
and inhibit the expression of PI3Kα, PI3Kδ and PI3Kβ. LY294002 can
enter cells and inhibit the PI3K and PI3K/Akt signaling pathways,
including inhibition of Akt phosphorylation, thereby inhibiting
cell division and induce G1 arrest of pancreatic cancer
and other cells (20). MK-2206 is a
highly selective AKT1/2/3 inhibitor, activated by the pleckstrin
homology domain. MK-2206 inhibits the autophosphorylation of AKT at
threonine 308 and serine 473 (21).
Preclinical studies have shown that MK-2206 has an inhibitory
effect on human cancer cell lines, such as breast, lung, colon and
liver cancer (22–25). PI3K/AKT/HIF-1α reportedly induces
chemoresistance in liver cancer cells (26,27),
and LY-294002 can enhance the chemosensitivity of liver cancer
(28). Therefore, the present study
used LY-294002 in combination with oxaliplatin.
The present study explored the effect of LY-294002
or MK-2206 in combination with oxaliplatin on the abnormal
activation of the PI3K/AKT pathway in oxaliplatin-resistant liver
cancer. The focus of the study was the more significant inhibitory
effect of LY-294002 and whether its effect is due to the inhibition
of HIF-1α expression. It was proposed that HIF-1α may be a target
to improve the sensitivity of chemotherapeutics, and that the
combination of LY-294002 and oxaliplatin will be beneficial in the
treatment of liver cancer and provide a basis for the development
of targeted therapeutic strategies against liver cancer.
Materials and methods
Cell source and culture
The human liver cancer cell line HepG2 (control
HepG2 cells) was purchased from Shanghai Mingjin Biological
Technology Co, Ltd. The oxaliplatin-resistant HepG2 cell line,
HepG2R, was induced in medium containing increasing
concentration of oxaliplatin (0–40 µM) for a total of seven months.
The two cell lines were cultured in RPMI-1640 medium (Hyclone;
Cyvita) containing 10% fetal bovine serum (Hangzhou Sijiqing
Biological Engineering Materials Co., Ltd.) and maintained in a 5%
CO2 incubator at 37°C.
Chemicals, antibodies and
reagents
LY-294002 (0.2 µM) and MK-2206 (50 nmol/l; MedChem
Express) were dissolved in dimethyl sulfoxide (DMSO) to prepare a 2
mM stock solution. Oxaliplatin (Sigma-Aldrich; Merck KGaA) was
dissolved in water to produce a 2 mM stock solution and stored at
−20°C. Phosphorylated (p-) retinoblastoma [Rb; anti-Rb (phospho
S807) antibody [EPR17732] (cat. no. ab184796)] and cyclin D1
[anti-Cyclin D1 antibody (SP4) cat. no. ab16663] antibodies were
purchased from Abcam. Antibodies against caspase-3 (cat. no. 9662),
cleaved caspase-3 (cat. no. 9661), caspase-9 (cat. no. 9502),
cleaved caspase-9 (cat. no. 20750), PARP (cat. no. 9532), cleaved
RARP (cat. no. 5625), PI3Kp110α (cat. no. 4249), PI3Kp110β (cat.
no. 3011), PI3Kp110γ (cat. no. 5405), PI3Kp85 (cat. no. 4292), Akt
(cat. no. 4691), p-Akt (Ser473; cat. no. 4060), Bad (cat. no.
9268), p-Bad (cat. no. 5284), Bax (cat. no. 14796) and Puma (cat.
no. 24633) were purchased from Cell Signaling Technology, Inc. The
p70S6K (cat. no. BM4240), p-S6K1 (T421+S424) (cat. no. BM4141) and
eukaryotic translation initiation factor 4E-binding protein 1
(eIF4EBP1) antibodies (cat. no. BM4851) were from Wuhan Boster
Biological Technology, Ltd. The p-eIF4EBP1 antibody (cat. no.
bs-14550R) was from BIOSS. β-actin antibody (cat. no. AF5003) and
BCA-200 protein detection kits were from Biosharp Life Sciences.
All antibodies were diluted 1:1,000 before use. Phosphatase
inhibitor mixture were provided by Beyotime Institute of
Biotechnology.
Western blot analysis
HepG2 and HepG2R [Oxaliplatin (2 µM) treated 0, 6,
12, 24, 48, 72 and 96 h] cells (5.0×106) were added with
lysis buffer (phosphatase inhibitor mixture) to extract protein.
SDS-PAGE gels (10%) were placed at room temperature for 30 min, and
20 µl of protein samples were added to each well. The protein
samples, separated by SDS-PAGE, were sequentially transferred on
PVDF membrane (EMD Millipore) according to molecular weight. After
being blocked with 5% skimmed milk at 37°C for 1 h, the
corresponding primary antibodies were incubated at 4°C overnight
After blocking with 5% skimmed milk at 37°C for 1 h, the
corresponding primary antibody was incubated overnight at 4°C, and
β-actin was used as a load control. The membrane was washed three
times with TBST [10 M Tris, 150 mM NaCl, 0.05% Tween 20 (pH 8.3)],
and secondary antibody was added and incubated at 37°C for 1 h. The
membrane was washed again three times with TBST, luminescent
solution (ECL Luminescence kit; EMD Millipore) was added for color
development. ImageJ 1.44p software (National Institutes of Health)
was used for immunoblotting quantitative analysis. The gray value
of each imprinted signal was compared with the control group to
analyze the expression of the target gene protein.
MTT assay
Cells were seeded in 96-well plates at a density of
5×103 cells/well and placed in an incubator for 24 h,
and drugs were added. After 24 h, the medium was discarded, 10 µl
of MTT medium was added to each well, and the plates were placed in
an incubator. After culturing for 4 h, the culture solution was
discarded, 100 µl of DMSO was added to each well and shaken for 10
min on a shaker. The absorbance at 490 nm was detected with an
enzyme-linked immunoassay detector (ELx800; Omega Bio-Tek,
Inc.).
Colony formation assay
The cells [HepG2R, plus oxaliplatin (2
µM) and/or LY-294002(0.2 µM)] were seeded in a six-well plate at a
density of 1×103 cells/well. After the cells had adhered
to the wall, drug was added. After 24 h, the drug-free medium was
replaced, and the culture was continued for ~3 weeks. When the cell
clones were clearly visible, the cell colonies were washed twice
with pre-cooled physiological saline and fixed with 4%
paraformaldehyde for 15 min at room temperature. After the fixing
solution was removed, the cells were stained with crystal violet
staining solution for 15 min at room temperature. The number of
clones formed was counted to reflect the proliferation ability of
cells under the action of each drug.
Cell cycle analysis
The cells were seeded in a six-well plate at a
density of 1×105 cells/well and placed in an incubator
for 24 h. Drugs were added for 24 h, and the cells were digested
with trypsin for collection of the cells, which were fixed with 70%
ethanol at room temperature for 1 h. Pre-cooled physiological
saline (1 ml) was added to the cells, the cells were centrifuged
(562.5 × g; room temperature; 5 min) and 100 µl of staining
solution from the AnnexinV-FITC/PI staining kit (Jiangsu KGI
Biotechnology Co., Ltd.) was added and incubated at 4°C in the dark
for 30 min (staining solution is a mixture of 50 µg/ml PI, 100
µg/ml RNaseA and 0.2% Triton X-100 solution). In total, 1 ml of
pre-cooled physiological saline was added to the staining solution,
and the cells were centrifuged (562.5 × g; room temperature; 5 min)
to obtain a cell pellet. Finally, the cells were resuspended in 300
µl of physiological saline, resuspended and analyzed with a flow
cytometer (BD FACSCalibur; BD Biosciences). The results were
analyzed with ModiFit LT 3.0 (Verity Software House, Inc.).
Annexin V FITC/PI and DAPI
staining
The cells were seeded in a 24-well plate containing
slides at a density of 1×104 cells/well, placed in an
incubator for 24 h, and treated with drug. After 24 h, the slides
were taken out, washed twice with normal saline, and stained with
Annexin V FITC/PI (Jiangsu KGI Biotechnology Co., Ltd.) and DAPI
(Shanghai Biyuntian Biotechnology Co., Ltd.) for 15 min at room
temperature in the dark. Apoptotic rate was determined with a
fluorescence microscope; 10 fields of view were evaluated at ×200
magnification. Apoptosis rate=(early apoptotic cells + late
apoptotic cells)/total number of cells.
Measurement of mitochondrial membrane
potential (JC-1)
The cells were inoculated into the well plates at a
density of 1×104 cells/well, placed in an incubator for
24 h, and treated with drug for another 24 h. The cells were rinsed
twice with saline, 500 µl JC-1 working solution was added to each
well, and the plates were placed in an incubator (5%
CO2, 37°C) and incubated for an additional 20 min. The
staining solution was discarded, and the cells were rinsed with
saline twice, then stained with DAPI solution at room temperature
in dark for 15 min. As JC-1 aggregates in the mitochondria when the
mitochondrial membrane potential is high, red fluorescence is
emitted; conversely, when mitochondria are depolarized, JC-1 is in
a monomer state and green fluorescence is emitted. Thus, the
mitochondrial membrane potential of the cell can be judged
according to the ratio of red and green fluorescence, and change of
JC-1 fluorescence color can indicate change of the mitochondrial
membrane potential. A decrease in mitochondrial membrane potential
is considered a sign of early apoptosis (29). Red and green fluorescence were
analyzed with a flow cytometer (BD FACSCalibur; BD Biosciences).
The results were analyzed with CellQuest Pro 5.1.1 (Becton,
Dickinson and Company).
Statistical analysis
The experimental data were obtained through at least
three independent experiments. The data are expressed as mean ± SD,
one-way ANOVA to measure significant differences between the means
followed by Tukey's post hoc tests. P<0.05 was considered to
indicate a statistically significant difference. SPSS 17.0
statistical software (SPSS Inc.) was used for all analyses.
Results
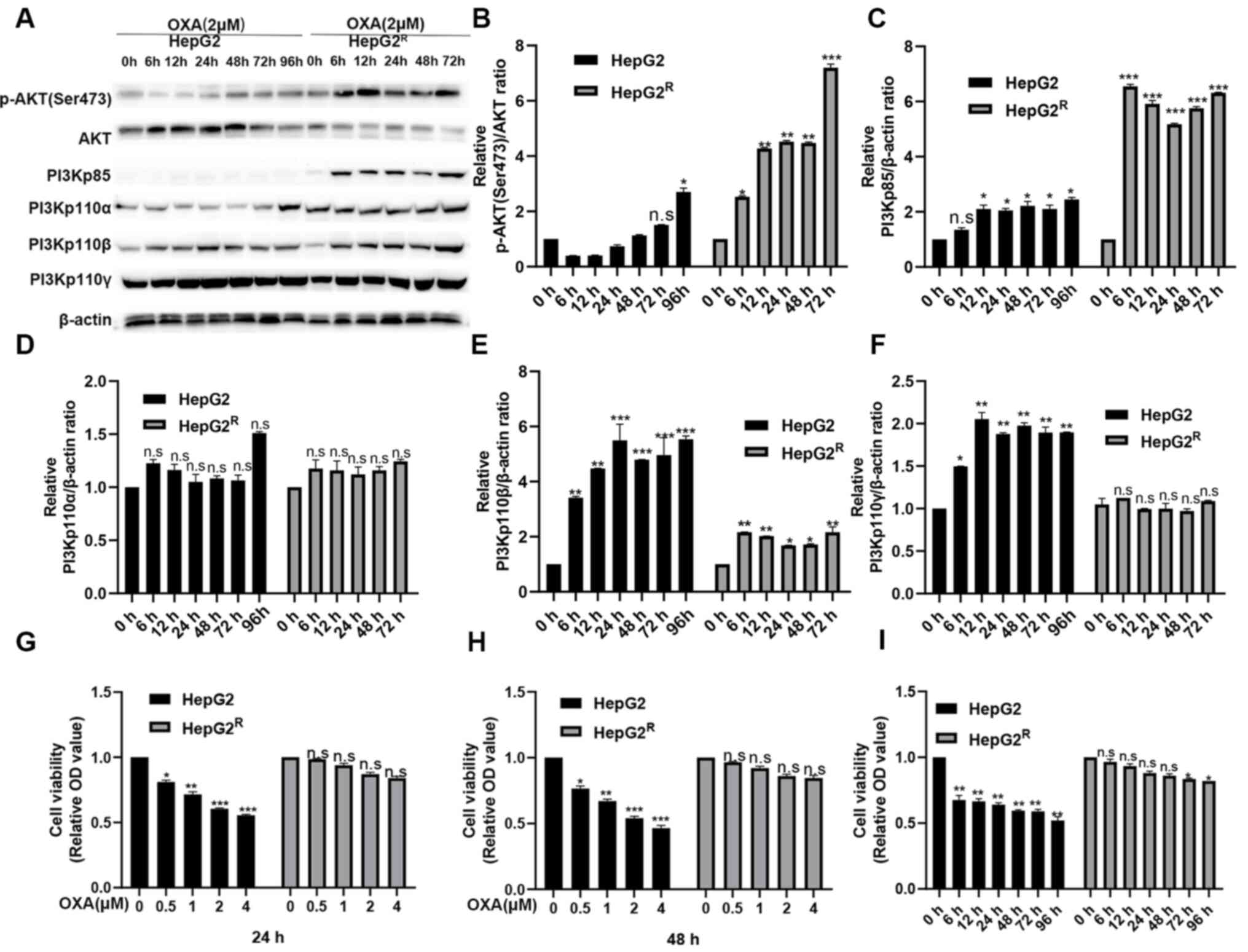
Stable activation of PI3K/AKT pathway
in HepG2R cells induced by oxaliplatin
HepG2 and HepG2R cells were treated with
2 µM oxaliplatin. Protein was extracted after 0, 6, 12, 24, 48, 72
and 96 h and measured using western blotting. p-AKT (Ser473),
PI3Kp85, PI3Kp110α, PI3Kp110β were found all strongly expressed in
HepG2R but weakly expressed in HepG2 (Fig. 1A-F). However, the expression level
of PI3Kp110γ was not different between the HepG2 and
HepG2R cells; it is possible that PI3Kp110γ is not as
effective as PI3Kp110α and PI3Kp110β in the PI3K/AKT signaling
pathway. These results indicated that the PI3K/AKT signaling
pathway activation level was higher in HepG2R compared
with in HepG2 cells. The HepG2 and HepG2R cells were
treated with various concentrations of oxaliplatin for 24 or 48 h,
and cell viability of was analyzed using an MTT assay. As shown in
Fig. 1G-H, oxaliplatin inhibited
the cell viability of HepG2 in a time- and concentration-dependent
manner, but the proliferation of HepG2R was almost
unaffected. HepG2 and HepG2R cells were treated with 2
µmol/l oxaliplatin for the same time periods, and cell viability
was determined. The viability of HepG2 cells was found gradually
decreased over time, but the viability of HepG2R did not
change significantly (Fig. 1I).
Since the abnormal activation of the PI3K/AKT signaling pathway
promoted the proliferation of HepG2R, it was
hypothesized that the PI3K/AKT pathway inhibitors (LY-294002 and
MK-2206) enhanced the sensitivity of liver cancer to oxaliplatin by
targeting the PI3K/AKT pathway.
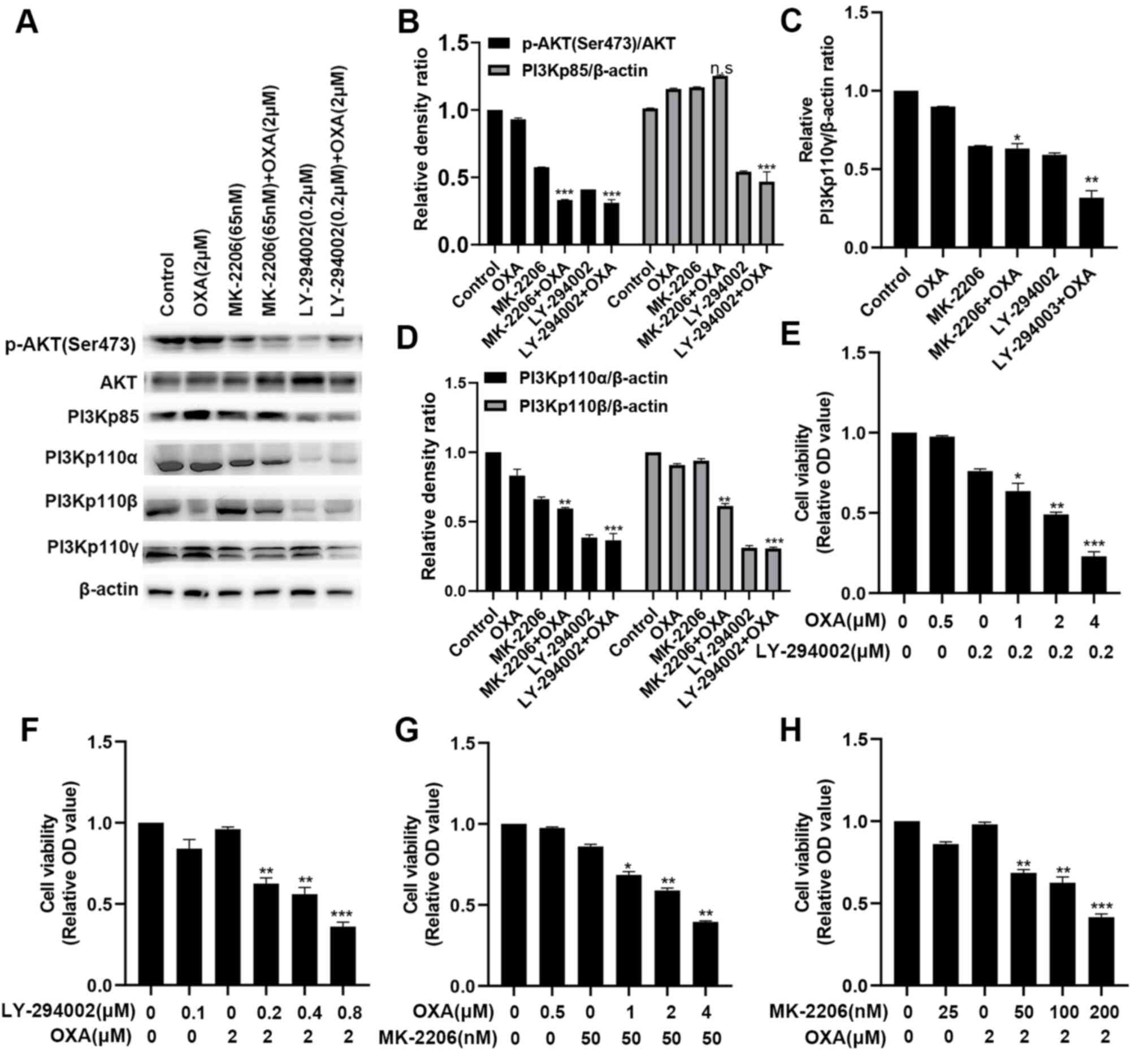
LY-294002 can more effectively inhibit
the PI3K/AKT pathway in the HepG2R compared with HepG2
cell line
To determine the effect of the PI3K site-specific
inhibitor (LY-294002) or the AKT site-specific inhibitor (MK-2206)
in combination with 2 µmol/l oxaliplatin on the PI3K/AKT pathway in
HepG2R, the expression levels of PI3Kp110α, PI3Kp110β,
PI3Kp110γ, AKT, p-AKT (Ser473) and PI3Kp85 were measured. It was
found that LY-294002 (0.2 µM) and MK-2206 (50 nmol/l) downregulated
the activation of the PI3K/AKT signaling pathway. The inhibitory
effect for p-AKT(Ser473) was similar, but for PI3Kp110α, PI3Kp110β
and PI3Kp110γ, the inhibitory effect of LY-294002 was greater
(Fig. 2A-D). MTT assays were also
conducted. As shown in Fig. 2E-H,
the combination treatment more effectively inhibited
HepG2R cell viability compared with single-drug
treatment, and LY-294002 combined with oxaliplatin had a stronger
inhibitory effect compared with MK-2206 with oxaliplatin. These
results demonstrated that LY-294002 targeting the PI3K/AKT pathway
can enhance the chemosensitivity of oxaliplatin.
LY-294002 combined with oxaliplatin
effectively inhibits the proliferation of HepG2R
cells
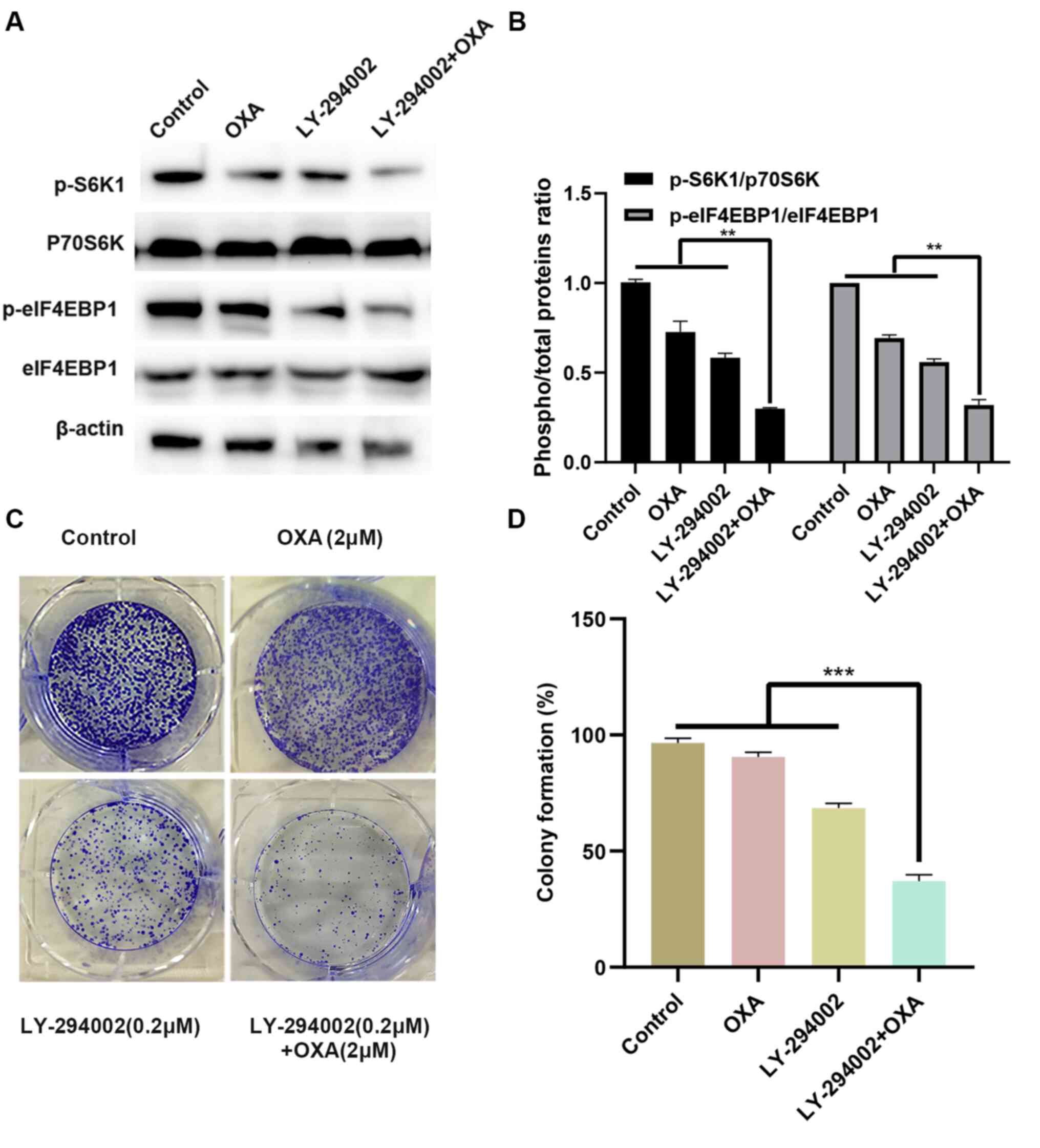
To improve our understanding of the molecular
mechanism by which LY-294002 inhibits cell proliferation,
HepG2R cells were treated with 2 µmol/l oxaliplatin
and/or 0.2 µmol/l LY-294002. As illustrated in Fig. 3A and B, the combination of LY-294002
and oxaliplatin significantly downregulated the phosphorylation
levels of the downstream proliferation signaling molecules S6K1 and
eIF4EBP1 in the PI3K/AKT signaling pathway. The effect of LY-294002
and oxaliplatin on tumorigenesis was also evaluated in the
HepG2R cell line with a colony formation assay. Compared
with LY-294002 or oxaliplatin alone, the combination of LY-294002
and oxaliplatin induced the least number of clone formation of
HepG2R cells and exerted a stronger inhibitory effect
(Fig. 3C and D). These results
indicated that LY-294002 inhibited the proliferation of
HepG2R by blocking the PI3K/AKT/S6K1/eIF4EBP1 signaling
pathway.
LY-294002 combined with oxaliplatin
blocks the cell cycle of HepG2R
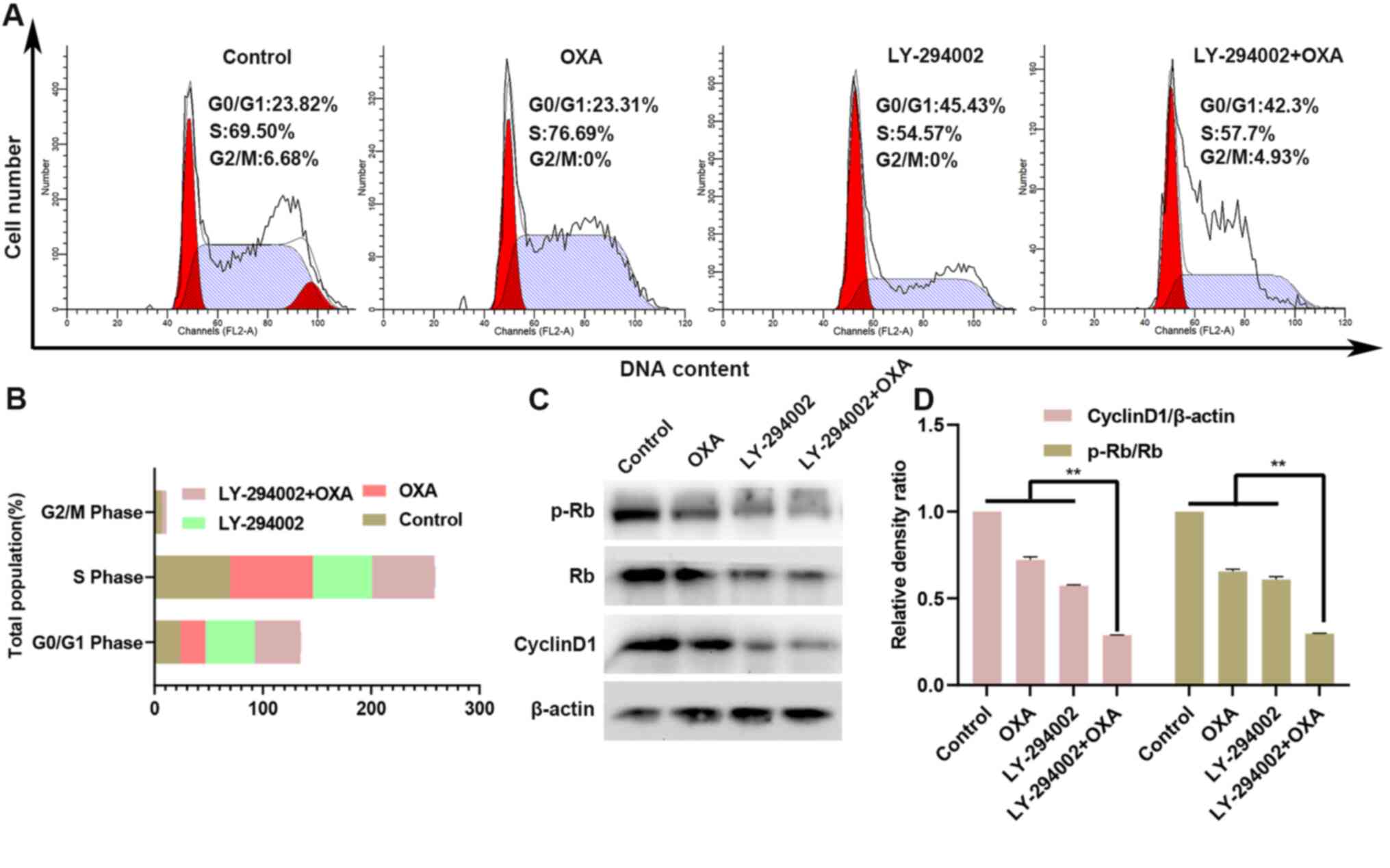
The HepG2R cell line was treated with
oxaliplatin and/or LY-294002 (0.2 µM) for 24 h, and cell-cycle
assays were performed. As shown in Fig.
4A and B, compared with the control group, the number of
G0/G1 phase cells was increased in the
LY-294002 group, while the number of G2/M phase and S
phase cells was decreased. However, there was almost no change in
the oxaliplatin-only treated group. Compared with treatment with
oxaliplatin alone, treatment of the HepG2R cell line
with LY-294002 and oxaliplatin for 24 h further increased the
number of cells in G0/G1 phase, while the
number of cells in G2/M phase and S phase was decreased.
Thus, the combination of LY-294002 and oxaliplatin induced
cell-cycle arrest in the G0/G1 phase. The
expression levels of the downstream cycle molecules cyclin D1 and
p-Rb in the PI3K/AKT signaling pathway were also measured in the
HepG2R cell line. As seen in Fig. 4C and D, compared with oxaliplatin
treatment alone, the treatment with the combination of LY-294002
and oxaliplatin downregulated the expression of cyclin D1 and the
p-Rb/Rb ratio.
Combination of LY-294002 and
oxaliplatin promotes the apoptosis of HepG2R cells
The effect of apoptosis on HepG2R cells
was determined with Annexin V FITC/PI staining and DAPI kit after
oxaliplatin and/or LY-294002 (0.2 µM) treatment for 24 h. As shown
in Fig. 5A and D, compared with the
oxaliplatin single-agent group, the LY-294002 and oxaliplatin
combination group had a higher apoptosis rate. After treatment of
the HepG2R cell line with oxaliplatin and/or LY-294002
for 24 h, the mitochondrial membrane potential was measured using
JC-1 staining and flow cytometry. The results presented in Fig. 5B and E illustrate that, compared
with the oxaliplatin single-agent group, the red fluorescence ratio
of the combination-drug group was lower, indicating that the
apoptosis rate of HepG2R cells was higher. At the same
time, the results presented in Fig. 5C
and F-G illustrate that, compared with the oxaliplatin
single-agent group, LY-294002 combined with oxaliplatin inhibited
the expression of downstream anti-apoptotic signaling molecules of
the PI3K/AKT signaling pathway, such as p-Bad, and increased the
expression of pro-apoptotic signaling molecules, such as Bad, Bax,
Puma, PARP, cleaved caspase-3 and −9 and promoted apoptosis of the
HepG2R cells.
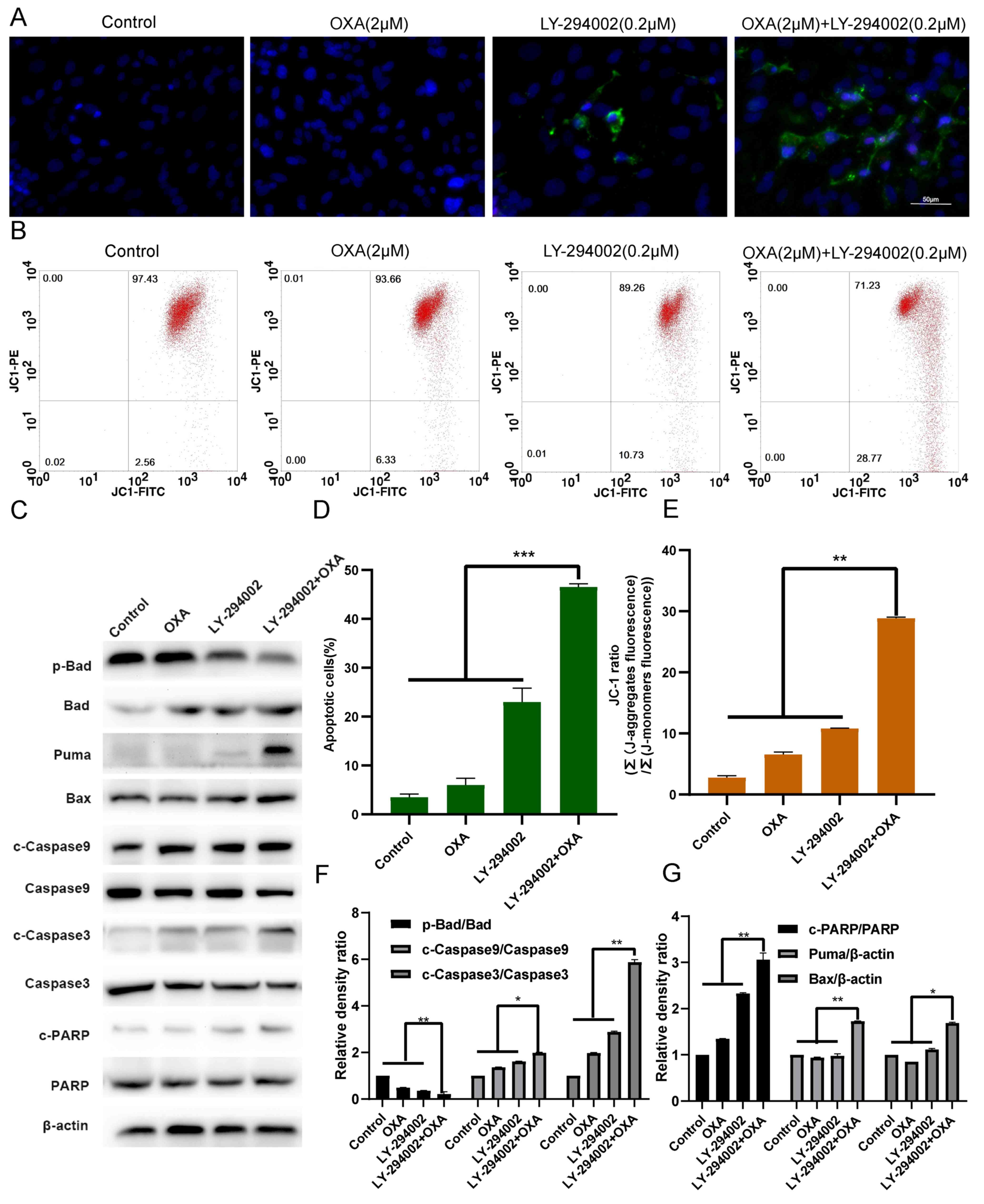 | Figure 5.LY-294002 promotes HepG2R
apoptosis induced by oxaliplatin. (A and D) LY-294002, oxaliplatin
or LY-294002 combined with oxaliplatin for 24 h. Effect of
LY-294002 on the apoptosis of liver cancer cells induced by
oxaliplatin was determined with Annexin V FITC/PI staining.
Magnification, ×400. (B and E) After 24 h treatment with LY-294002,
oxaliplatin, or LY-294002 combined with oxaliplatin, the
mitochondrial membrane potential was measured using JC-1 staining
and flow cytometry. (C and F, G) After 24 h of treatment with
LY-294002, oxaliplatin or LY-294002 combined with oxaliplatin,
western blot analysis was used to determine the effect of LY-294002
on oxaliplatin-induced apoptosis. *P<0.05, **P<0.01 and
***P<0.001. OXA, oxaliplatin; p-, phosphorylated; R,
resistant; c, cleaved. |
LY-294002 inhibits HIF-1α expression
through the PI3K/AKT signaling pathway
After HepG2 and HepG2R cells were treated
with oxaliplatin for 24 h, the expression level of HIF-1α was
increased in the HepG2 cell line, corresponding with the
oxaliplatin concentration, as illustrated in Fig. 6A and D-E. In contrast, the
expression level of HIF-1α in the HepG2R cell line was
not significantly changed with increasing concentrations of
oxaliplatin. The results of Fig. 6B and
F show that, compared with the single oxaliplatin group, the
combination of LY-294002 (0.2 µM) and oxaliplatin downregulated the
expression of HIF-1α. The results of Fig. 6C and G show that, compared with the
single oxaliplatin group, MK-2206 (50 nmol/l) combined with
oxaliplatin also downregulated the expression of HIF-1α.
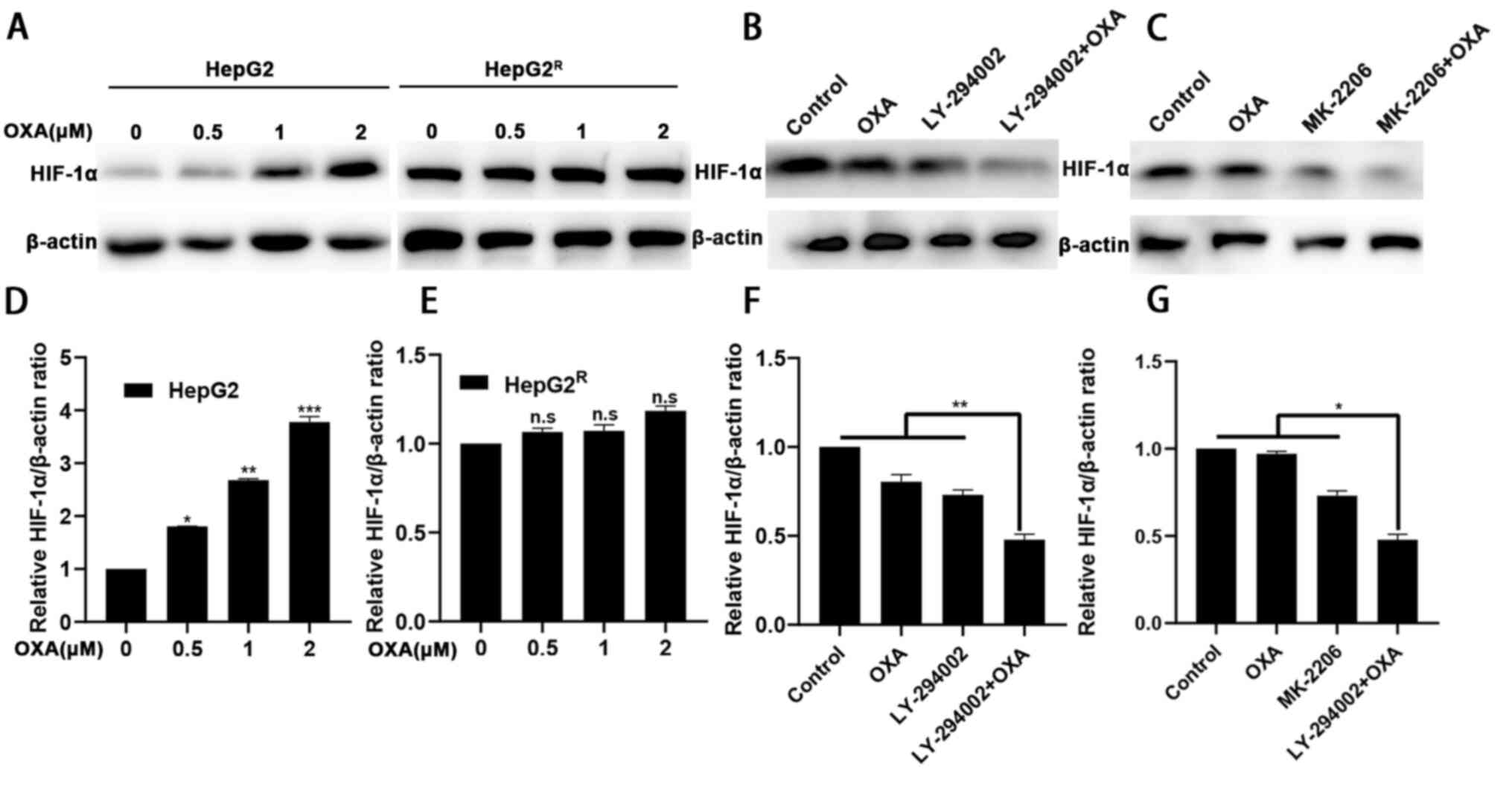 | Figure 6.LY-294002 downregulates the
expression level of HIF-1α through the PI3K/AKT signaling pathway.
(A and D, E) HepG2 and HepG2R cells were incubated with
various concentrations of oxaliplatin for 24 h. Cell lysates were
collected, and the designated proteins were detected using western
blotting. (B and F) After 24-h treatment with LY-294002,
oxaliplatin or LY-294002 combined with oxaliplatin, western blot
analysis was performed to determine the expression level of HIF-1α.
(C and G) After 24 h treatment with MK-2206, oxaliplatin, or
MK-2206 combined with oxaliplatin, western blot analysis was used
to determine that MK-2206 downregulated the expression level of
HIF-1α combined with oxaliplatin. *P<0.05, **P<0.01 vs. 0 µM
group or as indicated. OXA, oxaliplatin; n.s, not significant; HIF,
hypoxia-inducible factor; R, resistant. |
Discussion
Systemic chemotherapy is the main treatment of liver
cancer in its middle and late stages (30). For the treatment of liver cancer,
oxaliplatin has been approved as an effective systemic
chemotherapeutic agent (31,32).
Emerging resistance of oxaliplatin during chemotherapy; however,
has become a problem in the treatment of liver cancer (33,34).
The PI3K/AKT pathway is essential in a series of signal
transduction cascades that regulate cell survival and apoptosis.
Abnormal upregulation of the PI3K/AKT signaling pathway are
observed in autoimmune and infectious diseases such as human
immunodeficiency virus (HIV) and Covid-19. The PI3K/AKT signaling
pathway plays an important role in regulating the progression of
HIV and Covid-19 (35–38). In addition, abnormal upregulation of
PI3K/AKT signaling pathway usually occurs in cancer, including
liver cancer (39). For these
reasons, the present study investigated the PI3K/AKT signaling
pathway as a potential target for cancer treatment. PI3K/AKT/HIF-1α
can reportedly induce chemoresistance of liver cancer cells
(26,27). The current experimental results in
HepG2R cells indicated abnormal activation of the
PI3K/AKT/HIF-1α signaling pathway, which is consistent with
previous research results (40). In
the present study, the PI3K specific inhibitor LY-294002 was used,
which can downregulate the level of PI3K/AKT/HIF1α alienation.
LY-294002 can enhance the chemosensitivity of liver cancer
(28). Therefore, the present use
of LY-294002 in combination with oxaliplatin was well-founded. In
the current study, the PI3K/AKT signaling pathway in
HepG2R cells was in an abnormally activated state.
LY-294002 and MK-2206 downregulated the activation of the pathway,
with LY-294002 having a stronger effect compared with that of
MK-2206. In addition, compared with MK-2206, LY-294002 more
significantly inhibited the expression level of HIF-1α, which is
highly expressed in HepG2R cells, reduced cell
viability, promoted apoptosis, blocked the cell cycle and increased
the sensitivity of the cells to oxaliplatin.
In various liver cancer cell lines, oxaliplatin
activates the PI3K/AKT signaling pathway, which participates in the
most important chemotherapy resistance mechanism by inhibiting
apoptosis and promoting cell survival (41–43).
We previously reported that oxaliplatin induces abnormal activation
of the PI3K/AKT pathway to reduce the sensitivity of liver cancer
cells to oxaliplatin, which is evidence of the importance of the
PI3K/AKT pathway in anti-chemotherapy (44). The current study reported that the
PI3K/AKT signaling pathway was stably activated in
HepG2R cells, which may be the main reason for the
resistance of liver cancer cells to oxaliplatin. MK-2206 or
LY-294002 reduced the expression levels of p-AKT (Ser473) and
PI3Kp85 and inhibited the proliferation of HepG2R;
LY-294002 was more effective compared with MK-2206 in this
activity. It was also demonstrated that inhibiting the abnormal
activation of the PI3K/AKT signaling pathway increased the
sensitivity of HepG2R to oxaliplatin.
Activation of the PI3K/AKT signaling pathway can
promote the proliferation of liver cancer cells by promoting the
progression of the cell cycle (45). Activation of the pathway also can
activate the phosphorylation of the downstream effector molecules
S6K1 and elF4EBP1, which is conducive to protein synthesis and
proliferation of liver cancer cells (46). On the other hand, activation of the
PI3K/AKT signaling pathway inhibits apoptosis by phosphorylation
and inactivation of several targets (including Bad, c-Raf and
caspase-9), leading to prolonged cell survival (47). Compared with LY-294002 or
oxaliplatin alone, LY-294002 combined with oxaliplatin more
effectively inhibited the proliferation and survival of
HepG2R. In addition, the abnormal activation of the
PI3K/AKT signaling pathway participates in cell cycle regulation by
preventing GSK-3β-mediated phosphorylation and cyclin D1 and p-Rb
degradation, and negatively regulates the expression levels of
cyclin-dependent kinase inhibitors p27 and p21 (48). The present data revealed that the
combination of LY-294002 and oxaliplatin can downregulate cyclin D1
and p-Rb and arrest liver cancer cells in the
G0/G1 phase. This finding supports the view
that LY-294002 enhances the ability of oxaliplatin to inhibit liver
cancer proliferation.
Caspases are a family of cysteine proteases that are
the central regulator of apoptosis. Promoter caspases (including
caspase-2, −8, −9, −10, −11 and −12) are tightly integrated with
pro-apoptotic signals. Once activated, these caspases cleave and
activate downstream effector caspases (including caspase-3, −6, and
−7), which in turn cleave cell proteins at specific Asp residues,
thereby performing apoptosis. Members of the Bcl-2 family of
proteins can protect the integrity of mitochondria by preventing
damaged mitochondria from releasing cytochrome-c and the subsequent
combination with caspase-9 to form a complex, resulting in the
activation of caspase-9 (49,50).
Abnormal activation of the PI3K/AKT signaling pathway can inhibit
the activation of caspase-9 and activate pro-apoptotic Bcl-2 family
members (including Bad, Bax, GSK-3 and FoxO1), resulting in
anti-apoptosis (51,52). The current data showed that
LY-294002 downregulated the activation of PI3K/AKT signaling
pathway, upregulated the expression levels of Bad, Puma, Bax,
cleaved caspase-9 and −3 and c-PARP, promoted apoptosis and
increased the chemical sensitivity of liver cancer to oxaliplatin.
Therefore, oxaliplatin will activate the PI3K/AKT signaling pathway
when treating live cancer. HepG2R cells resistance to
oxaliplatin will cause abnormal activation of the PI3K/AKT
signaling pathway. LY-294002 downregulated the abnormal activation
level of PI3K/AKT signaling pathway, and improved the chemical
sensitivity of HepG2R to oxaliplatin. It was suspected
that the combination with oxaliplatin is more effective compared
with single drug treatment. The effects at the cell level have also
confirmed the reliability of the conjecture, such as reducing cell
viability, promoting apoptosis and blocking cell cycle progression.
LY-294002 enhances the toxicity of oxaliplatin to HepG2R
and plays a synergistic effect when combined with oxaliplatin.
HIF-1α expression is upregulated in liver tumors.
HIF-1α can activate the downstream target gene VEGF, induce tumor
cells to generate blood vessels, bring oxygen and nutrients to
tumor cell and promote tumor cell proliferation (53). HIF-1α is highly expressed in
colorectal cancer tissues in the early stage of colorectal cancer
and is positively correlated with the progression of the disease
(54). In liver cancer, HIF-1α can
upregulate the expression of VEGF, cyclin D1, TGF-β, insulin-like
growth factor 2 and other growth factors, promote the proliferation
and differentiation of liver cancer cells and promote
hyperproliferation (55). HIF-1α
also initiates transcription of downstream target genes, which
increases cell proliferation and decreases apoptosis (56). Since PI3K/AKT can regulate the
expression of HIF-1α (11), the
present study used PI3K/AKT signaling pathway inhibitors to
evaluate the effect on the expression level of HIF-1α. HIF-1α was
highly expressed in HepG2R cells, while MK-2206 and
LY-294002 reduced the expression level of HIF-1α protein, with
LY-294002 having the stronger effect. However, the current study
did not explore the mutual regulation between PI3K/AKT and HIF-1α,
which will be studied in the future. The downstream mTOR molecule
of PI3K/AKT is upregulated in a variety of cancer types, including
liver cancer. mTOR molecule is phosphorylated at Ser2448 and
auto-phosphorylated at Ser2481 through the PI3K/AKT signaling
pathway. The mTOR molecule plays an important role in cell
proliferation and homeostasis and may be abnormally regulated in
tumors (57). Besides, mTOR
inhibitors have been approved for the treatment of certain types of
cancer including gastrointestinal stromal tumors and breast cancer
(58,59). However, because of time constraints,
this was not investigated in the present study; it will be
discussed in depth in our future work. In addition, lack of in
vivo experimental data due to time restrictions and other
factors mean that the reliability of the results have not been
verified at the animal level. This is the limitation of this
article and will be supplemented in the future.
In summary, the PI3K/AKT signaling pathway inhibitor
LY-294002 increased the chemical sensitivity of liver cancer to
oxaliplatin by blocking cell cycle progression, reducing cell
viability and promoting apoptosis. These effects likely were
achieved by inhibiting the abnormal activation of the PI3K/AKT
signaling pathway in the cancer cells, thus reducing the expression
of HIF-1α. These findings suggested that the combination of
LY-294002 and oxaliplatin can be an effective treatment strategy
for liver cancer.
Acknowledgements
Not applicable.
Funding
The study was funded by The National Natural Science
Fund of China (grant nos. 82071862, 81872017 and 81572431), The
University Natural Science Research Project of Anhui Province
(grant nos. KJ2018ZD011 and KJ2019A0093), The Anhui Provincial
Science and Technology program (grant nos. 202004j07020053 and
1604a0802094), Research Foundation of the Institute of
Environment-friendly Materials and Occupational Health (Wuhu),
Anhui University of Science and Technology (grant no. ALW2020YF11)
and The Huainan Science and Technology Project (grant no. 2017B41)
funded this research.
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
RYX performed the experiments, analyzed the data and
was the main contributor to writing the manuscript. YCZ analyzed
the data. AML was responsible for the English revision work and
performed some of the experiments. YFM, WPC, LS, YHX, SPZ and WYC
performed the experiments. XLT was the project leader and was
responsible for the design of the project, the revision of the
manuscript and performed some of the experiments. RYX and XLT
confirm the authenticity of all the raw data. All authors read and
approved the final manuscript.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Feng RM, Zong YN, Cao SM and Xu RH:
Current cancer situation in China: Good or bad news from the 2018
Global cancer statistics? Cancer Commun (Lond). 39:222019.
View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Miller KD, Goding Sauer A, Ortiz AP,
Fedewa SA, Pinheiro PS, Tortolero-Luna G, Martinez-Tyson D, Jemal A
and Siegel RL: Cancer statistics for Hispanics/Latinos, 2018. CA
Cancer J Clin. 68:425–445. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Li A, Zhang R, Zhang Y, Liu X, Wang R, Liu
J, Liu X, Xie Y, Cao W, Xu R, et al: BEZ235 increases sorafenib
inhibition of hepatocellular carcinoma cells by suppressing the
PI3K/AKT/mTOR pathway. Am J Transl Res. 11:5573–5585.
2019.PubMed/NCBI
|
|
4
|
Smith DK and Murphy BA: Lower levels of
education and household income mediate lower dental care
utilization among survivors of early life cancers. Prev Med Rep.
14:1008682019. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Liu X, Xie C, Li A, Zhang Y, Liu X, Zhou
S, Shen J, Huo Z, Cao W, Ma Y, et al: BEZ235 enhances
chemosensitivity of paclitaxel in hepatocellular carcinoma through
inhibiting the PI3K/Akt/mTOR pathway. Am J Transl Res.
11:7255–7271. 2019.PubMed/NCBI
|
|
6
|
Yang Y, Yao JH, Du QY, Zhou YC, Yao TJ, Wu
Q, Liu J and Ou YR: Connexin 32 downregulation is critical for
chemoresistance in oxaliplatin-resistant HCC cells associated with
EMT. Cancer Manag Res. 11:5133–5146. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Ye JZ, Yan SM, Yuan CL, Wu HN, Zhang JY,
Liu ZH, Li YQ, Luo XL, Lin Y and Liang R: GP73 level determines
chemotherapeutic resistance in human hepatocellular carcinoma
cells. J Cancer. 9:415–423. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Zhu YJ, Zheng B, Wang HY and Chen L: New
knowledge of the mechanisms of sorafenib resistance in liver
cancer. Acta Pharmacol Sin. 38:614–622. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Zhang Y, Liu X, Zhang J, Xu Y, Shao J, Hu
Y, Shu P and Cheng H: Inhibition of miR-19a partially reversed the
resistance of colorectal cancer to oxaliplatin via PTEN/PI3K/AKT
pathway. Aging (Albany NY). 12:5640–5650. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Kang HG, Wang BZ, Zhang J, Liu MR and Li
YX: Combination of temsirolimus and Adriamycin exhibits an enhanced
antitumor effect in hepatocellular carcinoma. Clin Res Hepatol
Gastroenterol. 41:197–203. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Sun X, Su Y, He Y, Zhang J, Liu W, Zhang
H, Hou Z, Liu J and Li J: New strategy for in vitro activation of
primordial follicles with mTOR and PI3K stimulators. Cell Cycle.
14:721–731. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Fruman DA, Chiu H, Hopkins BD, Bagrodia S,
Cantley LC and Abraham RT: The PI3K pathway in human disease. Cell.
170:605–635. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Aoki M and Fujishita T: Oncogenic roles of
the PI3K/AKT/mTOR axis. Curr Top Microbiol Immunol. 407:153–189.
2017.PubMed/NCBI
|
|
14
|
Grosbois J and Demeestere I: Dynamics of
PI3K and Hippo signaling pathways during in vitro human follicle
activation. Hum Reprod. 33:1705–1714. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Chang L, Graham PH, Ni J, Hao J, Bucci J,
Cozzi PJ and Li Y: Targeting PI3K/Akt/mTOR signaling pathway in the
treatment of prostate cancer radioresistance. Crit Rev Oncol
Hematol. 96:507–517. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Brown JS and Banerji U: Maximising the
potential of AKT inhibitors as anti-cancer treatments. Pharmacol
Ther. 172:101–115. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Mehal W: NASH and HCC are driven by
different signaling pathways with a common regulator. Cell Metab.
29:3–4. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Lee JH, Lee HJ, Sim DY, Jung JH, Kim KR
and Kim SH: Apoptotic effect of lambertianic acid through
AMPK/FOXM1 signaling in MDA-MB231 breast cancer cells. Phytother
Res. 32:1755–1763. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Laughner E, Taghavi P, Chiles K, Mahon PC
and Semenza GL: HER2 (neu) signaling increases the rate of
hypoxia-inducible factor 1alpha (HIF-1alpha) synthesis: Novel
mechanism for HIF-1-mediated vascular endothelial growth factor
expression. Mol Cell Biol. 21:3995–4004. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Qu D, Weygant N, Yao J, Chandrakesan P,
Berry WL, May R, Pitts K, Husain S, Lightfoot S, Li M, et al:
Overexpression of DCLK1-AL increases tumor cell invasion, drug
resistance, and KRAS activation and can be targeted to inhibit
tumorigenesis in pancreatic cancer. J Oncol. 2019:64029252019.
View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Simioni C, Martelli AM, Cani A,
Cetin-Atalay R, McCubrey JA, Capitani S and Neri LM: The AKT
inhibitor MK-2206 is cytotoxic in hepatocarcinoma cells displaying
hyperphosphorylated AKT-1 and synergizes with conventional
chemotherapy. Oncotarget. 4:1496–1506. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Yu X, Liu J, Qiu H, Hao H, Zhu J and Peng
S: Combined inhibition of ACK1 and AKT shows potential toward
targeted therapy against KRAS-mutant non-small-cell lung cancer.
Bosn J Basic Med Sci. 21:198–207. 2021.PubMed/NCBI
|
|
23
|
Li YL, Weng HC, Hsu JL, Lin SW, Guh JH and
Hsu LC: The combination of MK-2206 and WZB117 exerts a synergistic
cytotoxic effect against breast cancer cells. Front Pharmacol.
10:13112019. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Zhu Y, Zhong Y, Long X, Zhu Z, Zhou Y, Ye
H, Zeng X and Zheng X: Deoxyshikonin isolated from Arnebia
euchroma inhibits colorectal cancer by down-regulating the
PI3K/Akt/mTOR pathway. Pharm Biol. 57:412–423. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Wang X, Wang X, Xu Y, Yan M, Li W, Chen J
and Chen T: Effect of nicastrin on hepatocellular carcinoma
proliferation and apoptosis through PI3K/AKT signalling pathway
modulation. Cancer Cell Int. 20:912020. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Jiao M and Nan KJ: Activation of PI3
kinase/Akt/HIF-1α pathway contributes to hypoxia-induced
epithelial-mesenchymal transition and chemoresistance in
hepatocellular carcinoma. Int J Oncol. 40:461–468. 2012.PubMed/NCBI
|
|
27
|
Ling S, Li J, Shan Q, Dai H, Lu D, Wen X,
Song P, Xie H, Zhou L, Liu J, et al: USP22 mediates the multidrug
resistance of hepatocellular carcinoma via the SIRT1/AKT/MRP1
signaling pathway. Mol Oncol. 11:682–695. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Ma J, Xie SL, Geng YJ, Jin S, Wang GY and
Lv GY: In vitro regulation of hepatocellular carcinoma cell
viability, apoptosis, invasion, and AEG-1 expression by LY294002.
Clin Res Hepatol Gastroenterol. 38:73–80. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Elefantova K, Lakatos B, Kubickova J,
Sulova Z and Breier A: Detection of the mitochondrial membrane
potential by the cationic Dye JC-1 in L1210 cells with massive
overexpression of the plasma membrane ABCB1 drug transporter. Int J
Mol Sci. 19:19852018. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Tang X, Li A, Xie C, Zhang Y, Liu X, Xie
Y, Wu B, Zhou S, Huang X, Ma Y, et al: The PI3K/mTOR dual inhibitor
BEZ235 nanoparticles improve radiosensitization of hepatoma cells
through apoptosis and regulation DNA repair pathway. Nanoscale Res
Lett. 15:632020. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Devanabanda B and Kasi A: Oxaliplatin.
StatPearls. Treasure Island (FL): StatPearls Publishing Copyright©
2020, StatPearls Publishing LLC; 2020
|
|
32
|
Tang X, Chen L, Li A, Cai S, Zhang Y, Liu
X, Jiang Z, Liu X, Liang Y and Ma D: Anti-GPC3 antibody-modified
sorafenib-loaded nanoparticles significantly inhibited HepG2
hepatocellular carcinoma. Drug Deliv. 25:1484–1494. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Shen JH, Chen PH, Liu HD, Huang DA, Li MM
and Guo K: HSF1/AMPKα2 mediated alteration of metabolic phenotypes
confers increased oxaliplatin resistance in HCC cells. Am J Cancer
Res. 9:2349–2363. 2019.PubMed/NCBI
|
|
34
|
Liao X, Song G, Xu Z, Bu Y, Chang F, Jia
F, Xiao X, Ren X, Zhang M and Jia Q: Oxaliplatin resistance is
enhanced by saracatinib via upregulation Wnt-ABCG1 signaling in
hepatocellular carcinoma. BMC Cancer. 20:312020. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
He J, Ma J, Ren B and Liu A: Advances in
systemic lupus erythematosus pathogenesis via mTOR signaling
pathway. Semin Arthritis Rheum. 50:314–320. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Mammana S, Bramanti P, Mazzon E, Cavalli
E, Basile MS, Fagone P, Petralia MC, McCubrey JA, Nicoletti F and
Mangano K: Preclinical evaluation of the PI3K/Akt/mTOR pathway in
animal models of multiple sclerosis. Oncotarget. 9:8263–8277. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Nicoletti F, Fagone P, Meroni P, McCubrey
J and Bendtzen K: mTOR as a multifunctional therapeutic target in
HIV infection. Drug Discov Today. 16:715–721. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Fagone P, Ciurleo R, Lombardo SD,
Iacobello C, Palermo CI, Shoenfeld Y, Bendtzen K, Bramanti P and
Nicoletti F: Transcriptional landscape of SARS-CoV-2 infection
dismantles pathogenic pathways activated by the virus, proposes
unique sex-specific differences and predicts tailored therapeutic
strategies. Autoimmun Rev. 19:1025712020. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Gong C, Ai J, Fan Y, Gao J, Liu W, Feng Q,
Liao W and Wu L: NCAPG promotes the proliferation of hepatocellular
carcinoma through PI3K/AKT signaling. Onco Targets Ther.
12:8537–8552. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Xie Y and Zhong DW: AEG-1 is associated
with hypoxia-induced hepatocellular carcinoma chemoresistance via
regulating PI3K/AKT/HIF-1alpha/MDR-1 pathway. EXCLI J. 15:745–757.
2016.PubMed/NCBI
|
|
41
|
Li XZ, Sun YL, Cao LQ and Li MJ:
Oxaliplatin-rapamycin combination was superior to mono-drug in
treatment of hepatocellular carcinoma both in vitro and in vivo.
Neoplasma. 63:880–887. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Liao B, Zhang Y, Sun Q and Jiang P:
Vorinostat enhances the anticancer effect of oxaliplatin on
hepatocellular carcinoma cells. Cancer Med. 7:196–207. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Fu X, Wen H, Jing L, Yang Y, Wang W, Liang
X, Nan K, Yao Y and Tian T: MicroRNA-155-5p promotes hepatocellular
carcinoma progression by suppressing PTEN through the PI3K/Akt
pathway. Cancer Sci. 108:620–631. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Zhang Y, Xie C, Li A, Liu X, Xing Y, Shen
J, Huo Z, Zhou S, Liu X, Xie Y, et al: PKI-587 enhances
chemosensitivity of oxaliplatin in hepatocellular carcinoma through
suppressing DNA damage repair pathway (NHEJ and HR) and
PI3K/AKT/mTOR pathway. Am J Transl Res. 11:5134–5149.
2019.PubMed/NCBI
|
|
45
|
Wang Y, Nie H, Zhao X, Qin Y and Gong X:
Bicyclol induces cell cycle arrest and autophagy in HepG2 human
hepatocellular carcinoma cells through the PI3K/AKT and
Ras/Raf/MEK/ERK pathways. BMC Cancer. 16:7422016. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Li TT, Zhu D, Mou T, Guo Z, Pu JL, Chen
QS, Wei XF and Wu ZJ: IL-37 induces autophagy in hepatocellular
carcinoma cells by inhibiting the PI3K/AKT/mTOR pathway. Mol
Immunol. 87:132–140. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Zhu F, Jiang D, Zhang M and Zhao B:
2,4-Dihydroxy- 3′-methoxy-4′-ethoxychalcone suppresses cell
proliferation and induces apoptosis of multiple myeloma via the
PI3K/akt/mTOR signaling pathway. Pharm Biol. 57:641–648. 2019.
View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Fang X, Yang D, Luo H, Wu S, Dong W, Xiao
J, Yuan S, Ni A, Zhang KJ, Liu XY and Chu L: SNORD126 promotes HCC
and CRC cell growth by activating the PI3K-AKT pathway through
FGFR2. J Mol Cell Biol. 9:243–255. 2017.PubMed/NCBI
|
|
49
|
Zhou M, Zhang Q, Zhao J, Liao M, Wen S and
Yang M: Phosphorylation of Bcl-2 plays an important role in
glycochenodeoxycholate-induced survival and chemoresistance in HCC.
Oncol Rep. 38:1742–1750. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Pan W, Li W, Zhao J, Huang Z, Zhao J, Chen
S, Wang C, Xue Y, Huang F, Fang Q, et al: lncRNA-PDPK2P promotes
hepatocellular carcinoma progression through the PDK1/AKT/Caspase 3
pathway. Mol Oncol. 13:2246–2258. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Jiang S, Wang Q, Feng M, Li J, Guan Z, An
D, Dong M, Peng Y, Kuerban K and Ye L: C2-ceramide enhances
sorafenib-induced caspase-dependent apoptosis via PI3K/AKT/mTOR and
Erk signaling pathways in HCC cells. Appl Microbiol Biotechnol.
101:1535–1546. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Sui Y, Zheng X and Zhao D: Rab31 promoted
hepatocellular carcinoma (HCC) progression via inhibition of cell
apoptosis induced by PI3K/AKT/Bcl-2/BAX pathway. Tumour Biol.
36:8661–8670. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
53
|
Mendez-Blanco C, Fondevila F,
Garcia-Palomo A, Gonzalez-Gallego J and Mauriz JL: Sorafenib
resistance in hepatocarcinoma: Role of hypoxia-inducible factors.
Exp Mol Med. 50:1–9. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
54
|
Gong J, Zhou S and Yang S: Vanillic Acid
Suppresses HIF-1α expression via inhibition of mTOR/p70S6K/4E-BP1
and Raf/MEK/ERK pathways in human colon cancer HCT116 cells. Int J
Mol Sci. 20:4652019. View Article : Google Scholar : PubMed/NCBI
|
|
55
|
Xu LF, Ni JY, Sun HL, Chen YT and Wu YD:
Effects of hypoxia-inducible factor-1α silencing on the
proliferation of CBRH-7919 hepatoma cells. World J Gastroenterol.
19:1749–1759. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
56
|
Chen J, Bai M, Ning C, Xie B, Zhang J,
Liao H, Xiong J, Tao X, Yan D, Xi X, et al: Gankyrin facilitates
follicle-stimulating hormone-driven ovarian cancer cell
proliferation through the PI3K/AKT/HIF-1α/cyclin D1 pathway.
Oncogene. 35:2506–2517. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
57
|
Ferrin G, Guerrero M, Amado V,
Rodriguez-Peralvarez M and De la Mata M: Activation of mTOR
signaling pathway in hepatocellular carcinoma. Int J Mol Sci.
21:12662020. View Article : Google Scholar : PubMed/NCBI
|
|
58
|
Duan Y, Haybaeck J and Yang Z: Therapeutic
potential of PI3K/AKT/mTOR pathway in gastrointestinal stromal
tumors: Rationale and progress. Cancers (Basel). 12:29722020.
View Article : Google Scholar : PubMed/NCBI
|
|
59
|
Steelman LS, Martelli AM, Cocco L, Libra
M, Nicoletti F, Abrams SL and McCubrey JA: The therapeutic
potential of mTOR inhibitors in breast cancer. Br J Clin Pharmacol.
82:1189–1212. 2016. View Article : Google Scholar : PubMed/NCBI
|