Introduction
Spontaneous intracerebral hemorrhage (ICH) has the
highest mortality rate among stroke subtypes, accounts for 15–20%
of all stroke types, and has an increased incidence in elderly
patients (1–3). Acute ICH due to a large intracranial
hematoma is associated with high morbidity and mortality, as it can
lead to primary brain injury through the destruction of brain
tissue and the high intracranial pressure (ICP) that results from
the large hematoma (4,5). Previous studies revealed that
craniotomy for hematoma evacuation is an effective therapy for
limiting primary brain damage and decreasing ICP following ICH,
which is of substantial interest (4,6,7).
However, craniotomy for hematoma evacuation shows no clinical
benefit to patients, no improvement in long-term outcomes, and
rarely affects neurological recovery (8). Increasing evidence shows that red
blood cell debris, hemoglobin, its degradation products, and blood
components trigger secondary brain injury following ICH and
contribute to a series of damaging events, including
neuroinflammation, brain edema, oxidative stress, blood-brain
barrier damage, and neuronal death (9–14). In
recent years, an increasing number of studies have been conducted
with a focus on the mechanisms underlying ICH-induced secondary
injury to investigate better therapeutic targets for ICH (6,15).
A previous study has reported that neuroinflammation
has a key role in the progression of ICH-induced brain injury and
is characterized by an innate immune response and subsequent
inflammatory cytokine production (16). The neuroinflammatory mechanisms
include activation of microglia, the resident macrophages in the
brain, and other blood-derived immune cells (9). Activated microglia/macrophages and
peripheral blood-derived immune cells release proinflammatory
cytokines, including tumor necrosis factor (TNF-) α, interleukin
(IL-) 1β, chemokines, free radicals, and other toxic chemicals,
which ultimately leads to brain edema and neuronal death (17,18).
ICH-induced neuronal death was initially divided into two different
categories, necrosis and apoptosis. Necroptosis, a recently
discovered form of necrosis, is a caspase-independent mechanism of
programmed cell death and serves a vital role in neuronal cell
death following ICH (19,20). Previous studies have demonstrated
that necroptosis inhibition can significantly improve brain injury
outcomes after ICH (21,22). Therefore, targeting
neuroinflammation and necroptosis has become a major goal of ICH
treatment.
Glutamate is the most important and widely
distributed excitatory neurotransmitter in the brain and
predominantly binds to
alpha-amino-3-hydroxy-5-methyl-4-isoxazolepropionic acid (AMPA),
kainate, and N-methyl-D-aspartate ionotropic receptors (23). Overexpression of glutamate and
overactivation of its receptors contribute to neuroinflammation,
tissue damage, or neuronal death following ICH, traumatic brain
injury, or acute and chronic pain (24,25).
Perampanel is a noncompetitive antagonist of AMPA receptor (AMPAR),
a subtype of ionotropic glutamate receptor (26). Previous studies have revealed that
perampanel can exert neuroprotective effects and improve brain
injury outcomes after traumatic brain injury, and its mechanisms of
action may be related to neuronal necroptosis and antioxidative and
anti-inflammatory activity (24,27,28).
However, the effects of perampanel on ICH in vitro or in
vivo remain unclear, and whether the underlying molecular
mechanisms involve neuronal necroptosis and neuroinflammation has
not been investigated. Wang et al (29) reported that the PTEN signaling
pathway was crucial for regulating cell death by necroptosis and
apoptosis. Guido et al (30)
also demonstrated that the estrogen receptor (ER)β/PTEN signaling
pathway induced cell death in TCAM2 cells by autophagy and
necroptosis. In a subarachnoid hemorrhage model, mesenchymal stem
cell-derived extracellular vesicles promoted neuronal survival and
alleviated early brain injury via the PTEN/AKT pathway (31).
The present study therefore investigated the effects
of perampanel on ICH in vitro and in vivo, and
explored its potential molecular mechanisms with a focus on the
PTEN signaling pathway.
Materials and methods
Animals
All animal experiments performed for the present
study complied with the National Institutes of Health guidelines
for the handling of laboratory animals and were approved by the
Ethics Committee of the Wuxi Medical College of Anhui Medical
University (Wuxi, China). All experiments were conducted on healthy
adult male C57BL/6J mice (22–25 g; age, 8–12-weeks; Anhui Medical
University, Hefei, China). The mice were housed in animal care
facilities with a 12-h light/dark cycle and had free access to food
and water.
ICH animal model
The ICH mouse model was generated based on a
previously described protocol involving autologous blood injection
(32). Briefly, male C57BL6/J mice
were anesthetized by intraperitoneal (i.p.) injection of 50 mg/kg
pentobarbital sodium and placed in a prone position with a
stereotactic head frame. The rectal temperature was kept at
37.0±0.5°C during the operation using a heating pad. An artificial
tear ointment was used to protect the eyes from injury during
surgery. A midline scalp incision was made, and a cranial burr hole
with a 1-mm diameter was made at the following coordinates relative
to bregma: 0.2 mm posterior, 2.2 mm lateral to bregma, and 3.5 mm
below the dura. A total of 30 µl of autologous blood without
anticoagulation was collected from the caudal artery and rapidly
injected into the basal ganglia through the burr hole via the
26-gauge needle of a 10-µl Hamilton syringe. First, 5 µl of
arterial blood was injected at a depth of 2.8 mm from the dura
(injection speed, 3 µl/min). Five minutes later, the other 25 µl
blood was injected at a depth of 3.5 mm (injection speed, 3
µl/min). After the injection of autologous blood, the needle was
kept in the brain for 10 min to prevent blood backflow along the
needle tract. Finally, the hole was covered with medical bone wax.
The animals in the Sham group received similar surgical procedures
but were injected at the same site with an equal volume of sterile
saline instead of blood. At 72 h following ICH, the mice were
sacrificed with 100 mg/kg sodium pentobarbital via i.p. injection.
Death was confirmed by observing respiration and by using the
corneal reflection method. Brain tissue samples were collected
after the mice were sacrificed. Fresh specimens were stored in
liquid nitrogen (−196°C) or 4% formalin-fixed (4°C) for ≥48 h.
Drug administration
Perampanel was purchased from Santa Cruz
Biotechnology, Inc. (cat. no. sc-477647). The mice were orally
administered with saline water or perampanel (5 mg/kg) for 24 h
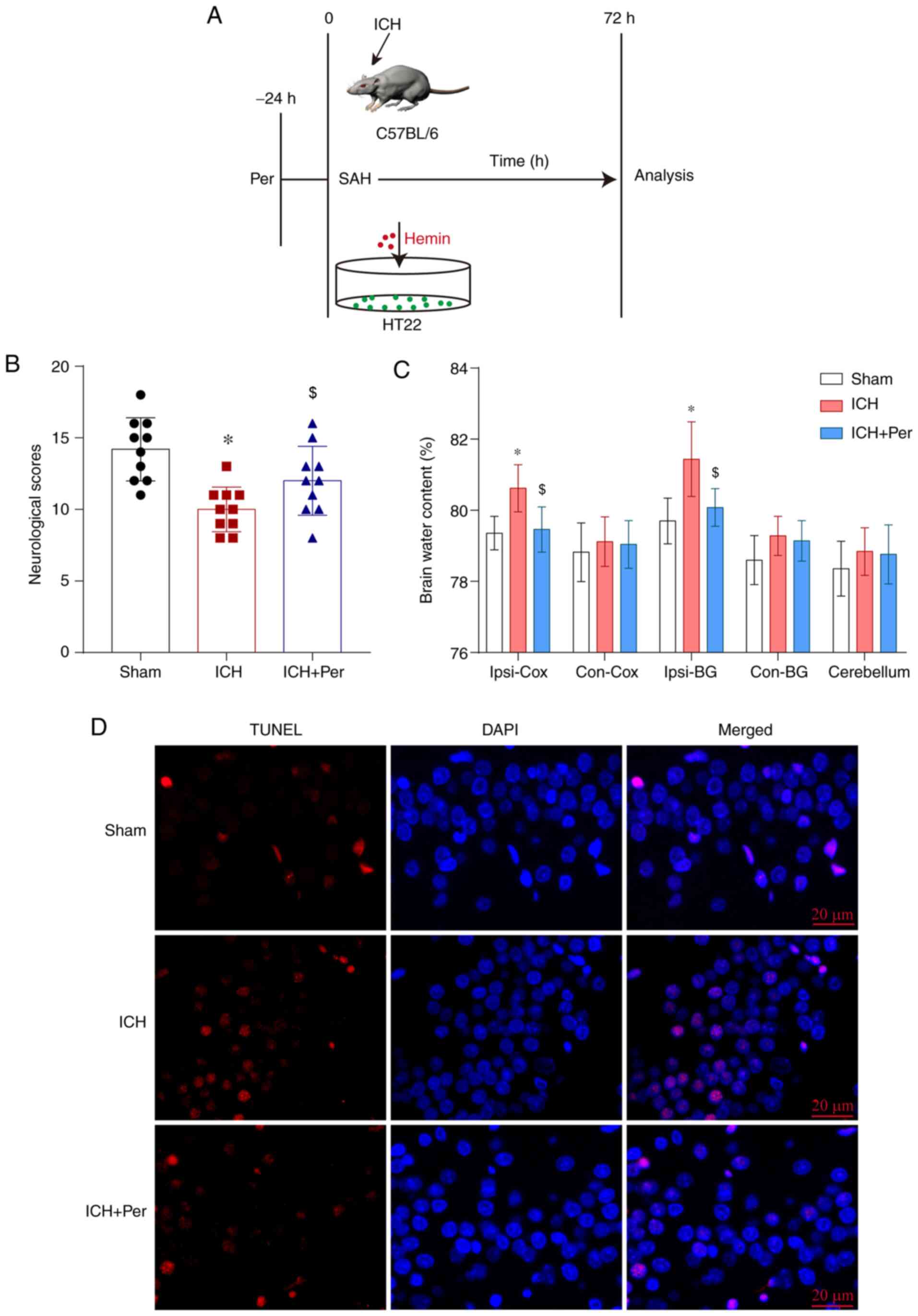
before ICH (Fig. 1A). HT22 cells
(Procell Life Science & Technology Co., Ltd.) were cultured in
Dulbecco's modified Eagle's medium containing 10% fetal bovine
serum and 1% penicillin/streptomycin and until 70% confluent at
37°C with 5% CO2, then were treated with perampanel (2
mmol/l) for 24 h prior to hemin insult (Fig. 1A). z-VAD-fmk (cat. no. 187389-52-2),
3-Methyladenine (3-MA; cat. no. 5142-23-4), and Necrostatin-1 (cat.
no. 4311-88-0) were purchased from Selleck Chemicals.
Small interfering RNA (siRNA)
treatment and transfection
Mice were anesthetized with pentobarbital sodium and
placed on stereotactic apparatus. Then, a burr hole was made in the
left hemisphere using the following coordinates: 0.2 mm posterior,
1 mm lateral, and 2.2 mm below the horizontal plane of the bregma.
Then, 5 µl (4 nM/µl) siRNA molecules were injected into the left
lateral ventricle at a rate of 0.5 µl/min. To enhance the silencing
effect, the injection was performed 48 h prior to ICH.
For in vitro cell transfection, Lipofectamine
RNAiMax reagent (Invitrogen; Thermo Fisher Sceintific, Inc.) was
used in Opti-MEM medium, according to the manufacturer's
instructions. The transfection was performed at 37°C in a
humidified incubator with 5% CO2. Cells were transfected
with siRNA at a concentration of 50 nM. Cells were incubated with
siRNA and Lipofectamine® 2000 for 6 h in transfection
medium. Cells were harvested for further studies 72 h after the
beginning of transfection. The following targeted and control
siRNAs were synthesized by JiKai Biotechnology, Inc.: si-PTEN
(Cy3-labeled siR-996) sense, 5′-GGUGUAUACAGGAACAAUATT-3′ and
antisense, 5′-UAUUGUUCCUGUAUACACCTT-3′; and negative control siRNA
sense, 5′-UUCUCCGAACGUGUCACGUTT-3′ and antisense,
5′-ACGUGACACGUUCGGAGAATT-3′.
Neurobehavioral assessment
The severity of brain injury was determined by
evaluating the neurological function of mice at 72 h after ICH,
using our previously described neurological grading system
(33). The scoring system consisted
of six tests, and the specific standards are listed in the Table SI. The neurological scores ranged
from 3 to 18 and included spontaneous activities (0–3), movement
symmetry of all limbs (0–3), forelimb outstretching (0–3), body
proprioception (1–3), response to vibrissae touch (1–3), and
climbing (1–3). All mice from every group received a
behavioral assessment, and a higher score represented better
neurological function.
Brain water content measurement
The severity of brain edema was evaluated by
measuring the brain water content using the standard wet-dry
method, as previously reported (15,33,34).
The mice were sacrificed 72 h following ICH, and the entire brain
was harvested and separated into the ipsilateral and contralateral
cortex, ipsilateral and contralateral basal ganglia, and cerebellum
(wet weight). Then, brain specimens from each group were dehydrated
at 105°C for 24 h to acquire the dry weight. The percentage of
brain water content was calculated using the following formula:
(wet weight-dry weight)/wet weight ×100.
TdT-mediated dUTP-biotin nick end
labeling (TUNEL) assay
A TUNEL assay was conducted to assess neuronal death
in the brain cortex and were fixed with 4% paraformaldehyde for 1 h
at 25°C. Paraffin-embedded sections (10 µm) were cut from
formalin-fixed tissue and stained with TUNEL staining. Specimens
were examined under a light microscope (magnification, ×200). TUNEL
reaction mixture (50 µl) was added to each sample, and the slides
were incubated in a humidified dark chamber for 60 min at 37°C. The
slides were then incubated with DAPI for 5 min at room temperature
in the dark to stain the nuclei, followed by imaging with a
fluorescence microscope. The procedure was performed according to
the manufacturer's instructions with a TUNEL staining kit. A
negative control (without the TUNEL reaction mixture) was used. The
apoptotic index (%) was the ratio of the number of TUNEL-positive
cells/total number of cells ×100. The cell count was confirmed in
four randomly selected high-power fields, and the data obtained
from each field were averaged.
Cytokine measurements of ipsilateral
cortex tissue
The levels of IL-1β (cat. no. ab197742; Abcam), IL-6
(cat. no. ab222503; Abcam), TNF-α (cat. no. ab208348; Abcam), and
NF-κB (cat. no. ab176663; Abcam) were measured by ELISA, according
to the manufacturer's instructions.
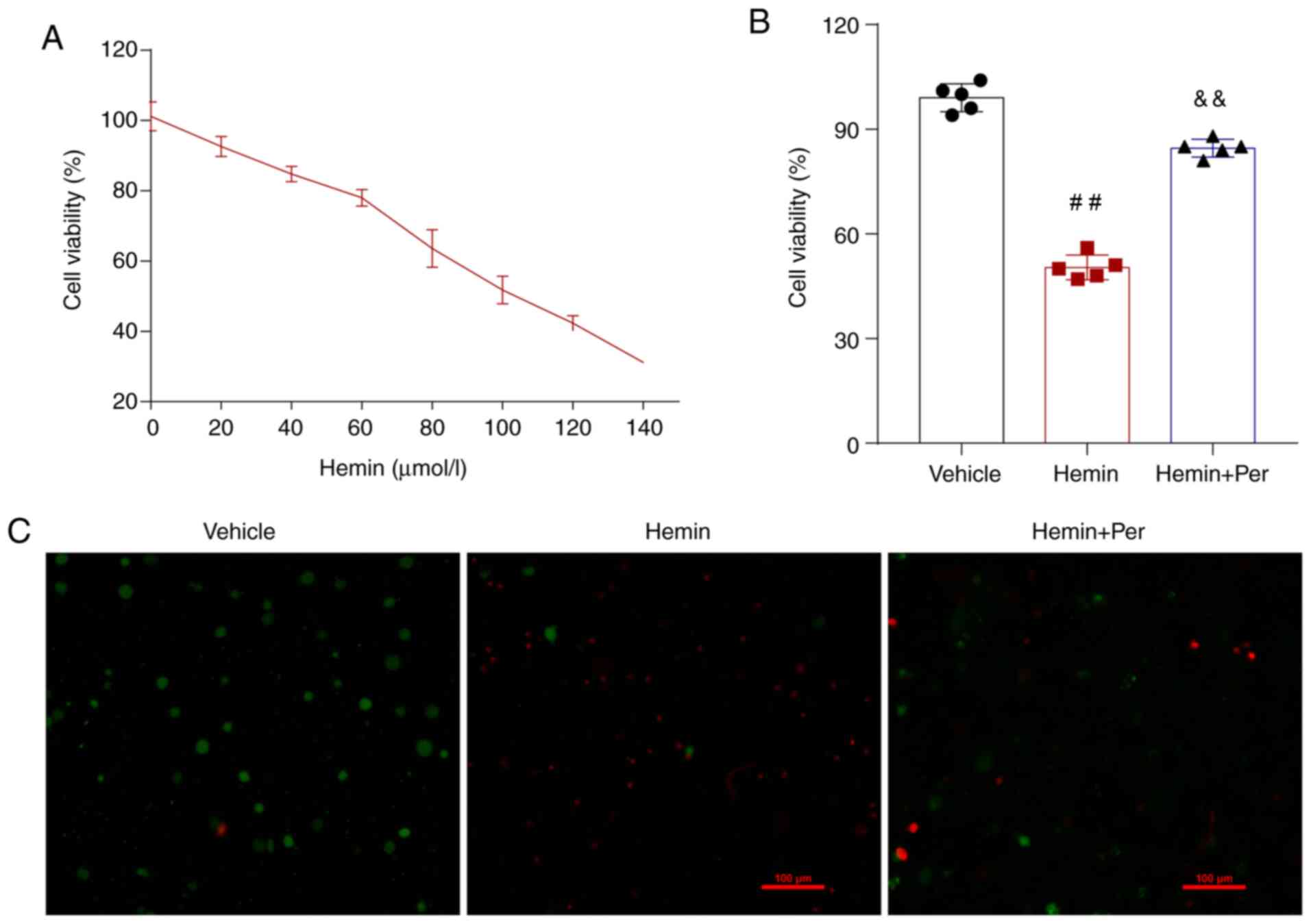
In vitro ICH model in HT22 cells
The hemin-induced ICH method was used to construct
the in vitro cell ICH model as previously described
(35). For in vitro ICH
experiments, cell death was induced in hippocampal HT22 cells by
treatment with hemin (dose-response, 20–140 mM for 48 h;
Sigma-Aldrich; Merck KGaA; cat. no. H9039). In the present study,
cells were treated with 100 mM hemin (median lethal dose) in the
presence of sodium selenite (Sigma-Aldrich; Merck KGaA) dissolved
in ddH2O. Hemin was prepared and diluted to a final
concentration of 80 mM with culture medium and sterilized by
filtration through a 0.22-µm sterile filter. Cell viability was
analyzed 72 h after treatment. Cells were then washed in prewarmed
PBS (37°C) and assessed by an MTT assay as per the manufacturer's
instructions (Promega Corporation). As determined in a prior study
(15), neurons were stimulated with
80 mM hemin for 72 h to mimic intracranial hemorrhage
conditions.
MTT assay for cell viability
Cell viability was assessed with a
3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyl tetrazolium bromide (MTT)
assay and dead/live assay. For the MTT assay, HT22 cells were
seeded into 96-well plates, then 20 µl MTT (Sigma-Aldrich; Merck
KGaA) was added to each well and the plate was incubated at 37°C
for 4 h. The culture medium was removed, and formazan crystals were
lysed in 150 µl dimethylsulfoxide (DMSO) by gently shaking the
plate. Subsequently, the optical density was determined in a
microplate reader (Bio-Rad Laboratories, Inc.) at 490 nm.
Dead/live assay
For the dead/live assay, HT22 cells were washed
twice and resuspended in PBS, and then samples were exposed to
propidium iodide (PI; 4 µM) and calcein-AM (3 µM) for 20 min in the
dark at room temperature. Cells were washed and resuspended in PBS
and analyzed under a fluorescence microscope system (DMIL 4000B;
Leica Microsystems GmbH). Bound PI produces red fluorescence and
indicates dead cells, whereas calcein-AM produces green
fluorescence and indicates live cells. Cell viability was expressed
as a percentage relative to the values obtained from the control
cultures. All experiments were independently repeated in
triplicate.
Reverse transcription-quantitative PCR
(RT-qPCR)
RT-qPCR analysis was performed as indicated
previously (36). Total RNA was
extracted from either cell cultures or hippocampal brain samples
using TRIzol reagent (Thermo Fisher Scientific, Inc.), according to
the manufacturer's instructions. Then, RNA was reverse transcribed
to cDNA using the RevertAid First Strand cDNA Synthesis kit (cat.
no. K1622; Thermo Fisher Scientific Inc.). Receptor interacting
serine/threonine kinase (RIP) 1, RIP3, mixed lineage kinase domain
like pseudokinase (MLKL), and PTEN mRNA expression levels in each
sample were measured by qPCR using SYBR Green Master Mix (Toyobo
Life Science). GAPDH was used as an internal control. The qPCR
thermocycling conditions were as follows: 45°C for 2 min and 95°C
for 10 min, followed by 40 cycles of denaturation at 95°C for 15
sec, annealing at 60°C for 1 min, and extension at 72°C for 1 min.
The 2−ΔΔCq method was used to assess the relative RNA
expression levels (37). All
samples were analyzed in triplicate. The primers used are listed in
Table SII.
Western blot analysis
Western blot analysis was performed as indicated
previously (34). Briefly, cerebral
cortex samples or HT22 cell homogenates were collected using RIPA
lysis buffer (Beyotime Institute of Biotechnology) containing
protease inhibitor cocktail (Sigma-Aldrich; Merck KGaA), dissolved,
and separated by 10% SDS-PAGE. A BCA Protein Assay kit (Beyotime
Institute of Biotechnology) was used to measure protein
concentrations by the bicinchoninic acid method. Then, protein
samples (30 µg/lane) were transferred onto immobilon nitrocellulose
membranes. The membranes were blocked at room temperature for 1 h
with 5% non-fat milk. The membranes were then incubated with the
following primary antibodies overnight at 4°C: Rabbit anti-β-actin
(1:1,000; rabbit polyclonal; Abcam; cat. no. ab8227), rabbit
anti-RIP1 (1:1,000; rabbit polyclonal; Abcam; cat. no. ab106393),
rabbit anti-RIP3 (1:1,000; rabbit polyclonal; Abcam; cat. no.
ab62344), rabbit anti-MLKL (1:1,000; rabbit monoclonal; Abcam; cat.
no. ab196436), and rabbit anti-PTEN (1:1,000; rabbit monoclonal;
Abcam; cat. no. ab267787). After washing the membranes with TBST
three times, HRP-conjugated goat anti-rabbit IgG or goat anti-mouse
IgG secondary antibodies (1:2,000; cat. no. 7074s; Cell Signaling
Technology, Inc.) were applied, and the membranes were incubated
with the secondary antibodies at room temperature for 1.5 h. The
protein bands were detected using a Bio-Rad imaging system (Bio-Rad
Laboratories, Inc.) and quantified with ImageJ (version 1.52;
National Institutes of Health).
Statistical analysis
All experiments were repeated at least three times,
and the data are expressed as the means ± SEM. SPSS 14.0 (SPSS,
Inc.) and GraphPad Prism 6 (GraphPad Software, Inc.) were used for
the statistical analyses. Student's t-test was used if two groups
were compared, and one-way analysis of variance followed by
Bonferroni's post hoc test was used if two independent variables
were compared. For nonnormally distributed data and/or
nonhomogeneous variance, the Kruskal-Wallis test was used followed
by Dunn's post hoc test. P<0.05 was considered to indicate a
statistically significant difference.
Results
Perampanel alleviates early brain
injury following ICH in vivo
To explore the neuroprotective effects of
perampanel, the autologous blood injection method was used to
construct an ICH model in vivo (Fig. 1A). To investigate early brain injury
following ICH, the present study determined the levels of brain
water content, neurological scores, and apoptosis rates by TUNEL
assay at 72 h after ICH. The results demonstrated that neurological
scores were decreased significantly following ICH, and that
perampanel administration significantly ameliorated this effect
(P<0.05; Fig. 1B). In addition,
the brain water content in the ipsilateral cortex and basal ganglia
was significantly increased following ICH (P<0.05), and
perampanel treatment attenuated this effect (P<0.05) (Fig. 1C). There was no significant
difference in the brain water content among the three experimental
groups in the contralateral cortex, basal ganglia, or cerebellum
(Fig. 1C). To investigate
hippocampal neuron death following ICH and the neuroprotective
effects of perampanel, the present study used a TUNEL assay to
quantify the level of cell death in treated and untreated ICH mice
at 72 h after model construction. The present results indicated
increased hippocampal neuron death following ICH, which was
markedly decreased by perampanel administration (Fig. 1D). Thus, the present data confirmed
the neuroprotective effects of perampanel in ICH.
Perampanel relieves hemin-induced HT22
neuronal injury in vitro
To assess the neuroprotective effect of perampanel
in vitro, a hemin-induced method was used to construct an
ICH model in cultured cells (Fig.
1A). Then, an MTT assay was used to evaluate HT22 neuron
viability following hemin-induced ICH. The results demonstrated
that hemin administration induced HT22 neuron death in a
dose-dependent manner (Fig. 2A). Of
note, the hemin-induced neuronal cell death was significantly
attenuated by perampanel at 72 h (Fig.
2B). A dead/live immunofluorescent staining assay was also used
to detect neuronal death at 72 h. The results indicated that
neuronal death increased following hemin-induced ICH, and that
perampanel treatment relieved hemin-induced neuronal injury in
vitro (Fig. 2C).
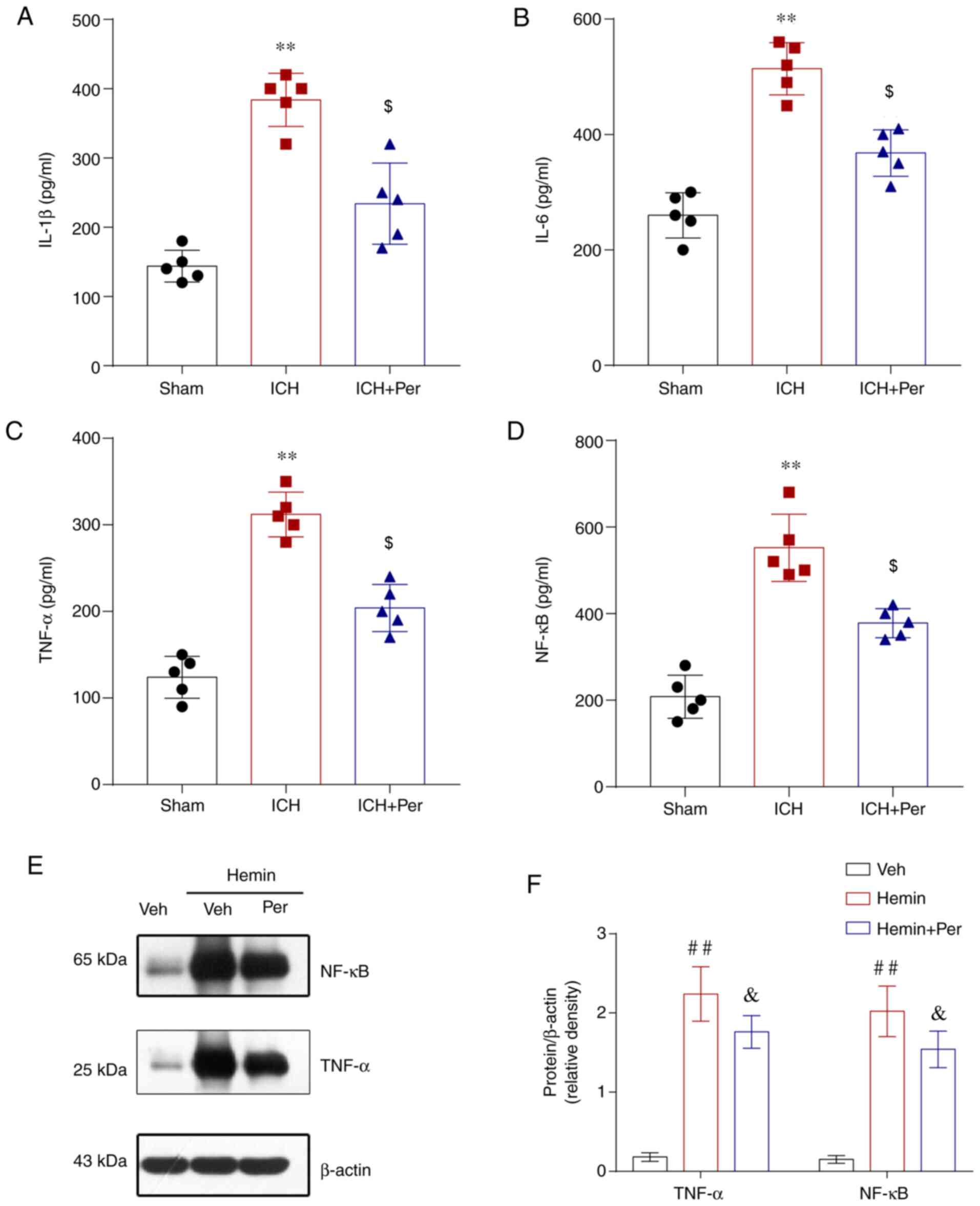
Perampanel relieves neuroinflammation
following ICH
Neuroinflammation serves a key role in the
progression of ICH-induced brain injury, and a previous study
demonstrated that the neuroprotective effect of perampanel after
traumatic brain injury (TBI) may be associated with the
anti-inflammatory response (27).
Therefore, the present study measured the levels of inflammatory
cytokines IL-1β, IL-6, TNF-α, and NF-κB by ELISA. The results
showed that the levels of IL-1β, IL-6, TNF-α, and NF-κB in the
ipsilateral cortex were increased significantly following ICH and
this increase was attenuated by perampanel treatment in vivo
(Fig. 3A-D). Furtermore, the
protein expression levels of TNF-α and NF-κB were measured by
western blot analysis in vitro, and the results confirmed
that perampanel treatment significantly alleviated the
hemin-induced TNF-α and NF-κB upregulation in HT22 cells (Fig. 3E-F).
Perampanel suppresses necroptosis
following ICH
Previous studies have indicated that perampanel can
improve brain injury outcomes following traumatic brain injury by
regulating neuronal necroptosis (27,28).
Therefore, the present study also investigated whether perampanel
improves brain injury after ICH via the necroptosis pathway. First,
the mRNA expression levels of the necroptosis-related genes RIP1,
RIP3, and MLKL in the cerebral cortex were determined by RT-qPCR.
As expected, the mRNA expression levels of RIP1, RIP3, and MLKL
mRNA increased after ICH and decreased after perampanel treatment
(Fig. 4A-C). The protein expression
levels of RIP1, RIP3, and MLKL in the cerebral cortex were also
measured by western blot analysis. The results were similar to the
mRNA analysis results (Fig. 4D-G).
The specific role of necroptosis, a process that is different from
apoptosis and autophagy, was explored next in hemin-induced ICH
in vitro. The results demonstrated that inhibitors of
caspase-dependent apoptosis (z-VAD-fmk) and autophagy (3-MA) did
not affect hemin-induced neuronal injury in vitro, while an
inhibitor of necroptosis-dependent apoptosis (necrostatin-1)
significantly attenuated neuronal death (Fig. 4H).
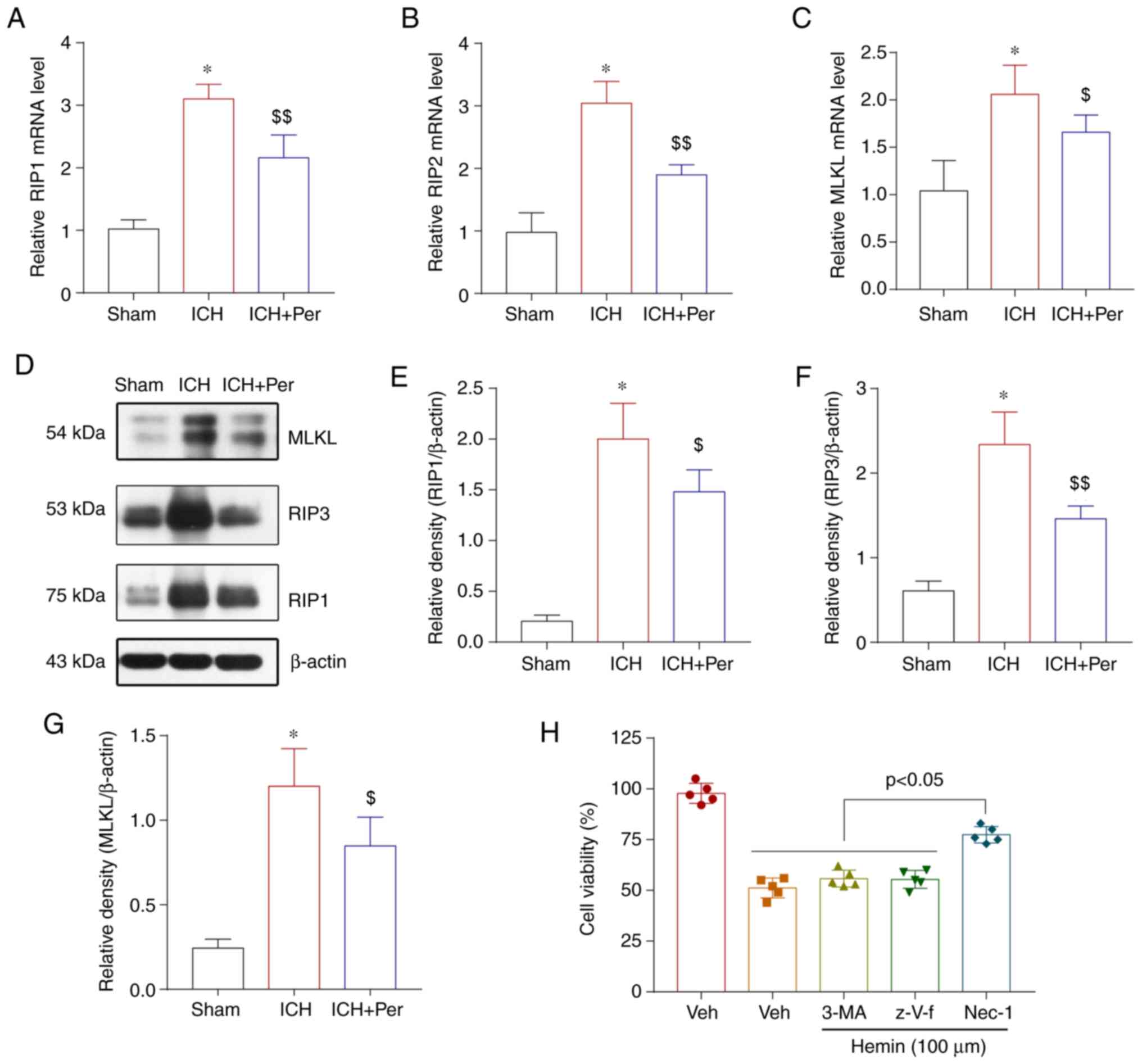 | Figure 4.Perampanel suppresses necroptosis
following ICH. (A) mRNA expression levels of RIP1, (B) RIP3, and
(C) MLKL were determined after ICH by reverse
transcription-quantitative PCR analysis. (D) Representative blots
and (E-G) quantification of RIP1, RIP3, and MLKL protein expression
levels after ICH, as determined by western blotting. (H) MTT assay
of HT22 cells treated in vitro with hemin in the absence or
presence of inhibitors of caspase-dependent apoptosis (z-VAD-fmk),
autophagy (3-MA), or necroptosis (Nec-1). Error bars indicate SEM
(n=5). *P<0.01 vs. sham group; $P<0.05 and
$$P<0.01 vs. ICH group. ICH, intracerebral
hemorrhage; RIP, receptor interacting serine/threonine kinase;
MLKL, mixed lineage kinase domain like pseudokinase; Nec-1,
necrostatin-1. |
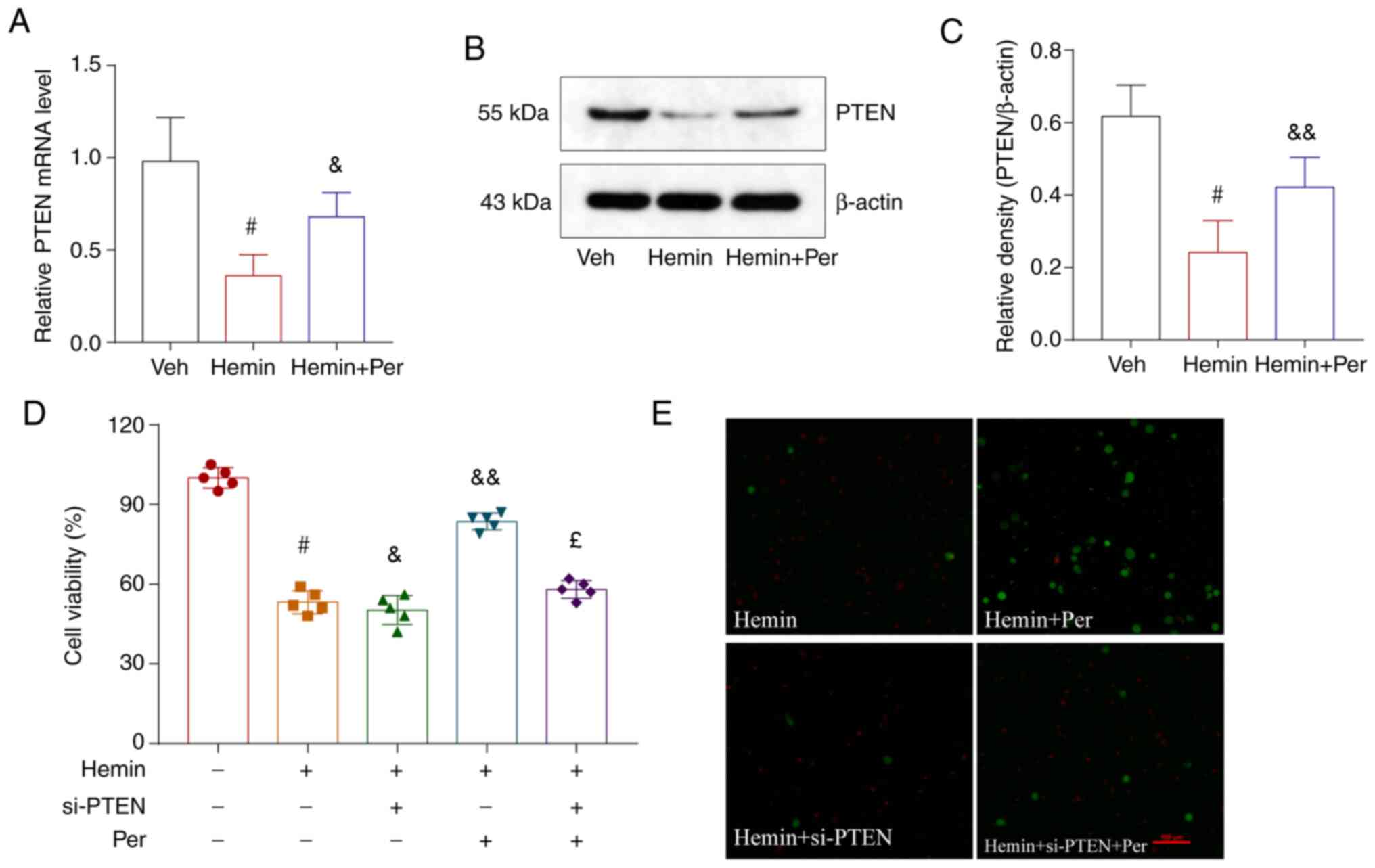
The neuroprotective effect of
perampanel is mediated via the PTEN signaling pathway
As Kim et al (38) reported that AMPAR-mediated
spontaneous seizure activity may abolish PTEN activity in the
epileptic hippocampus, PTEN may be required for the antiepileptic
effects of AMPAR antagonists (25).
Therefore, the present study explored whether the antinecroptotic
role of perampanel may occur via the PTEN signaling pathway. First,
the mRNA expression levels of PTEN were measured after
hemin-induced ICH and perampanel treatment in vitro. The
results demonstrated that PTEN mRNA expression was reduced after
ICH, and this reduction was alleviated by perampanel treatment
(Fig. 5A). Similar results were
observed at the protein level, as determined by western blot
analysis (Fig. 5B and C).
Next, transfection with PTEN-targeting siRNA was
performed to silence the expression of PTEN in the hemin-induced
ICH model in vitro. RT-qPCR was used to confirm that the
transfections were successful and PTEN expression was efficiently
silenced (Fig. S1). Following PTEN
knockdown, the hemin-induced neuronal death was not improved after
perampanel treatment (Fig. 5D and
E). Therefore, the neuroprotective effect of perampanel was
reversed by knockdown of PTEN in the ICH in vitro model.
Perampanel alleviates PTEN-dependent
brain injury in vivo
The in vitro results demonstrated that
hemin-induced ICH decreased the expression of PTEN, perampanel
treatment increased PTEN expression, and PTEN knockdown blocked the
neuroprotective effects of perampanel. To further investigate the
role of PTEN in the in vivo ICH model, si-PTEN was infused
into the left lateral ventricle of mice to downregulate the
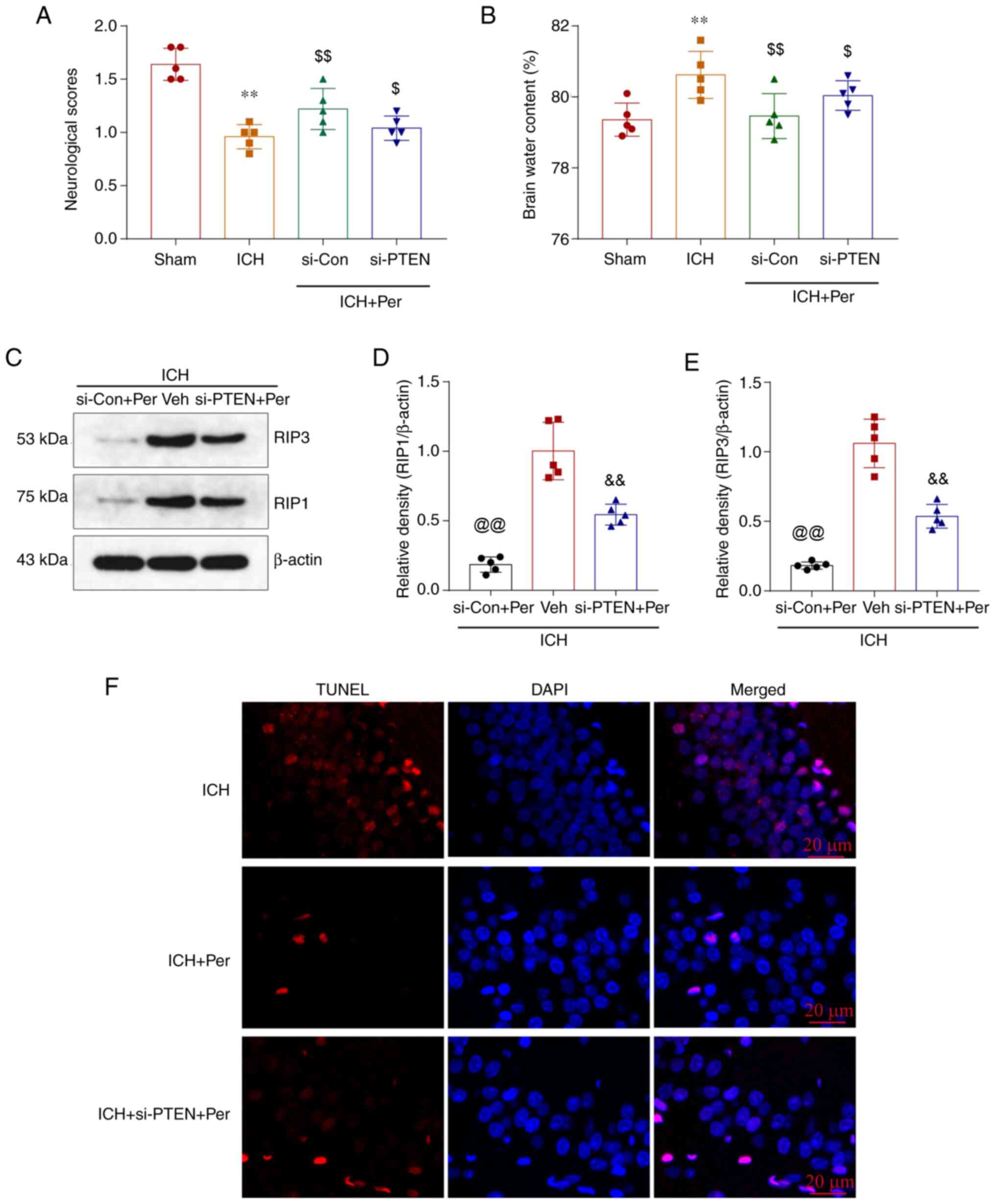
expression of PTEN. The results showed that the behavioral score
and ipsilateral basal ganglia brain edema were improved by
perampanel treatment, but this effect was partially blocked by PTEN
knockdown (Fig. 6A and B). The
expression levels of the necroptosis-related proteins RIP1 and RIP3
in the ipsilateral cortex tissue were also decreased after
perampanel treatment, and this effect was reduced following PTEN
knockdown (Fig. 6C-E). Finally,
TUNEL staining of brain sections was performed (Fig. 6F), and the results showed that the
antinecroptotic role of perampanel was partially inhibited by PTEN
knockdown.
Discussion
The present study evaluated the therapeutic
potential of perampanel in alleviating early brain injury in an
in vivo ICH mouse model and an in vitro hemin-induced
ICH cell culture model. The current results demonstrated that
perampanel acted as a neuroprotective agent that attenuated early
brain injury and neuronal death following ICH. In specific, it was
found that: i) Perampanel improved neurological dysfunction
following ICH; ii) perampanel alleviated brain damage in a mouse
ICH model and in a hemin-induced HT22 cell neuronal injury model;
iii) perampanel relieved neuroinflammation after ICH and decreased
brain inflammatory damage; iv) perampanel prevented necroptosis
after ICH and alleviated neuronal death; and v) the
antineuroinflammatory and antinecroptotic roles of perampanel may
be mediated by the PTEN pathway.
Perampanel, an orally active, noncompetitive,
selective AMPAR antagonist, has been widely used for the treatment
of epilepsy (23). Previous studies
have revealed that AMPAR antagonists are important potential
neuroprotective agents in preclinical models of ischemic stroke
(39), spinal cord injury (40), and traumatic brain injury (27). However, while these early studies
revealed that AMPAR antagonists could minimize brain injury and
neuronal death following ICH, the potential molecular mechanisms
and consequent effects were not explored. Chen et al
(27) first reported that the AMPAR
antagonist perampanel could alleviate TBI-induced neuronal injury
and neurological dysfunction by suppressing the expression of
proinflammatory cytokines and increasing the levels of
anti-inflammatory cytokines. The underlying mechanisms may be
related to antioxidative and anti-inflammatory activities. Aida
et al (41) also
demonstrated that pre- or postinjury administration of perampanel
could reduce impairments in learning and memory, and impairments in
reward-seeking behavior, by inhibiting hippocampal cell apoptosis.
In brain hemorrhage, Dohare et al (25) first found that AMPA-specific
inhibition by FDA-approved perampanel can decrease oligodendrocyte
precursor cell injury, rebuild myelination, and improve neurologic
recovery in rabbits with ICH. In addition to reporting mechanisms
of apoptosis and oxidative and inflammatory activity, Chen et
al (28) first indicated that
necroptosis had a critical role in neuronal death following
traumatic neuronal injury in an in vitro model, and that
perampanel could regulate necroptosis via the AKT/GSK3β signaling
pathway.
Necroptosis is a newly discovered pathway of
regulated necrosis, a caspase-independent programmed cell death
mechanism that requires the proteins RIP3 and MLKL and is induced
by death receptors (42).
Increasing evidence suggests that necroptosis serves a critical
role in central nervous system diseases, including traumatic brain
injury (28,43,44),
ICH (45,46), ischemic stroke (47), amyotrophic lateral sclerosis,
Parkinson's disease, and Alzheimer's disease (48). The most upstream signaling activity
required for the induction of necroptosis is the activation of a
TNF ligand family member (such as RIP1 and MLKL); RIPK1 activation
leads to necroptosis through the formation of a RIP1/RIP3/MLKL
complex (49). The present study
revealed that the protein and mRNA expression levels of RIP1, RIP3,
and MLKL were elevated 72 h after the ICH procedure, and perampanel
decreased the expression of RIP1, RIP3, and MLKL. Necrostatin-1 is
a specific inhibitor of RIP1 that prevents necroptosis and
ameliorates cell death, brain edema, and neuroinflammation and
improves behavioral scores following ischemic brain injury and
traumatic brain injury (28,45,47).
Furthermore, Xu et al (50)
reported that necrostatin-1 could prevent glutamate-induced
glutathione depletion and decrease caspase-independent cell death
in HT22 cells. The present results also revealed that necrostatin-1
significantly decreased neuronal death after hemin-induced HT22
neuronal injury.
Although the results of many studies have confirmed
that necroptosis and RIP1 are involved in multiple central nervous
system diseases, the regulatory mechanisms and molecular networks
of necroptosis remain unclear. Chen et al (43) reported that activity regulated
cytoskeleton associated protein (Arc) silencing aggravated neuronal
injury via the metabotropic glutamate receptor 1-mediated
endoplasmic reticulum stress pathway and RIP1-dependent
necroptosis. Recent studies have revealed that iron overload can
induce necroptosis of osteoblastic cells in vitro partly
through the RIP1/RIP3/MLKL pathway, and reactive oxygen species
(ROS) have an important role in regulating necroptosis (51–53).
The present study demonstrated that necroptosis lead to neuronal
death after ICH and hemin-induced ICH, that neuronal death was
inhibited by perampanel treatment, and that the neuroprotectice
effects of perampanel were dependent on the PTEN pathway. Hence,
the current results indicated that perampanel could reduce neuronal
death and brain injury by protecting against neural necroptosis
through the PTEN pathway. Wang et al (29) also reported that Chlorpyrifos
induced fish liver cell necroptosis by regulating the
ROS/PTEN/PI3K/AKT axis. The role of PTEN in the necroptosis
molecular regulatory network was also confirmed in human seminoma
and prostate cancer cells (30,54).
Previous studies have revealed that the expression levels of PTEN
are high in mouse brain neurons and can regulate survival and
death, and that PTEN inhibition has a vital role in neuroprotection
following ICH (55,56). Additionally, the PTEN pathway is
involved in regulating neuroinflammation and mediating microglial
and astrocyte activation and neuronal degeneration (57,58).
Together, the present findings indicated that the antinecroptosis
and antineuroinflammation effects of perampanel occur at least in
part through the regulation of the PTEN pathway after ICH. However,
further studies are needed to fully elucidate the specific
underlying regulatory mechanisms of necroptosis in ICH-induced
early brain injury.
In summary, the present study demonstrated that the
noncompetitive AMPAR antagonist perampanel exerted potent
neuroprotective effects in early brain injury and protected against
ICH-induced neuronal death in both in vivo and in
vitro models. Perampanel alleviated brain injury and inhibited
neuroinflammation and necroptosis. The neuroprotective effects of
perampanel were found to be mediated by PTEN. The present findings
indicated that perampanel is a neuroprotective agent that might
improve the clinical management of early brain injury after
ICH.
Supplementary Material
Supporting Data
Acknowledgements
The authors would like to thank Dr Tao Chen (904th
Hospital of Joint Logistic Support Force of PLA, Wuxi, China) for
providing technical and theoretical assistance.
Funding
No funding was received.
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
LY performed the experiments and wrote the
manuscript. LY, YW and CZ assisted in performing the experiments
and processed the figures. HC designed the study and revised the
manuscript. LY and HC confirm the authenticity of all the raw data.
All authors read and approved the final manuscript.
Ethics approval and consent to
participate
All protocols involving animals were approved by the
Ethics Committee of the Wuxi Medical College of Anhui Medical
University (Wuxi, China; approval no. YXLL-2020-009).
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Zhang Y, Zhang X, Wei Q, Leng S, Li C, Han
B, Bai Y, Zhang H and Yao H: Activation of Sigma-1 receptor
enhanced pericyte survival via the interplay between apoptosis and
autophagy: Implications for blood-brain barrier integrity in
stroke. Transl Stroke Res. 11:267–287. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Zhang Z, Cho S, Rehni AK, Quero HN, Dave
KR and Zhao W: Automated assessment of hematoma volume of rodents
subjected to experimental intracerebral hemorrhagic stroke by bayes
segmentation approach. Transl Stroke Res. 11:789–798. 2020.
View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Gross BA, Jankowitz BT and Friedlander RM:
Cerebral intraparenchymal hemorrhage: A Review. JAMA.
321:1295–1303. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Wu X, Luo J, Liu H, Cui W, Guo K, Zhao L,
Bai H, Guo W, Guo H, Feng D and Qu Y: Recombinant adiponectin
peptide ameliorates brain injury following intracerebral hemorrhage
by suppressing astrocyte-derived inflammation via the inhibition of
Drp1-mediated mitochondrial fission. Transl Stroke Res. 11:924–939.
2020. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Hanley DF, Thompson RE, Rosenblum M,
Yenokyan G, Lane K, McBee N, Mayo SW, Bistran-Hall AJ, Gandhi D,
Mould WA, et al: Efficacy and safety of minimally invasive surgery
with thrombolysis in intracerebral haemorrhage evacuation (MISTIE
III): A randomised, controlled, open-label, blinded endpoint phase
3 trial. Lancet. 393:1021–1032. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Chen J, Wang Y, Wu J, Yang J, Li M and
Chen Q: The potential value of targeting ferroptosis in early brain
injury after acute CNS disease. Front Mol Neurosci. 13:1102020.
View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Adeoye O and Broderick JP: Advances in the
management of intracerebral hemorrhage. Nat Rev Neurol. 6:593–601.
2010. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Mendelow AD, Gregson BA, Rowan EN, Murray
GD, Gholkar A and Mitchell PM: Early surgery versus initial
conservative treatment in patients with spontaneous supratentorial
lobar intracerebral haematomas (STICH II): A randomised trial.
Lancet. 382:397–408. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Zhou Y, Wang Y, Wang J, Anne Stetler R and
Yang QW: Inflammation in intracerebral hemorrhage: From mechanisms
to clinical translation. Prog Neurobiol. 115:25–44. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Xue M and Yong VW: Neuroinflammation in
intracerebral haemorrhage: Immunotherapies with potential for
translation. Lancet Neurol. 19:1023–1032. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Chen JH, Yang LK, Chen L, Wang YH, Wu Y,
Jiang BJ, Zhu J and Li PP: Atorvastatin ameliorates early brain
injury after subarachnoid hemorrhage via inhibition of AQP4
expression in rabbits. Int J Mol Med. 37:1059–1066. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Bao WD, Zhou XT, Zhou LT, Wang F, Yin X,
Lu Y, Zhu LQ and Liu D: Targeting miR-124/Ferroportin signaling
ameliorated neuronal cell death through inhibiting apoptosis and
ferroptosis in aged intracerebral hemorrhage murine model. Aging
Cell. 19:e132352020. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Gautam J, Xu L, Nirwane A, Nguyen B and
Yao Y: Loss of mural cell-derived laminin aggravates hemorrhagic
brain injury. J Neuroinflammation. 17:1032020. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Karuppagounder SS, Alim I, Khim SJ,
Bourassa MW, Sleiman SF, John R, Thinnes CC, Yeh TL, Demetriades M,
Neitemeier S, et al: Therapeutic targeting of oxygen-sensing prolyl
hydroxylases abrogates ATF4-dependent neuronal death and improves
outcomes after brain hemorrhage in several rodent models. Sci
Transl Med. 8:328ra3292016. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Chen J, Zhang C, Yan T, Yang L, Wang Y,
Shi Z, Li M and Chen Q: Atorvastatin ameliorates early brain injury
after subarachnoid hemorrhage via inhibition of pyroptosis and
neuroinflammation. J Cell Physiol. Mar 31–2021.(Epub ahead of
print). doi: 10.1002/jcp.30351. View Article : Google Scholar
|
|
16
|
Wang T, Nowrangi D, Yu L, Lu T, Tang J,
Han B, Ding Y, Fu F and Zhang JH: Activation of dopamine D1
receptor decreased NLRP3-mediated inflammation in intracerebral
hemorrhage mice. J Neuroinflammation. 15:22018. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Wang M, Ye X, Hu J, Zhao Q, Lv B, Ma W,
Wang W, Yin H, Hao Q, Zhou C, et al: NOD1/RIP2 signalling enhances
the microglia-driven inflammatory response and undergoes crosstalk
with inflammatory cytokines to exacerbate brain damage following
intracerebral haemorrhage in mice. J Neuroinflammation. 17:3642020.
View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Chen S, Peng J, Sherchan P, Ma Y, Xiang S,
Yan F, Zhao H, Jiang Y, Wang N, Zhang JH and Zhang H: TREM2
activation attenuates neuroinflammation and neuronal apoptosis via
PI3K/Akt pathway after intracerebral hemorrhage in mice. J
Neuroinflammation. 17:1682020. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Chen AQ, Fang Z, Chen XL, Yang S, Zhou YF,
Mao L, Xia YP, Jin HJ, Li YN, You MF, et al: Microglia-derived
TNF-α mediates endothelial necroptosis aggravating blood
brain-barrier disruption after ischemic stroke. Cell Death Dis.
10:4872019. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Chu X, Wu X, Feng H, Zhao H, Tan Y, Wang
L, Ran H, Yi L, Peng Y, Tong H, et al: Coupling between
Interleukin-1R1 and necrosome complex involves in hemin-induced
neuronal necroptosis after intracranial hemorrhage. Stroke.
49:2473–2482. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Su X, Wang H, Kang D, Zhu J, Sun Q, Li T
and Ding K: Necrostatin-1 ameliorates intracerebral
hemorrhage-induced brain injury in mice through inhibiting
RIP1/RIP3 pathway. Neurochem Res. 40:643–650. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Lule S, Wu L, Sarro-Schwartz A, Edmiston
Iii WJ, Izzy S, Songtachalert T, Ahn SH, Fernandes ND, Jin G, Chung
JY, et al: Cell-specific activation of RIPK1 and MLKL after
intracerebral hemorrhage in mice. J Cereb Blood Flow Metab. Nov
19–2020.(Epub ahead of print). doi: 10.1177/0271678X20973609.
PubMed/NCBI
|
|
23
|
Weiping L, Dong Z, Zhen H, Patten A, Dash
A and Malhotra M: Efficacy, safety, and tolerability of adjunctive
perampanel in patients from China with focal seizures or
generalized tonic-clonic seizures: Post hoc analysis of phase III
double-blind and open-label extension studies. CNS Neurosci Ther.
27:330–340. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Chen T, Liu WB, Qian X, Xie KL and Wang
YH: The AMPAR antagonist perampanel protects the neurovascular unit
against traumatic injury via regulating Sirt3. CNS Neurosci Ther.
27:134–144. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Dohare P, Zia MT, Ahmed E, Ahmed A, Yadala
V, Schober AL, Ortega JA, Kayton R, Ungvari Z, Mongin AA and
Ballabh P: AMPA-kainate receptor inhibition promotes neurologic
recovery in premature rabbits with intraventricular hemorrhage. J
Neurosci. 36:3363–3377. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Wright SK, Wilson MA, Walsh R, Lo WB,
Mundil N, Agrawal S, Philip S, Seri S, Greenhill SD and Woodhall
GL: Abolishing spontaneous epileptiform activity in human brain
tissue through AMPA receptor inhibition. Ann Clin Transl Neurol.
7:883–890. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Chen T, Dai SH, Jiang ZQ, Luo P, Jiang XF,
Fei Z, Gui SB and Qi YL: The AMPAR antagonist perampanel attenuates
traumatic brain injury through anti-oxidative and anti-inflammatory
activity. Cell Mol Neurobiol. 37:43–52. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Chen T, Yang LK, Zhu J, Hang CH and Wang
YH: The AMPAR antagonist perampanel regulates neuronal necroptosis
via Akt/GSK3β signaling after acute traumatic injury in cortical
neurons. CNS Neurol Disord Drug Targets. Oct 1–2020.(Epub ahead of
print). doi: 10.2174/1871527319666201001110937. View Article : Google Scholar
|
|
29
|
Wang L, Wang L, Shi X and Xu S:
Chlorpyrifos induces the apoptosis and necroptosis of L8824 cells
through the ROS/PTEN/PI3K/AKT axis. J Hazard Mater. 398:1229052020.
View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Guido C, Panza S, Santoro M, Avena P,
Panno ML, Perrotta I, Giordano F, Casaburi I, Catalano S, De Amicis
F, et al: Estrogen receptor beta (ERβ) produces autophagy and
necroptosis in human seminoma cell line through the binding of the
Sp1 on the phosphatase and tensin homolog deleted from chromosome
10 (PTEN) promoter gene. Cell Cycle. 11:2911–2921. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Gao X, Xiong Y, Li Q, Han M, Shan D, Yang
G, Zhang S, Xin D, Zhao R, Wang Z, et al: Extracellular
vesicle-mediated transfer of miR-21-5p from mesenchymal stromal
cells to neurons alleviates early brain injury to improve cognitive
function via the PTEN/Akt pathway after subarachnoid hemorrhage.
Cell Death Dis. 11:3632020. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Deng S, Sherchan P, Jin P, Huang L, Travis
Z, Zhang JH, Gong Y and Tang J: Recombinant CCL17 enhances hematoma
resolution and activation of CCR4/ERK/Nrf2/CD163 signaling pathway
after intracerebral hemorrhage in mice. Neurotherapeutics.
17:1940–1953. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Chen J, Xuan Y, Chen Y, Wu T, Chen L, Guan
H, Yang S, He J, Shi D and Wang Y: Netrin-1 alleviates subarachnoid
haemorrhage-induced brain injury via the PPARγ/NF-κB signalling
pathway. J Cell Mol Med. 23:2256–2262. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Chen JH, Wu T, Xia WY, Shi ZH, Zhang CL,
Chen L, Chen QX and Wang YH: An early neuroprotective effect of
atorvastatin against subarachnoid hemorrhage. Neural Regen Res.
15:1947–1954. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Mo Y, Duan L, Yang Y, Liu W, Zhang Y, Zhou
L, Su S, Lo PC, Cai J, Gao L, et al: Nanoparticles improved
resveratrol brain delivery and its therapeutic efficacy against
intracerebral hemorrhage. Nanoscale. 13:3827–3840. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Chen JH, Wu T, Yang LK, Chen L, Zhu J, Li
PP, Hu X and Wang YH: Protective effects of atorvastatin on
cerebral vessel autoregulation in an experimental rabbit model of
subarachnoid hemorrhage. Mol Med Rep. 17:1651–1659. 2018.PubMed/NCBI
|
|
37
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Kim JE, Park H, Lee JE, Kim TH and Kang
TC: PTEN is required for the anti-epileptic effects of AMPA
receptor antagonists in chronic epileptic rats. Int J Mol Sci.
21:56432020. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Mazzocchetti P, Mancini A, Sciaccaluga M,
Megaro A, Bellingacci L, Di Filippo M, Cesarini EN, Romoli M,
Carrano N, Gardoni F, et al: Low doses of Perampanel protect
striatal and hippocampal neurons against in vitro ischemia by
reversing the ischemia-induced alteration of AMPA receptor subunit
composition. Neurobiol Dis. 140:1048482020. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
De Caro C, Cristiano C, Avagliano C,
Cuozzo M, La Rana G, Aviello G, De Sarro G, Calignano A, Russo E
and Russo R: Analgesic and Anti-inflammatory effects of perampanel
in acute and chronic pain models in mice: Interaction with the
cannabinergic system. Front Pharmacol. 11:6202212021. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Aida V, Niedzielko TL, Szaflarski JP and
Floyd CL: Acute administration of perampanel, an AMPA receptor
antagonist, reduces cognitive impairments after traumatic brain
injury in rats. Exp Neurol. 327:1132222020. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Vandenabeele P, Galluzzi L, Vanden Berghe
T and Kroemer G: Molecular mechanisms of necroptosis: An ordered
cellular explosion. Nat Rev Mol Cell Biol. 11:700–714. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Chen T, Zhu J, Wang YH and Hang CH: Arc
silence aggravates traumatic neuronal injury via mGluR1-mediated ER
stress and necroptosis. Cell Death Dis. 11:42020. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Bao Z, Fan L, Zhao L, Xu X, Liu Y, Chao H,
Liu N, You Y, Liu Y, Wang X and Ji J: Silencing of A20 aggravates
neuronal death and inflammation after traumatic brain injury: A
potential trigger of necroptosis. Front Mol Neurosci. 12:2222019.
View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Laird MD, Wakade C, Alleyne CH Jr and
Dhandapani KM: Hemin-induced necroptosis involves glutathione
depletion in mouse astrocytes. Free Radic Biol Med. 45:1103–1114.
2008. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Shen H, Liu C, Zhang D, Yao X, Zhang K, Li
H and Chen G: Role for RIP1 in mediating necroptosis in
experimental intracerebral hemorrhage model both in vivo and in
vitro. Cell Death Dis. 8:e26412017. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Zhang Y, Li M, Li X, Zhang H, Wang L, Wu
X, Zhang H and Luo Y: Catalytically inactive RIP1 and RIP3
deficiency protect against acute ischemic stroke by inhibiting
necroptosis and neuroinflammation. Cell Death Dis. 11:5652020.
View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Yuan J, Amin P and Ofengeim D: Necroptosis
and RIPK1-mediated neuroinflammation in CNS diseases. Nat Rev
Neurosci. 20:19–33. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Linkermann A and Green DR: Necroptosis. N
Engl J Med. 370:455–465. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Xu X, Chua CC, Kong J, Kostrzewa RM,
Kumaraguru U, Hamdy RC and Chua BH: Necrostatin-1 protects against
glutamate-induced glutathione depletion and caspase-independent
cell death in HT-22 cells. J Neurochem. 103:2004–2014. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Liu L, Liu Y, Cheng X and Qiao X: The
alleviative effects of quercetin on Cadmium-induced necroptosis via
inhibition ROS/iNOS/NF-κB pathway in the Chicken brain. Biol Trace
Elem Res. 199:1584–1594. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Wang KJ, Meng XY, Chen JF, Wang KY, Zhou
C, Yu R and Ma Q: Emodin induced necroptosis and inhibited
glycolysis in the renal cancer cells by enhancing ROS. Oxid Med
Cell Longev. 2021:88405902021.PubMed/NCBI
|
|
53
|
Tian Q, Qin B, Gu Y, Zhou L, Chen S, Zhang
S, Zhang S, Han Q, Liu Y and Wu X: ROS-mediated necroptosis is
involved in iron overload-induced osteoblastic cell death. Oxid Med
Cell Longev. 2020:12953822020. View Article : Google Scholar : PubMed/NCBI
|
|
54
|
Liu X, Zhang N, Wang D, Zhu D, Yuan Q,
Zhang X, Qian L, Niu H, Lu Y, Ren G, et al: Downregulation of
reticulocalbin-1 differentially facilitates apoptosis and
necroptosis in human prostate cancer cells. Cancer Sci.
109:1147–1157. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
55
|
Zhao D, Qin XP, Chen SF, Liao XY, Cheng J,
Liu R, Lei Y, Zhang ZF and Wan Q: PTEN inhibition protects against
experimental intracerebral hemorrhage-induced brain injury through
PTEN/E2F1/β-catenin pathway. Front Mol Neurosci. 12:2812019.
View Article : Google Scholar : PubMed/NCBI
|
|
56
|
Chang N, El-Hayek YH, Gomez E and Wan Q:
Phosphatase PTEN in neuronal injury and brain disorders. Trends
Neurosci. 30:581–586. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
57
|
Huang H, Hu C, Xu L, Zhu X, Zhao L and Min
J: The effects of hesperidin on neuronal apoptosis and cognitive
impairment in the sevoflurane anesthetized rat are mediated through
the PI3/Akt/PTEN and nuclear Factor-κB (NF-κB) signaling pathways.
Med Sci Monit. 26:e9205222020. View Article : Google Scholar : PubMed/NCBI
|
|
58
|
Huang SY, Sung CS, Chen WF, Chen CH, Feng
CW, Yang SN, Hung HC, Chen NF, Lin PR, Chen SC, et al: Involvement
of phosphatase and tensin homolog deleted from chromosome 10 in
rodent model of neuropathic pain. J Neuroinflammation. 12:592015.
View Article : Google Scholar : PubMed/NCBI
|