Introduction
Myocardial infarction (MI) remains one of the
leading causes of mortality and disability worldwide (1). At present, the most effective approach
for reducing MI injury and size is timely myocardial reperfusion
(2). However, unsatisfactorily,
reperfusion causes further myocardial injury and cardiomyocyte
death, causing a phenomenon denominated myocardial ischemia and
reperfusion (MI/R) injury (3). MI/R
injury contributes to disease progression in patients with MI.
However, some mechanical and pharmacological therapeutic strategies
for preventing MI/R injury have been shown to be effective in
improving clinical outcomes (2).
Recently, MI pathology has been increasingly
associated with long non-coding RNAs (lncRNAs) (4,5), a
novel class of endogenous cellular RNAs that regulate gene
expression through various means, including acting as microRNA
(miRNA) sponges (6). The antisense
non-coding RNA in the INK4 locus (ANRIL) can regulate cell
proliferation, senescence and apoptosis involved in some
cardiovascular diseases and human cancers (7). In particular, ANRIL was newly
identified as a dysregulated lncRNA in patients with MI compared
with healthy controls (8). In
addition, an ANRIL polymorphism (rs1333049:C>G) is associated
with MI (9). Consistently, another
previous study also demonstrated that ANRIL gene variants
contribute to MI risk (10). These
reports suggest that ANRIL may play a functional role in the
pathology of MI. However, to the best of our knowledge, at present,
no evidence has been provided to mechanistically link ANRIL with
MI/R injury.
In the present study, the role of ANRIL in the
regulation of MI/R injury was investigated using a
hypoxia/reoxygenation (H/R) in vitro experimental model. It
was found that the expression of ANRIL was downregulated after H/R
injury and that ANRIL positively regulated sirtuin 1 (SIRT1)
expression by sponging miR-181a. This regulatory mechanism may be
responsible for the protective role of ANRIL against H/R-induced
cardiomyocyte injury. These data could offer a novel mechanistic
insight into understanding the association between ANRIL and MI
pathology.
Materials and methods
Cell culture and H/R treatment
H9c2 cardiomyocytes (cat. no. CRL-1446) were
purchased from the American Type Culture Collection and then
cultured in DMEM (Gibco; Thermo Fisher Scientific, Inc.)
supplemented with 10% (v/v) FBS (Gibco; Thermo Fisher Scientific,
Inc.), 100 U/ml penicillin and 100 mg/ml streptomycin (Gibco;
Thermo Fisher Scientific, Inc.) in an incubator at 37°C with 5%
CO2. In vitro H/R treatment was used to mimic
MI/R injury, as described previously (11). Briefly, H9c2 cardiomyocytes were
cultured in glucose-free DMEM with an oxygen-free atmosphere (95%
N2 and 5% CO2) at 37°C for 4 h, followed by
incubation with normal medium (4.5 mg/ml glucose) and atmosphere
(95% air and 5% CO2) at 37°C for 2 to 24 h, depending on
the experimental purposes.
Transfection
ANRIL-overexpressing plasmids were constructed by
inserting the ANRIL coding sequence into the pcDNA3.1 vector
(Invitrogen; Thermo Fisher Scientific, Inc.). Oligonucleotides,
including miR-181a mimic (5′-AACAUUCAACGCUGUCGGUGAGU-3′), negative
control (NC) mimic (5′-UUCUCCGAACGUGUCACGUTT−3′), miR-181a
antagomir (5′-ACUCACCGACAGCGUUGAAUGUU-3′) and NC antagomir
(5′-CAGUACUUUUGUGUAGUACAA−3′), were purchased from Guangzhou
RiboBio Co., Ltd. Small interfering (si)RNA targeting SIRT1
(siSIRT1, 5′-CCCUGUAAAGCUUUCAGAATT−3′) and NC siRNA (siNC;
5′-UUCUCCGAACGUGUCACGUTT−3′) were obtained from Shanghai GenePharma
Co., Ltd. When the cell density reached 50–60% confluence, cell
transfection was performed with Lipofectamine® 2000
reagent (Invitrogen; Thermo Fisher Scientific, Inc.) according to
the manufacturer's protocols. Equal amounts of empty pcDNA3.1
vector (4 µg/well), NC mimic, NC antagomir (50 or 150 nM) or siNC
(10 or 20 nM) diluted in the same volume of transfection reagents
were transfected as controls, depending on the experimental
purposes. The efficiency was examined 2 days after transfection.
For H/R treatment, cells were cultured under normoxia or exposed to
H/R (hypoxia for 4 h followed by reoxygenation for 8 h) at 2 days
after transfection. The transfection efficiency for all
transfections was effective under normoxic or H/R treatment
conditions.
Reverse transcription-quantitative PCR
(RT-qPCR) analysis
Total RNA from H9c2 cells was extracted using
TRIzol® reagent (Invitrogen; Thermo Fisher Scientific,
Inc.) according to the manufacturer's protocol. ANRIL and SIRT1
expression levels were determined via RT-qPCR analysis using the
One Step PrimeScript™ RT-RNA kit (Takara Biotechnology Co., Ltd.)
according to the manufacturer's protocol. miR-181a expression was
determined using the TaqMan™ MicroRNA Reverse Transcription kit and
TaqMan Universal Master Mix II (Applied Biosystems; Thermo Fisher
Scientific, Inc.), according to the manufacturer's protocols. The
thermocycling conditions were as follows: Initial denaturation at
95°C for 5 min, followed by 40 cycles of 95°C for 15 sec and 60°C
for 1 min. β-actin served as an internal control for ANRIL and
SIRT1. U6 served as an internal control for miR-181a. Data were
calculated using the 2−ΔΔCq method (12). The following primer sequences were
used for the qPCR: ANRIL forward, 5′-CAAGCCACGTTGGAAGATGC-3′ and
reverse, 5′-AGAGTGTGTAGCAGCTGACG-3′; miR-181a forward,
5′-CACTCCAGCTGGGAACATTCAACGCTGTCGG-3′ and reverse,
5′-TGGTGTCGTGGAGTCG-3′; U6 forward, 5′-TGGGGTTATACATTGTGAGAGGA-3′
and reverse, 5′-GTGTGCTACGGAGTTCAGAGGTT-3′; SIRT1 forward,
5′-CCAGATCCTCAAGCCATG−3′ and reverse, 5′-TTGGATTCCTGCAACCTG−3′; and
β-actin forward, 5′-CATGCCATCCTGCGTCTGGA-3′ and reverse,
5′-CCACATCTGCTGGAAGGTGG-3′.
Immunoblotting
Total cellular proteins were extracted using RIPA
lysis buffer (Beyotime Institute of Biotechnology) supplemented
with the protease inhibitor cocktail (Roche Diagnostics GmbH). The
protein concentration was determined using a BCA protein assay
(Pierce; Thermo Fisher Scientific, Inc.). Total proteins (30 µg per
well) were separated via 10% SDS-PAGE electrophoresis, and then
separated proteins were transferred to polyvinylidene difluoride
membranes, which were then blocked for 1 h with 5% non-fat milk at
room temperature and incubated overnight at 4°C with primary
antibodies against SIRT1 (1:1,000; cat. no. ab7343; Abcam), β-actin
(1:5,000; cat. no. sc-47778; Santa Cruz Biotechnology, Inc.),
cleaved caspase-3 (1:500; cat. no. ab49822; Abcam), Bcl-2 (1:500;
cat. no. sc-7382; Santa Cruz Biotechnology, Inc.) and Bax (1:2,000;
cat. no. NBP1-28566; Novus Biologicals, LLC). After washing, the
membranes were further incubated at room temperature for 1 h with a
horseradish peroxidase-conjugated secondary antibody (1:10,000;
cat. no. ab6721; Abcam). Protein bands were visualized with the
Immobilon Western Chemiluminescent HRP Substrate (EMD Millipore).
Protein expression was semi-quantified using Image Lab software
(version 5.1; Bio-Rad Laboratories, Inc.).
H/R injury detection
The terminal deoxynucleotidyl-transferase mediated
dUTP nick-end labeling (TUNEL) method was carried out to evaluate
cell apoptosis using the TUNEL assay kit (OriGene Technologies,
Inc.) according to the manufacturer's protocols. Briefly, cells
were fixed with 4% paraformaldehyde solution for 1 h at room
temperature and then stained with TUNEL reagent for 1 h at 37°C.
The nuclei were stained with 1 µg/ml DAPI for 15 min at room
temperature. Cells were mounted with mounting solution (Beyotime
Institute of Biotechnology), and images were captured with the
LSM510META fluorescence microscope (Carl Zeiss AG; magnification,
×100). TUNEL-positive cells were considered as apoptotic cells and
counted in 12 random fields from each treatment group. Cell
apoptosis was expressed as the number of apoptotic cells divided by
the total number of cells. Lactate dehydrogenase (LDH) release was
determined using an LDH cytotoxicity detection kit (cat. no. A020;
Nanjing Jiancheng Bioengineering Institute) following the
manufacturer's instructions. The absorbance at 450 nm was measured
using the Synergy HT microplate reader (BioTek Instruments, Inc.).
The results were calculated as the LDH activity in the medium
divided by the LDH activity in total cell lysates.
RNA immunoprecipitation (RIP)
assay
RIP experiments were performed using a Magna RIP RNA
Binding Protein Immunoprecipitation kit (cat. no. 17-700; EMD
Millipore) according to the manufacturer's protocol. The Argonaute
2 (AGO2)-RIP assay was conducted using lysates of H9c2 cells.
Briefly, 1×107 cells were pelleted at 3,000 × g for 10
min at 4°C and resuspended with an equal volume of RIP lysis buffer
(100 µl) containing protease and RNase inhibitors. Cell lysates
(100 µl) were incubated with 5 µg control IgG antibody (cat. no.
ab182931; Abcam) or anti-AGO2 antibody (cat. no. ab32381;
Abcam)-coated protein A/G magnetic beads at 4°C overnight, with
constant rotation. After treating with proteinase K at 55°C for 30
min, the immunoprecipitated RNAs were extracted using a RNeasy
MinElute Cleanup kit (Qiagen, Inc.) and reverse transcribed as
described above for the RT-qPCR experiments. The expression levels
of miR-181a and ANRIL were analyzed using RT-qPCR as described
above.
Luciferase reporter assay
The putative targets of miRNAs were predicted using
in silico computational tools, including starBase (v2.0;
http://starbase.sysu.edu.cn/starbase2) and TargetScan
(http://www.targetscan.org/vert_72).
The wild-type (wt) 3′untranslated region (UTR) of ANRIL and SIRT1
containing the putative miR-181a binding sites was amplified by PCR
and inserted into the pmirGLO Dual-luciferase miRNA Target
Expression Vector (Promega Corporation). The clone primers are as
follows: ANRIL forward, 5′-ATCGATAGCGATAAGATCTCATTGCTCTATCCGCCAA-3′
and reverse, 5′-CGCAAGCGCAAAGAGTAGTTCAAAACTGACATTCAGC-3′; and SIRT1
forward, 5′-CAAACAAATCATAGTGTAATAA-3′ and reverse,
5′-CGCAAGCGCAAAGAGACAATCTATTTTACCAACCTAT−3′. DNA was synthesized
from 293T cells via TaqFast DNA polymerase (cat. no. T828421;
MACKLIN) and isolated using a DNA purification kit (cat. no. DP304;
Tiagen Biotech Co., Ltd.) according to the manufacturer's protocol.
Those with putative mutated binding sites (ANRIL-mut and SIRT1-mut)
were generated using the Site-Directed Mutagenesis System
(Invitrogen; Thermo Fisher Scientific, Inc.) according to the
manufacturer's instructions. The luciferase assay was performed by
co-transfecting reporter vectors along with NC mimic (NC-mimic) or
miR-181a mimic into 293T cells using Lipofectamine 2000 reagent
according to the manufacturer's protocols. Each treatment was
performed five times. Then, 2 days after transfection, firefly
luciferase activity was measured using the Dual-Luciferase Reporter
Assay System (Promega Corporation), according to the manufacturer's
instructions. The Renilla luciferase activity served as a
normalization control for each well.
Statistical analysis
All results from at least three replicates are
expressed as the mean ± SD. P-values were calculated using one-way
analysis of variance followed by Tukey's post hoc test using SPSS
19.0 software (IBM Corp.). P<0.05 was considered to indicate a
statistically significant difference.
Results
ANRIL and SIRT1 transcription levels
are decreased following H/R injury
MI/R injury is an insurmountable problem that arises
during reperfusion therapy for patients with MI (2). Despite the discovery of the
association between ANRIL polymorphisms and the risk of MI
(9,10), the expression pattern of ANRIL after
MI/R injury is still unknown. To explore this issue, MI/R injury
was mimicked using a H/R model with H9c2 cardiomyocytes cultured
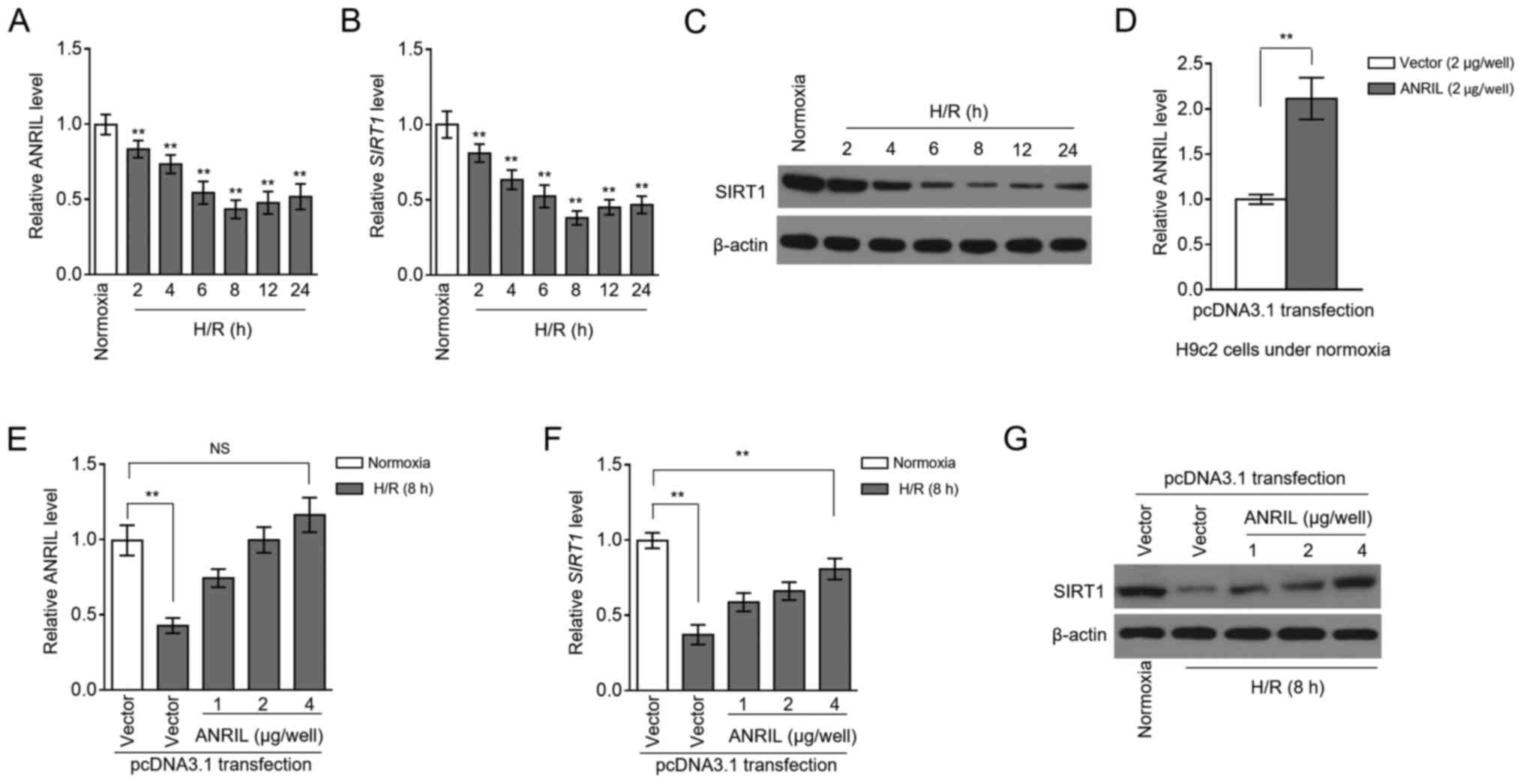
in vitro (13). RT-qPCR
analysis showed that ANRIL transcription was decreased in H9c2
cells after H/R treatment, compared with the normoxia control
(Fig. 1A). This result suggested
that ANRIL may be functionally involved in H/R-induced
cardiomyocyte pathology. Similarly, the transcription of SIRT1 was
also downregulated in H/R-treated H9c2 cells (Fig. 1B). This was consistent with the
immunoblotting analysis, which revealed that SIRT1 protein
expression also decreased following H/R treatment (Fig. 1C). In order to determine whether
ANRIL was associated with SIRT1 downregulation in H/R-treated H9c2
cells, ANRIL was overexpressed in H9c2 cells by transfecting
pcDNA3.1-ANRIL plasmids. In fact, the decreased ANRIL level in
H/R-treated H9c2 cells was completely recovered after transfection
compared with the control (Fig. 1D and
E). Moreover, consistent with restoring ANRIL expression, the
downregulation of SIRT1 expression was significantly reversed at
both the mRNA (Fig. 1F) and protein
(Fig. 1G) expression levels
following ANRIL overexpression. Together, these results suggested
that a reduction in the expression of ANRIL contributed to the
downregulation of SIRT1 expression in H9c2 cardiomyocytes following
H/R injury.
ANRIL upregulates SIRT1 expression via
sponging miR-181a
LncRNAs can serve as competitive endogenous RNAs to
sponge miRNAs and, thus, regulate the expression of downstream
targets (14). To understand how
ANRIL regulates SIRT1 expression, the computational tools starBase
(v2.0) (15) and TargetScan
(http://www.targetscan.org/vert_72/)
were used to predict candidate miRNAs. miR-181a was found to share
common complementary binding sites with the 3′-UTR of both ANRIL
and SIRT1 (Fig. 2A). Besides, RIP
showed that ANRIL and miR-181a were simultaneously enriched in
beads conjugated to Ago2 antibodies in contrast to IgG control
antibodies (Fig. 2B). Luciferase
reporter assays revealed that, compared with the NC mimics, the
miR-181a mimics transfection significantly reduced the luciferase
activity of the wt, but not from the mut ANRIL construct (Fig. 2C). The efficiency of siRNA targeting
ANRIL was confirmed in H9c2 cells under normoxia (Fig. 2D, left panel). In addition, ANRIL
overexpression in H9c2 cells resulted in decreased miR-181a
expression, and inversely, ANRIL knockdown via siRNA transfection
elevated miR-181a expression (Fig.
2D, right panel). Therefore, these results revealed that ANRIL
is capable of sponging miR-181a.
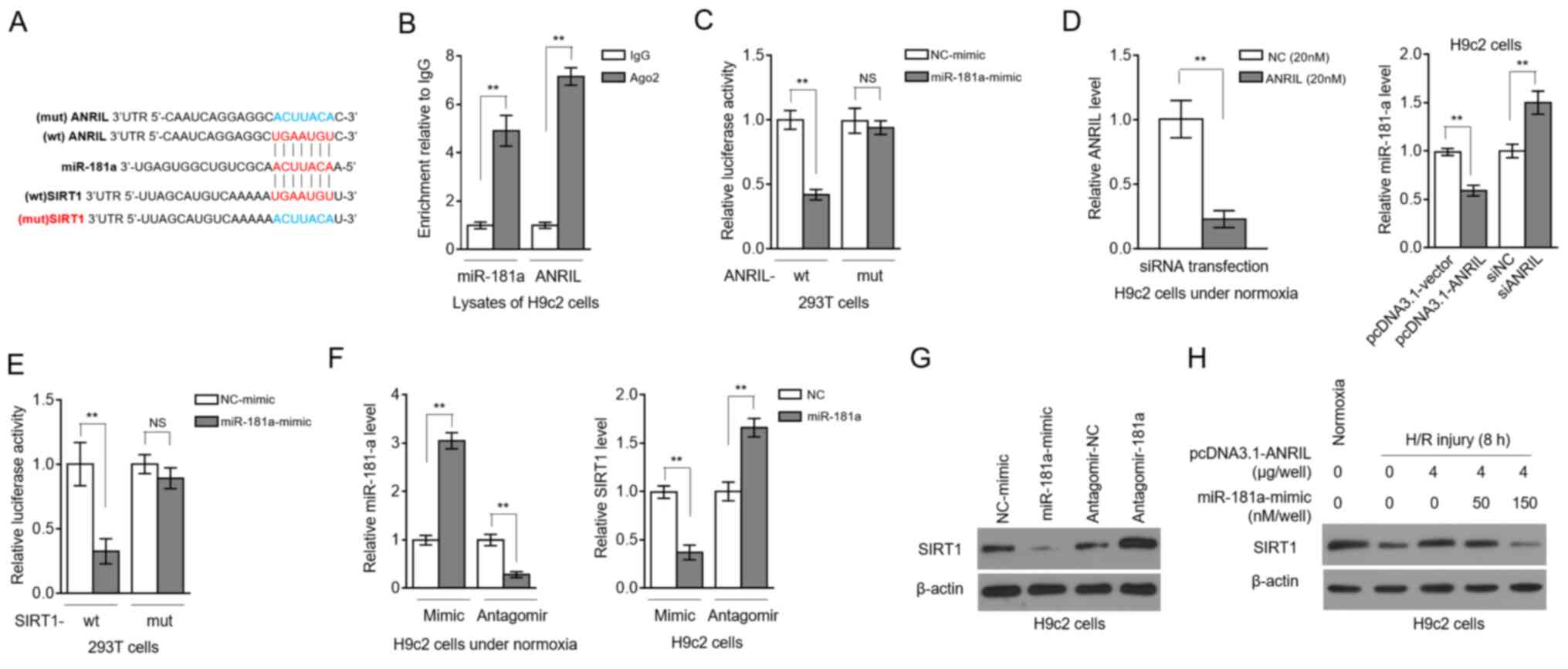 | Figure 2.ANRIL upregulates SIRT1 expression by
sponging miR-181a. (A) Putative binding sites (red) between
miR-181a and the 3′-UTR of ANRIL and SIRT1. Mutant sequences (blue)
of ANRIL were depicted at the top. (B) RNA-binding protein
immunoprecipitation assay was conducted to detect the enrichment of
miR-181a and ANRIL by using Ago2 antibody to precipitate the
lysates of H9c2 cells. Isotype IgG antibody was used as a control.
Results are expressed as relative to IgG control (n=3). (C) 293T
cells were co-transfected with NC-mimic or miR-181a mimic along
with wt or mut ANRIL luciferase reporter plasmids. Luciferase
activity was measured at 2 days after transfection. Results are
expressed as relative to NC transfection (n=5). (D) H9c2 cells were
transfected with pcDNA3.1-vector or pcDNA3.1-ANRIL plasmids, or
si-NC or siANRIL as indicated. ANRIL (left) and miR-181a (right)
expression levels were determined at 2 days after transfection. (E)
293T cells were co-transfected with NC-mimic or miR-181a mimic
along with wt or mut SIRT1 luciferase reporter plasmids. Luciferase
activity was measured at 2 days after transfection. Results are
expressed as relative to NC transfection (n=5). (F and G) H9c2
cells were transfected with NC-mimic or miR-181a mimic, or
antagomir-NC or antagomir-181a. At 2 days after transfection, the
(F) mRNA expression levels of miR-181a and SIRT1, and the (G)
protein expression levels of SIRT1 were determined (n=3). (H) H9c2
cells were transfected with pcDNA3.1-vector or pcDNA3.1-ANRIL
plasmids (4 µg per well) along with or without 50 or 150 nM
miR-181a mimic. At 2 days after transfection, cells were cultured
under normoxia or exposed to H/R (4 h hypoxia followed by 8 h
reoxygenation). SIRT1 protein expression was determined by an
immunoblotting assay (n=3). Data are presented as the mean ± SD.
**P<0.01. NS, not significant; ANRIL, antisense non-coding RNA
in the INK4 locus; SIRT1, sirtuin 1; miR, microRNA; UTR,
untranslated region; NC, negative control; wt, wild-type; mut,
mutant; si/siRNA, small interfering RNA; H/R,
hypoxia/reoxygenation. |
Luciferase reporter assays also showed that miR-181a
targeted the 3′-UTR of SIRT1 (Fig.
2E), indicating that SIRT1 is a target for miR-181 in H9c2
cells. Then, miR-181a was overexpressed or inhibited in H9c2 cells
by transfecting mimics or antagomir, respectively, and transfection
efficiency was confirmed by RT-qPCR analysis (Fig. 2F, left). The transcription level of
SIRT1 was decreased by miR-181a overexpression, and, conversely,
SIRT1 transcription was elevated upon miR-181a inhibition (Fig. 2F, right). Similar results were
obtained at the SIRT1 protein level (Fig. 2G). These results reinforced the
evidence that miR-181a targets and suppresses SIRT1 expression, at
least in H9c2 cells. Remarkably, the ANRIL-attenuated decline of
SIRT1 expression in H/R-treated H9c2 cells was completely reversed
in the presence of miR-181a overexpression (Fig. 2H). Thus, these data showed that
miR-181a was an intermediary that controlled ANRIL positive
regulation over SIRT1 after H/R injury, as observed in Fig. 1D-F.
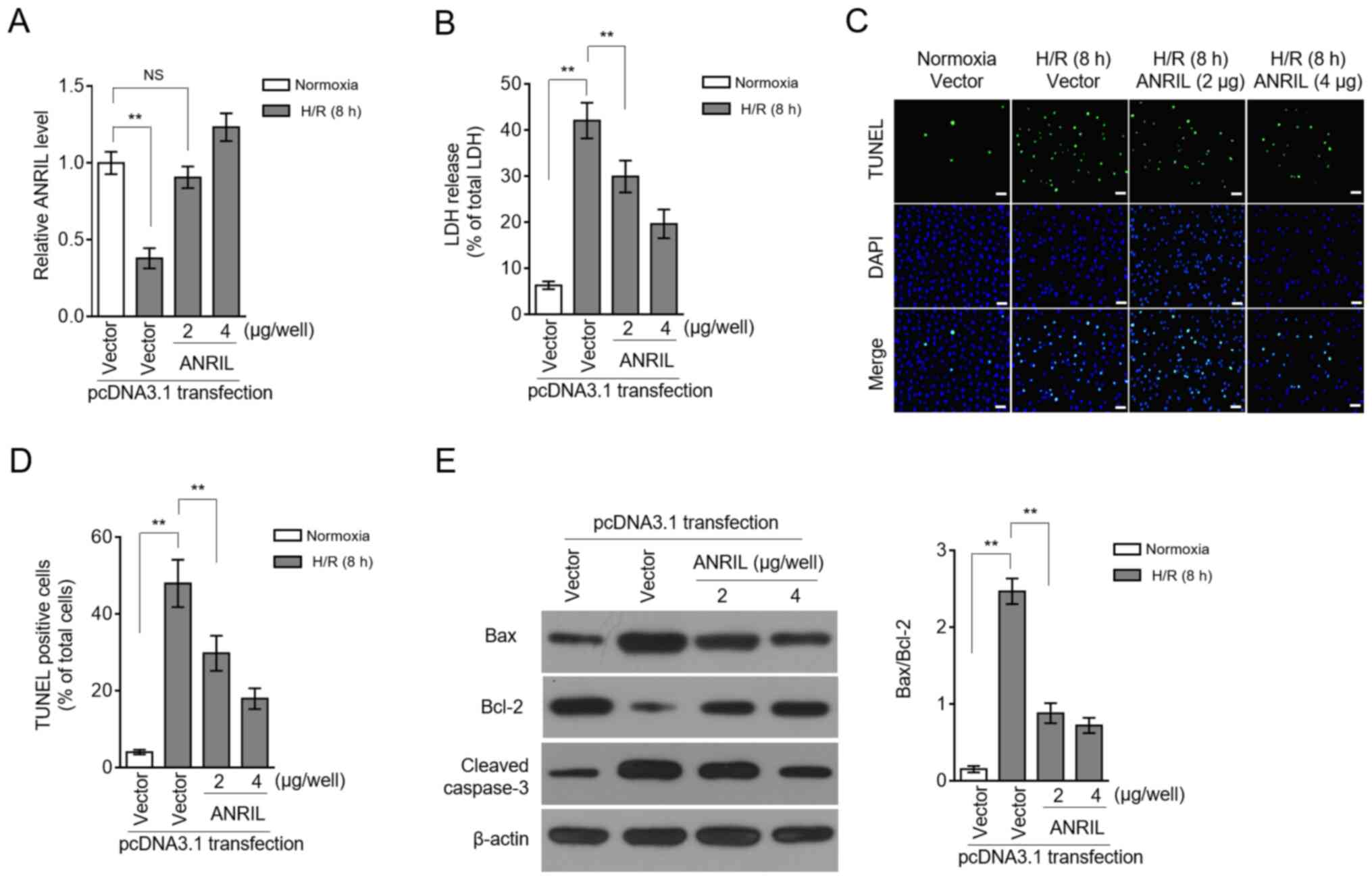
ANRIL exhibits protective effects
against H/R injury
LDH release and apoptosis are two commonly used
indicators for H/R injury (16,17).
Restoration of the ANRIL transcription level by enforced
overexpression (Fig. 3A) caused a
large decrease in the LDH release induced by H/R injury (Fig. 3B), suggesting that ANRIL may have
the ability to reduce H/R injury. This was reinforced by the
observation that ANRIL overexpression decreased the apoptotic cell
number, which had been increased in H9c2 cells upon H/R injury
(Fig. 3C and D). The
ANRIL-inhibited apoptosis of H9c2 cells was evidenced by the
reduced expression of Bax and cleaved caspase-3, and
simultaneously, by the elevated expression of Bcl-2 (Fig. 3E). These data showed that ANRIL
reduced H/R-induced LDH release and apoptosis in H9c2 cells and,
therefore, that ANRIL had protective effects against H/R
injury.
ANRIL protects against H/R injury by
regulating the miR-181a/SIRT1 axis
SIRT1 was previously reported to exhibit protective
activities against MI/R injury in various pathological scenarios
(18–21). Based on the SIRT1 regulation via
miR-181a and ANRIL revealed by the present study, it was
investigated whether miR-181a and SIRT1 could contribute to the
protective effects of ANRIL against H/R injury. Indeed, in H9c2
cells induced with H/R injury, miR-181a overexpression
significantly reversed the ANRIL-induced decrease in LDH release
(Fig. 4A), as well as the ANRIL
inhibitory effects on the apoptotic cell number (Fig. 4B) and on the expression of
pro-apoptotic markers (Fig. 4C).
Moreover, SIRT1 knockdown via siRNA (Fig. 4D) also reversed the ANRIL
alleviating effects on H/R-induced LDH release (Fig. 4E) and cell apoptosis (Fig. 4F and G) in H9c2 cells. These results
indicated that miR-181a could antagonize the protective effects of
ANRIL against H/R injury, which was associated with decreased SIRT1
expression. Along with the aforementioned observation that ANRIL
upregulated SIRT1 expression via sponging miR-181a, these data
established that ANRIL exerted its protective effects against H/R
injury by regulating the miR-181a/SIRT1 axis.
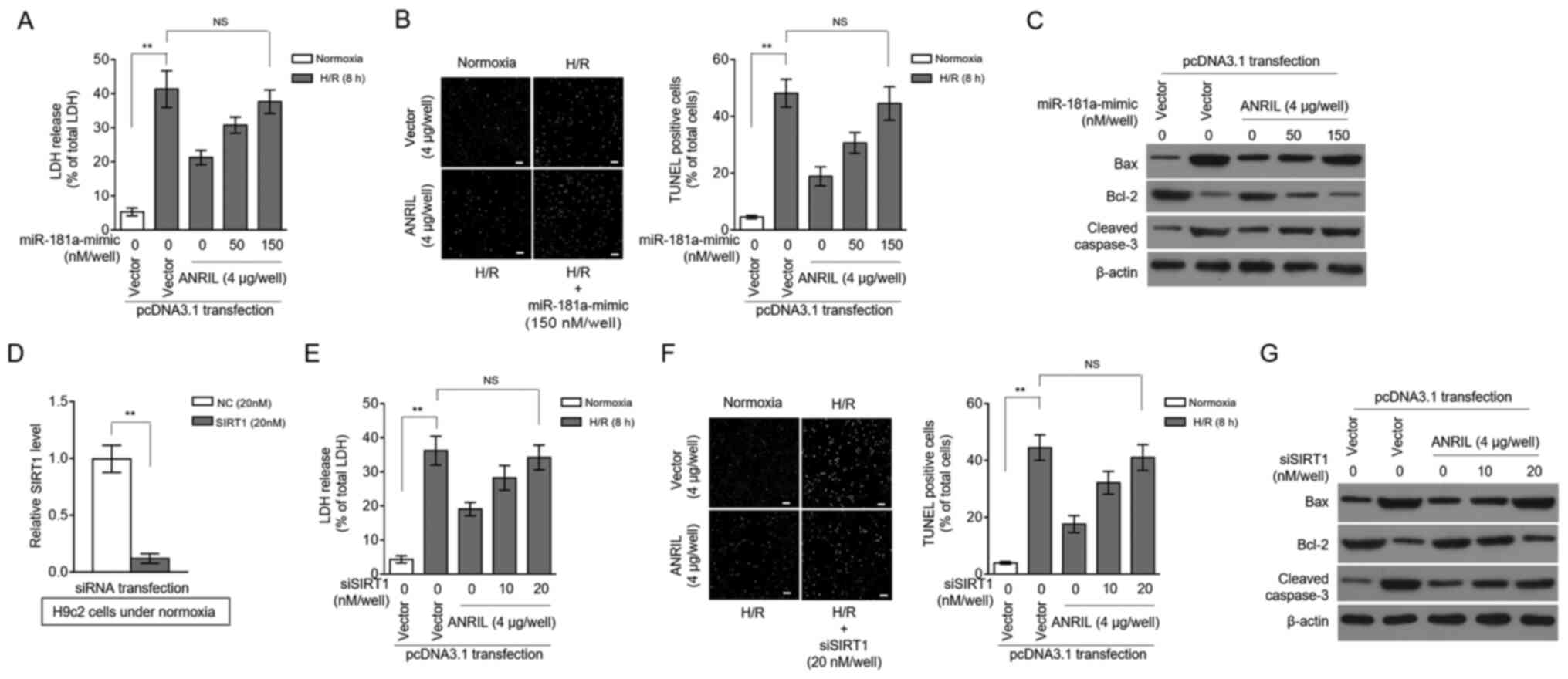 | Figure 4.ANRIL protects against H/R injury via
the miR-181a/SIRT1 axis. (A-C) H9c2 cells were transiently
transfected with pcDNA3.1-vector or pcDNA3.1-ANRIL plasmids (4 µg
per well) along with or without 50 or 150 nM miR-181a mimic. At 2
days after transfection, cells were cultured under normoxia or
exposed to H/R (4 h hypoxia followed by 8 h reoxygenation). (A) The
release of LDH from cells was measured and expressed as a
percentage of the total LDH activity (n=5). (B) Apoptosis was
detected using a TUNEL assay and defined as a percentage of TUNEL
positive cells (green) of the total number of cells (DAPI, blue)
(n=12). Scale bar, 100 µm. (C) The expression levels of Bax, Bcl-2
and cleaved caspase-3 were analyzed. (D) The efficiency of siRNA
targeting SIRT1 was confirmed in H9c2 cells under normoxia. (E-G)
H9c2 cells were transiently transfected with pcDNA3.1-vector or
pcDNA3.1-ANRIL plasmids (4 µg per well) along with or without 10 or
20 nM siSIRT1. At 2 days after transfection, cells were cultured
under normoxia or exposed to H/R (4 h hypoxia followed by 8 h
reoxygenation). (E) LDH release, (F) cell apoptosis (scale bar, 100
µm) and (G) expression of Bax, Bcl-2 and cleaved caspase-3 were
analyzed as described in A-C. Data are presented as the mean ± SD.
**P<0.01. NS, not significant; ANRIL, antisense non-coding RNA
in the INK4 locus; H/R, hypoxia/reoxygenation; SIRT1, sirtuin 1;
miR, microRNA; LDH, lactate dehydrogenase; si/siRNA, small
interfering RNA; NC, negative control. |
Discussion
In recent years, the association between lncRNAs and
MI/R injury has been increasingly recognized, which offers novel
perspectives in the discovery of potential therapeutic targets to
reduce MI/R injury (22). In the
present study, it was found that the expression levels of the
lncRNA ANRIL and SIRT1 were synchronously decreased in H9c2
cardiomyocytes after H/R-induced injury in vitro. Besides,
it was demonstrated that ANRIL upregulated SIRT1 expression by
sponging miR-181a. These data, from a mechanistic point of view,
may explain how ANRIL regulates SIRT1 expression in this in
vitro experimental system used to mimic MI/R injury.
Furthermore, the present study linked the protective effects of
ANRIL to miR-181a/SIRT1 regulation, providing a novel mechanistic
insight into the protective role of ANRIL observed following H/R
injury. As a whole, this study may have identified ANRIL as being a
novel regulator in MI/R injury pathogenesis and also highlighted
its potential to be explored as a therapeutic target for reducing
MI/R injury.
The present study observed that ANRIL was
downregulated in H9c2 cells after H/R-induced injury, which was in
line with a previous clinical study that reported lower ANRIL
expression in 414 patients with MI compared with healthy volunteers
(8). However, the exact underlying
mechanism of how ANRIL expression is regulated after H/R injury is
still unclear and needs to be elucidated by further investigation.
Of note, in the present study a decrease in the expression of SIRT
was observed to be in sync with the downregulation of ANRIL. Thus,
it was concluded that the decrease in SIRT1 expression following
H/R injury could be attributed, at least partially, to ANRIL, since
the recovery of ANRIL normal expression level restored its
expression. Consistently, the repressed SIRT1 expression was also
found in the mouse myocardium following I/R injury and
cardiomyocytes exposed to H/R in vitro (23). In addition, it was previously found
that SUV39H1 binding to the SIRT1 promoter suppresses SIRT1
transcription (24). Therefore, the
present findings uncovered a novel regulatory mechanism by which
SIRT1 was modulated in response to H/R injury.
ANRIL can regulate gene expression through a direct
epigenetic mechanism or by recruiting the polycomb repression
complex 2 (25,26). Besides, ANRIL can also act as a
miRNA sponge to execute its biological activities, such as miR-186
(27) and let-7 (28). It was demonstrated in the present
study that ANRIL upregulated SIRT1 by sponging miR-181a, thus
identifying another ANRIL target. Remarkably, two recent
investigations reinforce the present data by also documenting that
ANRIL inhibits cell senescence of vascular smooth muscle cells
(29) and promotes
lymphangiogenesis (30) by
regulating miR-181a. In conjunction with these previous studies,
the present results suggested that the negative regulation of
miR-181a by ANRIL may be a critical mechanism by which ANRIL exerts
its versatile activities.
ANRIL knockdown has been demonstrated to aggravate
H2O2-induced cell injury (31). On the other hand, ANRIL upregulation
protects lens epithelial cells against
H2O2-induced cell injury (32). Moreover, in PC-12 cells, ANRIL
reduces oxygen and glucose deprivation-induced injury (33), indicating a protective role of ANRIL
implicated in the pathogenesis of cellular oxidative and ischemic
damage. In line with these studies, the present study found that
ANRIL protected against H/R injury in H9c2 cells. This function of
ANRIL may be associated with its modulation of the miR-181a/SIRT1
axis since both miR-181a and SIRT1 knockdown diminished the
protective effects of ANRIL. These data highlighted the important
role of SIRT1 in mediating ANRIL protection against H/R injury. To
date, it is known that SIRT1 exhibits cardioprotective effects on
H/R injury by comprehensive means, including inhibition of
oxidative and endoplasmic reticulum stress (34,35),
and orchestration of autophagy induction (36). Examining the specific contribution
of these underlying mechanisms to the effects of ANRIL is one of
the future research topics that deserves further investigation.
Finally, a major limitation of the present study is
that all the observations were obtained only from H9c2 cells. The
differences in physiological aspects between mice and humans
combined with the complexity of cell involvement during MI/R
injury, require future studies to explore ANRIL activities involved
in MI/R injury using other physiologically relevant approaches and
materials in order to evaluate effects within a more complex system
than just in vitro cultured cardiomyocytes, such as other
appropriate types of cardiac cells and tissues and animal
models.
In conclusion, the present study established a key
role of the downstream regulation of the miR-181a/SIRT1 signaling
axis in mediating ANRIL protection against H/R injury in
cardiomyocytes, suggesting that targeting this axis may have a
potential therapeutic benefit for patients with MI/R injury.
Acknowledgements
Not applicable.
Funding
No funding was received.
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
BS conceived and designed the study. BS and DW
prepared the manuscript, ensured its legitimacy and revised the
final draft of the manuscript. BS, DW, GY, XS, SW, SJ, JZ, LL and
XW performed the experiments and analyzed the data. BS and DW
confirm the authenticity of all the raw data. All authors read and
approved the final manuscript.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Anderson JL and Morrow DA: Acute
myocardial infarction. N Engl J Med. 376:2053–2064. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Hausenloy DJ and Yellon DM: Myocardial
ischemia-reperfusion injury: A neglected therapeutic target. J Clin
Invest. 123:92–100. 2013. View
Article : Google Scholar : PubMed/NCBI
|
|
3
|
Yellon DM and Hausenloy DJ: Myocardial
reperfusion injury. N Engl J Med. 357:1121–1135. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Guo Y, Luo F, Liu Q and Xu D: Regulatory
non-coding RNAs in acute myocardial infarction. J Cell Mol Med.
21:1013–1023. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Ong SB, Katwadi K, Kwek XY, Ismail NI,
Chinda K, Ong SG and Hausenloy DJ: Non-coding RNAs as therapeutic
targets for preventing myocardial ischemia-reperfusion injury.
Expert Opin Ther Targets. 22:247–261. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Long Y, Wang X, Youmans DT and Cech TR:
How do lncRNAs regulate transcription? Sci Adv. 3:eaao21102017.
View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Congrains A, Kamide K, Ohishi M and Rakugi
H: ANRIL: Molecular mechanisms and implications in human health.
Int J Mol Sci. 14:1278–1292. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Vausort M, Wagner DR and Devaux Y: Long
noncoding RNAs in patients with acute myocardial infarction. Circ
Res. 115:668–677. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Ahmed W, Ali IS, Riaz M, Younas A, Sadeque
A, Niazi AK, Niazi SH, Ali SH, Azam M and Qamar R: Association of
ANRIL polymorphism (rs1333049:C>G) with myocardial infarction
and its pharmacogenomic role in hypercholesterolemia. Gene.
515:416–420. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Cheng J, Cai MY, Chen YN, Li ZC, Tang SS,
Yang XL, Chen C, Liu X and Xiong XD: Variants in ANRIL gene
correlated with its expression contribute to myocardial infarction
risk. Oncotarget. 8:12607–12619. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Li X, Dai Y, Yan S, Shi Y, Han B, Li J,
Cha L and Mu J: Down-regulation of lncRNA KCNQ1OT1 protects against
myocardial ischemia/reperfusion injury following acute myocardial
infarction. Biochem Biophys Res Commun. 491:1026–1033. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Zhang YQ, Tang Y, Wu AL and Zhu HB:
Salvianolic acid A displays cardioprotective effects in in vitro
models of heart hypoxia/reoxygenation injury. J Asian Nat Prod Res.
12:899–915. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Tay Y, Rinn J and Pandolfi PP: The
multilayered complexity of ceRNA crosstalk and competition. Nature.
505:344–352. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Li JH, Liu S, Zhou H, Qu LH and Yang JH:
StarBase v2.0: Decoding miRNA-ceRNA, miRNA-ncRNA and protein-RNA
interaction networks from large-scale CLIP-Seq data. Nucleic Acids
Res. 42((Database Issue)): D92–D97. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
He Y, Li C, Ma Q and Chen S: Esculetin
inhibits oxidative stress and apoptosis in H9c2 cardiomyocytes
following hypoxia/reoxygenation injury. Biochem Biophys Res Commun.
501:139–144. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Wang X, Ha T, Hu Y, Lu C, Liu L, Zhang X,
Kao R, Kalbfleisch J, Williams D and Li C: MicroRNA-214 protects
against hypoxia/reoxygenation induced cell damage and myocardial
ischemia/reperfusion injury via suppression of PTEN and Bim1
expression. Oncotarget. 7:86926–86936. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Becatti M, Taddei N, Cecchi C, Nassi N,
Nassi PA and Fiorillo C: SIRT1 modulates MAPK pathways in
ischemic-reperfused cardiomyocytes. Cell Mol Life Sci.
69:2245–2260. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Yang Y, Duan W, Lin Y, Yi W, Liang Z, Yan
J, Wang N, Deng C, Zhang S, Li Y, et al: SIRT1 activation by
curcumin pretreatment attenuates mitochondrial oxidative damage
induced by myocardial ischemia reperfusion injury. Free Radic Biol
Med. 65:667–679. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Shalwala M, Zhu SG, Das A, Salloum FN, Xi
L and Kukreja RC: Sirtuin 1 (SIRT1) activation mediates sildenafil
induced delayed cardioprotection against ischemia-reperfusion
injury in mice. PLoS One. 9:e869772014. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Ding M, Lei J, Han H, Li W, Qu Y, Fu E, Fu
F and Wang X: SIRT1 protects against myocardial
ischemia-reperfusion injury via activating eNOS in diabetic rats.
Cardiovasc Diabetol. 14:1432015. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Yu SY, Tang L and Zhou SH: Long noncoding
RNAs: New players in ischaemia-reperfusion injury. Heart Lung Circ.
27:322–332. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Guo Y, Zhang L, Li F, Hu CP and Zhang Z:
Restoration of sirt1 function by pterostilbene attenuates
hypoxia-reoxygenation injury in cardiomyocytes. Eur J Pharmacol.
776:26–33. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Yang G, Zhang X, Weng X, Liang P, Dai X,
Zeng S, Xu H, Huan H, Fang M, Li Y, et al: SUV39H1 mediated SIRT1
trans-repression contributes to cardiac ischemia-reperfusion
injury. Basic Res Cardiol. 112:222017. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Yap KL, Li S, Muñoz-Cabello AM, Raguz S,
Zeng L, Mujtaba S, Gil J, Walsh MJ and Zhou MM: Molecular interplay
of the noncoding RNA ANRIL and methylated histone H3 lysine 27 by
polycomb CBX7 in transcriptional silencing of INK4a. Mol Cell.
38:662–674. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Kotake Y, Nakagawa T, Kitagawa K, Suzuki
S, Liu N, Kitagawa M and Xiong Y: Long non-coding RNA ANRIL is
required for the PRC2 recruitment to and silencing of p15(INK4B)
tumor suppressor gene. Oncogene. 30:1956–1962. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Zhang JJ, Wang DD, Du CX and Wang Y: Long
noncoding RNA ANRIL promotes cervical cancer development by acting
as a sponge of miR-186. Oncol Res. 26:345–352. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Wang Y, Cheng N and Luo J: Downregulation
of lncRNA ANRIL represses tumorigenicity and enhances
cisplatin-induced cytotoxicity via regulating microRNA let-7a in
nasopharyngeal carcinoma. J Biochem Mol Toxicol. 312017.doi:
10.1002/jbt.21904.
|
|
29
|
Tan P, Guo YH, Zhan JK, Long LM, Xu ML, Ye
L, Ma XY, Cui XJ and Wang HQ: LncRNA-ANRIL inhibits cell senescence
of vascular smooth muscle cells by regulating miR-181a/Sirt1.
Biochem Cell Biol. 97:571–580. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
He ZY, Wei TH, Zhang PH, Zhou J and Huang
XY: Long noncoding RNA-antisense noncoding RNA in the INK4 locus
accelerates wound healing in diabetes by promoting
lymphangiogenesis via regulating miR-181a/Prox1 axis. J Cell
Physiol. 234:4627–4640. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Li R, Yin F, Guo YY, Zhao KC, Ruan Q and
Qi YM: Knockdown of ANRIL aggravates
H2O2-induced injury in PC-12 cells by
targeting microRNA-125a. Biomed Pharmacother. 92:952–961. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Qi D, Wang M, Zhang D and Li H: Tanshinone
IIA protects lens epithelial cells from
H2O2-induced injury by upregulation of lncRNA
ANRIL. J Cell Physiol. Jan 30–2019.(Epub ahead of print).
View Article : Google Scholar
|
|
33
|
Liu B, Cao W and Xue J: LncRNA ANRIL
protects against oxygen and glucose deprivation (OGD)-induced
injury in PC-12 cells: Potential role in ischaemic stroke. Artif
Cells Nanomed Biotechnol. 47:1384–1395. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Li YP, Wang SL, Liu B, Tang L, Kuang RR,
Wang XB, Zhao C, Song XD, Cao XM, Wu X, et al: Sulforaphane
prevents rat cardiomyocytes from hypoxia/reoxygenation injury in
vitro via activating SIRT1 and subsequently inhibiting ER stress.
Acta Pharmacol Sin. 37:344–353. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Yang H, Wang C, Zhang L, Lv J and Ni H:
Rutin alleviates hypoxia/reoxygenation-induced injury in myocardial
cells by up-regulating SIRT1 expression. Chem Biol Interact.
297:44–49. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Qiu R, Li W and Liu Y: MicroRNA-204
protects H9C2 cells against hypoxia/reoxygenation-induced injury
through regulating SIRT1-mediated autophagy. Biomed Pharmacother.
100:15–19. 2018. View Article : Google Scholar : PubMed/NCBI
|