Introduction
Lung cancer is one of the most common types of
cancer and is the leading cause of cancer-associated death
worldwide (1–3). Lung cancer is classified into two
groups based on the pathological features: Non-small cell lung
cancer (NSCLC) and small cell lung cancer (SCLC) (4,5). NSCLC
contributes to ~80% of lung cancer diagnoses (6,7).
Additionally, ~75% of patients with lung cancer are already at the
advanced stage at diagnosis as they are usually asymptomatic at an
early stage. Despite improvements having been made in diagnosis and
treatment, the prognosis of lung cancer remains poor and the 5-year
survival rate is low (8,9). Therefore, there is an urgent need to
develop effective strategies for the treatment of lung cancer.
Long non-coding RNAs (lncRNAs) are defined as
non-protein coding transcripts >200 nucleotides in length.
Previous studies have shown that lncRNAs serve a crucial role in a
variety of biological processes, including energy metabolism,
apoptosis, cell proliferation and differentiation, through
transcriptional and posttranscriptional regulation (10,11).
Notably, an increasing number of studies have revealed that lncRNAs
are involved in the development and progression of multiple types
of cancer, including breast cancer, gastric cancer, prostate
cancer, lung cancer and leukemia (12,13).
Although several lncRNAs, including ZEB1 antisense RNA 1 (14), breast cancer anti-estrogen
resistance 4 (15), small nucleolar
RNA host gene 1 (16), growth
arrest specific 5 (GAS5) (17) and
deleted in lymphocytic leukemia 2 (18) have been explored previously, the
functions and underlying mechanisms of lncRNAs in NSCLC have not
been fully revealed.
LncRNA HLA complex group 11 (HCG11) is located on
chromosome 6 and is a member of the lncRNA family. It has been
reported that HCG11 functions as a tumor suppressor in
hepatocellular carcinoma and prostate cancer (19) by inhibiting cell proliferation,
invasion and metastasis, and promoting cell apoptosis.
Additionally, abnormal HCG11 expression has been observed in
gastric cancer (20). In
particular, downregulation of HCG11 expression is predictive of a
poor prognosis in prostate cancer (21). However, whether HCG11 is involved in
the development and progression of NSCLC remains unclear.
The present study primarily explored the role of
HCG11 in NSCLC. First, it was identified that HCG11 expression was
significantly reduced in patients with NSCLC. Further
investigations revealed that low HCG11 expression was associated
with a poor prognosis in patients with NSCLC. In terms of the
mechanism, it was demonstrated that HCG11 could inhibit the
malignant phenotype of NSCLC cells, at least partially, by
targeting a microRNA-875 (miR-875)/special AT-rich sequence-binding
protein 2 (SATB2) signaling pathway. Collectively, the results of
the present study indicated that HCG11 is an important tumor
suppressor, and thus, provides a novel potential avenue for the
treatment of NSCLC.
Materials and methods
Patients and specimens
In the present study, 85 pairs of NSCLC tissues and
adjacent normal tissues were collected from patients. The patients
included 47 males and 38 females, with a median age of 59.5 years
(range, 43–79 years), and were enrolled from The Sichuan Mianyang
404 Hospital (Mianyang, China) between January 2012 and June 2015.
The patients did not receive any preoperative treatments, and
patients who had received anticancer treatment were excluded from
the present study. Tumor-node-metastasis (TNM) stage was determined
as previously described (22). All
patients provided informed consent. The present study was approved
by the Ethics Committee of The Sichuan Mianyang 404 Hospital
(Mianyang, China).
Cell culture and transfection
A total of five NSCLC cell lines (A549, H1299, H466,
H460 and H358) and 16HBE normal lung epithelial cells were obtained
from The Cell Bank of Type Culture Collection of The Chinese
Academy of Sciences. All cell lines were cultured in DMEM (HyClone;
Cytiva) supplemented with 10% FBS (Gibco; Thermo Fisher Scientific,
Inc.), 100 U/ml penicillin and 100 µg/ml streptomycin (Gibco;
Thermo Fisher Scientific, Inc.). All cells were cultured in a 37°C
incubator with 5% CO2. pcDNA3.1-HCG11 (pcDNA-HCG11) and
corresponding negative control (pcDNA3.1-NC; pcDNA3.1-vector) were
synthesized by Shanghai Gene Pharma Co., Ltd. miR-875-mimic, as
well as the corresponding negative control (miR-NC), were purchased
from Guangzhou RiboBio Co., Ltd. Cells were transfected using
Lipofectamine® 3000 (Invitrogen; Thermo Fisher
Scientific, Inc.) according to the manufacturer's protocols.
MTT assay
A549 cells were seeded in 96-well plates
(2×103 cells/well). After 24 h of culture, 500 µg/ml MTT
solution (Sigma-Aldrich; Merck KGaA) was added to each well,
followed by incubation at 37°C for 4 h. Subsequently, 200 µl DMSO
(Sigma-Aldrich; Merck KGaA) was added to each well. Finally, the
absorbance of the samples was measured at 570 nm using a microplate
reader (Bio-Rad Laboratories, Inc.).
Cell invasion assay
Cell invasion assays were performed using a
Transwell insert (6.5 mm in diameter) with an 8-µm pore size
(Corning, Inc.). Briefly, 5×104 cells in DMEM without
FBS were seeded onto the Matrigel-coated membrane matrix in the
upper chamber, and the lower chamber was filled with medium
supplemented with 10% FBS. Following incubation at 37°C for 24 h,
cells which had not invaded were removed, and cells on the bottom
of the filter, which had invaded, were fixed in 4% paraformaldehyde
at room temperature and stained with crystal violet at room
temperature for 30 min. The number of invaded cells were counted
using a microscope.
Cell migration assay
Migration assays were performed using
Matrigel-coated Transwell chambers (BD Biosciences). Briefly,
1×105 cells were seeded in the upper chamber in FBS free
DMEM, and DMEM supplemented with 10% FBS was added to the lower
chamber. Following incubation at 37°C for 24 h, cells in the upper
chamber were removed using a cotton swab. The membrane was then
soaked in 90% ethanol, fixed at 37°C for 10 min, and stained with
0.2% crystal violet at room temperature for 30 min. The cells which
had migrated were counted under an inverted light microscope
(magnification, ×200).
Cell apoptosis analysis
Cell apoptosis was detected using an Annexin
V-FITC/7-AAD kit (BD Biosciences) according to the manufacturer's
protocol, followed by flow cytometry (FACSCantoII; BD Biosciences).
Flow data were analyzed using FlowJo version 10.0 (FlowJo LLC).
Luciferase reporter assay
A luciferase reporter assay was performed using a
dual luciferase reporter assay kit (Promega Corporation) according
to the manufacturer's protocol. Briefly, pmirGLO vector containing
wild-type or mutant HCG11 or SATB2 (synthesized by Sangon Biotech
Co., Ltd.) and miR-875-mimic or miR-NC were co-transfected into
293T cells obtained from The Cell Bank of Type Culture Collection
of The Chinese Academy of Sciences using Lipofectamine®
3000 (Invitrogen; Thermo Fisher Scientific, Inc.). The luciferase
activity was measured 48 h after transfection.
Reverse transcription-quantitative PCR
(RT-qPCR)
Total RNA was extracted using TRIzol®
reagent (Invitrogen; Thermo Fisher Scientific, Inc.) according to
the manufacturer's protocols, and then converted to cDNA using the
PrimeScript RT kit (Takara Bio, Inc.) at 37°C for 15 min followed
by 85°C for 5 sec. Next, qPCR was performed using the SYBR Premix
Ex Taq kit (Takara Bio, Inc.) with the following thermocycling
conditions: Initial denaturation at 95°C for 3 min, followed by 40
cycles of denaturation at 95°C for 5 sec, annealing at 60°C for 30
sec and extension at 72°C for 30 sec. GAPDH was used as the
internal control. The primer sequences used were: HCG11 forward,
5′-GCTCTATGCCATCCTGCTT-3′ and reverse, 5′-TCCCATCTCCATCAACCC-3′;
GAPDH forward, 5′-TGTTCGTCATGGGTGTGAAC-3′ and reverse,
5′-ATGGCATGGACTGTGGTCAT-3′; and SATB2 forward,
5′-GGAGAACGACAGCGAGGAA-3′ and reverse,
5′-CCGATGTATTGCTTTGCCTAGT-3′. The expressions levels of miRNAs were
measured using the mirVana RT-qPCR miRNA Detection kit, according
to the manufacturer's protocol (Invitrogen; Thermo Fisher
Scientific, Inc.). The miRNA primers were purchased from Guangzhou
RiboBio Co., Ltd. Small RNA U6 was used as an internal reference
gene. The miRNA primer sequences used were: miR-875 forward
5′-CGAATGGGCCTAAGATCCCG-3′ and reverse 5′-GGAGCCCAGCACTTTGATCT-3′;
miR-522 forward, 5′-ACACTCCAGCTGGGCTCTAGAGGGAAGCGC-3′ and reverse,
5′-TGGTGTCGTGGAGTCG-3′; miR-499a forward,
5′-TGCGGTGGCAGTGTATTGTTAGC-3′ and reverse,
5′-CCAGTGCAGGGTCCGAGGT-3′; miR-224 forward,
5′-GTATACTAAGTCACTAGTGGT-3′ and reverse, 5′-GTGCAGGGTCCGAGGT-3′;
miR-483 forward, 5′-GCTGACTCACTCCTCCCCTC-3′ and reverse,
5′-TATGGTTGTTCACGACTCCTTCAC-3′; miR-214 forward,
5′-TCCTTCAATCACCAAATCTGG-3′ and reverse,
5′-CCCAAGCTTTCATTCAGGCTGGGTTG-3′; miR-26a forward,
5′-GGATCCGCAGAAACTCCAGAGAGAAGGA-3′ and reverse,
5′-AAGCTTGCCTTTAGCAGAAAGGAGGTT-3′; miR-455 forward,
5′-ACACTCCAGCTGGGGCAGTCCATGGGCAT-3′ and reverse,
5′-TGGTGTCGTGGAGTCG-3′; miR-543 forward, 5′-GTGCTCGGTTTGTAGGCAGT-3′
and reverse, 5′-GTGCCTTGTTTTGATGGCAG-3′; miR-155 forward,
5′-TTAATGCTAATCGTGATAG-3′ and reverse, 5′-ACCTGAGAGTAGACCAGA-3′;
miR-105 forward, 5′-GCCCTCGAGATACCATATCTATCCCCTTTTTCA-3′ and
reverse, 5′-GCCGAATTCCAACCATGAAGATACGAATTGATG-3′; and U6 forward,
5′-CTCGCTTCGGCAGCACA-3′ and reverse 5′-AACGCTTCACGAATTTGCGT-3′.
Western blotting
Cells were lysed using RIPA buffer (Invitrogen;
Thermo Fisher Scientific, Inc.). The concentration of proteins was
determined using a Bicinchoninic Acid Protein Assay kit (Beyotime
Institute of Biotechnology). Proteins (30 µg/lane) were loaded on a
10% SDS-gel, resolved using SDS-PAGE, and then electrotransferred
onto PVDF membranes for 90 min. Subsequently, 5% skimmed milk was
used to block the membranes at 37°C for 1 h, and the membranes were
incubated with the primary antibodies at 4°C overnight. The
following primary antibodies were used: Proliferation cell nuclear
antigen (PCNA; cat. no. 13110S; 1:1,000; Cell Signaling Technology,
Inc.), cleaved-caspase 3 (cat. no. 9664S; 1:1,000; Cell Signaling
Technology, Inc.), cleaved-caspase-9 (cat. no. 9505S; 1:1,000; Cell
Signaling Technology, Inc.), caspase-3 (cat. no. 14220S; 1:1,000;
Cell Signaling Technology, Inc.), caspase-9 (cat. no. 9502S;
1:1,000; Cell Signaling Technology, Inc.), SATB2 (cat. no. ab92446;
1:1,000; Abcam) and β-actin (cat. no. ab8227; 1:2,000; Abcam).
Signals were visualized using ECL.
Animal experiments
A total of 16 male BALB/c nude mice (4–5 weeks old,
16–20 g) were obtained from Beijing Vital River Laboratory Animal
Technology Co., Ltd. All mice were housed in a SPF-grade animal
room (temperature 18–22°C; humidity 40–60%; light/dark cycle 12/12
h each day) and had free access to food and water. HCG11 was cloned
and inserted into pHBLV-U6-mcs-CMV-PURO lentiviral vectors (Hanbio
Biotechnology Co., Ltd.). A second-generation lentiviral vector
system was used. The recombinant lentivirus was generated by
co-transfection of 293T cells (The Cell Bank of Type Culture
Collection of The Chinese Academy of Sciences) with pHBLV
lentiviral plasmid (10 µg), psPAX2 packaging plasmid (10 µg) and
pMD2.G envelope plasmid (5 µg; Hanbio Biotechnology Co., Ltd.)
using Lipofectamine® 3000 (Invitrogen; Thermo Fisher
Scientific, Inc.). After 48 h, the virus was filtered through 0.22
µm cellulose acetate filters (Millipore Corp.) and concentrated by
ultracentrifugation (50,000 × g) for 2 h at 4°C. Lentivirus
carrying HCG11 or negative control was stably transfected into the
A549 cell line (MOI, 20) at 37°C in the presence of 5 µg/ml
polybrene (Hanbio Biotechnology Co., Ltd.). At 24 h after
transfection, cultures were washed with freshly DMEM containing 10%
FBS. After another 48 h, transfected cells were subcutaneously
injected into BALB/c nude mice. Euthanasia was performed using
pentobarbital sodium (100 mg/kg intraperitoneal injection) at the
end of the study. The tumor volume was measured once every 5 days.
A total of 30 days after implantation, all mice were sacrificed,
and tumor tissues were collected and weighed. Finally, tumor
tissues were used for hematoxylin and eosin (H&E) staining
(room temperature for 10 sec) and immunohistochemical analysis of
Ki-67 expression. All animal experiments performed in this study
were approved by the Animal Care and Use Committee of Sichuan
Mianyang 404 Hospital (Mianyang, China).
Immunohistochemical staining
Paraffin-embedded tumor tissues were sliced (0.25
µm), dewaxed using xylene and rehydrated using a series of
decreasing concentrations of ethanol solutions, and then incubated
with anti-Ki-67 antibody (cat. no. ab16667; 1:250; Abcam) at 4°C
overnight. Subsequently, tissue sections were incubated with the
goat anti-rabbit IgG secondary antibody (cat. no. A0208; 1:50;
Beyotime Institute of Biotechnology) at 37°C for 1 h. After
counterstaining with hematoxylin at room temperature for 3 min,
color development was performed using DAB (Beyotime Institute of
Biotechnology).
Bioinformatics
The prediction of the interaction between HCG11 and
miR-875 was performed using stabase3.0 (starbase.sysu.edu.cn). The prediction of the
interaction between miR-875 and SATB2 was performed using
TargetScan (targetscan.org/vert_71/).
Statistical analysis
Statistical analysis was performed using SPSS
version 19.0 (IBM Corp.) and GraphPad Prism version 6.0 (GraphPad
Software, Inc.) software. A Student's t-test (paired or
unpaired) or one-way ANOVA followed by a Tukey-Kramer post hoc test
were used to analyze differences amongst groups. A χ2
test was used to examine the association between HCG11 expression
and the clinical characteristics. Cox regression models (univariate
and multivariate) were used to evaluate the factors related to
overall survival. Patients were divided into high and low HCG11
expression groups based on the median expression levels; overall
survival was analyzed using the Kaplan-Meier method with the
log-rank test. The correlation between the expression levels of
HCG11 and miR-875 were analyzed using Pearson's correlation
coefficient. All experiments were independently performed at least
three times. Data are presented as the mean ± standard deviation.
P<0.05 was considered to indicate a statistically significant
difference.
Results
HCG11 expression is downregulated in
NSCLC tissues and cell lines
To assess the role of HCG11 in NSCLC, tumor and
adjacent normal tissues from NSCLC patients were collected.
Subsequently, RT-qPCR analysis showed that the expression levels of
HCG11 were significantly decreased in NSCLC tissues compared with
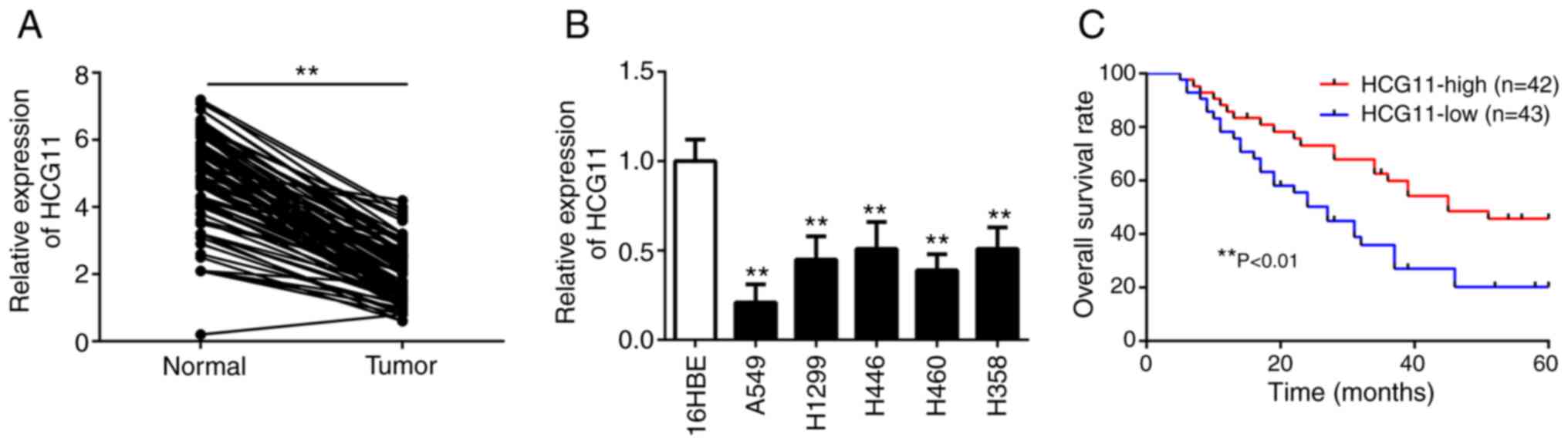
adjacent normal tissues (Fig. 1A).
Additionally, the expression levels of HCG11 were significantly
lower in NSCLC cell lines, particularly in A549 cells, compared
with the normal lung epithelial cells (16HBE) (Fig. 1B). To assess the clinical
significance of HCG11 expression in NSCLC, patients were further
divided into high and low HCG11 expression groups based on the
median expression levels. It was found that HCG11 expression was
significantly related to tumor size and TNM stage (Table I). However, there was no significant
association between HCG11 expression with age, sex, smoking
history, tumor differentiation and histological subtype (Table I). In addition, Kaplan-Meier
analysis showed that patients with high HCG11 expression had a
higher overall survival rate than those with low HCG11 expression
(Fig. 1C). In the multivariate
analysis, low HCG11 expression was an independent factor in
predicting a poor overall survival of NSCLC patients (Table II). These data suggest that HCG11
may serve a key role in regulating the development and progression
of NSCLC.
 | Table I.Correlation between HLA complex group
11 expression and clinical characteristics in non-small cell lung
cancer patients. |
Table I.
Correlation between HLA complex group
11 expression and clinical characteristics in non-small cell lung
cancer patients.
|
|
| HCG11
expression |
|
|---|
|
|
|
|
|
|---|
| Variable | n | High, n=42 | Low, n=43 | P-value |
|---|
| Age |
|
|
| 0.452 |
|
<60 | 37 | 20 | 17 |
|
|
≥60 | 48 | 22 | 26 |
|
| Sex |
|
|
| 0.593 |
|
Male | 47 | 22 | 25 |
|
|
Female | 38 | 20 | 18 |
|
| Smoking
history |
|
|
| 0.16 |
|
Yes | 45 | 19 | 26 |
|
| No | 40 | 23 | 17 |
|
| Tumor size |
|
|
| 0.006a |
| >3
cm | 35 | 11 | 24 |
|
| ≤3
cm | 50 | 31 | 19 |
|
|
Differentiation |
|
|
| 0.208 |
|
Well/moderate | 53 | 29 | 24 |
|
|
Poor | 32 | 13 | 19 |
|
| Histological
subtype |
|
|
| 0.622 |
|
Adenocarcinoma | 31 | 17 | 14 |
|
|
Squamous cell carcinoma | 43 | 19 | 24 |
|
|
Others | 11 | 6 | 5 |
|
|
Tumor-Node-Metastasis stage |
|
|
| 0.003a |
|
I/II | 51 | 32 | 19 |
|
|
III/IV | 34 | 10 | 24 |
|
| Table II.Univariate and multivariate analysis
of variables for overall survival in non-small cell lung cancer
patients. |
Table II.
Univariate and multivariate analysis
of variables for overall survival in non-small cell lung cancer
patients.
|
| Univariate
analysis | Multivariate
analysis |
|---|
|
|
|
|
|---|
| Variables | HR (95% CI) | P-value | HR (95% CI) | P-value |
|---|
| Age | 1.085
(0.683–1.945) | 0.282 | – | – |
| Sex | 0.967
(0.544–1.876) | 0.368 | – | – |
| Smoking
history | 1.636
(0.795–2.418) | 0.149 | – | – |
| Tumor size | 2.978
(1.513–4.751) | 0.004b | 2.668
(1.243–4.421) | 0.012a |
|
Differentiation | 1.785
(0.832–2.735) | 0.117 | – | – |
| Histological
subtype | 1.213
(0.508–2.047) | 0.456 | – | – |
|
Tumor-Node-Metastasis stage | 3.254
(1.362–4.356) | 0.011a | 2.874
(1.193–4.205) | 0.025a |
| HLA complex group
11 expression | 0.356
(0.224–0.643) | 0.001b | 0.435
(0.268–0.781) | 0.008b |
HCG11 inhibits proliferation, invasion
and migration, and induces apoptosis of NSCLC cells in vitro
To investigate the function of HCG11 in NSCLC, a
HCG11 overexpression plasmid was transfected into the A549 cell
line (Fig. 2A). MTT analysis
revealed that overexpression of HCG11 significantly reduced cell
proliferation (Fig. 2B).
Additionally, flow cytometry analysis showed that overexpression of
HCG11 increased the apoptosis of NSCLC cells (Fig. 2C). Consistent with these findings,
the expression levels of PCNA were decreased, and the expression
levels of apoptosis-associated proteins (cleaved-caspase-3 and
cleaved-caspase-9) were increased in A549 cells following
overexpression of HCG11 (Fig. 2D).
Furthermore, the results of Transwell assays showed that HCG11
overexpression significantly reduced the invasion and migration of
NSCLC cells (Fig. 2E and F).
Therefore, HCG11 may inhibit proliferation, invasion and migration,
and induce apoptosis of NSCLC cells.
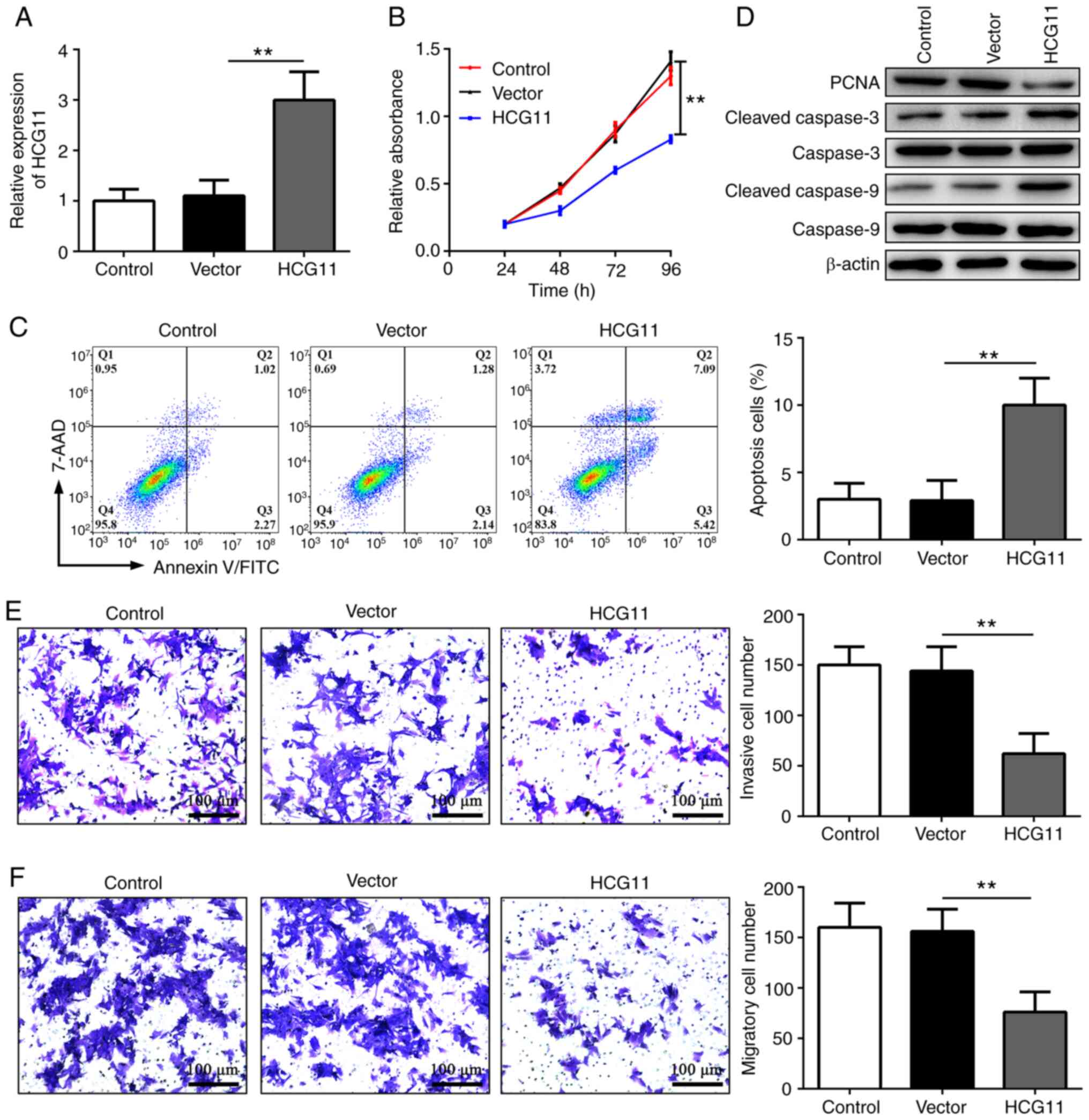 | Figure 2.HCG11 inhibits proliferation,
invasion and migration, and induces apoptosis of non-small cell
lung cancer cells in vitro. (A) Reverse transcription-quantitative
PCR analysis of the expression levels of HCG11 in the Control,
Vector and HCG11 groups. (B) MTT analysis of cell proliferation in
the Control, Vector and HCG11 groups. (C) Flow cytometry analysis
of cell apoptosis in the Control, Vector and HCG11 groups. (D)
Western blot analysis of the protein expression of PCNA,
cleaved-caspase-3 and cleaved-caspase-9 in the Control, Vector and
HCG11 groups. (E and F) Transwell invasion assays showing (E) cell
invasion and (F) migration in the Control, Vector and HCG11 groups.
**P<0.01. HCG11, HLA complex group 11; PCNA, proliferation cell
nuclear antigen; Vector, empty vector control. |
HCG11 suppresses NSCLC tumor growth in
vivo
To further validate the effects of HCG11 on NSCLC,
A549 cells transfected with HCG11 lentivirus were subcutaneously
injected into BALB/c nude mice (Fig.
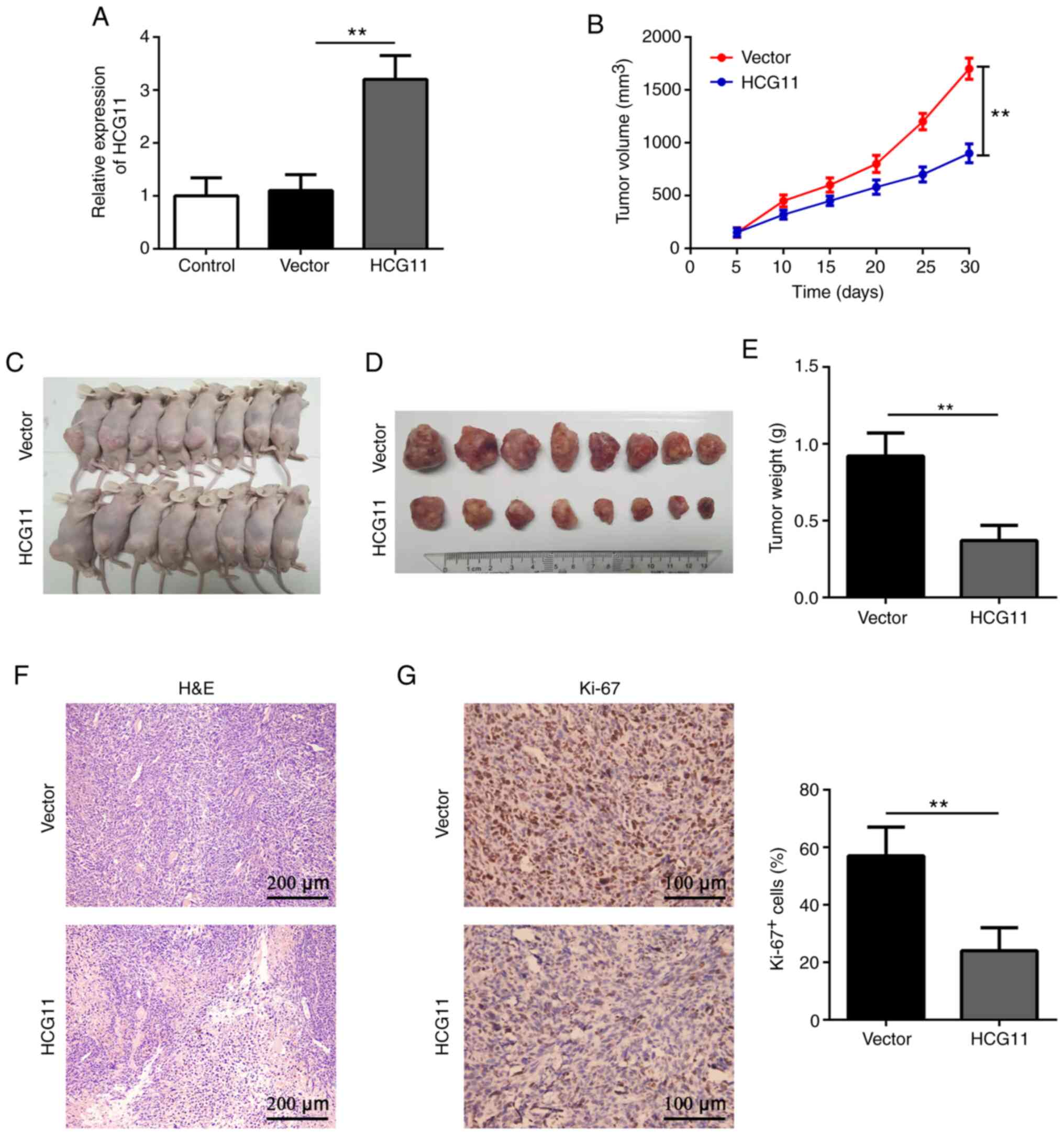
3A). The results revealed that the tumor volume was
significantly decreased following HCG11 overexpression compared
with the mice injected with the empty vector transfected cells
(Fig. 3B and C). The tumor weight
was also significantly lower in the HCG11-overexpression compared
with the control group 30 days after implantation (Fig. 3D and E). Further analysis showed
that HCG11-overexpressing tumors exhibited a loose tissue
structure, accompanied with lower expression of Ki-67, a marker of
proliferation (Fig. 3F, and G).
These data suggest that HCG11 may inhibit NSCLC tumor growth in
vivo.
HCG11 functions as a sponge of miR-875
in NSCLC cells
Given that lncRNAs can act as sponges of miRNAs, it
was hypothesized that HCG11 may inhibit a number of oncogenic
miRNAs in NSCLC cells. Bioinformatics analysis using StarBase
version 3.0 (starbase.sysu.edu.cn/) revealed that several miRNAs,
including miR-26a, miR-499a, miR-522, miR-224, miR-483, miR-214,
miR-455, miR-543, miR-155, miR-105 and miR-875, possessed potential
binding sites with HCG11. Interestingly, it was found that the
expression levels of miR-875, but not other miRNAs, were
significantly decreased in NSCLC cells following HCG11
overexpression (Fig. 4A).
Consistent with this finding, miR-875 expression was increased in
NSCLC tissues compared with adjacent normal tissues (Fig. 4B). Additionally, there was a
significant negative association between HCG11 and miR-875
expression in NSCLC tissues (Fig.
4C), indicating that HCG11 suppressed NSCLC progression,
potentially via inhibition of miR-875. Subsequently, a luciferase
reporter assay demonstrated that HCG11 could directly bind to
miR-875 (Fig. 4D). These results
suggest that HCG11 may inhibit the development of NSCLC by
inhibiting the expression of miR-875.
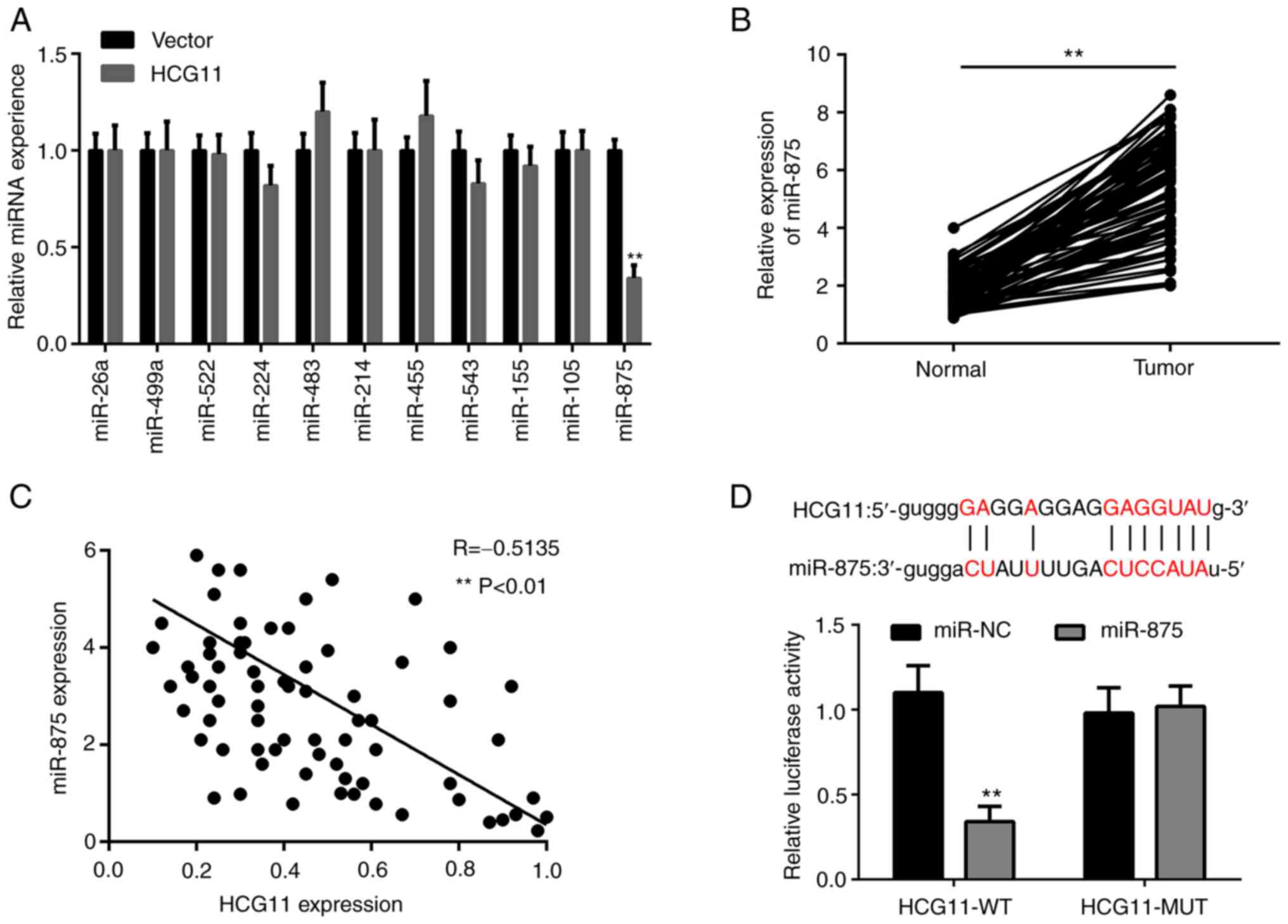 | Figure 4.HCG11 functions as a sponge of
miR-875 in NSCLC cells. (A) Relative expression levels of miR-26a,
miR-499a, miR-522, miR-224, miR-483, miR-214, miR-455, miR-534,
miR-155, miR-105 and miR-875 in A549 cells following overexpression
of HCG11. (B) Reverse transcription-quantitative PCR analysis of
the expression levels of miR-875 in NSCLC tissues and adjacent
normal tissues obtained from 85 patients. (C) Pearson correlation
analysis of HCG11 and miR-875 expression in NSCLC tissues obtained
from 85 patients. (D) Relative luciferase activity in 293T cells
after co-transfection of WT or MUT HCG11 with miR-875-mimic or
miR-NC. The specific binding site of miR-875 with HCG11 is shown in
the top panel, based on StarBase. **P<0.01. NSCLC, non-small
cell lung cancer; miR/miRNA, microRNA; NC, negative control; WT,
wild-type; MUT, mutant; HCG11, HLA complex group 11. |
HCG11 inhibits the malignant phenotype
of NSCLC cells by regulating a miR-875/SATB2 axis
TargetScan (targetscan.org/vert_71/) was used to search for
candidate targets with potential miR-875 binding sites. It was
observed that there was a common binding site between miR-875 and
the 3′-untranslated region of SATB2, a tumor suppressor (23), which was further verified using a
luciferase reporter assay (Fig.
5A). Furthermore, it was noticed that SATB2 protein expression
was significantly upregulated in HCG11-overexpressing A549 cells in
which miR-875 was substantially reduced (Fig. 5B). These findings suggested that
SATB2 may be a direct target of miR-875. To further confirm this
hypothesis, a miR-875-mimic was used and the transfection
efficiency was confirmed by RT-qPCR (Fig. 5C). Specifically, miR-875-mimic
treatment could increase miR-875 levels in HCG11-overexpressing
A549 cells (Fig. 5D).
Overexpression of miR-875 significantly decreased the expression
levels of SATB2, thereby reversing the effects of HCG11 on NSCLC
cells (Fig. 5E-I). Overall, these
findings show that HCG11 may serve a critical role in inhibiting
the malignant phenotype of NSCLC cells by regulating a
miR-875/SATB2 axis.
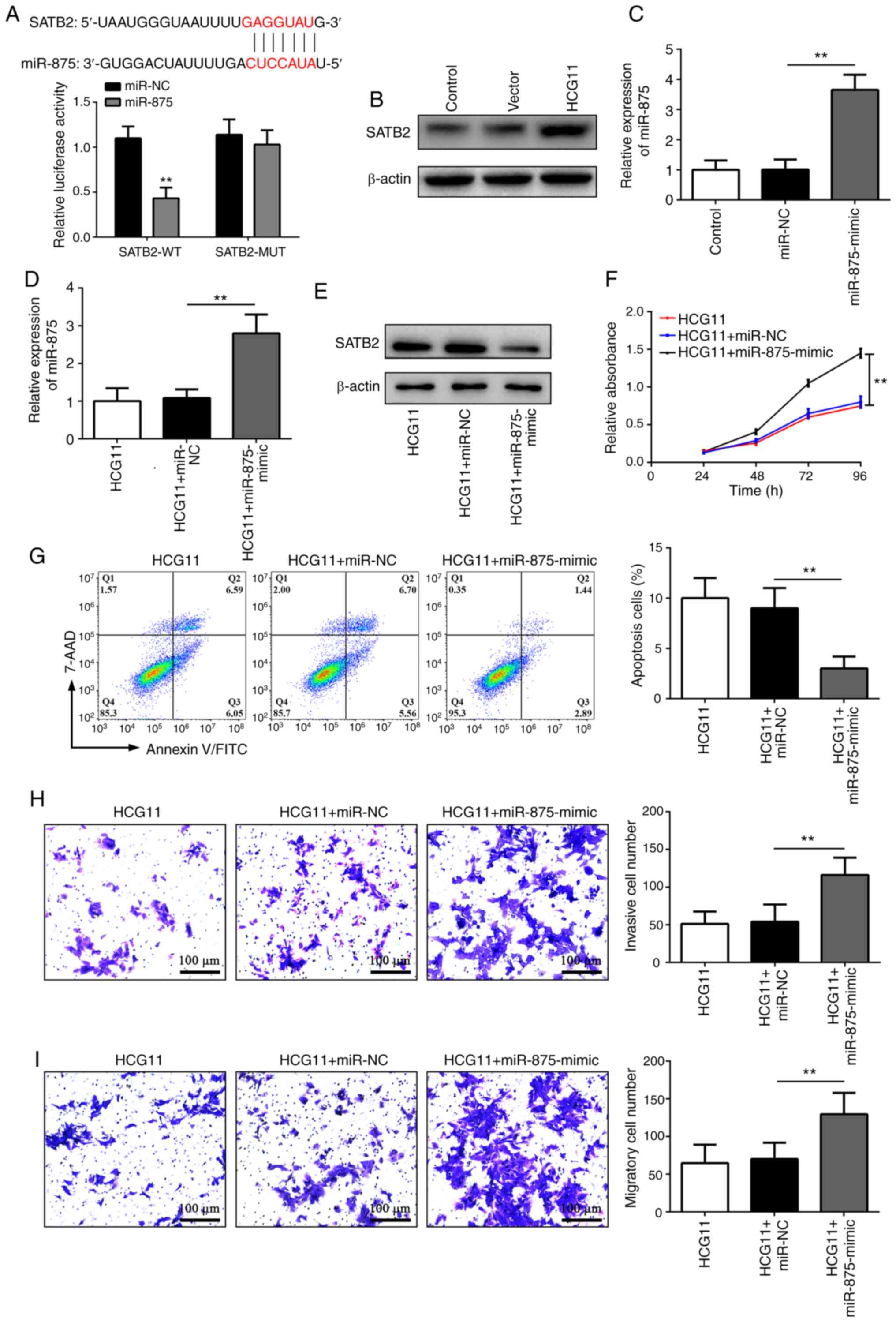 | Figure 5.HCG11 inhibits the malignant
phenotype of NSCLC cells by regulating a miR-875/SATB2 axis. (A)
Relative luciferase activity in 293T cells after co-transfection of
WT or MUT SATB2 with miR-875-mimic or miR-NC. The specific binding
site of miR-875 with SATB2 mRNA is shown in the top panel, based on
TargetScan. (B) Western blot analysis of the expression levels of
SATB2 in the Control, Vector and HCG11 groups. (C) Transfection
efficiency of miR-875-mimic, as determined by RT-qPCR. (D-I) A549
cells with overexpression of HCG11 were treated with miR-NC or
miR-875-mimic. Subsequently, the expression levels of (D) miR-875
were measured by RT-qPCR, and (E) SATB2 protein expression was
determined by western blotting. Finally, the (F) proliferation, (G)
apoptosis, (H) invasion and (I) migration were analyzed by an MTT
assay, flow cytometry or Transwell invasion and migration assays,
respectively. **P<0.01. NSCLC, non-small cell lung cancer;
miR/miRNA, microRNA; NC, negative control; WT, wild-type; MUT,
mutant; RT-qPCR, reverse transcription-quantitative PCR; SATB2,
special AT-rich sequence-binding protein 2; HCG11, HLA complex
group 11 Vector, empty vector control. |
Discussion
Lung cancer is characterized by a high recurrence
rate, high mortality rate and low 5-year survival rate, and
threatens the lives of individuals worldwide (24–26).
Although numerous advances have been made in our understanding of
the disease, the prognosis remains unsatisfactory. Therefore, it is
necessary to identify more effective strategies for early
prevention, detection and treatment of lung cancer. To the best of
our knowledge, the present study was the first to show that lncRNA
HCG11 functions as a tumor suppressor in NSCLC by regulating a
miR-875/SATB2 axis.
The role of lncRNAs in cancer is attracting
increasing attention (27–29). Due to the complexity of function and
heterogeneity of expression, they serve varying roles in the
development and metastasis dependent on the type of cancer. For
example, lncRNA H19 imprinted maternally expressed transcript is
upregulated and promotes carcinogenesis and metastasis in gastric
cancer (30). Additionally,
downregulation of lncRNA GAS5 contributes to cell proliferation and
predicts adverse consequences in cervical cancer (31). Previous studies have shown that the
lncRNA HCG11 affects the progression of glioma, prostate cancer and
hepatocellular carcinoma (32,33).
However, to the best of our knowledge, the expression levels and
role of HCG11 in NSCLC have not yet been explored. The present
study revealed that the expression levels of HCG11 were
significantly reduced postoperatively in tumor tissues compared
with adjacent normal tissues obtained from patients with NSCLC.
Reduction in HCG11 expression was observed in different lung cancer
cell lines. Of note, the survival rate of patients with high HCG11
expression was higher than that of patients with low HCG11
expression. Cox regression analysis showed that HCG11 expression
was an independent predictive factor for overall survival of NSCLC
patients. Therefore, these data suggest that HCG11 may exert an
inhibitory effect on lung cancer progression. In addition, HCG11
could be used as a prognostic marker of NSCLC.
Uncontrolled proliferation, increased invasion and
migration, and decreased apoptosis are the primary causes of the
development and progression of cancer (34–36).
In the present study, overexpression of HCG11 significantly reduced
cell proliferation, invasion and migration, and induced apoptosis
in NSCLC cells. In line with these findings, the expression levels
of PCNA were decreased, and the levels of cleaved-caspase 3 and
cleaved-caspase-9 were increased in NSCLC cells following HCG11
overexpression. More importantly, overexpression of HCG11 also
slowed NSCLC tumor growth in nude mice. These results illustrated
that HCG11 can inhibit the development and progression of
NSCLC.
Previous studies have revealed that HCG11 is
involved in various types of cancer via targeting of miRNAs or
direct regulation of gene expression (19,37,38).
For instance, HCG11 can affect the growth of glioma by regulating
the miR-496/cytoplasmic polyadenylation element binding protein 3
or miR-4425/metastasis associated 1 family member 3 axis.
Additionally, HCG11 is capable of promoting prostate cancer
progression by modulating miR-543 and the AKT/mTOR signaling
pathway (33). Furthermore, HCG11
is able to suppress the apoptosis of hepatocellular carcinoma cells
by regulating insulin like growth factor 2 mRNA binding protein 1
(19). The present study revealed
that HCG11 overexpression resulted in a significant reduction in
miR-875 expression, and HCG11 expression was negatively associated
with miR-875 expression in NSCLC tissues. In particular, a
luciferase reporter assay demonstrated that HCG11 could function as
a competing endogenous RNA to bind to miR-875. A recent study
reported that miR-875 is involved in facilitating proliferation and
inhibiting apoptosis of NSCLC cells (39). Hence, the results of the present
study indicated that upregulation of HCG11 can inhibit NSCLC tumor
growth, potentially by downregulating miR-875.
miRNAs regulate a wide range of cell biological
functions by inhibiting the translation of target genes (40–42).
Previous studies have shown that SATB2 is a cancer suppressor gene
and its expression is decreased in NSCLC cells, and that
downregulation of SATB2 promotes the progression of NSCLC (43–45).
However, how expression of SATB2 is regulated in NSCLC cells is
still not fully understood. In the present study, using
bioinformatics analysis and luciferase reporter assays, it was
demonstrated that SATB2 was a direct target gene of miR-875.
Consistently, it was noticed that SATB2 expression was
significantly increased in NSCLC cells following HCG11
overexpression. miR-875-mimic treatment significantly reduced the
HCG11 overexpression-induced upregulation of SATB2, thereby largely
abrogating the effects of HCG11 in NSCLC cells. Taken together,
these results suggested that HCG11 inhibited proliferation,
invasion and migration, and facilitated apoptosis of NSCLC cells,
at least in part, by regulating a miR-875/SATB2 axis. However,
lncRNAs may be involved in the regulation of cell biology via
numerous other mechanisms. Therefore, it is the possible that there
are other mechanisms which mediate the antitumor role of HCG11 in
NSCLC.
In conclusion, the present study highlighted the
importance of HCG11 in alleviating the malignant phenotype of NSCLC
cells by targeting a miR-875/SATB2 pathway. These data not only
further improve our understanding of the mechanism of regulation of
NSCLC, but also highlights a potential therapeutic target for
management of NSCLC. Nevertheless, it is necessary to further
explore the pathogenesis of NSCLC and treatment strategies.
Acknowledgements
Not applicable.
Funding
The current study was supported by funding from the
Scientific research project of Sichuan health and Family Planning
Commission (grant no. 17PJ598).
Availability of data and materials
The datasets used and analyzed during the present
study are available from the corresponding author on reasonable
request.
Authors' contributions
ZS and WL conceived and designed the study. MC and
RD analyzed and interpreted the results. LS and QZ performed the
experiments. ZS and WL confirmed the authenticity of all the raw
data. All authors have read and approved the final manuscript.
Ethics approval and consent to
participate
All animal experiments in this study were approved
by the Animal Care and Use Committee of Sichuan Mianyang 404
Hospital and performed in accordance with the Guide for the Care
and Use of Laboratory Animals. The use of human samples was
approved by the Ethics Committee of The Sichuan Mianyang 404
Hospital (Mianyang, China; approval no. 008), and each patient
provided a written informed consent.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Jemal A, Bray F, Center MM, Ferlay J, Ward
E and Forman D: Global cancer statistics. CA Cancer J Clin.
61:69–90. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Carr SR, Akerley W and Cannon-Albright LA:
Genetic contribution to nonsquamous, non-small cell lung cancer in
nonsmokers. J Thorac Oncol. 13:938–945. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Greathouse KL, White JR, Vargas AJ,
Bliskovsky VV, Beck JA, von Muhlinen N, Polley EC, Bowman ED, Khan
MA, Robles AI, et al: Interaction between the microbiome and TP53
in human lung cancer. Genome Biol. 19:1232018. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Velcheti V, Madison R, Ali SM and Schrock
AB: WAC/RET: A novel RET oncogenic fusion variant in non-small cell
lung carcinoma. J Thorac Oncol. 13:e122–e123. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Tsui DWY, Murtaza M, Wong ASC, Rueda OM,
Smith CG, Chandrananda D, Soo RA, Lim HL, Goh BC, Caldas C, et al:
Dynamics of multiple resistance mechanisms in plasma DNA during
EGFR-targeted therapies in non-small cell lung cancer. EMBO Mol
Med. 10:e79452018. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Li S, Yang J, Xia Y, Fan Q and Yang KP:
Long noncoding RNA NEAT1 promotes proliferation and invasion via
targeting miR-181a-5p in non-small cell lung cancer. Oncol Res.
26:289–296. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Thomas A, Rajan A, Lopez-Chavez A, Wang Y
and Giaccone G: From targets to targeted therapies and molecular
profiling in non-small cell lung carcinoma. Ann Oncol. 24:577–585.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Vijayvergia N, Shah PC and Denlinger CS:
Survivorship in non-small cell lung cancer: Challenges faced and
steps forward. J Natl Compr Canc Netw. 13:1151–1161. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Kudo Y, Haymaker C, Zhang J, Reuben A,
Duose DY, Fujimoto J, Roy-Chowdhuri S, Solis Soto LM, Dejima H,
Parra ER, et al: Suppressed immune microenvironment and repertoire
in brain metastases from patients with resected non-small-cell lung
cancer. Ann Oncol. 30:1521–1530. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Tang Y, Cao G, Zhao G, Wang C and Qin Q:
LncRNA differentiation antagonizing non-protein coding RNA promotes
proliferation and invasion through regulating miR-135a/NLRP37 axis
in pancreatic cancer. Invest New Drugs. 38:714–721. 2020.
View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Chen LL: Linking Long noncoding RNA
localization and function. Trends Biochem Sci. 41:761–772. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
12
|
St Laurent G, Wahlestedt C and Kapranov P:
The Landscape of long noncoding RNA classification. Trends Genet.
31:239–251. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Kopp F and Mendell JT: Functional
classification and experimental dissection of long noncoding RNAs.
Cell. 172:393–407. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Jin J, Wang H, Si J, Ni R, Liu Y and Wang
J: ZEB1-AS1 is associated with poor prognosis in non-small-cell
lung cancer and influences cell migration and apoptosis by
repressing ID1. Clin Sci (Lond). 133:381–392. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Yang H, Yan L, Sun K, Sun X, Zhang X, Cai
K and Song T: lncRNA BCAR4 increases viability, invasion, and
migration of non-small cell lung cancer cells by targeting
glioma-associated oncogene 2 (GLI2). Oncol Res. 27:359–369.
2019. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Shi SL and Zhang ZH: Long non-coding RNA
SNHG1 contributes to cisplatin resistance in non-small cell lung
cancer by regulating miR-140-5p/Wnt/β-catenin pathway. Neoplasma.
66:756–765. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Kamel LM, Atef DM, Mackawy AMH, Shalaby SM
and Abdelraheim N: Circulating long non-coding RNA GAS5 and SOX2OT
as potential biomarkers for diagnosis and prognosis of non-small
cell lung cancer. Biotechnol Appl Biochem. 66:634–642. 2019.
View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Zhou Y, Shi H, Du Y, Zhao G, Wang X, Li Q,
Liu J, Ye L, Shen Z, Guo Y and Huang Y: lncRNA DLEU2 modulates cell
proliferation and invasion of non-small cell lung cancer by
regulating miR-30c-5p/SOX9 axis. Aging (Albany NY). 11:7386–7401.
2019. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Xu Y, Zheng Y, Liu H and Li T: Modulation
of IGF2BP1 by long non-coding RNA HCG11 suppresses apoptosis of
hepatocellular carcinoma cells via MAPK signaling transduction. Int
J Oncol. 51:791–800. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Liu H, Li J, Koirala P, Ding X, Chen B,
Wang Y, Wang Z, Wang C, Zhang X and Mo YY: Long non-coding RNAs as
prognostic markers in human breast cancer. Oncotarget.
7:20584–20596. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Zhang Y, Zhang P, Wan X, Su X, Kong Z,
Zhai Q, Xiang X, Li L and Li Y: Downregulation of long non-coding
RNA HCG11 predicts a poor prognosis in prostate cancer. Biomed
Pharmacother. 83:936–941. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Costa GJ, Mello MJG, Bergmann A, Ferreira
CG and Thuler LCS: Tumor-node-metastasis staging and treatment
patterns of 73,167 patients with lung cancer in Brazil. J Bras
Pneumol. 46:e201802512020.(In English, Portuguese). View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Roy SK, Shrivastava A, Srivastav S,
Shankar S and Srivastava RK: SATB2 is a novel biomarker and
therapeutic target for cancer. J Cell Mol Med. 24:11064–11069.
2020. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Jemal A, Siegel R, Xu J and Ward E: Cancer
statistics, 2010. CA Cancer J Clin. 60:277–300. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
van Rens MT, Zanen P, Brutel de La Riviere
A, Elbers HR, van Swieten HA and van Den Bosch JM: Survival in
synchronous vs. single lung cancer: upstaging better reflects
prognosis. Chest. 118:952–958. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Marcus MW, Raji OY and Field JK: Lung
cancer screening: Identifying the high risk cohort. J Thorac Dis. 7
(Suppl 2):S156–S162. 2015.PubMed/NCBI
|
|
27
|
Li Y, Li L, Wang Z, Pan T, Sahni N, Jin X,
Wang G, Li J, Zheng X, Zhang Y, et al: LncMAP: Pan-cancer atlas of
long noncoding RNA-mediated transcriptional network perturbations.
Nucleic Acids Res. 46:1113–1123. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Zhang E, He X, Zhang C, Su J, Lu X, Si X,
Chen J, Yin D, Han L and De W: A novel long noncoding RNA HOXC-AS3
mediates tumorigenesis of gastric cancer by binding to YBX1. Genome
Biol. 19:1542018. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Khan MR, Xiang S, Song Z and Wu M: The
p53-inducible long noncoding RNA TRINGS protects cancer cells from
necrosis under glucose starvation. EMBO J. 36:3483–3500. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Yang F, Bi J, Xue X, Zheng L, Zhi K, Hua J
and Fang G: Up-regulated long non-coding RNA H19 contributes to
proliferation of gastric cancer cells. FEBS J. 279:3159–3165. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Cao S, Liu W, Li F, Zhao W and Qin C:
Decreased expression of lncRNA GAS5 predicts a poor prognosis in
cervical cancer. Int J Clin Exp Pathol. 7:6776–6783.
2014.PubMed/NCBI
|
|
32
|
Chen Y, Bao C, Zhang X, Lin X, Huang H and
Wang Z: Long non-coding RNA HCG11 modulates glioma progression
through cooperating with miR-496/CPEB3 axis. Cell Prolif.
52:e126152019. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Wang YC, He WY, Dong CH, Pei L and Ma YL:
lncRNA HCG11 regulates cell progression by targeting miR-543 and
regulating AKT/mTOR pathway in prostate cancer. Cell Biol Int. Nun
22–2019.(Epub ahead of print). doi: 10.1002/cbin.11194. View Article : Google Scholar
|
|
34
|
Cramer JD, Burtness B, Le QT and Ferris
RL: The changing therapeutic landscape of head and neck cancer. Nat
Rev Clin Oncol. 16:669–683. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Evans ER, Bugga P, Asthana V and Drezek R:
Metallic Nanoparticles for Cancer Immunotherapy. Mater Today
(Kidlington). 21:673–685. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Turajlic S, Sottoriva A, Graham T and
Swanton C: Resolving genetic heterogeneity in cancer. Nat Rev
Genet. 20:404–416. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Long R, Liu Z, Li J, Zhang Y and Yu H:
HCG11 up-regulation induced by ELK4 suppressed proliferation in
vestibular schwannoma by targeting miR-620/ELK4. Cancer Cell Int.
21:52021. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Li XF, Hu DM, Zhao YX, Zhang L and Jin Y:
Knockdown of lncRNA HCG11 suppresses cell progression in ovarian
cancer by modulating miR-144-3p/PBX3. Eur Rev Med Pharmacol Sci.
24:11032–11040. 2020.PubMed/NCBI
|
|
39
|
Tong JL, Wang LL, Ling XF, Wang MX, Cao W
and Liu YY: MiR-875 can regulate the proliferation and apoptosis of
non-small cell lung cancer cells via targeting SOCS2. Eur Rev Med
Pharmacol Sci. 23:5235–5241. 2019.PubMed/NCBI
|
|
40
|
Liang C, Liu H, Hao J, Li J and Luo L:
Expression profiling and regulatory network of cucumber microRNAs
and their putative target genes in response to cucumber green
mottle mosaic virus infection. Arch Virol. 164:1121–1134. 2019.
View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Li J, Huang L, Xiao X, Chen Y, Wang X,
Zhou Z, Zhang C and Zhang Y: Photoclickable microRNA for the
intracellular target identification of microRNAs. J Am Chem Soc.
138:15943–15949. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Wang Y, Luo J, Zhang H and Lu J: microRNAs
in the same clusters evolve to coordinately regulate functionally
related genes. Mol Biol Evol. 33:2232–2247. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Kucuksayan H, Ozes ON and Akca H:
Downregulation of SATB2 is critical for induction of
epithelial-to-mesenchymal transition and invasion of NSCLC cells.
Lung Cancer. 98:122–129. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Tian W, Wang G, Liu Y, Huang Z, Zhang C,
Ning K, Yu C, Shen Y, Wang M, Li Y, et al: The miR-599 promotes
non-small cell lung cancer cell invasion via SATB2. Biochem Biophys
Res Commun. 485:35–40. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Ma YN, Zhang HY, Fei LR, Zhang MY, Wang
CC, Luo Y and Han YC: SATB2 suppresses non-small cell lung cancer
invasiveness by G9a. Clin Exp Med. 18:37–44. 2018. View Article : Google Scholar : PubMed/NCBI
|