Introduction
Ovarian cancer is the most common cause of death
among patients with female genital tumors, and consists of a
variety of pathological types, of which the most common subtype is
epithelial ovarian cancer (EOC) (1,2). Due
to the ovary being located deep in the pelvic cavity, the early
diagnosis of EOC remains difficult, and currently, there is a lack
of effective screening methods; therefore, the majority of patients
are diagnosed with EOC upon reaching the advanced stages (3–5). In
addition, ovarian cancer is prone to extensive pelvic, abdominal
and lymph node metastasis, resulting in EOC displaying the highest
mortality rate among tumors of the reproductive system (6–8). Tumor
initiation and development are affected by numerous factors,
including cell movement, the aberrant expression of tumor
suppressor genes or oncogenes, and the abnormal regulation of
migration (9,10). The current treatment strategy for
ovarian cancer is surgery supplemented with platinum-based
chemotherapy; however, despite the majority of patients initially
responding to treatment, a significant proportion of patients
eventually die from recurrence and resistance (11,12).
Therefore, investigation of the molecular mechanisms underlying EOC
to identify novel treatment targets and improve the survival rate
of patients with EOC is required.
MicroRNAs (miRNAs/miRs) are non-coding, endogenous,
small single-stranded RNAs that are 20–24 nucleotides in length,
which regulate numerous basic biological processes in vivo
(13,14). Although miRNAs are non-coding RNAs,
by binding with the 3′-untranslated region (3′-UTR) of target
mRNAs, miRNAs promote mRNA cleavage or block mRNA translation,
thereby controlling the expression of certain target proteins in
cells (14,15). In addition, miRNAs also serve
important roles during the occurrence, development and metastasis
of tumors (16,17). Numerous studies have reported the
tumor-suppressive role of miR-193a-5p. For example, miR-193a-5p
inhibits HT-29 colon cancer cell metastasis (18). Moreover, miR-193a-5p targeted SPARC
(osteonectin), cwcv and kazal like domains proteoglycan 1 to
inhibit liver cancer cell migration and proliferation, while
promoting apoptosis (18).
miR-193a-5p overexpression in vitro and in vivo
inhibited the formation of pulmonary metastases in non-small cell
lung cancer, as well as inhibiting cell migration, invasion and
epithelial-mesenchymal transition (19). Additionally,
phosphoinositide-3-kinase regulatory subunit 3 (PIK3R3) and mTOR
were identified as direct target genes of miR-193a-5p, thereby
indicating that miR-193a-5p inhibited the AKT/mTOR signaling
pathway (19,20). However, to the best of our
knowledge, the role of miR-193a-5p in EOC has not been previously
reported. Therefore, the aim of the present study was to
investigate the expression levels of miR-193a-5p in serum samples
from patients with EOC and to determine the role of miR-193a-5p in
EOC.
Materials and methods
Clinical sample collection
A total of 60 blood samples were collected from 30
female patients with EOC (age, 41.33±3.01 years) and 30 female
healthy individuals (age, 43.21±2.47 years) who attended Beijing
Ditan Hospital Capital Medical University (Beijing, China) between
January 2018 and July 2019. The present study was approved by the
Ethics Committee of Beijing Ditan Hospital Capital Medical
University. Written informed consent was obtained from all
participants. The blood samples were left standing at 4°C for 1 h,
then centrifuged at 3,000 × g at 4°C for 5 min to obtain the serum
samples.
Cell culture and transfection
EOC cell lines (SKOV3, A2780, HEY, OVCAR3 and Es2)
and the normal cell line IOSE386 were purchased from the American
Type Culture Collection. Cells were cultured in RPMI-1640
(Invitrogen; Thermo Fisher Scientific, Inc.) supplemented with 10%
FBS (HyClone; Cytiva) and 100 U/ml penicillin and 100 µg/ml
streptomycin (Hyclone; Cytiva), and maintained in a humidified
atmosphere of 5% CO2 at 37°C.
After 6 h starvation, SKOV3 cells were cultured in
6-well plates to 70–80% confluence and transfected with
miR-negative control (NC) mimic (100 nM;
5′-UUCUCCGAACGUGUCACGUTT-3′; Guangzhou RiboBio Co., Ltd.) or
miR-193a-5p mimic (50 nM; 5′- UGGGUCUUUGCGGGCGAGAUGA−3′; Guangzhou
RiboBio Co., Ltd.). The pcDNA3.1 RBBP6 overexpression vector
(oe-RBBP6; 100 nM; Guangzhou RiboBio Co., Ltd.) or an empty
pcDNA3.1 vector (oe-NC; 100 nM; Guangzhou RiboBio Co., Ltd.) using
Lipofectamine® 2000 (Invitrogen; Thermo Fisher
Scientific, Inc.). Following transfection for 48 h in a humidified
atmosphere of 5% CO2 at 37°C, cells were harvested for
use in subsequent experiments.
Dual-luciferase reporter assay
To determine the targets of miR-193a-5p, TargetScan
(http://www.targetscan.org/mamm_31)
was used to predict the target gene of RBBP6. Subsequently, a
dual-luciferase reporter assay was performed to verify the
findings. Briefly, SKOV3 cells (4×104) were seeded into
24-well plates and cultured for 24 h in a humidified atmosphere of
5% CO2 at 37°C. A total of 4×104 cells/well
were plated into 60-mm cell culture dishes and cultured for 24 h
until reaching 60–80% confluence. The putative miR-193a-5p binding
site in the 3′-UTR of RBBP6 [wild-type (WT) or mutant (MUT)] was
cloned into psi-CHECK (Promega Corporation) downstream of the
firefly luciferase 3′-UTR. The psi-CHECK vector also provided
Renilla luciferase as the normalization signal.
Subsequently, 100 ng psi-CHECK-RBBP6-WT or psi-CHECK-RBBP6-MUT
luciferase plasmids were co-transfected into SKOV3 cells alongside
100 nM miR-193a-5p mimic or mimic-NC using
Lipofectamine® 2000 (Invitrogen; Thermo Fisher
Scientific, Inc.). Following incubation for 24 h at 37°C,
luciferase activities were detected using a Dual-Luciferase
Reporter Assay system (Promega Corporation). Firefly luciferase
activities were normalized to Renilla luciferase
activities.
Cell Counting Kit-8 (CCK-8) assay
SKOV3 cells were plated (3×103
cells/ml/well) into 96-well plates and separated into two groups:
i) Mimic-NC group and ii) miR-193a-5p mimic group. To assess cell
proliferation, 10 µl CCK-8 solution was added to each well and
incubated for 2 h at 37°C. The absorbance of each well was measured
at a wavelength of 450 nm using a microplate reader.
Colony formation assay
SKOV3 cells were plated (3×103
cells/well) into 6-well plates and incubated at 37°C with 5%
CO2 for 8–16 days. Following incubation, cells were
fixed with 75% methanol for 30 min and stained with 0.5% crystal
violet for 15 min, both at 37°C. Stained cells were visualized
using an IX51 optical microscope (magnification, ×10; Olympus
Corporation) to calculate the number of cell colonies (>50
cells) formed.
Wound healing assay and Transwell
assay for determining of cell migration and invasion,
respectively
Cell migration and invasion were measured by
performing a wound healing and Transwell assay, respectively, as
previously described (21).
For the wound healing assay, 5×105 SKOV3
cells/well were plated into six-well plates and cultured until 100%
confluence in DMEM supplemented with 10% FBS at 37°C. Subsequently,
the monolayer of cells was scratched with a 200-µl sterile pipette
tip and the cells were incubated in serum-free DMEM for 24 h at
37°C. The migratory distance of the cells was observed under a
light microscope (magnification, ×200; Olympus Corporation) and
analyzed using ImageJ version 1.49 software (National Institutes of
Health).
For the cell invasion assay, the upper chambers of
Transwell plates (BD Biosciences) were precoated with Matrigel (BD
Biosciences) at 37°C for 2 h, then cells (5×106
cells/ml) were seeded into the upper chambers in serum-free DMEM
(Invitrogen; Thermo Fisher Scientific, Inc.) supplemented with 2.5%
FBS was plated into the lower chambers. Following culture for 24 h
at 37°C, cells on the upper surface of the Transwell membrane were
removed with a cotton swab. Cells on the lower surface of the
Transwell membrane were washed with PBS and fixed with 4%
paraformaldehyde at room temperature for 30 min. Subsequently, the
paraformaldehyde was discarded, and cells were stained with 0.1%
crystal violet at room temperature for 15 min, prior to being
observed in five fields of view using a microscope (Olympus
Corporation; magnification, ×200). The total number of cells in
each field of view was recorded and the mean number of cells was
calculated.
TUNEL assay
SKOV3 cells (3×103 cells/well) were fixed
in 4% formaldehyde at room temperature for 10 min and washed with
PBS buffer solution twice. SKOV3 cell apoptosis was investigated
using a TUNEL assay kit (cat. no. ab206386; Abcam), according to
the manufacturer's protocol. TUNEL-positive cells presented with
yellow nuclei following visualization using a fluorescence
microscope (Nikon Eclipse 80i; Nikon Corporation; magnification,
×200) in five randomly selected fields of view.
Western blotting
Total protein was extracted from SKOV3 cells using
RIPA lysis buffer (Beyotime Institute of Biotechnology) at 4°C for
1 h. Total protein was quantified using a Pierce™ BCA Protein assay
kit (Thermo Fisher Scientific, Inc.) and protein (30 µg/lane) was
separated via 10% SDS-PAGE and transferred onto PVDF membranes. The
membranes were blocked with 5% skim milk in TBS-0.05% Tween-20
(TBST) for 1 h at room temperature. Subsequently, the membranes
were incubated overnight with the following primary antibodies in
blocking buffer at 4°C: Bcl2 (1:1,000; cat. no. ab182858; Abcam),
Bax (1:1,000; cat. no. ab32503; Abcam), cleaved-caspase 3 (1:1,000;
cat. no. ab49822; Abcam), cleaved-caspase 7 (1:1,000; cat. no.
ab32042; Abcam), cleaved-caspase 9 (1:1,000; cat. no. ab2324;
Abcam), RBBP6 (1:1,000; cat. no. ab237514; Abcam), Ki67 (1:1,000;
cat. no. ab15580; Abcam), proliferating cell nuclear antigen (PCNA;
1:1,000; cat. no. 13110; Cell Signaling Technology, Inc.), matrix
metallopeptidase (MMP)-2 (1:1,000; cat. no. 40994; Cell Signaling
Technology, Inc.), MMP-9 (1:1,000; cat. no. 13667; Cell Signaling
Technology, Inc.) and GAPDH (1:2,000; cat. no. ab181602; Abcam).
Following primary incubation, the membranes were washed with TBST
and incubated with a horseradish peroxidase-conjugated secondary
antibody (1:10,000; cat. no. ab205718; Abcam) for 2 h at room
temperature. Proteins bands were visualized using an ECL reagent
(Invitrogen; Thermo Fisher Scientific, Inc.). Densitometric
analysis was performed using ImageJ software (version 1.49v;
National Institutes of Health).
Reverse transcription-quantitative PCR
(RT-qPCR)
Total RNA was extracted from serum samples and SKOV3
cells with TRIzol® reagent (Invitrogen; Thermo Fisher
Scientific, Inc.). Total RNA was reverse transcribed using
RevertAid reverse transcriptase (Invitrogen; Thermo Fisher
Scientific, Inc.) at 42°C for 1 h, according to the manufacturer's
protocol. qPCR was performed using the LightCycler 480 SYBR Green I
Master kit (Roche Diagnostics) on a LightCycler 480 II (Roche
Diagnostics). The following thermocycling conditions were used for
qPCR: Initial denaturation at 95°C for 5 min; followed by 45 cycles
of amplification, including denaturation at 94°C for 10 sec,
annealing at 60°C for 20 sec and a final extension at 72°C for 30
sec. The following primers were used for qPCR: miR-193a-5p forward,
5′-ACACTCCAGCTGGGTGGGTCTTTGCGGGCG-3′ and reverse,
5′-TGGTGTCGTGGAGTCG-3′; RBBP6 forward, 5′-CTCCCCATACACTTCCTCTCC-3′
and reverse, 5′-TTCTTTTAGTCGTCGCTGCTC-3′; GAPDH forward,
5′-GAGCCATGAGGGAGGCTG-3′ and reverse, 5′-CAGTTGAATCAGATGGATCC-3′;
and U6 forward, 5′-ATCGTCCGTGATCAGCGT-3′ and reverse,
5′-GCAGGTGGTCGGTCGA-3′. miRNA and mRNA expression levels were
quantified using the 2−∆∆Cq method and normalized to the
internal reference genes U6 and GAPDH, respectively (22).
Statistical analysis
Statistical analyses were performed using SPSS
software (version 18.0; SPSS, Inc.). Data are presented as the mean
± SD. Statistical differences among groups were determined using a
one-way ANOVA followed by a Tukey's or Dunnett's post hoc test.
P<0.05 was considered to indicate a statistically significant
difference. All experiments were repeated at least three times.
Results
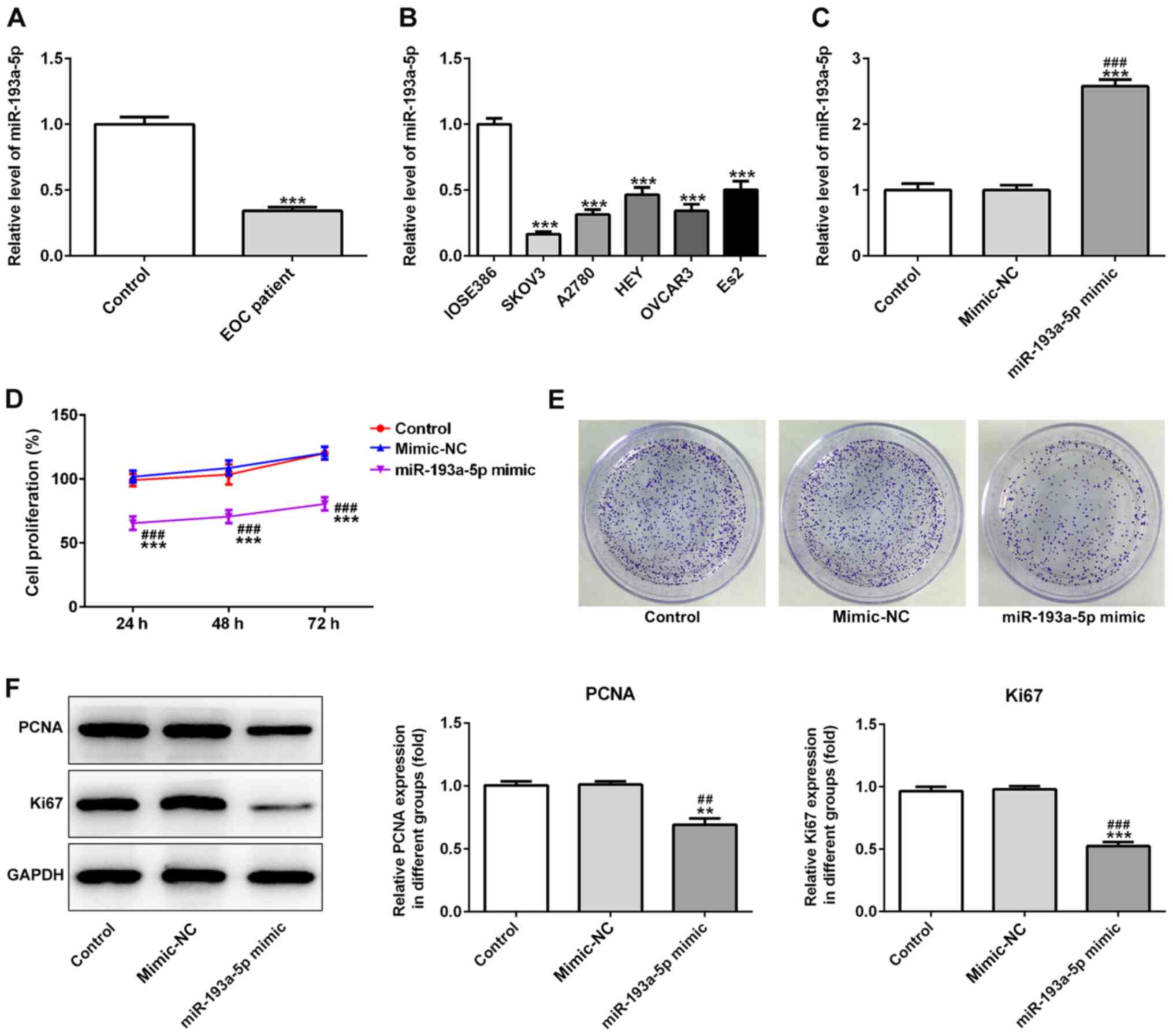
miR-193a-5p is abnormally expressed in
EOC
The expression levels of miR-193a-5p in 30 serum
samples obtained from patients with EOC were significantly
decreased compared with serum samples obtained from healthy
individuals (Fig. 1A). To
investigate the effects of miR-193a-5p, the expression levels of
miR-193a-5p were analyzed in EOC cells. The results indicated that
miR-193a-5p expression levels were decreased in EOC cells compared
with normal cells (Fig. 1B).
miR-193a-5p regulates EOC cell
progression
miR-193a-5p mimic significantly increased the
expression levels of miR-193a-5p in SKOV3 cells compared with the
control and mimic-NC groups (Fig.
1C). In addition, the CCK-8 assay indicated that miR-193a-5p
overexpression significantly decreased SKOV3 cell proliferation
compared with the control and mimic-NC groups (Fig. 1D). Moreover, the colony formation
assay indicated that miR-193a-5p overexpression notably decreased
SKOV3 colony formation compared with the control and mimic-NC
groups (Fig. 1E). Furthermore,
miR-193a-5p overexpression significantly decreased the expression
levels of proliferation-related proteins, PCNA and Ki67, compared
with the control and mimic-NC groups, which also suggested that
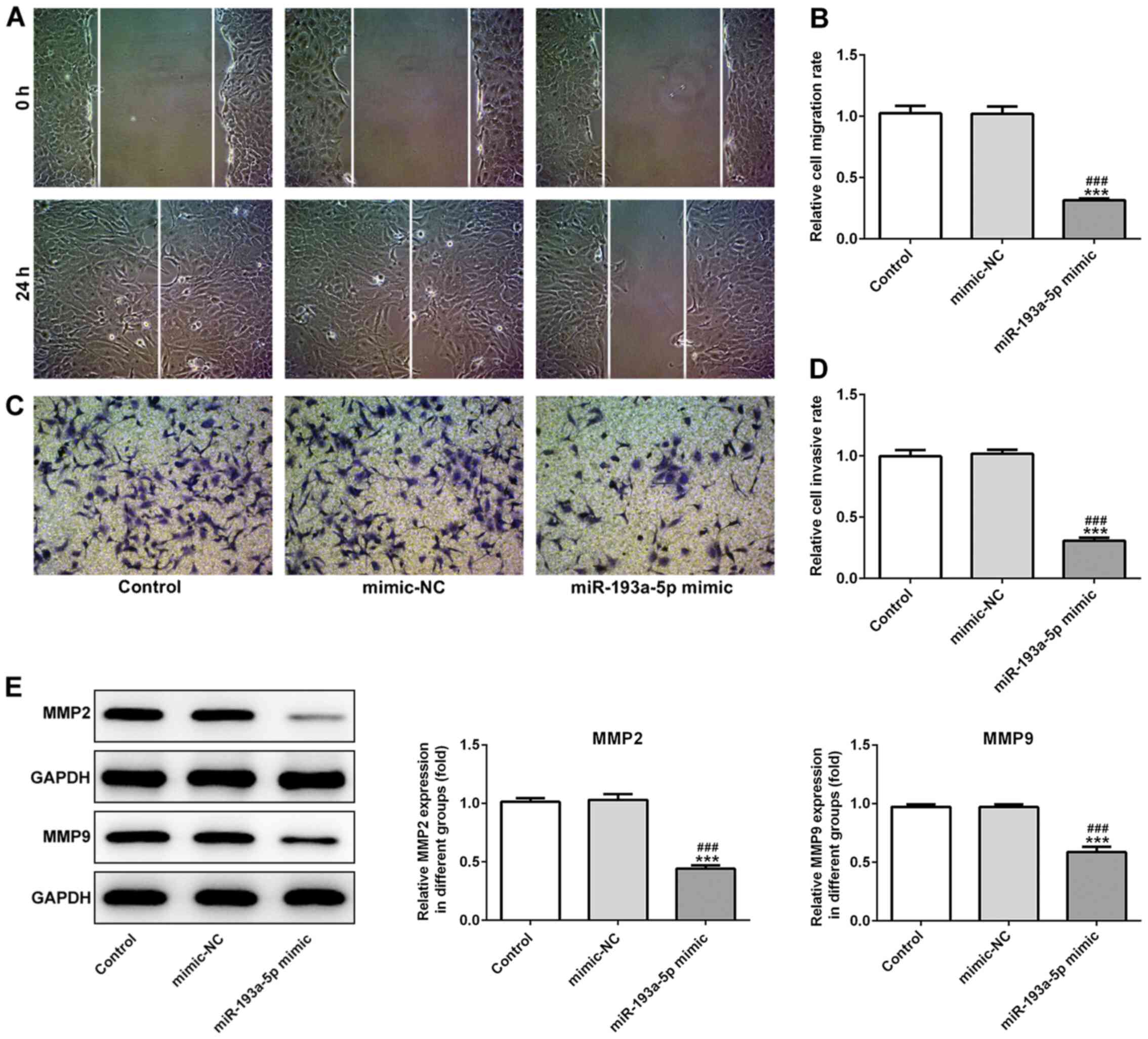
miR-193a-5p regulated EOC cell proliferation (Fig. 1F). Similarly, miR-193a-5p
overexpression significantly inhibited SKOV3 cell migration
compared with the control and mimic-NC groups, as indicated by the
Transwell and wound healing assays (Fig. 2A-D). Moreover, miR-193a-5p
overexpression significantly decreased the expression levels of
MMP2 and MMP9 compared with the control and mimic-NC groups
(Fig. 2E), suggesting that
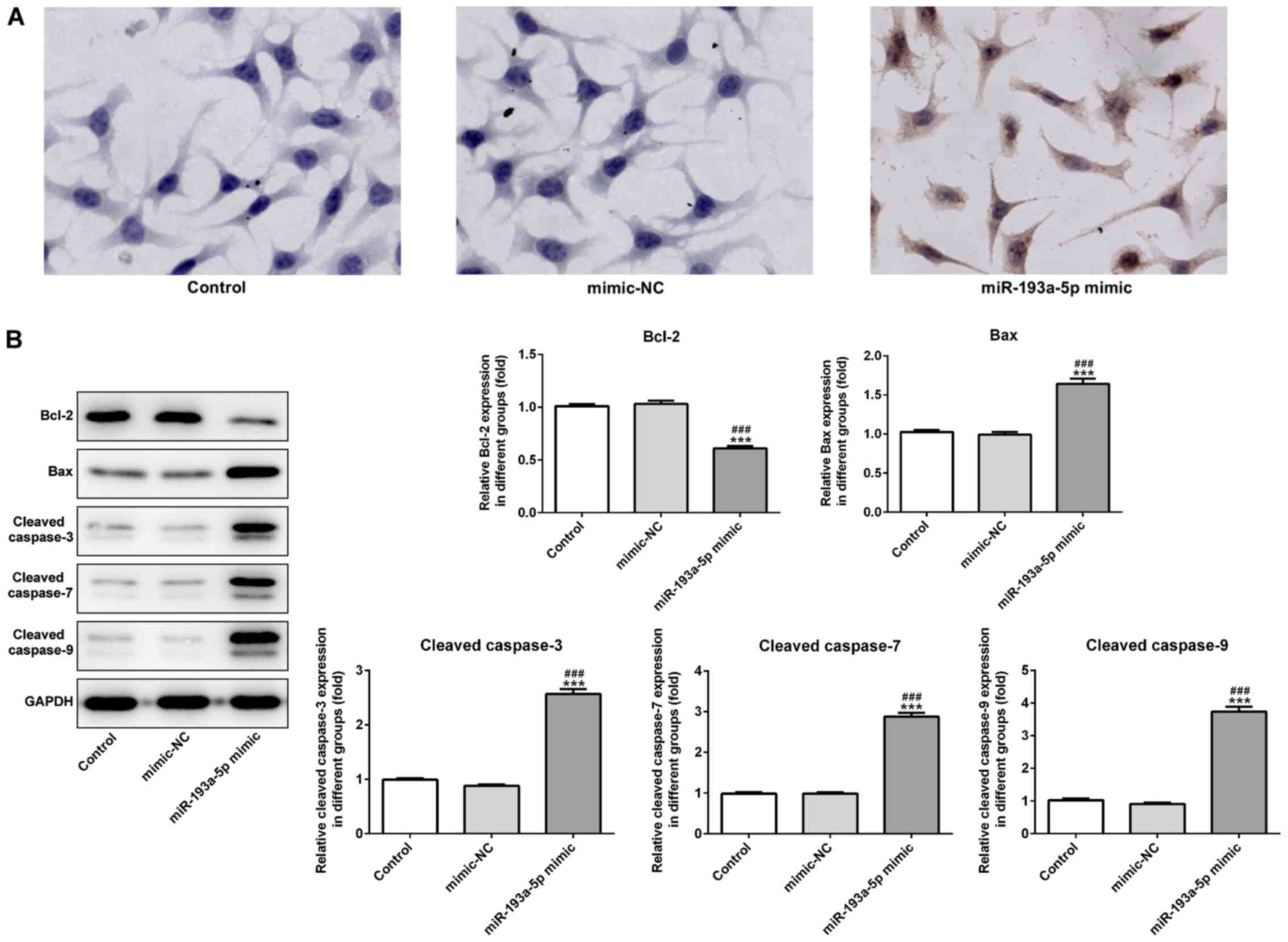
miR-193a-5p may potentially modulate cell migration. Furthermore,
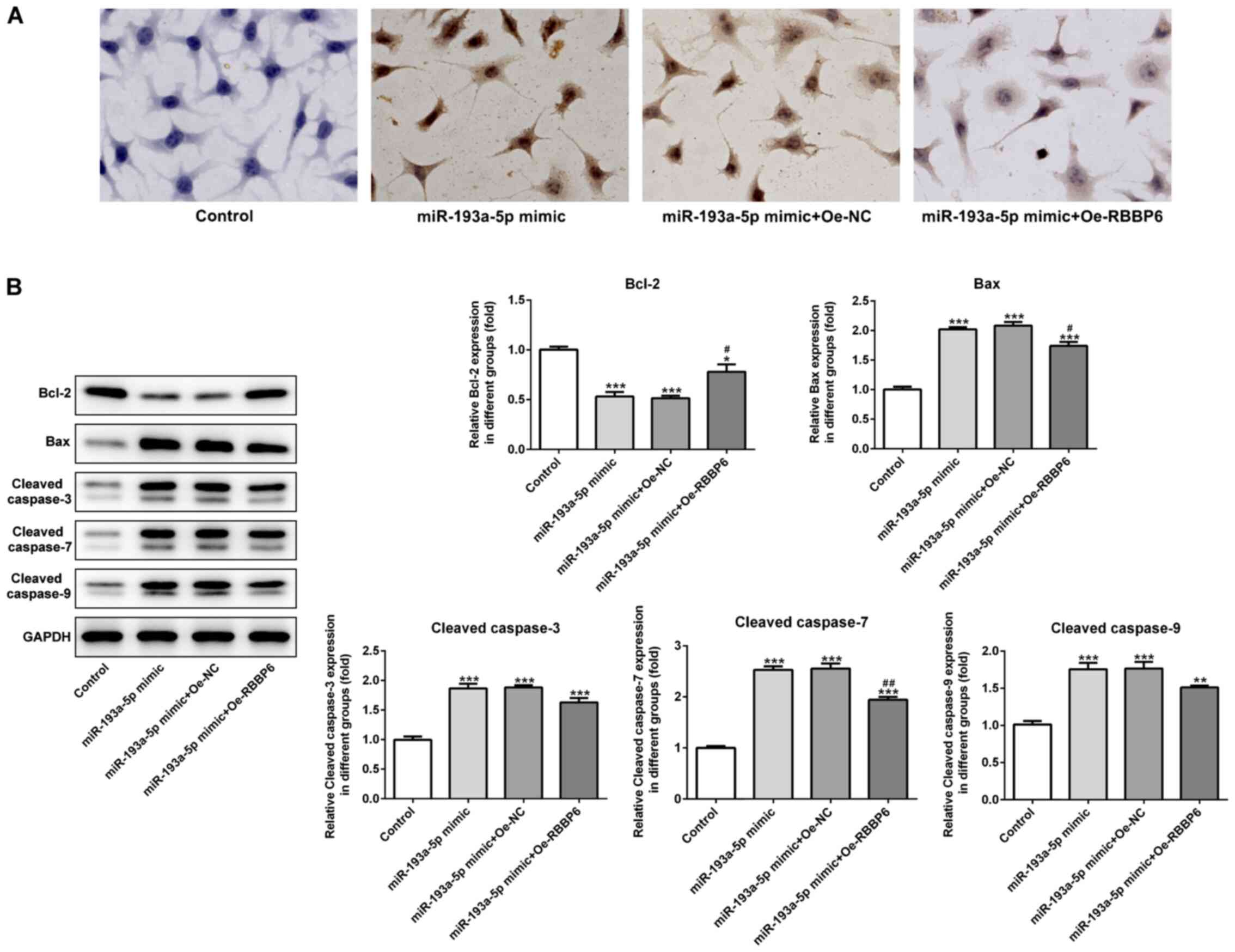
the effects of miR-193a-5p on SKOV3 cell apoptosis were analyzed by
performing a TUNEL assay and western blotting. The TUNEL assay
indicated that miR-193a-5p overexpression notably increased SKOV3
cell apoptosis compared with the control and mimic-NC groups
(Fig. 3A). Furthermore, the
expression levels of proapoptotic proteins, Bax, cleaved caspase-3,
−7 and −9, were significantly increased in the miR-193a-5p mimic
group compared with the control and mimic-NC group. By contrast,
miR-193a-5p overexpression significantly decreased the expression
levels of the antiapoptotic protein Bcl2 compared with the control
and mimic-NC groups (Fig. 3B).
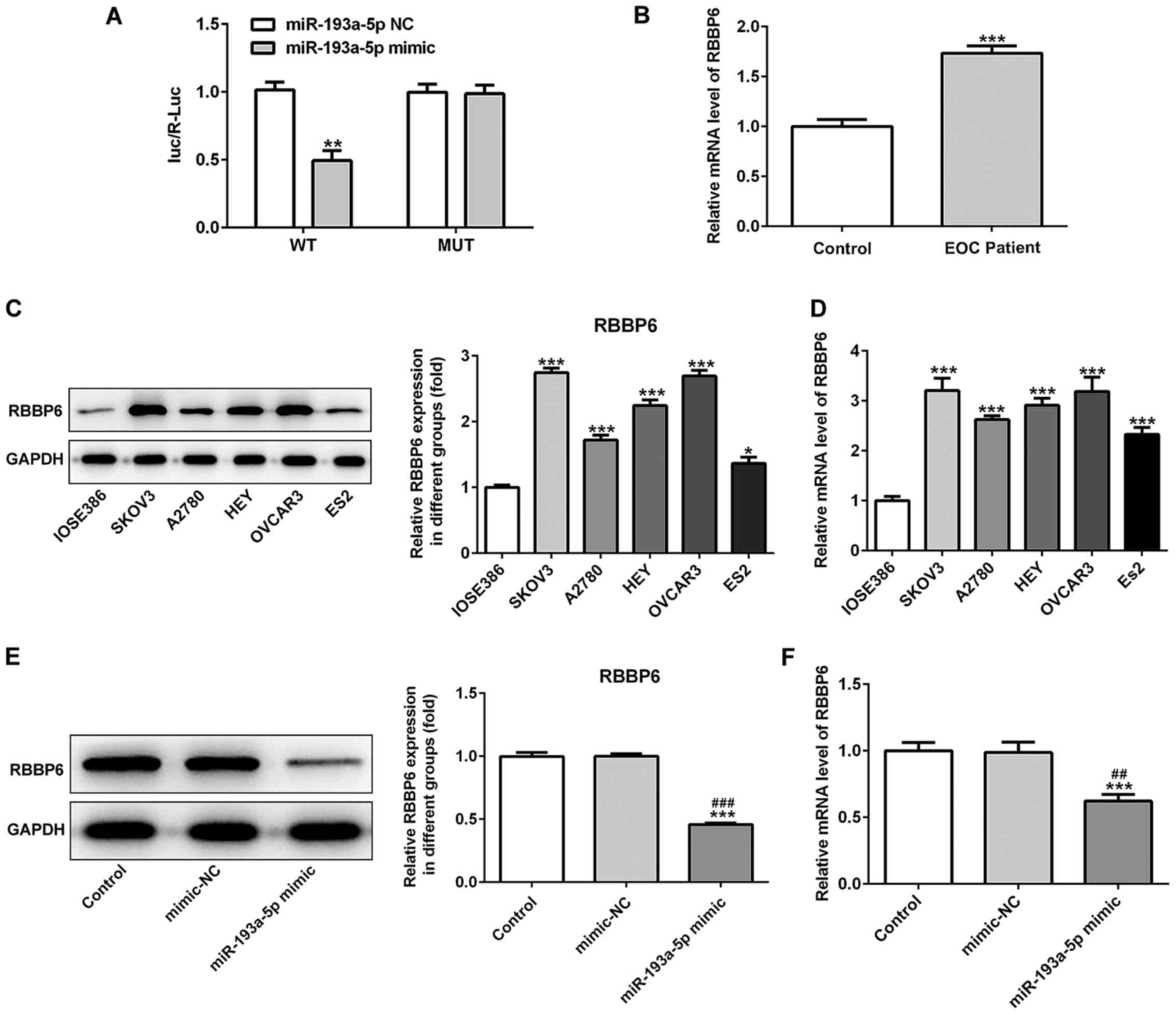
RBBP6 is a target gene of
miR-193a-5p
RBBP6 was predicted as the target gene of
miR-193a-5p using TargetScan. Subsequently, a dual-luciferase
reporter assay was performed to verify RBBP6 as a target gene of
miR-193a-5p. The reporter vector containing RBBP6-WT exhibited
markedly decreased luciferase activity in SKOV3 cells
co-transfected with the miR-193a-5p mimic compared with cells
co-transfected with the miR-193a-5p NC (Fig. 4A). However, the statistical
difference between cells co-transfected with the miR-193a-5p mimic
or miR-193a-5p NC and MUT reporters was not significant. Taken
together, these results showed that RBBP6 was the direct target
gene of miR-193a-5p. The mRNA expression levels of RBBP6 in serum
samples obtained from patients with EOC were significantly
increased compared with blood samples obtained from healthy
individuals (Fig. 4B). Furthermore,
the protein and mRNA expression levels of RBBP6 in EOC cell lines
were significantly increased compared with IOSE386 cells (Fig. 4C and D). In addition, RBBP6
expression levels were significantly decreased in the miR-193a-5p
mimic group compared with the control and mimic-NC groups (Fig. 4E and F).
miR-193a-5p exerts antitumor effects
following RBBP6 overexpression
Subsequently, the RBBP6 overexpression vector was
transfected into SKOV3 cells to determine whether RBBP6 served a
role in the effects of miR-193a-5p on cell proliferation, migration
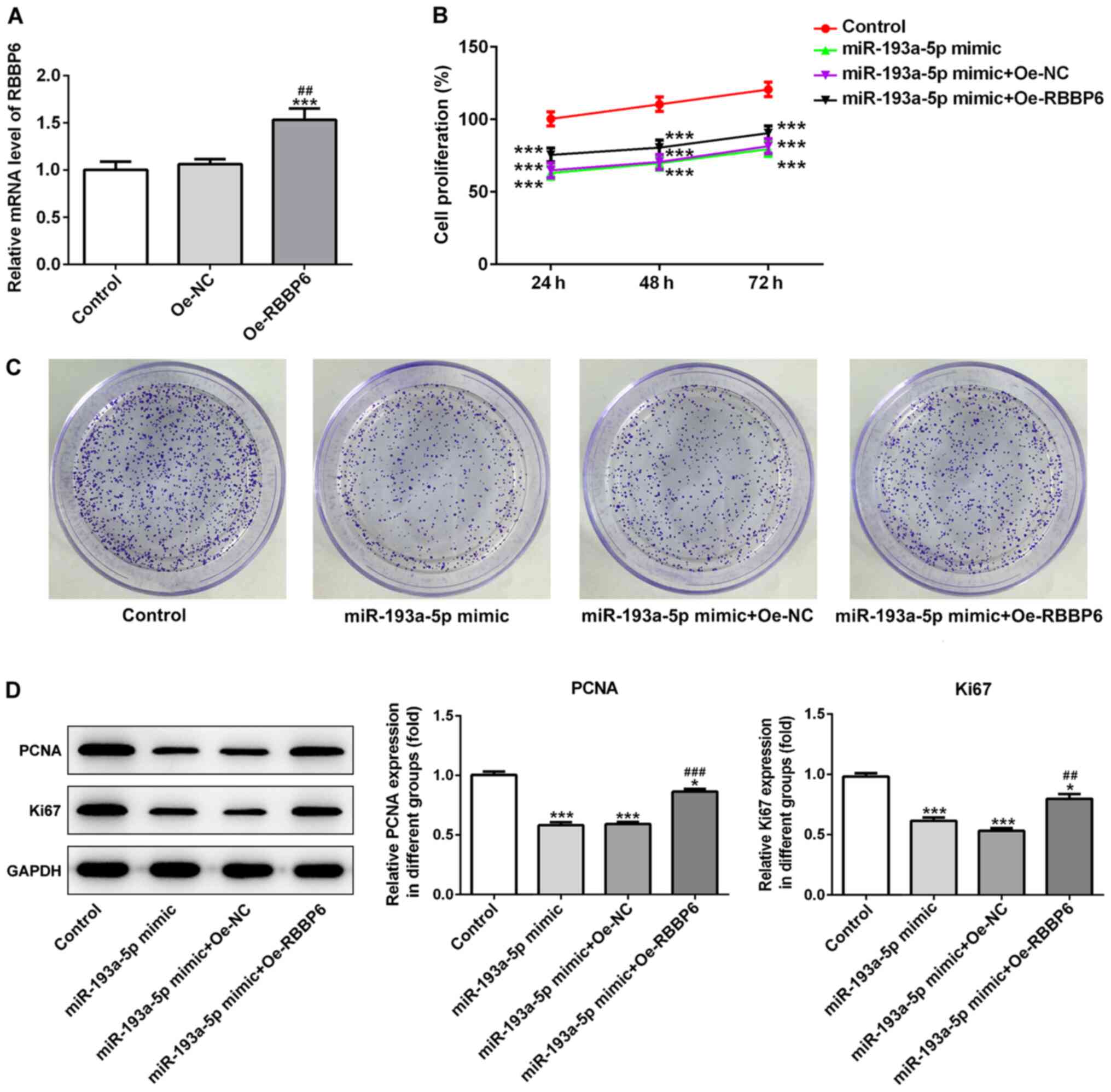
and apoptosis. The transfection efficacy of RBBP6 overexpression in
SKOV3 cells was determined via RT-qPCR (Fig. 5A). RBBP6 overexpression partially
reversed miR-193a-5p mimic-mediated effects on cell proliferation
(Fig. 5B-D), migration (Fig. 6A-C) and apoptosis (Fig. 7A and B). Collectively, the results
suggested that miR-193a-5p may inhibit cell proliferation,
migration and apoptosis by regulating RBBP6.
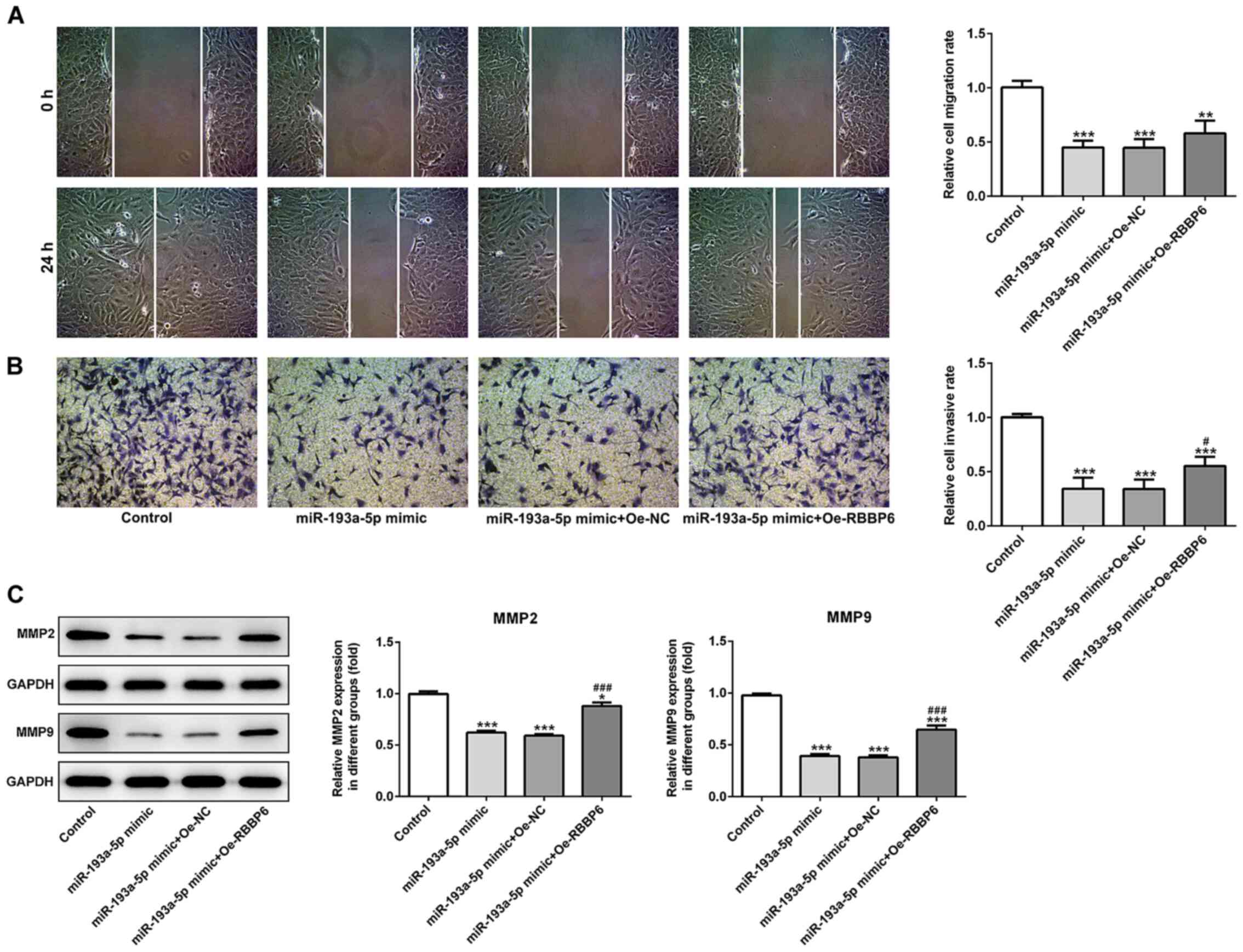 | Figure 6.miR-193a-5p-mediated regulation of
SKOV3 cell migration and invasion is mediated by RBBP6
overexpression. (A) Wound healing and (B) Transwell assays were
conducted to assess cell migration and invasion, respectively.
Magnification, ×200. (C) Western blotting was performed to assess
the expression levels of MMP2 and MMP9. *P<0.05, **P<0.01 and
***P<0.001 vs. control; #P<0.05 and
###P<0.001 vs. miR-193a-5p mimic + Oe-NC. miR,
microRNA; RBBP6, RB binding protein 6, ubiquitin ligase; MMP,
matrix metallopeptidase; Oe, overexpression; NC, negative control.
Magnification, ×200. |
Discussion
The poor prognosis of ovarian cancer is associated
with the fact that the majority of patients with EOC are diagnosed
at an advanced stage; therefore, the early diagnosis of ovarian
cancer may aid with controlling disease progression and reducing
mortality rates (23). miRNAs
affect tumor cell proliferation and apoptosis by regulating
cytokines (24,25). For example, miR-193a-5p expression
levels are downregulated in numerous types of cancer, including
colon cancer, non-small cell lung cancer, human endometrioid
endometrial adenocarcinoma and prostate cancer, which has been
reported to be associated with tumor progression (18–20,26,27). A
previous study also reported an improvement to membranous
nephropathy following miR-193a inhibition, which affected
podocytosis by targeting WT1 transcription factor (WT1) (28). Jin et al (29) demonstrated that miR-193a-5p exerted
a tumor-suppressive role in glioblastoma via modulating NOVA
alternative splicing regulator 1. In addition, Shirafkan et
al (18) identified that
miR-193a-5p inhibited human HT-29 colon cancer cell migration by
suppressing the metastatic pathway. miR-193a-5p also suppressed
human non-small-cell lung cancer metastasis by downregulating the
erb-b2 receptor tyrosine kinase 4/PIK3R3/mTOR/ribosomal protein S6
kinase B2 signaling pathway (20).
In human endometrioid endometrial carcinoma, a novel
miR-193a-5p-YY1 transcription factor- APC regulator of WNT
signaling pathway regulatory axis was identified (26). By contrast, miR-193a-5p knockdown
increased the chemosensitivity of prostate cancer cells to
docetaxel (30). Furthermore,
miR-193a downregulation contributes to non-small cell lung cancer
metastasis by targeting the WT1/E-cadherin axis (31).
miRNAs serve a role in cells and other parts of the
body via circulating in peripheral blood and other body fluids
(32). Previous studies have
demonstrated that serum and plasma miRNAs may serve as non-invasive
biomarkers due to their availability and long-term stability
(33–35); therefore, miRNAs may be used for the
early diagnosis of various types of cancer (36). miR-193a-3p is associated with
cancer. For example, miR-193a-3p overexpression in esophageal
squamous cell carcinoma regulates cancer cell proliferation,
migration and apoptosis. Lin et al (37) demonstrated that the expression level
of miR-193a-5p was negatively correlated with metastasis and poor
prognosis in patients with esophageal squamous cell carcinoma. The
serum expression level of miR-193a-5p can reflect the expression
level of miR-193a-5p in tissues and cells; therefore, serum
miR-193a-5p expression levels can be used as a noninvasive
diagnostic indicator for the prognosis of esophageal cancer
(37). The present study indicated
that miR-193a-5p expression levels were significantly decreased in
serum samples obtained from patients with EOC and EOC cell lines
compared with serum samples obtained from healthy individuals and
normal cells, respectively. The results also suggested a potential
diagnostic value and molecular mechanism underlying miR-193a-5p in
EOC. Further investigations predicted that RBBP6 was a target gene
of miR-193a-5p. RBBP6 has been reported to be associated with
various types of cancer. For example, Yoshitake et al
(38) reported that RBBP6
expression levels were significantly upregulated in invasive
esophageal cancer, suggesting that RBBP6 may promote tumor cell
proliferation and may serve as a promising target for
immunotherapy. In addition, RBBP6 knockdown in breast cancer cells
significantly inhibited cell proliferation (39). Similarly, the increased expression
levels of RBBP6 reported in lung cancer tissues suggested that
RBBP6 may be involved in promoting malignancy in lung tissues, and
inhibition of RBBP6 target genes may render cancer cells sensitive
to apoptosis (40). RBBP6 has also
been identified as a potential marker of apoptosis and cell cycle
arrest in cervical cancer (41).
The present study demonstrated that the expression levels of RBBP6
were significantly increased in patients with EOC compared with
healthy individuals. Moreover, in vitro analysis indicated
that miR-193a-5p mimic significantly decreased SKOV3 cell
viability, migration and invasion, but promoted SKOV3 cell
apoptosis compared with the control and mimic-NC groups. However,
RBBP6 overexpression reversed miR-193a-5p overexpression-mediated
effects.
In conclusion, the results of the present study
suggested that upregulated expression levels of miR-193a-5p may
serve an inhibitory role in EOC via inhibiting cell proliferation
and migration, and promoting apoptosis. Moreover, the effects of
miR-193a-5p may be partly mediated by RBBP6. Collectively, the
results indicated that miR-193a-5p may serve as a potential
therapeutic target for EOC.
Acknowledgements
Not applicable.
Funding
No funding was received.
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
JL and JH collected and analyzed the data. SZ and NY
performed the experiments, conceived and designed the study,
confirm the authenticity of all the raw data and drafted and
revised the manuscript. All authors read and approved the final
manuscript.
Ethics approval and consent to
participate
The present study was approved by the Ethics
Committee of Beijing Ditan Hospital Capital Medical University.
Written informed consent was obtained from all participants.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
De A, De A, Sharma R, Suo W and Sharma M:
Sensitization of Carboplatinum- and Taxol-Resistant High-Grade
Serous Ovarian Cancer Cells Carrying p53, BRCA1/2 Mutations by
Emblica officinalis (Amla) via Multiple Targets. J Cancer.
11:1927–1939. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Chua KJC, Patel RD, Trivedi R, Greenberg
P, Beiter K, Magliaro T, Patel U and Varughese J: Accuracy in
Referrals to Gynecologic Oncologists Based on Clinical Presentation
for Ovarian Mass. Diagnostics (Basel). 10:1062020. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Huang D, Gaul DA, Nan H, Kim J and
Fernández FM: Deep Metabolomics of a High-Grade Serous Ovarian
Cancer Triple-Knockout Mouse Model. J Proteome Res. 18:3184–3194.
2019. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Giampaolino P, Della Corte L, Foreste V,
Vitale SG, Chiofalo B, Cianci S, Zullo F and Bifulco G: Unraveling
a difficult diagnosis: The tricks for early recognition of ovarian
cancer. Minerva Med. 110:279–291. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Leskela S, Romero I, Cristobal E,
Pérez-Mies B, Rosa-Rosa JM, Gutierrez-Pecharroman A, Santón A,
Gonzalez BO, López-Reig R, Hardisson D, et al: The Frequency and
Prognostic Significance of the Histologic Type in Early-stage
Ovarian Carcinoma: A Reclassification Study by the Spanish Group
for Ovarian Cancer Research (GEICO). Am J Surg Pathol. 44:149–161.
2020. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Della Corte L, Giampaolino P, Fabozzi A,
Cieri M, Zizolfi B, Morra I and Bifulco G: Breast metastasis two
years after pelvic surgery and adjuvant chemotherapy for serous
ovarian cancer. Gynecol Endocrinol. 35:211–213. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Buensuceso A, Ramos-Valdes Y, DiMattia GE
and Shepherd TG: AMPK-Independent LKB1 Activity Is Required for
Efficient Epithelial Ovarian Cancer Metastasis. Mol Cancer Res.
18:488–500. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Tato-Varela S and Kuhn W: Impact of
retroperitoneal lymph node dissection in ovarian cancer - time for
paradigm shift? Horm Mol Biol Clin Investig. 41:201900202019.
View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Kontomanolis EN, Fasoulakis Z, Papamanolis
V, Koliantzaki S, Dimopoulos G and Kambas NJ: The Impact of
microRNAs in Breast Cancer Angiogenesis and Progression. MicroRNA.
8:101–109. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Mucha O, Podkalicka P, Mikulski M, Barwacz
S, Andrysiak K, Biela A, Mieczkowski M, Kachamakova-Trojanowska N,
Ryszawy D, Białas A, et al: Development and characterization of a
new inhibitor of heme oxygenase activity for cancer treatment. Arch
Biochem Biophys. 671:130–142. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Grayson K, Gregory E, Khan G and Guinn BA:
Urine Biomarkers for the Early Detection of Ovarian Cancer - Are We
There Yet? Biomark Cancer. 11:1179299X198309772019. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Stewart D and Cristea M: Antibody-drug
conjugates for ovarian cancer: Current clinical development. Curr
Opin Obstet Gynecol. 31:18–23. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Zou X, Zhu D, Zhang H, Zhang S, Zhou X, He
X, Zhu J and Zhu W: MicroRNA expression profiling analysis in serum
for nasopharyngeal carcinoma diagnosis. Gene. 727:1442432020.
View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Jain N, Das B and Mallick B: Restoration
of microRNA-197 expression suppresses oncogenicity in fibrosarcoma
through negative regulation of RAN. IUBMB Life. 72:1034–1044. 2020.
View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Tani S, Kusakabe R, Naruse K, Sakamoto H
and Inoue K: Genomic organization and embryonic expression of
miR-430 in medaka (Oryzias latipes): Insights into the
post-transcriptional gene regulation in early development. Gene.
449:41–49. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Liu Z, Wang Y, Dou C, Sun L, Li Q, Wang L,
Xu Q, Yang W, Liu Q and Tu K: MicroRNA-1468 promotes tumor
progression by activating PPAR-γ-mediated AKT signaling in human
hepatocellular carcinoma. J Exp Clin Cancer Res. 37:492018.
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Dong D, Mu Z, Wei N, Sun M, Wang W, Xin N,
Shao Y and Zhao C: Long non-coding RNA ZFAS1 promotes proliferation
and metastasis of clear cell renal cell carcinoma via targeting
miR-10a/SKA1 pathway. Biomed Pharmacother. 111:917–925. 2019.
View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Shirafkan N, Shomali N, Kazemi T,
Shanehbandi D, Ghasabi M, Baghbani E, Ganji M, Khaze V, Mansoori B
and Baradaran B: MicroRNA-193a-5p inhibits migration of human HT-29
colon cancer cells via suppression of metastasis pathway. J Cell
Biochem. Dec 2–2018.(Epub ahead of print). doi:
10.1002/jcb.28164.
|
|
19
|
Chen J, Gao S, Wang C, Wang Z, Zhang H,
Huang K, Zhou B, Li H, Yu Z, Wu J, et al: Pathologically decreased
expression of miR-193a contributes to metastasis by targeting
WT1-E-cadherin axis in non-small cell lung cancers. J Exp Clin
Cancer Res. 35:1732016. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Yu T, Li J, Yan M, Liu L, Lin H, Zhao F,
Sun L, Zhang Y, Cui Y, Zhang F, et al: MicroRNA-193a-3p and −5p
suppress the metastasis of human non-small-cell lung cancer by
downregulating the ERBB4/PIK3R3/mTOR/S6K2 signaling pathway.
Oncogene. 34:413–423. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Ding N, Sun X, Wang T, Huang L, Wen J and
Zhou Y: miR-378a-3p exerts tumor suppressive function on the
tumorigenesis of esophageal squamous cell carcinoma by targeting
Rab10. Int J Mol Med. 42:381–391. 2018.PubMed/NCBI
|
|
22
|
Livak KJ and Schmittgen Td: Analysis of
relative gene expression data using real-time quantitative Pcr and
the 2(−delta deltac(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Hou M, Cheng Z, Shen H, He S, Li Y, Pan Y,
Feng C, Chen X, Zhang Y, Lin M, et al: Correction: High expression
of CTHRC1 promotes EMT of epithelial ovarian cancer (EOC) and is
associated with poor prognosis. Oncotarget. 11:825–826. 2020.
View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Xie Y, Zhao J, Liang Y, Chen M, Luo Y, Cui
X, Jiang B, Peng L and Wang X: MicroRNA-10b controls the metastasis
and proliferation of colorectal cancer cells by regulating
Krüppel-like factor 4. Artif Cells Nanomed Biotechnol.
47:1722–1729. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Hu S, Zang R, Wang Y, Liang Y, Mu J, Zhang
Y and Ma J: Highly expressed microRNA-124 inhibits migration and
promotes apoptosis of esophageal cancer cells by degrading PDCD6. J
BUON. 24:805–812. 2019.PubMed/NCBI
|
|
26
|
Yang Y, Zhou L, Lu L, Wang L, Li X, Jiang
P, Chan LK, Zhang T, Yu J, Kwong J, et al: A novel
miR-193a-5p-YY1-APC regulatory axis in human endometrioid
endometrial adenocarcinoma. Oncogene. 32:3432–3442. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Zhang Y, Jiang F, He H, Ye J, Mao X, Guo
Q, Wu SL, Zhong W, Wu CL and Lin N: Identification of a novel
microRNA-mRNA regulatory biomodule in human prostate cancer. Cell
Death Dis. 9:3012018. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Li J, Chen Y, Shen L and Deng Y:
Improvement of membranous nephropathy by inhibition of miR-193a to
affect podocytosis via targeting WT1. J Cell Biochem.
120:3438–3446. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Jin L, Li H, Wang J, Lin D, Yin K, Lin L,
Lin Z, Lin G, Wang H, Ying X, et al: MicroRNA-193a-5p exerts a
tumor suppressor role in glioblastoma via modulating NOVA1. J Cell
Biochem. 120:6188–6197. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Yang Z, Chen JS, Wen JK, Gao HT, Zheng B,
Qu CB, Liu KL, Zhang ML, Gu JF, Li JD, et al: Silencing of
miR-193a-5p increases the chemosensitivity of prostate cancer cells
to docetaxel. J Exp Clin Cancer Res. 36:1782017. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Finetti F, Moglia A, Schiavo I, Donnini S,
Berta GN, Di Scipio F, Perrelli A, Fornelli C, Trabalzini L and
Retta SF: Yeast-Derived Recombinant Avenanthramides Inhibit
Proliferation, Migration and Epithelial Mesenchymal Transition of
Colon Cancer Cells. Nutrients. 10:11592018. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Stepien A, Knop K, Dolata J, Taube M,
Bajczyk M, Barciszewska-Pacak M, Pacak A, Jarmolowski A and
Szweykowska-Kulinska Z: Posttranscriptional coordination of
splicing and miRNA biogenesis in plants. Wiley Interdiscip Rev RNA.
May 8–2017.(Epub ahead of print). doi: 10.1002/wrna.1403.
View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Movafagh S, Crook S and Vo K: Regulation
of hypoxia-inducible factor-1a by reactive oxygen species: New
developments in an old debate. J Cell Biochem. 116:696–703. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Sideris M and Papagrigoriadis S: Molecular
biomarkers and classification models in the evaluation of the
prognosis of colorectal cancer. Anticancer Res. 34:2061–2068.
2014.PubMed/NCBI
|
|
35
|
Maroof H, Salajegheh A, Smith RA and Lam
AK: Role of microRNA-34 family in cancer with particular reference
to cancer angiogenesis. Exp Mol Pathol. 97:298–304. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Wang MJ, Xu YY, Huang RY, Chen XM, Chen
HM, Han L, Yan YH and Lu CJ: Role of an imbalanced miRNAs axis in
pathogenesis of psoriasis: Novel perspectives based on review of
the literature. Oncotarget. 8:5498–5507. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Lin CH, Tsai CH, Yeh CT, Liang JL, Hung
WC, Lin FC, Chang WL, Li HY, Yao YC, Hsu TI, et al:
MiR-193a-5p/ERBB2 act as concurrent chemoradiation therapy response
indicator of esophageal squamous cell carcinoma. Oncotarget.
7:39680–39693. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Yoshitake Y, Nakatsura T, Monji M, Senju
S, Matsuyoshi H, Tsukamoto H, Hosaka S, Komori H, Fukuma D, Ikuta
Y, et al: Proliferation potential-related protein, an ideal
esophageal cancer antigen for immunotherapy, identified using
complementary DNA microarray analysis. Clin Cancer Res.
10:6437–6448. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Moela P, Choene MM and Motadi LR:
Silencing RBBP6 (Retinoblastoma Binding Protein 6) sensitises
breast cancer cells MCF7 to staurosporine and camptothecin-induced
cell death. Immunobiology. 219:593–601. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Motadi LR, Bhoola KD and Dlamini Z:
Expression and function of retinoblastoma binding protein 6 (RBBP6)
in human lung cancer. Immunobiology. 216:1065–1073. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Moela P and Motadi LR: RBBP6: A potential
biomarker of apoptosis induction in human cervical cancer cell
lines. OncoTargets Ther. 9:4721–4735. 2016. View Article : Google Scholar : PubMed/NCBI
|