Introduction
Colorectal cancer (CRC) is a primary cause of
cancer-related deaths, with >1.2 million newly diagnosed cases
every year worldwide (1). Although
great achievements have been made in diagnostic approaches and
therapeutic strategies, the clinical outcome of advanced CRC
remains poor (2). It has been
identified that the survival rate of patients with CRC is highly
associated with the stage, with ~90% for stage I and only 25% for
stage II (3). Therefore, there is
an urgent requirement to identify reliable biomarkers of CRC
progression and provide potential therapeutic targets.
Long non-coding (lnc)RNAs are a group of non-coding
(nc)RNAs with a length of >200 nucleotides (4). For a long time, lncRNAs were
considered the ‘junk’ of the genome due to their inability to
encode proteins (5). In previous
years, accumulating evidence has demonstrated that lncRNAs serve
essential roles in the initiation and progression of cancer, and
can therefore be used as reliable diagnostic and prognostic
biomarkers (6). Notably, lncRNAs
are relatively stable and it is feasible to detect cancer-related
lncRNAs in blood and/or other body fluids (7). lncRNA nicotinamide nucleotide
transhydrogenase antisense RNA 1 (NNT-AS1) is a newly-identified
lncRNA that is located in the chromosome 5p12 region, with a full
length of 3,304 bp (8). NNT-AS1 is
abnormally expressed in various types of cancer, including cervical
cancer, osteosarcoma, gastric cancer and lung cancer (9–11).
NNT-AS1 is also identified to be highly upregulated in CRC tissues
compared with adjacent normal tissues and can contribute to the
migration and invasion of CRC cells (12). However, the clinical significance of
circulating NNT-AS1 expression in CRC has not yet been
investigated.
Previous studies have identified that nanosized
vesicles, also known as exosomes, may function as natural carriers
to transport proteins, mRNAs and various ncRNAs (13,14).
Exosomes can be identified in various human body fluids, such as
plasma, saliva, serum and urine (14). Their double layer construction makes
exosomes ideal vehicles that protect biomolecules from lysosomal
and/or other enzymes. Exosomes are involved not only in
cell-to-cell communication, but also in the progression of
tumorigenesis (15). Certain
lncRNAs can be selectively packaged into exosomes, which are
correlated with the clinicopathological features of cancer, and
therefore may have useful applications as cancer-related biomarkers
(16).
MicroRNAs (miRNAs/miRs) are another type of ncRNA
with a length of ~20 nucleotides. A number of studies have
identified that miRNAs are involved in the pathogenesis of various
human diseases, including types of human cancers (17). Among such miRNAs, miR-496 has been
identified as a regulator in the progression of different cancers,
including lung cancer, breast cancer and osteosarcoma (18,19).
However, there are few studies on the role of miR-496 in CRC.
Ras-related protein Rap-2c (RAP2C) belongs to the GTP-binding
protein family and has been identified to participate in the
development of different types of cancer. For instance, RAP2C
promotes the migration and invasion of osteosarcoma cells (20). In addition, RAP2C has also been
identified to promote the tumorigenesis of breast cancer and
laryngeal squamous cell carcinoma (21,22).
However, there remains a lack of studies regarding the role of
RAP2C in CRC.
The current study evaluated the expression levels of
NNT-AS1 in CRC tissues and adjacent normal tissues, followed by
analysis of serum NNT-AS expression in patients with CRC and
healthy controls. The presence of serum NNT-AS1 in exosomes was
also determined and it indicated that exosomal NNT-AS1 may be a
novel biomarker for CRC. In addition, the role of NNT-AS1 in CRC
was also investigated and it was demonstrated that NNT-AS1 exerted
its effects on CRC cells via regulation of the miR-496/RAP2C
axis.
Materials and methods
Patient and healthy control
specimens
The present study was approved by the Ethical
Committee of Hwamei Hospital, University of Chinese Academy of
Sciences (Ningbo, China; approval no. 43321764) and all procedures
were conducted according to the Declaration of Helsinki. Written
informed consent was obtained from each patient. All patients and
healthy controls involved in this study were recruited from Hwamei
Hospital between January 2017 and August 2019 according to the
following criteria: i) Patients who underwent surgery with curative
intent; ii) relevant clinical features (Table I) are available; iii) patients who
did not receive radiotherapy or chemotherapy; iv) patients without
other tumors. A total of 40 resected tumor tissues and adjacent
normal tissues (≥1 cm from tumor tissue) were obtained at the
Department of General Surgery. The diagnosis of CRC was
histopathologically confirmed and other clinical data, including
age, sex, tumor location, tumor size, differentiation, lymph node
metastasis and distant metastasis, were collected. The
postoperative pathological staging was evaluated based on the 7th
edition of the Union for International Cancer Control
tumor-node-metastasis (TNM) staging system (23). Serum samples were obtained from 40
age- and sex-matched healthy volunteers. All patients underwent
physical examinations at Hwamei Hospital before recruitment. All
resected tissue samples were immediately snap-frozen in liquid
nitrogen. Venous blood was collected and prepared as previously
described and stored at −80°C until analysis (14).
 | Table I.Relationship between NNT-AS1
expression and clinical characteristics of patients with CRC. |
Table I.
Relationship between NNT-AS1
expression and clinical characteristics of patients with CRC.
|
|
| Tissue expression
of NNT-AS1 |
| Serum expression of
NNT-AS1 |
|
|---|
|
|
|
|
|
|
|
|---|
|
Characteristics | n | High | Low | P-value | High | Low | P-value |
|---|
| Sex |
|
|
| 0.371 |
|
| 0.446 |
|
Female | 14 | 6 | 8 |
| 7 | 7 |
|
|
Male | 26 | 14 | 12 |
| 11 | 15 |
|
| Age, years |
|
|
| 0.292 |
|
| 0.503 |
|
≤60 | 10 | 4 | 6 |
| 4 | 6 |
|
|
>60 | 30 | 17 | 13 |
| 14 | 16 |
|
| Tumor location |
|
|
| 0.578 |
|
| 0.252 |
|
Colon | 18 | 7 | 11 |
| 11 | 7 |
|
|
Rectum | 22 | 9 | 13 |
| 10 | 12 |
|
| Tumor size, cm |
|
|
| 0.184 |
|
| 0.564 |
| ≤4 | 25 | 10 | 15 |
| 11 | 14 |
|
|
>4 | 15 | 9 | 6 |
| 7 | 8 |
|
| TNM stage |
|
|
| 0.033 |
|
| 0.576 |
|
I–II | 23 | 7 | 16 |
| 9 | 14 |
|
|
III–IV | 17 | 11 | 6 |
| 7 | 10 |
|
| Local invasion |
|
|
| 0.025 |
|
| 0.252 |
|
T1-T2 | 18 | 5 | 13 |
| 11 | 7 |
|
|
T3-T4 | 22 | 14 | 8 |
| 10 | 12 |
|
| Lymph node |
|
|
| 0.004 |
|
| 0.501 |
| No | 21 | 7 | 14 |
| 11 | 10 |
|
|
Yes | 19 | 15 | 4 |
| 9 | 10 |
|
Serum exosome isolation
Exosomes were isolated and purified from 0.5 ml
serum using a Total Exosome Isolation Kit (cat. no. 4484450, Thermo
Fisher Scientific, Inc.) according to the manufacturer's
instructions. The exosomes were then suspended in 1X
phosphate-buffered saline (PBS) and stored at −80°C before use.
Cell culture and transfection
Human normal colorectal epithelial cells (HCnEpC)
and human CRC cells (LoVo, RKO, SW48 and HCT116) were purchased
from The Cell Bank of Type Culture Collection of the Chinese
Academy of Sciences. All cells were cultured at a density of
1×106 cells/ml in RPMI-1640 medium (HyClone; Cytiva)
supplemented with 10% fetal bovine serum (FBS; Gibco; Thermo Fisher
Scientific, Inc.), 100 U/ml penicillin and 100 µg/ml streptomycin
(Thermo Fisher Scientific, Inc.) at 37°C with 5% CO2.
For transfection, small interfering (si)RNA against NNT-AS1
(si-NNT-AS1, 5′-GAAAAGAAAAAGAAGCUUATT-3′), scramble siRNA negative
control (si-NC, 5′-UUCUCCGAACGUGUCACG-3′), pcDNA3.1 vector (empty
control), NNT-AS1 overexpression vector (pcDNA. NNT-AS1), miR-496
mimic (5′-UGAGUAUUACAUGGCCAAUCUC-3′), scramble negative control
miRNA mimic (miR-NC) (5′-ACCGUACACGUUACCGUCACU-3′), miR-496
inhibitor (5′-GAGAUUGGCCAUGUGUAAUACUCA-3′), miR-NC inhibitor
(5′-CAUAACGACUACGGUACCAUCA-3′) and RAP2C overexpression vector
(pcDNA. RAP2C) were all synthesized by Shanghai GenePharma Co.,
Ltd. All transfections were conducted using
Lipofectamine® 2000 (Invitrogen; Thermo Fisher
Scientific, Inc.) according to the manufacturer's instructions. The
working concentration of siRNA was 20 nM and that for miRNA mimics
and inhibitors was 50 nM. The concentration used for plasmids was
100 nM. At 24 h after the transfection, following assays were
conducted.
Total RNA purification and reverse
transcription-quantitative (RT-q)PCR
Total RNA was purified from tissues (5 g)/serum (5
ml) using TRIzol® reagent (Thermo Fisher Scientific,
Inc.) and exosomal RNA of serum was extracted using the mirVana™
PARIS™ RNA and Native Protein Purification Kit (Thermo Fisher
Scientific, Inc.) according to the manufacturer's instructions. The
quantity and quality of RNA were measured using a NanoDrop™ 2000
Spectrophotometer (Thermo Fisher Scientific, Inc.). cDNA was
generated by reverse transcription using a PrimeScript RT Reagent
Kit (Takara Biotechnology Co., Ltd.) according to the
manufacturer's instructions. SYBR® Premix Ex Taq™ II
(Takara Biotechnology Co., Ltd.) was used to perform RT-qPCR and
reactions were run in duplicate at 95°C for 30 sec for 1 cycle,
followed by 40 cycles of 95°C for 3 sec and 60°C for 30 sec. The
total reaction volume was 50 µl. Primers were designed as follows:
NNT-AS1, forward 5′-ACGTGCAGACAACATCTACCT-3′ and reverse
5′-TACAACACCTTCCCGCAT-3′; miR-496, forward
5′-GCAATCCTGAGGAAGAGGATGTGGA-3′ and reverse
5′-TCTGAGCTGCGAGTCATT-3′; GAPDH, forward
5′-CCCACTTGAAGGGTGGAGCCAA-3′ and reverse
5′-TGGCATGGACTGTGGTCATGA-3′; Tubulin, forward
5′-ATGCGTGAAATAGTTCATATCC-3′ and reverse
5′-TTAGATTTCTTCTTCCTCGTC-3′; and U6, forward
5′-GCTTCACGAATTTGCGTGTCAT-3′ and reverse
5′-GCTTCGGCAGCACATATACTAAAAT-3′. The expression of lncRNA NNT-AS1
was expressed as the 2−ΔΔCq value, with GAPDH and
Tubulin as internal controls for NNT-AS1 and U6 as the internal
control for miR-496 (24).
NanoSight assay flow cytometry
analysis
Exosome pellets were resuspended in 1 ml PBS and the
particle size distribution was analyzed under a NanoSight NS300
(Malvern Instruments, Ltd).
Flow cytometry analysis of
exosomes
For the analysis of exosomes by
fluorescence-activated cell sorting, the exosomes were mixed with
100 µl PBS at room temperature for 5 min and then analyzed with
monoclonal antibodies against CD9 (1:200; cat. no. ab2215; Abcam),
CD63 (1:200; cat. no. ab193349; Abcam) and CD81 (1:200; cat. no.
ab219209; Abcam) by flow cytometry (BD FACSCalibur; BD
Biosciences). The results were analyzed with FlowJo 11.0 (TreeStar
Ltd.).
Cell viability assay
Cell viability was measured using an MTT assay as
described previously (24).
Briefly, cells were seeded in 6-well plates at a density of
1×105 cells/well. Following transfection, 20 µl of 0.5
mg/ml MTT (Sigma-Aldrich; Merck KGaA) was added to the medium and
incubated for another 4 h at 37°C. Before measurement, 150 µl DMSO
was added and a microplate reader (BioTek Instruments, Inc.) was
used to measure the optical density value at 450 nm.
Dual-luciferase assays
The wild-type (wt) and mutant (mut) fragments of
NNT-AS1 and the 3′ untranslated region of RAP2C containing the
miR-496 targeted sequences were synthesized by Shanghai GenePharma
Co., Ltd. and inserted into the pGL3 promoter vector (Promega
Corporation). The transfection was conducted using
Lipofectamine® 2000 (Thermo Fisher Scientific, Inc.)
according to the manufacturer's guide. The concentration of plasmid
was 100 ng. Then, 48 h after transfection, cells were collected and
the Dual-Luciferase Reporter Assay System (Promega Corporation) was
used to measure luminescence. Renilla luciferase activity
was used to normalize the firefly luciferase activity.
Cell migration and invasion assay
The wound healing assay was performed using 6-well
plates (Corning Life Sciences). A total of 1×106 cells
were seeded into each well and cultured with 5% serum (Gibco;
Thermo Fisher Scientific, Inc.) medium overnight until 90%
confluent. A 200-µl pipette tip was used to make a straight
scratch. The floating cells were gently washed off using PBS and
fresh full medium was added. The initial gap length (0 h) and the
residual gap length at 24 h after wounding were calculated from
photomicrographs as previously described (25). The results were observed using the
IX70 inverted light microscope (magnification, ×100; Olympus
Corporation). For the cell invasion assay, a Transwell chamber
(Corning Life Sciences) was used. The upper chamber was coated with
Matrigel at room temperature for 4 h (BD Biosciences) for
filtering, and culture medium containing 20% FBS (Gibco; Thermo
Fisher Scientific, Inc.) was added to the lower chamber. Then,
1×105 glioma cells were cultured in 2 ml serum-free
medium and placed in the upper chamber. After 48 h at 37°C,
non-invasive cells were removed and invasive cells were fixed with
70% ethanol at room temperature for 0.5 h and stained with 5%
crystal violet (Beyotime Institute of Biotechnology) at room
temperature for 0.5 h. The number of invading cells to the lower
chamber was counted under the IX70 inverted optical microscope
(magnification, ×100; Olympus Corporation); five fields were
randomly selected from each sample. This experiment was repeated
three times.
Western blot analysis
Total protein was collected with RIPA buffer
(Beyotime Institute of Biotechnology) and the concentration of
protein was measured using a BCA protein assay kit (Beyotime
Institute of Biotechnology). Protein (20 µg) was separated via
SDS-PAGE on a 10% gel, and then transferred to a PVDF membrane (EMD
Millipore). After blocking with 5% skimmed milk for 1 h at room
temperature, the membranes were incubated with primary antibodies
overnight at 4°C. The following primary antibodies were used:
Anti-CD63 (1:1,000; cat. no. ab193349; Abcam), anti-tumor
susceptibility gene 101 protein (1:1,000; cat. no. ab125011;
Abcam), anti-CD9 (1:1,000; cat. no. ab92726; Abcam); and
anti-Flotillin-1 (1:1,000; cat. no. ab41927; Abcam). Anti-RAP2C
(1:1,000; cat. no. ab97805; Abcam), GAPDH (1:5,000; cat. no.
ab9485; Abcam). Subsequently, the membrane was washed three times
with TBS with 1% Tween-20 for 10 min, and then incubated with goat
anti-rabbit HRP-conjugated secondary antibody (1:5,000; cat. no.
ab6721; Abcam) for 1 h at room temperature. Then, the membrane was
visualized with enhanced chemiluminescent solution (Beijing
Solarbio Science & Technology Co., Ltd.). The results were
visualized using the ChemiDoc XRS+ GelDoc™ 5.0 (BioRad
Laboratories, Inc.).
Bioinformatical analysis
Prediction of the NNT-AS1-miRNA-target gene was
performed using the StarBase 3.0 (starbase.sysu.edu.cn) and TargetScan 2.0 (https://targetscan.org/).
Statistical analysis
Statistical analyses were performed using SPSS 22.0
(IBM Corp.) and GraphPad Prism 6.0 (GraphPad Software, Inc.).
Comparisons of continuous outcomes between groups were performed
using a two-tailed, paired Student's t-test and multiple groups
were analyzed with one-way analysis of variance (ANOVA), followed
by Tukey's post hoc test. The receiver operating characteristic
(ROC) curves and area under the curve (AUC) values were calculated
using algorithms described previously (26). P<0.05 was considered to indicate
a statistically significant difference.
Results
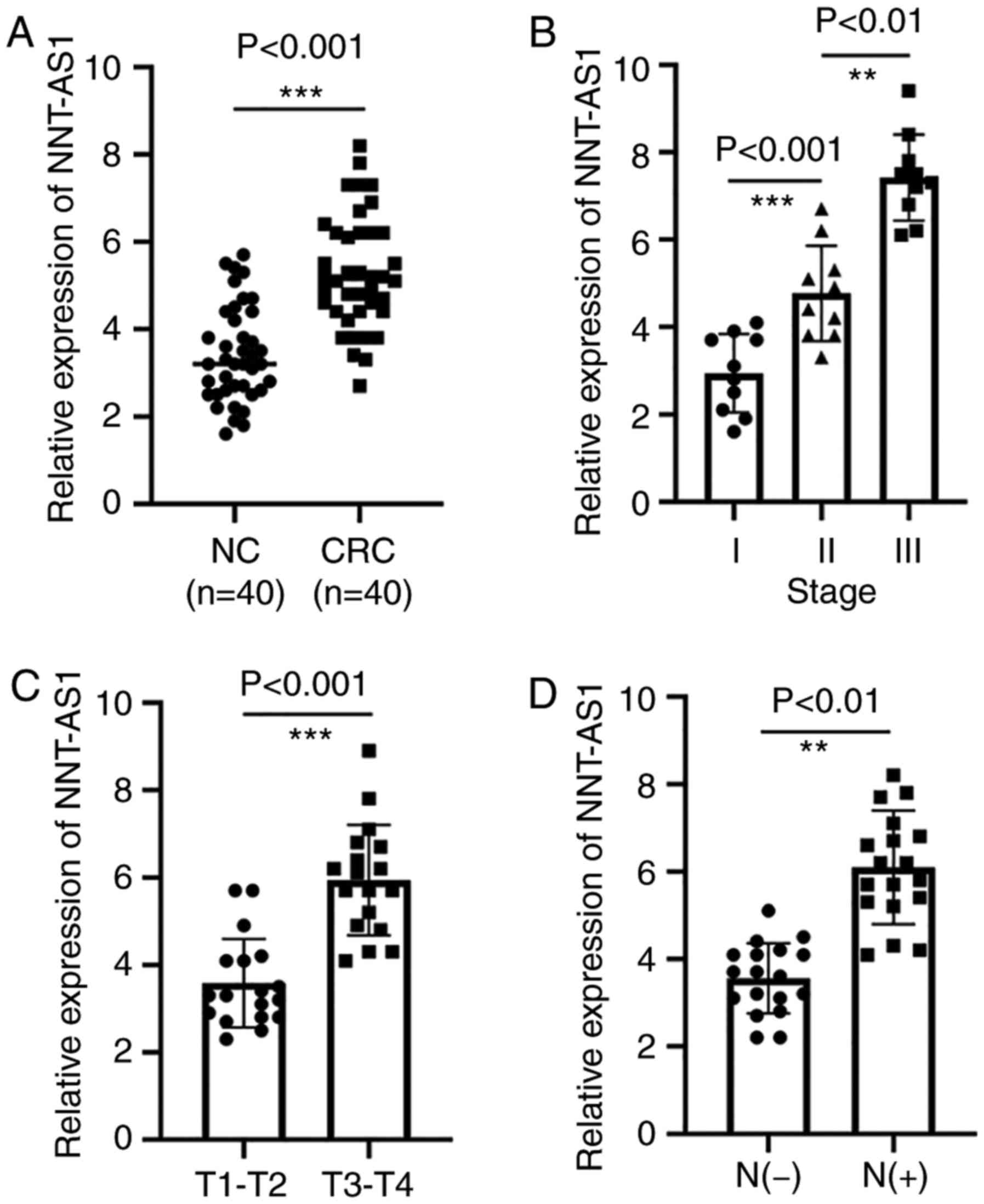
NNT-AS1 is upregulated in CRC
tissues
The expression levels of NNT-AS1 were measured in a
set of 40 paired tissues by RT-qPCR. The expression of NNT-AS1 in
tumor tissues was significantly higher than that in adjacent normal
tissues (P<0.001, Fig. 1A). The
association of NNT-AS1 levels with several clinicopathological
features was then examined (Table
I). It was identified that patients with CRC at advanced stages
had higher NNT-AS1 levels (P<0.01, Fig. 1B). It was also identified that the
level of NNT-AS1 was significantly associated with local invasion
(P<0.001, Fig. 1C). In addition,
high expression of NNT-AS1 was associated with lymph node
metastasis (P<0.001, Fig. 1D).
The expression of NNT-AS1 was not related to gender, age, tumor
size or tumor location (P>0.05, Table I).
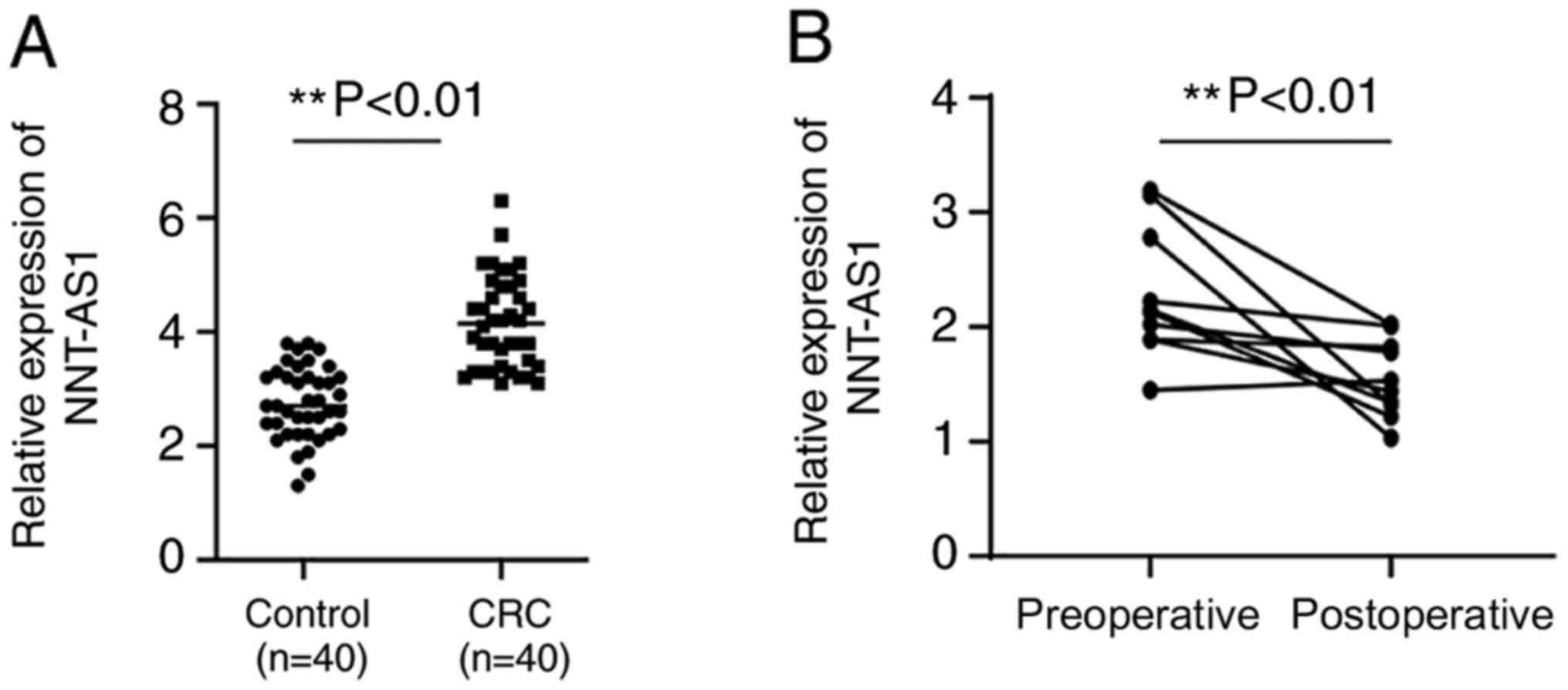
Expression levels of serum NNT-AS1 in
patients with CRC
Serum NNT-AS1 expression levels were assayed in
patients with CRC and healthy controls. Serum levels of NNT-AS1
were significantly higher in patients compared with healthy
controls (P<0.01, Fig. 2A). No
significant associations were observed between serum NNT-AS1 levels
and clinical features such as gender, age, tumor size and location,
local invasion, lymph node metastasis and stage (P>0.05,
Table I). The diagnostic value of
serum NNT-AS1 levels was then assayed in patients with CRC.
Serum NNT-AS1 levels between
preoperative and postoperative samples
To examine the changes in NNT-AS1 expression levels
after surgery, the expression levels of NNT-AS1 in eight paired
preoperative and postoperative serum samples were compared. The
results indicated that the expression levels of NNT-AS1 were
significantly decreased in postoperative samples compared with
preoperative samples (P<0.01, Fig.
2B).
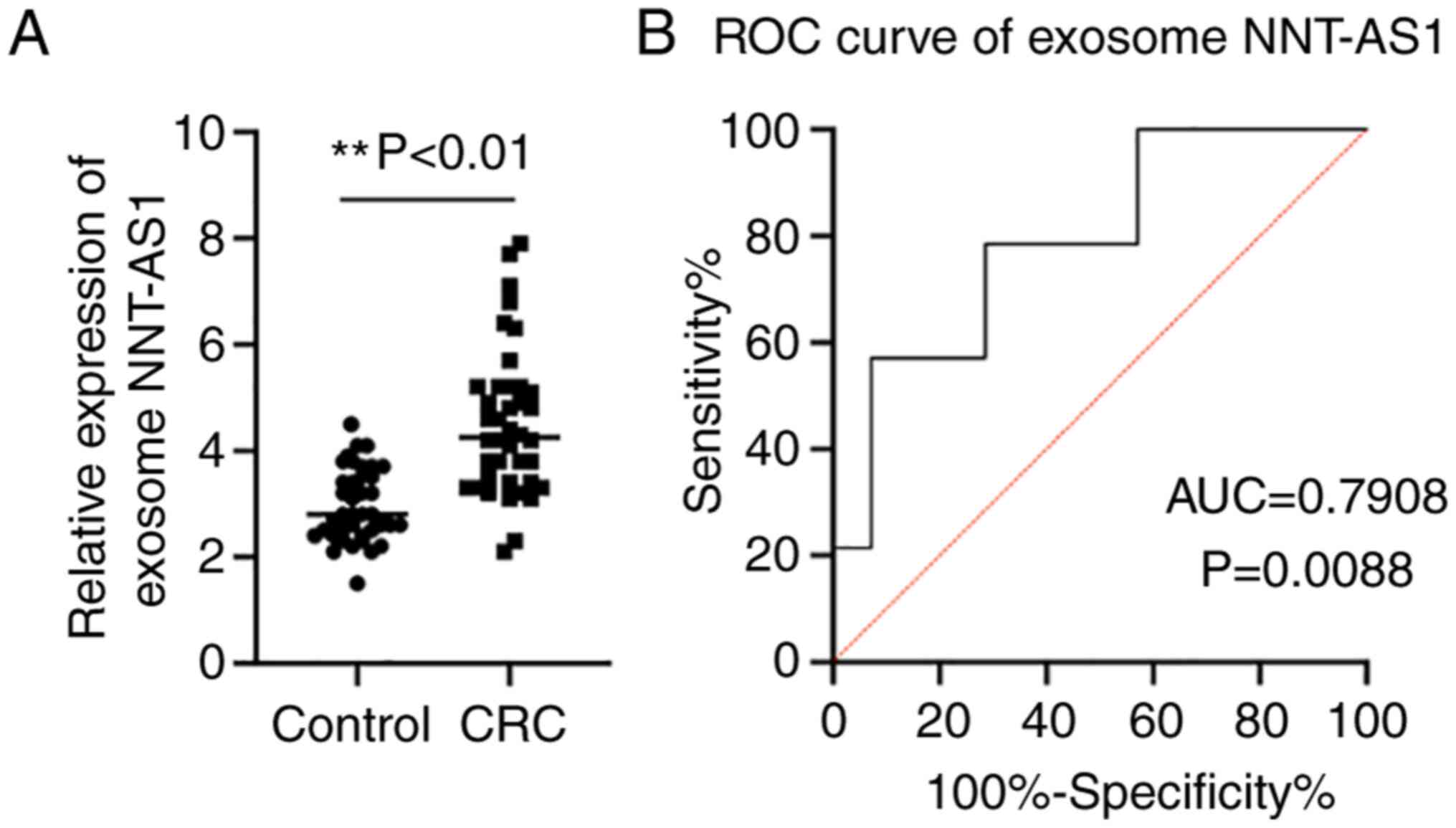
Exosome NNT-AS1 expression levels in
patients with CRC and healthy controls
Accumulating evidence has demonstrated that exosomes
serve an essential role in maintaining the stability of lncRNAs
(13). Thus, the expression levels
of NNT-AS1 in exosomes was assayed. The exosome levels of NNT-AS1
in the same cohort of patients with CRC and healthy controls were
compared by RT-qPCR. The results demonstrated that NNT-AS1
expression was upregulated in CRC exosomes compared with the
controls (P<0.01, Fig. 3A).
Then, ROC curve analyses were performed to further elucidate the
potential diagnostic value of serum exosomal NNT-AS1. The AUC of
serum exosomal NNT-AS1 was 0.7908 (95% CI, 0.6226-0.9590; P=0.0088;
Fig. 3B).
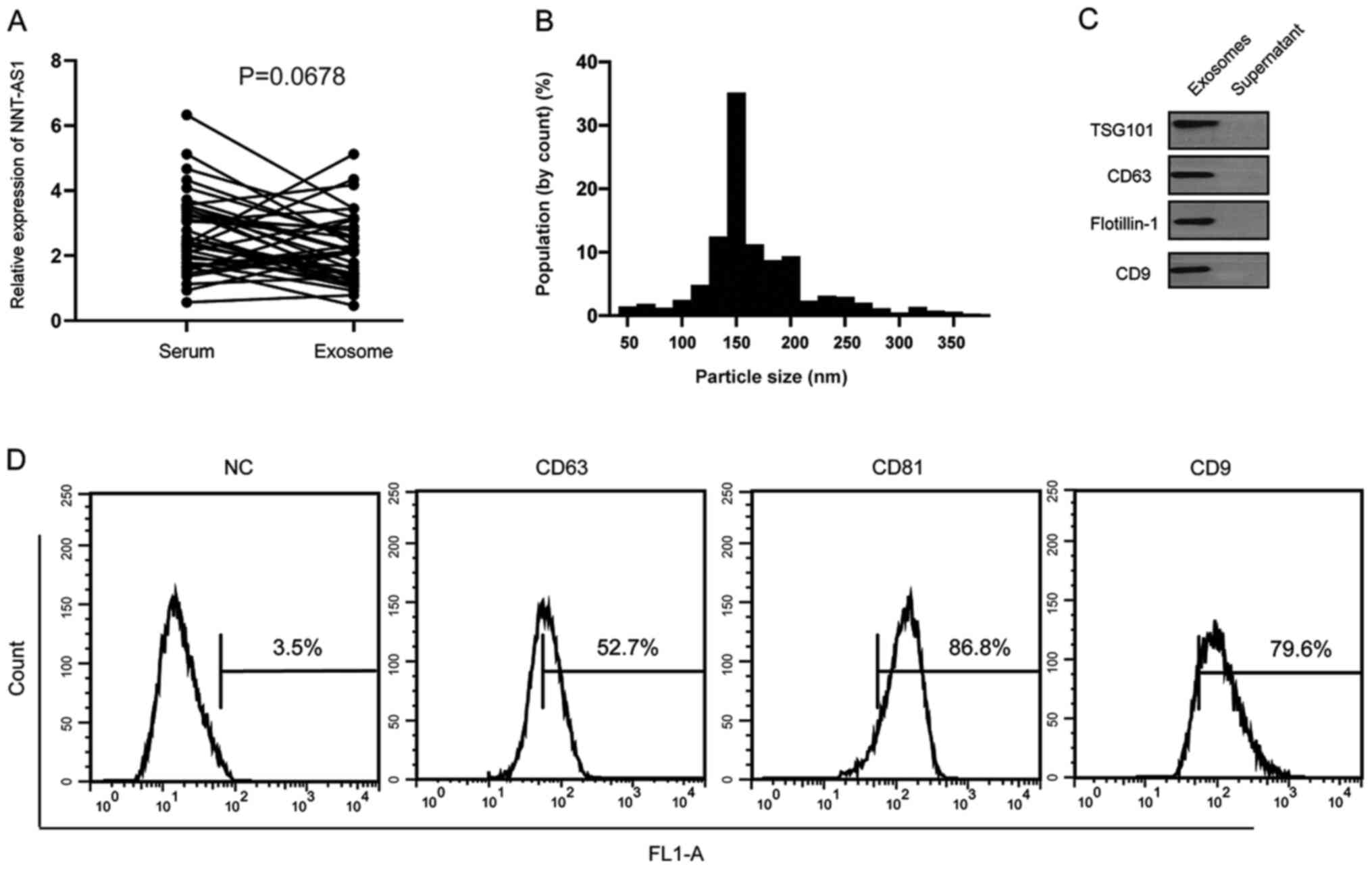
Serum NNT-AS1 is contained in
exosomes
It was hypothesized that serum NNT-AS1 is secreted
from tumor cells or is protected by exosomes. The levels of NNT-AS1
were measured in 40 pairs of serum samples and exosomes isolated
from the same volume of serum. The results demonstrated that there
were no significant differences in NNT-AS1 expression levels
between serum and exosomes (P=0.0678; Fig. 4A). To further identify the isolated
particles, nanoparticle tracking analysis was performed to measure
the concentration and size range of the particles. The data
demonstrated that the diameters of 10% of the particles were ~100
nm (Fig. 4B). Western blotting and
flow cytometry analyses further confirmed the presence of the
surface biomarkers of exosomes (Fig. 4C
and D).
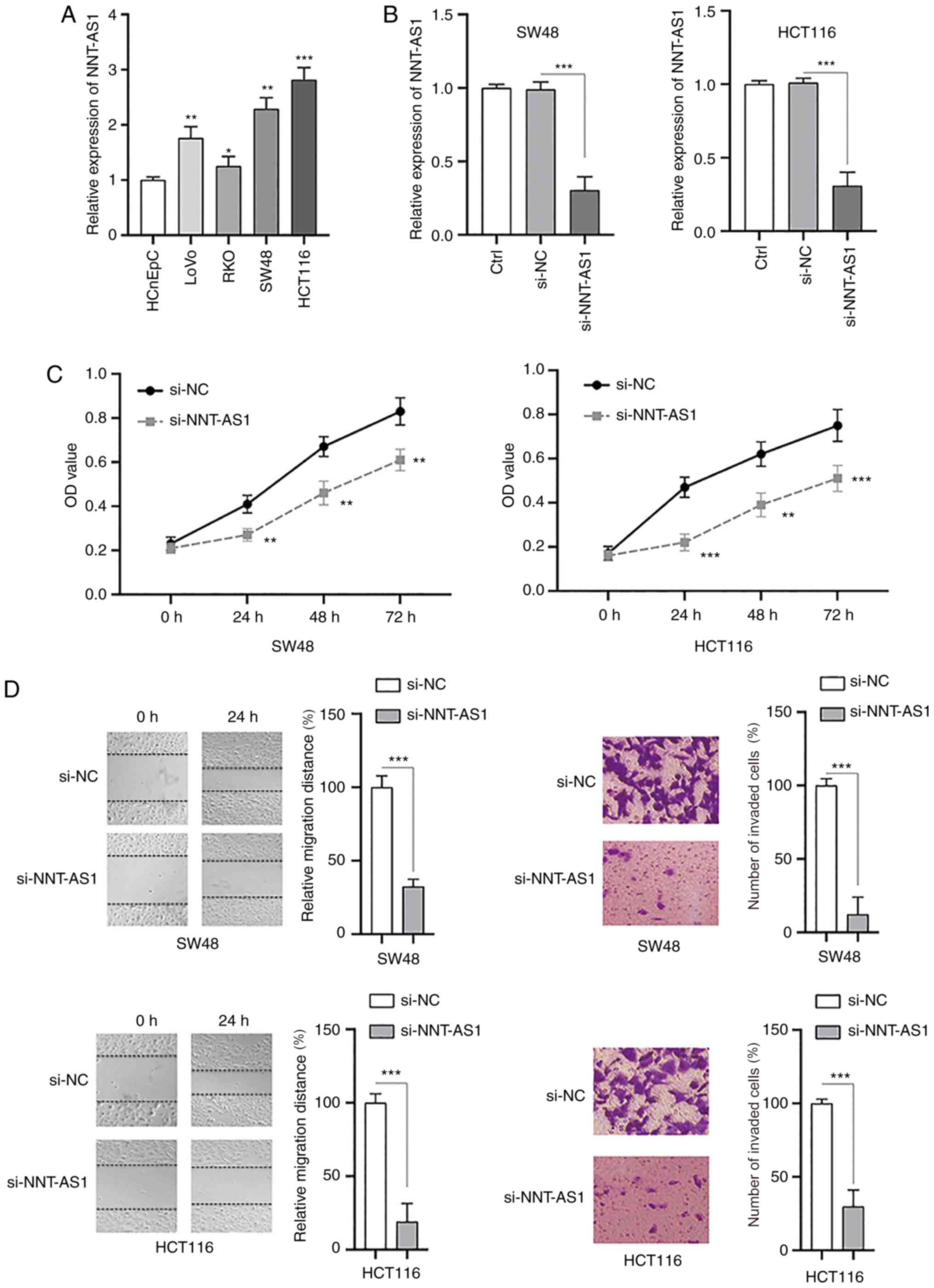
Knockdown of NNT-AS1 inhibits the
proliferation, migration and invasion of CRC cells
The biological functions of NNT-AS1 in CRC cells
were investigated in vitro. The levels of NNT-AS1 in human
colorectal normal cells (HCnEpC) and CRC cell lines (LoVo, RKO,
SW48 and HCT116) were measured by RT-qPCR. It was identified that
the levels of NNT-AS1 were significantly higher in CRC cells
compared with normal cells (P<0.05, Fig. 5A). Next, siRNA (si-NNT-AS1) was used
to successfully silence the expression of NNT-AS1 in SW48 and
HCT116 cells compared with the controls (P<0.001; Fig. 5B). MTT assays demonstrated that the
proliferation of both SW48 and HCT116 cells was significantly
inhibited after silencing NNT-AS1 (P<0.01; Fig. 5C). The migration and invasion of CRC
cells were also significantly decreased after knockdown of NNT-AS1
(P<0.001; Fig. 5D). Taken
together, these data suggested that NNT-AS1 may function as an
oncogene in CRC cells.
NNT-AS1 acts as a sponge of
miR-496
Substantial evidence has indicated that various
lncRNAs can act as a sponge to bind and negatively regulate the
expression of miRNAs (7).
Therefore, bioinformatic tools (StarBase 2.0 and TargetScan) were
used to predict the potential targets of NNT-AS1. miR-496 was
identified and the putative binding sites between NNT-AS1 and
miR-496 are indicated in Fig. 6A.
Transfection of miR-496 mimics successfully upregulated the
expression of miR-496 in both SW48 and HCT116 cells (P<0.01;
Fig. 6B). The interaction between
NNT-AS1 and miR-496 was then examined by the luciferase activity
assay. The data indicated that miR-496 mimics significantly
suppressed the luciferase activity of CRC cells transfected with wt
NNT-AS1, whereas those transfected with mut NNT-AS1 were not
affected (P>0.05; Fig. 6C).
Next, NNT-AS1 was overexpressed by transfection of pcDNA3.NNT-AS1
in SW48 and HCT116 cells (P<0.01; Fig. 6D). In addition, the expression of
miR-496 was significantly inhibited by the overexpression of
NNT-AS1 and increased by the silencing of NNT-AS1 in SW48 and
HCT116 cells (P<0.05; Fig. 6E).
MTT assay demonstrated that overexpression of miR-496 inhibited the
proliferation of CRC cells, which was similar to the effects of
NNT-AS1 downregulation (P<0.05; Fig.
6F). Overexpression of miR-496 also inhibited the migration
(P<0.001; Figs. 6G and S1A) and invasion (P<0.001; Figs. 6H and S1B) of CRC cells. To further investigate
the function of miR-496 in CRC cells, transfection with an miR-496
inhibitor, which successfully decreased the expression of miR-496,
was conducted in CRC cells (P<0.001; Fig. 6I). The effects of silencing NNT-AS1
on the migration, invasion (P<0.01; Figs. 6J and S2A and B) and proliferation (P<0.01;
Fig. 6K) of CRC cells could be
partially attenuated by inhibition of miR-496. A previous study
identified that NNT-AS1 could also bind and inhibit the expression
of miR-320 (26). However, no
significant difference was observed between the expression of
miR-320 in CRC and adjacent normal tissues (data not shown). Taken
together, these data suggested that NNT-AS1 functions as a sponge
of miR-496.
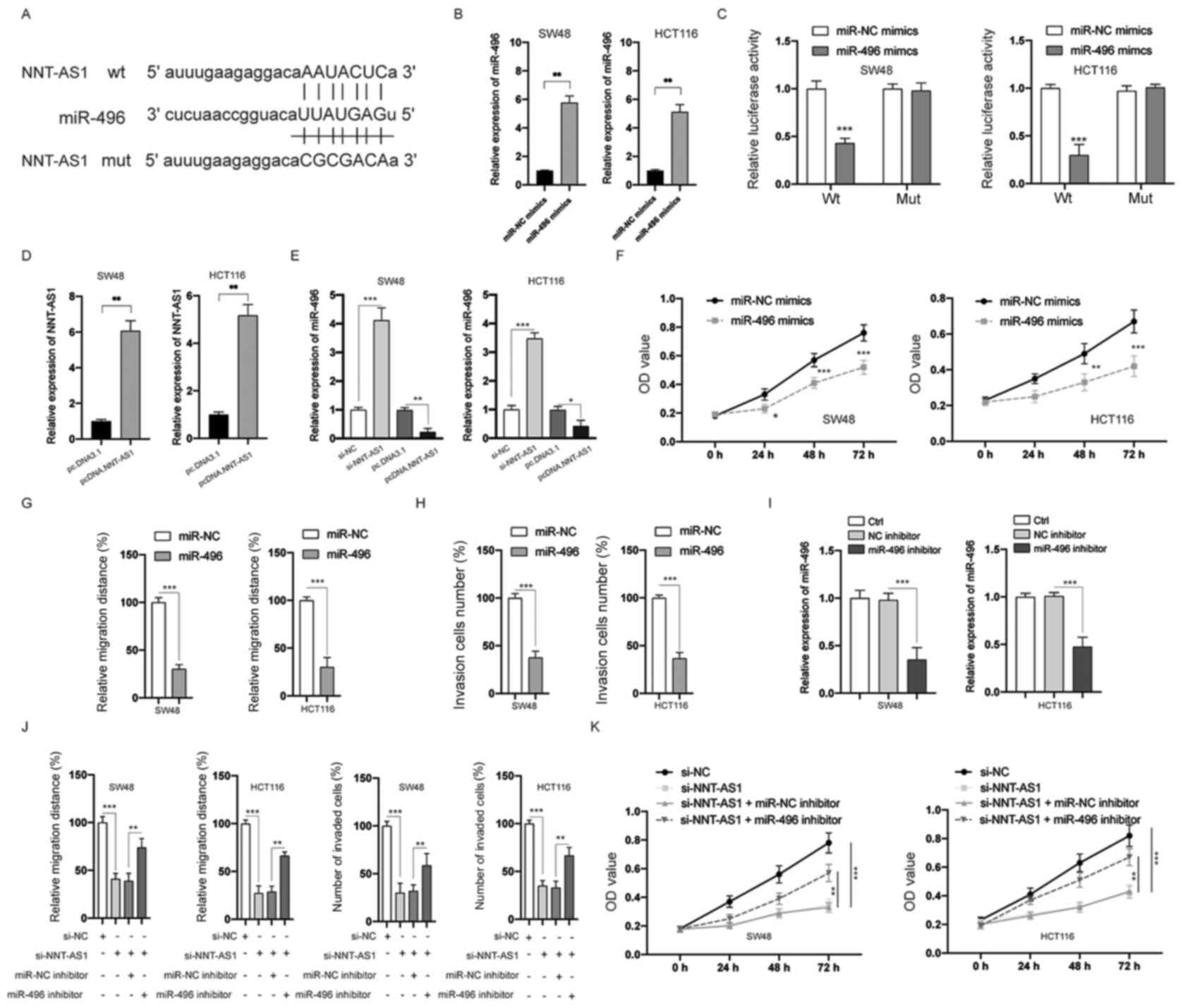 | Figure 6.NNT-AS1 is a sponge of miR-496. (A)
The predicted miR-496 binding sites on wt or mut NNT-AS1. (B) CRC
cells were transfected as indicated, and the expression of miR-496
was measured via RT-qPCR. (C) A luciferase reporter assay was
performed to measure the luciferase activity in CRC cells after
co-transfection of miRNA mimics and NNT-AS1 wt or mut. (D) CRC
cells were transfected as indicated, and the expression of NNT-AS1
was measured via RT-qPCR. (E) CRC cells were transfected as
indicated and the expression of miR-496 was measured. (F) CRC cells
were transfected as indicated and cell viability was measured by
the MTT assay. (G) Cell migration was measured. (H) Cell invasion
was measured. (I) The levels of miR-496 in CRC were measured. (J)
CRC cells were transfected as indicated and the migration and
invasion of cells were measured. (K) CRC cells were transfected as
indicated and cell viability was measured. Data are presented as
the mean ± standard deviation. *P<0.05, **P<0.01 and
***P<0.001 vs. miR-NC mimics or as indicated. All experiments
were conducted at least three times. NNT-AS1, lncRNA nicotinamide
nucleotide transhydrogenase antisense RNA 1; CRC, colorectal
cancer; miR, microRNA; NC, negative control; si-, small interfering
RNA; wt, wild-type; mut, mutant; pcD, overexpression vector;
RT-qPCR, reverse transcription-quantitative PCR. |
RAP2C is a direct target of
miR-496
Next, the possible targets of miR-496 were examined.
Through bioinformatics analysis, RAP2C was identified as a possible
direct target of miR-496 and the putative binding sites are
indicated in Fig. 7A. The possible
interaction between miR-496 and RAP2C was then validated by the
dual-luciferase reporter assay. The results demonstrated that the
luciferase activity of wt RAP2C was significantly inhibited by
miR-496 mimics, whereas the luciferase activity of mut RAP2C was
not affected (Fig. 7B).
Furthermore, transfection of miR-496 mimics suppressed the
expression of RAP2C (Fig. 7C, top
panel) and downregulation or upregulation of NNT-AS1 resulted in
the inhibition or increase of RAP2C (Fig. 7C, bottom panel), respectively. These
findings suggested that NNT-AS1 may exert its effects via
regulation of the miR-496/RAP2C axis. To further examine the
function of RAP2C, RAP2C was overexpressed in CRC cells via
transfection of a vector containing RAP2C (Fig. 7D). It was identified that the
upregulation of RAP2C partially abrogated the effects of NNT-AS1
silencing on the proliferation of CRC cells (Fig. 7E). In addition, overexpression of
RAP2C also partially rescued the inhibition of migration (Figs. 7F and S3A) and invasion (Figs. 7G and S3B) of CRC cells caused by NNT-AS1
knockdown. Taken together, these data suggested that RAP2C was a
direct target of miR-496 and that NNT-AS1 exerted its effects via
regulation of the miR-496/RAP2C axis.
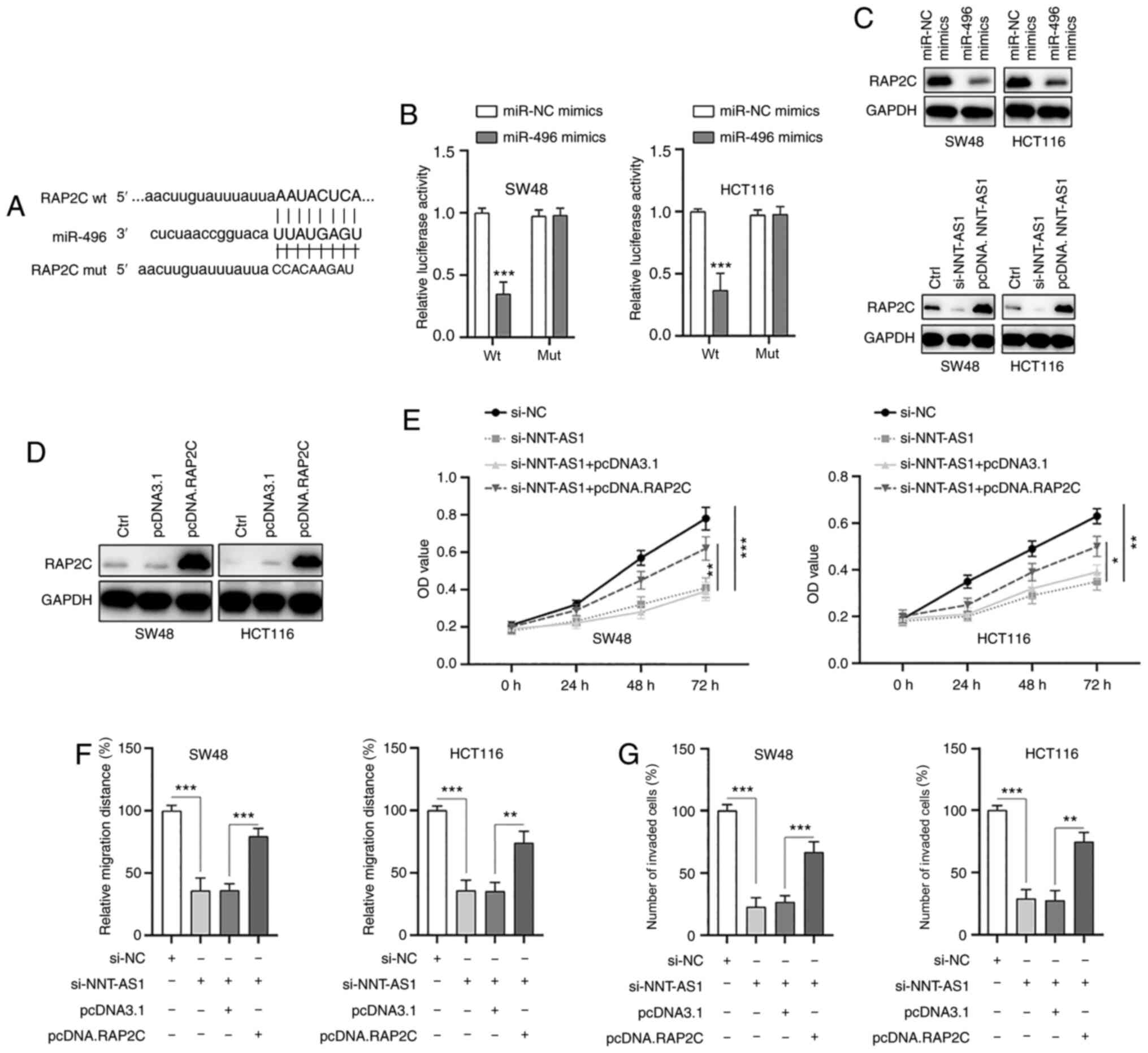 | Figure 7.RAP2C is a direct target of miR-496.
(A) The constructed luciferase reporter plasmids containing the
predicted wt or mut miR-496 binding sites on RAP2C. (B) A
luciferase reporter assay was performed to measure the luciferase
activity in CRC cells after co-transfection of miRNA mimics and
RAP2C wt or mut. (C) CRC cells were transfected with miR-496
mimics, si-NNT-ASI or pcDNA.NNT-ASI and the levels of the indicated
proteins were measured by western blotting. (D) RAP2C was
overexpressed in CRC cells by transfection with pcDNA.RAP2C. (E)
CRC cells were transfected as indicated and cell viability was
measured by an MTT assay. CRC cells were transfected as indicated
and (F) migration and (G) invasion were measured. Data are
presented as the mean ± standard deviation. *P<0.05, **P<0.01
and ***P<0.001 vs. miR-NC mimics or as indicated. All
experiments were conducted at least three times. RAP2C, Ras-related
protein Rap-2c; miR, microRNA; wt, wild-type; mut, mutant; NC,
negative control; si-, small interfering RNA; pcDNA, overexpression
vector; CRC, colorectal cancer; NNT-AS1, lncRNA nicotinamide
nucleotide transhydrogenase antisense RNA 1. |
Discussion
CRC is one of the most common malignancies
worldwide, with considerable mortality rates. A lack of early
diagnosis results in limited effective therapeutic options and poor
clinical outcomes (3). Therefore,
the identification of novel sensitive and specific biomarkers for
CRC is urgently needed. Accumulating evidence has indicated that
due to different expression patterns in various cancers, lncRNAs
can be applied as effective diagnostic and prognostic biomarkers
(6–8). In addition, the detection of
circulating lncRNAs can be convenient and non-invasive, making
lncRNAs an ideal tool for tumor diagnosis. For instance, it was
reported that lncRNA AFAP1-AS1 is upregulated in patients with
non-small cell lung cancer and demonstrates a higher sensitivity
and specificity when compared with healthy controls (27). Wang et al (28) identified that circulating lncRNA
CCAT2 is significantly upregulated in the serum of patients with
CRC and could be applied as a potential predictor in CRC. The
present study focused on NNT-AS1 to study the potential clinical
value of circulating NNT-AS1 in CRC.
The lncRNA NNT-AS1, located on chromosome 5p12, has
been identified to be differentially expressed in different types
of cancers. For example, NNT-AS1 has been identified to contribute
to the carcinogenesis of hepatocellular cancer cells through the
regulation of miR-363 (29). High
expression of NNT-AS1 also leads to the proliferation and invasion
of cervical cancer via activation of the Wnt/β-catenin signaling
pathway (8). By contrast, Huang
et al (30) reported that
NNT-AS1 is downregulated in ovarian cancer tissues and cells. These
discrepancies indicate the complex role of NNT-AS1 in various
cancers and further investigation is necessary to reveal the
biological function of NNT-AS1. The present study identified that
NNT-AS1 was significantly upregulated in CRC tissues compared with
adjacent normal tissues. Notably, the higher expression levels of
NNT-AS1 were associated with advanced TNM stage, increased local
invasion and positive lymph node metastasis. The findings of the
present study are similar to those of a previous study that
demonstrated that NNT-AS1 is upregulated in CRC cells and serves an
essential role in proliferation, migration and invasion (12). Accumulating evidence has
demonstrated that circulating lncRNAs, such as H19, ATB, CCAT1,
HOTAIR and GAS5, can be stably detected and used as non-invasive
biomarkers for various tumors (31–33).
However, to the best of the authors' knowledge, the present study
was the first to investigate the levels of NNT-AS1 in the serum of
patients with CRC. As a result, it was identified that serum
NNT-AS1 levels were also significantly higher in patients with CRC
compared with healthy controls. However, no significant association
was identified between serum NNT-AS1 levels and other
clinicopathological features. The present study also investigated
the serum NNT-AS1 expression levels in paired preoperative and
postoperative samples and identified that the levels of NNT-AS1 in
serum were significantly decreased in the postoperative
samples.
A number of studies have reported that lncRNAs
stably exist in circulating blood and are protected by exosomes
(34,35). Exosomes are type of membrane vesicle
with a diameter of 40–150 nm and can be purified by sucrose
gradients at a density range of 1.13-1.19 g/ml (35). Exosomes also possess a bilayer lipid
membrane that is structurally similar to the cell membrane
(36). Exosomes have become a focus
of investigation due to their potential as a novel approach for the
diagnosis of cancers (37). ncRNAs
(including miRNAs and lncRNAs) and proteins from exosomes have been
proven to be useful as biomarkers (37). Thus far, only a few lncRNAs have
been identified to be contained in exosomes and possess biomarker
functions in CRC (28,38). To further confirm the vital function
of NNT-AS1 in the diagnosis of CRC, the expression of NNT-AS1 was
assayed in exosomes and it was identified that NNT-AS1 was also
upregulated in CRC exosomes. The expression of NNT-AS1 in serum
samples and in exosomes isolated from the same volume of serum was
also compared. No significant differences were observed; therefore,
NNT-AS1 might exist in exosomes, which may provide protection to
increase the stability of NNT-AS1 in serum.
The functional roles of NNT-AS1 in CRC cells were
also examined. The data demonstrated that silencing NNT-AS1
inhibited the proliferation, migration and invasion of CRC cells.
These findings are in line with previous studies showing that
NNT-AS1 promotes the proliferation and metastasis of various
cancers, such as cervical cancer, CRC and liver cancer (8,12,29).
Increasing evidence has confirmed that lncRNAs can bind and
negatively regulate the function of miRNAs (5–7). The
present study revealed that NNT-AS1 acted as a sponge of miR-496,
which acted as a tumor suppressor in CRC cells. Previous studies
have suggested that miR-496 functions as a tumor suppressor in
various cancers, such as CRC, lung cancer and osteosarcoma
(18,39,40). A
recent study identified that miR-496 acted as an oncogene and
promoted malignancy in CRC cells (41). This discrepancy might be due to
different cancer types and further investigation is needed to
further verify the role of miR-496 in more cancer types. The
present study also revealed that RAP2C might be a direct target of
miR-496. RAP2C, a member of the Ras family, has been identified to
participate in the tumorigenesis of various cancers. For instance,
RAP2C acts as an oncogene in human osteosarcoma cells (20). Inhibition of RAP2C suppresses the
proliferation and promotes the apoptosis of breast cancer cells
(21). However, there is little
information concerning the role of RAP2C in CRC. Overexpression of
RAP2C partially abrogated the effects of NNT-AS1 knockdown and
rescued the proliferation, migration and invasion of CRC cells. The
data from the present study suggested that RAP2C acted as an
oncogene in CRC cells and further investigation is needed to
confirm this.
In summary, the data from the present study
suggested that NNT-AS1 expression is significantly upregulated in
tissue and serum samples of patients with CRC. Circulating NNT-AS1
may be protected by exosomes and could be used as a potential
predictor in CRC. It was also identified that NNT-AS1 regulated the
proliferation, migration and invasion of CRC cells via the
miR-496/RAP2C axis. The results from the present study identified
novel pathways regulating CRC progression, which may facilitate the
identification of novel therapeutic targets for CRC.
Supplementary Material
Supporting Data
Acknowledgements
Not applicable.
Funding
The present study was supported by Medical
Scientific Research Identification of Zhejiang Province, China
(grant. no. 2019KY595), Provincial Natural Fund of Zhejiang
Province (grant. no. LGC20H160002) and Ningbo Clinical Research
Center for Digestive System Tumors (grant. no. 2019A21003).
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
HY and JH performed the experiments. ZY repeated the
experiments (data not shown), validated the data and conducted the
statistical analysis. SC analyzed and interpreted the data. ZY and
SC confirm the authenticity of all the raw data. YC conceived and
designed the study, and drafted and revised the manuscript. All
authors read and approved the final manuscript.
Ethics approval and consent to
participate
The present study was approved by the Ethical
Committee of Hwamei Hospital, University of Chinese Academy of
Sciences (Ningbo, China) and all procedures were conducted
according to the Declaration of Helsinki. Written informed consent
was obtained from each patient.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Siegel RL, Miller KD, Fedewa SA, Ahnen DJ,
Meester RGS, Barzi A and Jemal A: Colorectal cancer statistics,
2017. CA Cancer J Clin. 67:177–193. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Brenner H, Kloor M and Pox CP: Colorectal
cancer. Lancet. 383:1490–1502. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
The Lancet: Toward better control of
colorectal cancer. Lancet. 383:14372014. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Peng WX, Koirala P and Mo YY:
LncRNA-mediated regulation of cell signaling in cancer. Oncogene.
36:5661–5667. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Wang KC and Chang HY: Molecular mechanisms
of long noncoding RNAs. Mol Cell. 43:904–914. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Han D, Wang M, Ma N, Xu Y, Jiang Y and Gao
X: Long noncoding RNAs: Novel players in colorectal cancer. Cancer
Lett. 361:13–21. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Schmitt AM and Chang HY: Long noncoding
RNAs in cancer pathways. Cancer Cell. 29:452–463. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Hua F, Liu S, Zhu L, Ma N, Jiang S and
Yang J: Highly expressed long non-coding RNA NNT-AS1 promotes cell
proliferation and invasion through Wnt/β-catenin signaling pathway
in cervical cancer. Biomed Pharmacother. 92:1128–1134. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Huang Q, Wang S, Li X, Yang F, Feng C,
Zhong K, Qiu M and Wang J: Circular RNA ATXN7 is upregulated in
non-small cell lung cancer and promotes disease progression. Oncol
Lett. 17:4803–4810. 2019.PubMed/NCBI
|
|
10
|
Wang L, Ma H, Kong W, Liu B and Zhang X:
Up-regulated circular RNA VANGL1 contributes to progression of
non-small cell lung cancer through inhibition of miR-195 and
activation of Bcl-2. Biosci Rep. 39:BSR201824332019. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
He W, Zhang Y and Xia S: LncRNA NNT-AS1
promotes non-small cell lung cancer progression through regulating
miR-22-3p/YAP1 axis. Thorac Cancer. 11:549–560. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Wang Q, Yang L, Hu X, Jiang Y, Hu Y, Liu
Z, Liu J, Wen T, Ma Y, An G and Feng G: Upregulated NNT-AS1, a long
noncoding RNA, contributes to proliferation and migration of
colorectal cancer cells in vitro and in vivo. Oncotarget.
8:3441–3453. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Sun T, Kalionis B, Lv G, Xia S and Gao W:
Role of exosomal noncoding RNAs in lung carcinogenesis. Biomed Res
Int. 2015:1258072015. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Gallo A, Tandon M, Alevizos I and Illei
GG: The majority of microRNAs detectable in serum and saliva is
concentrated in exosomes. PLoS One. 7:e306792012. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Luga V, Zhang L, Viloria-Petit AM,
Ogunjimi AA, Inanlou MR, Chiu E, Buchanan M, Hosein AN, Basik M and
Wrana JL: Exosomes mediate stromal mobilization of autocrine
Wnt-PCP signaling in breast cancer cell migration. Cell.
151:1542–1556. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Hewson C and Morris KV: Form and function
of exosome-associated long non-coding RNAs in cancer. Curr Top
Microbiol Immunol. 394:41–56. 2016.PubMed/NCBI
|
|
17
|
Lee YS and Dutta A: MicroRNAs in cancer.
Annu Rev Pathol. 4:199–227. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Ma R, Zhu P, Liu S, Gao B and Wang W:
MiR-496 suppress tumorigenesis via targeting BDNF-mediated PI3K/Akt
signaling pathway in non-small cell lung cancer. Biochem Biophys
Res Commun. 518:273–277. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Alvarado S, Wyglinski J, Suderman M,
Andrews SA and Szyf M: Methylated DNA binding domain protein 2
(MBD2) coordinately silences gene expression through activation of
the microRNA hsa-mir-496 promoter in breast cancer cell line. PLoS
One. 8:e740092013. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Wu J, Du W, Wang X, Wei L, Pan Y, Wu X,
Zhang J and Pei D: Ras-related protein Rap2c promotes the migration
and invasion of human osteosarcoma cells. Oncol Lett. 15:5352–5358.
2018.PubMed/NCBI
|
|
21
|
Zhu X, Qiu J, Zhang T, Yang Y, Guo S, Li
T, Jiang K, Zahoor A, Deng G and Qiu C: MicroRNA-188-5p promotes
apoptosis and inhibits cell proliferation of breast cancer cells
via the MAPK signaling pathway by targeting Rap2c. J Cell Physiol.
235:2389–2402. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Wang Z, Huang C, Zhang A, Lu C and Liu L:
Overexpression of circRNA_100290 promotes the progression of
laryngeal squamous cell carcinoma through the miR-136-5p/RAP2C
axis. Biomed Pharmacother. 125:1098742020. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Yu R, Yu BX, Chen JF, Lv XY, Yan ZJ, Cheng
Y and Ma Q: Anti-tumor effects of Atractylenolide I on bladder
cancer cells. J Exp Clin Cancer Res. 35:402016. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Muschelli J: ROC and AUC with a binary
predictor: Potentially a misleading metric. J Classif. 37:696–708.
2002. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Li C, Zhang S, Qiu T, Wang Y, Ricketts DM
and Qi C: Upregulation of long non-coding RNA NNT-AS1 promotes
osteosarcoma progression by inhibiting the tumor suppressive
miR-320a. Cancer Biol Ther. 20:413–422. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Li T, Ding ZL, Zheng YL and Wang W:
MiR-484 promotes non-small-cell lung cancer (NSCLC) progression
through inhibiting Apaf-1 associated with the suppression of
apoptosis. Biomed Pharmacother. 96:153–164. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Wang L, Duan W, Yan S, Xie Y and Wang C:
Circulating long non-coding RNA colon cancer-associated transcript
2 protected by exosome as a potential biomarker for colorectal
cancer. Biomed Pharmacother. 113:1087582019. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Lu YB, Jiang Q, Yang MY, Zhou JX and Zhang
Q: Long noncoding RNA NNT-AS1 promotes hepatocellular carcinoma
progression and metastasis through miR-363/CDK6 axis. Oncotarget.
8:88804–88814. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Huang Y, Shi J and Xu Y: Long non-coding
RNA NNT-AS1 contributes to cell proliferation, metastasis and
apoptosis in human ovarian cancer. Oncol Lett. 15:9264–9270.
2018.PubMed/NCBI
|
|
31
|
Abedini P, Fattahi A, Agah S, Talebi A,
Beygi AH, Amini SM, Mirzaei A and Akbari A: Expression analysis of
circulating plasma long noncoding RNAs in colorectal cancer: The
relevance of lncRNAs ATB and CCAT1 as potential clinical hallmarks.
J Cell Physiol. 234:22028–22033. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Zhang R, Xia Y, Wang Z, Zheng J, Chen Y,
Li X, Wang Y and Ming H: Serum long non coding RNA MALAT-1
protected by exosomes is up-regulated and promotes cell
proliferation and migration in non-small cell lung cancer. Biochem
Biophys Res Commun. 490:406–414. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Shen J, Hodges TR, Song R, Gong Y, Calin
GA, Heimberger AB and Zhao H: Serum HOTAIR and GAS5 levels as
predictors of survival in patients with glioblastoma. Mol Carcinog.
57:137–141. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Arita T, Ichikawa D, Konishi H, Komatsu S,
Shiozaki A, Shoda K, Kawaguchi T, Hirajima S, Nagata H, Kubota T,
et al: Circulating long non-coding RNAs in plasma of patients with
gastric cancer. Anticancer Res. 33:3185–3193. 2013.PubMed/NCBI
|
|
35
|
Niu L, Song X, Wang N, Xue L, Song X and
Xie L: Tumor-derived exosomal proteins as diagnostic biomarkers in
non-small cell lung cancer. Cancer Sci. 110:433–442. 2019.
View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Pant S, Hilton H and Burczynski ME: The
multifaceted exosome: Biogenesis, role in normal and aberrant
cellular function and frontiers for pharmacological and biomarker
opportunities. Biochem Pharmacol. 83:1484–1494. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Zhou R, Chen KK, Zhang J, Xiao B, Huang Z,
Ju C, Sun J, Zhang F, Lv XB and Huang G: The decade of exosomal
long RNA species: An emerging cancer antagonist. Mol Cancer.
17:752018. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Liu T, Zhang X, Gao S, Jing F, Yang Y, Du
L, Zheng G, Li P, Li C and Wang C: Exosomal long noncoding RNA
CRNDE-h as a novel serum-based biomarker for diagnosis and
prognosis of colorectal cancer. Oncotarget. 7:85551–85563. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Qi NN, Tian S, Li X, Wang FL and Liu B:
Up-regulation of microRNA-496 suppresses proliferation, invasion,
migration and in vivo tumorigenicity of human osteosarcoma cells by
targeting eIF4E. Biochimie. 163:1–11. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Ye J, Xie W, Zuo Y, Jing G and Tong J:
MicroRNA-496 suppresses tumor cell proliferation by targeting BDNF
in osteosarcoma. Exp Ther Med. 19:1425–1431. 2020.PubMed/NCBI
|
|
41
|
Wang H, Yan B, Zhang P, Liu S, Li Q, Yang
J, Yang F and Chen E: MiR-496 promotes migration and
epithelial-mesenchymal transition by targeting RASSF6 in colorectal
cancer. J Cell Physiol. 235:1469–1479. 2020. View Article : Google Scholar : PubMed/NCBI
|