Introduction
Ischemia/reperfusion (I/R) injury refers to the
destruction of the normal structure and physiological function of
the tissues and organs after the interruption and restoration of
the blood supply (1). I/R injury
often occurs when tissues and organs are traumatized, such as after
thrombosis, frostbite burns, organ transplantation and surgery
(2). Hepatic I/R injury (HIRI) is
also a common complication after liver tissue injury and includes
two stages of ischemia injury and reperfusion injury, and is
associated with calcium overload, oxidative stress, cell apoptosis
and metabolic acidosis (3,4). However, there are currently no ideal
treatments or prevention interventions that can effectively avoid
HIRI.
Accumulating evidence has suggested that stem cells
secrete a variety of cytokines, improve the local microenvironment,
regulate the process of cell apoptosis and exert anti-inflammatory
and anti-apoptotic effects, as well as promote the endogenous
repair of tissues in the injured area (5–7). With
the development of stem cells in the field of disease treatment,
bone marrow-derived mesenchymal stem cells (BM-MSCs) have been
attracting increased attention due to their wide-ranging sources,
weak immunogenicity and multidirectional differentiation across
germ layers (8,9). After continuous subculturing and
cryopreservation, BM-MSCs maintain their multidirectional
differentiation ability, as well as their normal karyotype and
telomerase activity, and can be induced to differentiate into
hepatocytes, neuronal cells, cartilage cells and adult cells under
specific conditions in vitro (10,11).
Moreover, BM-MSCs can attenuate injury of the brain, heart, kidney,
liver and other organs, which activates strong chemical signals
from damaged tissues, via differentiating into a variety of cells,
secreting cytokines and chemokines, and through cell fusion
(12).
A number of studies have reported that MSCs can
alleviate liver histological damage, promote hepatocyte
proliferation and accelerate liver tissue recovery (13,14).
Accumulating evidence has indicated that BM-MSC-derived
hepatocyte-like exosomes reduce HIRI by enhancing autophagy
(13). In a rat HIRI model, MSCs
improve PTEN induced kinase 1-dependent mitochondrial-mediated
apoptosis of hepatocytes via AMP-activated protein kinase α subunit
activation (15). Other studies
have further confirmed that allogeneic BM-MSCs can reduce HIRI by
inhibiting oxidative stress and apoptosis (16). These findings indicate that BM-MSCs
serve an important role in HIRI. Therefore, BM-MSCs may have
potential therapeutic prospects for the treatment of HIRI.
It is worth noting that BM-MSCs can also be used as
vectors to deliver protective genes by overexpressing the
transfected genes in damaged parts. This not only improves the
therapeutic effect, but also promotes local tissue repair (17). Superoxide dismutase 2 (SOD2) gene
transfection of BM-MSCs inhibits inflammation, and it has been
found that the transfected gene can be stably expressed in the body
and improve glucose tolerance (18). Furthermore, upregulation of SOD2 can
promote the proliferation of human umbilical cord blood-derived
MSCs, and improve the oxidative stress and apoptosis of bone tissue
(19). These findings suggest that
BM-MSCs can be used as cell therapy, and that they are also optimal
gene carriers.
Based on the aforementioned research findings, the
present study aimed to establish an animal model of HIRI that
closely simulated hepatic transplantation injury to investigate the
protective role of SOD2-overexpressing BM-MSCs and the underlying
molecular mechanisms during HIRI.
Materials and methods
Animals
All animal procedures were conducted in accordance
with the International Guidelines for the Care and Use of
Laboratory Animals and local ethics committee approval (20), and were also approved by the
Laboratory Animal Center, The Second Xiangya Hospital of Central
South University (Changsha, China). A total of 80 male Wistar rats
(weight, 200–220 g; age, 8–10 weeks) were used for this study. All
rats were kept in a temperature and humidity-controlled environment
with a 12-h light/dark cycle at 22–25°C, with 50–65% humidity and
free access to food and water.
Isolation of BM-MSCs
BM-MSCs were obtained using density centrifugation
as previously described (18).
Briefly, MSCs were flushed from the femurs and tibias of male
4-week-old Wistar rats, and were cultured with MEM (Sigma-Aldrich;
Merck KGaA) supplemented with 10% FBS (cat. no. F8687;
Sigma-Aldrich; Merck KGaA), 100 U/ml penicillin and 100 mg/l
streptomycin. Cultures were maintained at 37°C in a humidified
atmosphere containing 5% CO2. Non-adherent hematopoietic
cells were removed from the adherent BM-MSCs. The culture medium
was changed every 3 days. The cells were subcultured when they
reached 70–80% confluence. Third-passage BM-MSCs were used in all
the experiments.
Characterization of MSCs
When the cultured cells reached 90% confluence,
adherent cells were trypsinized and passaged. Cells underwent five
passages before use in subsequent experiments. BM-MSCs were
analyzed via antibody staining using CD90 (cat. no. SAB4700719;
Sigma-Aldrich; Merck KGaA), CD105 (cat. no. MABT117; Sigma-Aldrich;
Merck KGaA), CD34 (cat.no. RAB1334; Sigma-Aldrich; Merck KGaA) and
CD45 (cat. no. APREST79682; Sigma-Aldrich; Merck KGaA). MSC marker
antibody staining was performed using a FACSCanto II flow cytometer
(BD Biosciences).
Adenovirus production and
transduction
Empty adenovirus vectors (Ad.null) and vectors
encoding SOD2 (Ad.SOD2) were purchased from Shanghai Liangtai
Biotechnology Company, and transductions were performed as
previously described (21). To
yield the adenovirus, the linearized construct DNA was transfected
into 293 cells (American Type Culture Collection) using
Lipofectamine® 2000 (Invitrogen; Thermo Fisher
Scientific, Inc.), according to the manufacturer's instructions.
After 24 h, the cells were supplied with fresh medium, and the
incubation was continued for an additional 5 days. The virus was
released from the cells by freezing and thawing for three
consecutive cycles. After the third freeze-thaw cycle, the cells
were briefly centrifuged at 4,000 × g to pellet the debris at 4°C
for 10 min, and the lysate was collected in sterile centrifuge
tubes and stored at −20°C for subsequent use. For adenovirus
transduction, the MSCs were plated into 6-well plates, and the next
day, the adenovirus was added into 2 ml serum-free DMEM at a MOI of
10. The plate was centrifuged at 220 × g for 90 min at 37°C. Then,
the cells were incubated in a 5% CO2 incubator at 37°C
for an additional 4 h. Next, the medium was removed, and fresh
complete growth medium (cat. no. 12558011; Gibco; Thermo Fisher
Scientific, Inc.) was added. The cells were incubated for another
24 h at 37°C prior to analysis.
Animal model of HIRI
Rats were divided into the following groups (n=8 per
group): Sham group, rats were subjected to laparotomy only; I/R
group, rats were induced with I/R and received PBS; +MSCs, rats
were induced with I/R and then transplanted with MSCs; and
+SOD2-MSCs, rats were induced with I/R and then transplanted with
SOD2-overexpressing MSCs. HIRI was induced as previously described
(17). All surgical procedures were
performed under anesthesia with pentobarbital sodium (60 mg/kg). A
midline laparotomy was performed to expose the portal circulation,
and a microaneurysm clamp was placed on the hepatic artery and
portal vein to block the blood supply. This method resulted in 70%
of segmental liver ischemia and prevented mesenteric veins. After
60 min, the clamp was removed, and 1×106 PKH26-labeled
MSCs were immediately resuspended in 200 µl PBS with a 30-gauge
needle. Sham-operated rats only underwent laparotomy. After the
operation, a 4/0 silk suture was used. After the animals were fully
awake, they were provided with free access to food and water.
During the entire procedure, the core body temperature of each rat
was continuously monitored and maintained at 37.0±0.4°C with a
heating lamp. All animals were anesthetized by intraperitoneal
injection of sodium pentobarbital (30 mg/kg) until the animals lost
consciousness, and then sacrificed by exsanguination.
Assessment of liver functions
To assess the severity of HIRI, the levels of serum
aspartate aminotransferase (AST) and alanine aminotransferase (ALT)
were determined. Rats were anesthetized with pentobarbital sodium
(60 mg/kg), and 2 ml blood was collected from the inferior vena
cava with a 20-gauge needle, which was then placed in a microtainer
tube with serum separator (Eppendorf), and centrifuged at 4°C at
3,000 × g for 12 min. AST (cat. no. C010-2-1; Nanjing Jiancheng
Bioengineering Institute) and ALT (cat. no. C009-2-1; Nanjing
Jiancheng Bioengineering Institute) levels were measured using
commercial kits.
Hematoxylin and eosin (H&E)
staining
Liver tissues were collected, fixed in 4%
paraformaldehyde for 24 h at room temperature and dehydrated until
transparent. The tissues were embedded in paraffin, and cut into
5-µm sections. Sections were stained with H&E at room
temperature and observed in three random areas under a light
microscope (Zeiss GmbH) connected to a digital camera.
The severity of hepatic injury was evaluated in
accordance with the modified Suzuki classification (22). Scores for the corresponding
indicators of liver severity were determined as follows: None, 0;
minimal, 1; moderate, 2; and severe, 3. For each rat, three liver
sections were examined and three randomly selected high-power
fields (magnification, ×200) were analyzed in each section. The
mean score for each animal was then determined by a summation of
all scores.
Determination of SOD, glutathione
peroxidase (GSH-Px) and malondialdehyde (MDA) in the liver
Liver tissues were collected and homogenized in
ice-cold 0.9% saline. Following centrifugation at 3,000 × g for 10
min at 4°C, the supernatant was collected and the activities of SOD
(cat. no. 19160; Sigma-Aldrich; Merck KGaA), GSH-Px (cat. no.
CS0260; Sigma-Aldrich; Merck KGaA) and MDA (cat. no. MAK085;
Sigma-Aldrich; Merck KGaA) content were determined using commercial
kits (Nanjing Jiancheng Bioengineering Institute).
Measurement of ROS
The total ROS level was measured using a ROS assay
kit (cat. no. MAK143; Sigma-Aldrich; Merck KGaA), according to the
manufacturer's protocol. Briefly, intracellular ROS levels were
determined by measuring the oxidative conversion of cell permeable
2V, 7V-dichlorofluorescein diacetate (DCFH-DA) to fluorescent
dichlorofluorescein (DCF) using a fluorospectrophotometer (Thermo
Fisher Scientific, Inc.) at an excitation wavelength of 488 nm and
an emission wavelength of 535 nm.
Detection of apoptotic cells in liver
tissues
A TUNEL kit was used to detect the apoptotic cells.
Liver sections (thickness, 5 µm) were excised and fixed with 4%
paraformaldehyde in PBS at room temperature for 24 h. Fixed tissues
were embedded in paraffin and stained using a TUNEL kit (cat. no.
11684795910; Roche Diagnostics), according to the manufacturer's
protocol, and six sections were analyzed for each rat. The numbers
of apoptotic cells and total hepatic cells in each section were
counted in three randomly selected fields (magnification, ×400).
The apoptosis index (AI) was expressed as the mean percentage of
apoptotic cells within the total number of hepatic cells for each
animal.
Western blotting
A total of 100 mg liver tissue was homogenized with
lysate (cat.no. P0013; Beyotime Institute of Biotechnology), and
the supernatant was centrifuged at 4,000 × g at 4°C for 10 min.
Protein quantification was performed using a BCA protein assay
(Abcam) and protein samples (20 µg) were collected and subjected to
SDS-PAGE (10% separation gel and 6% concentration gel). Proteins in
the gel were subsequently transferred to PVDF membranes and blocked
with 5% skimmed milk in TBS-0.1% Tween-20 at 37°C for 1 h. The
membranes were then incubated with the following primary
antibodies: Rabbit anti-Bcl-2 (1:1,000; cat. no. SAB4500003;
Sigma-Aldrich; Merck KGaA), rabbit anti-Bax (1:1,000; cat. no.
SAB4502546; Sigma-Aldrich; Merck KGaA) and rabbit anti-caspase-3
(1:1,000; cat. no. C8487; Sigma-Aldrich; Merck KGaA) in blocking
solution at 4°C overnight. Membranes were then washed and incubated
for 5 min at room temperature with HRP-conjugated anti-mouse
(1:5,000; cat. no. AP160P; Sigma-Aldrich; Merck KGaA) or
anti-rabbit IgG secondary antibodies (1:2,000; cat. no. 31402;
Invitrogen; Thermo Fisher Scientific, Inc.). The bound secondary
antibodies were analyzed with an Odyssey Infrared Imaging system
(LI-COR Biosciences), and proteins were normalized to β-actin
(1:5,000; cat. no. SAB3500350; Sigma-Aldrich; Merck KGaA).
Densitometric analysis was performed using ImageJ version 2
software (National Institutes of Health).
Reverse transcription-quantitative
(RT-q)PCR
Total RNA was extracted using a RNeasy Mini kit
(Qiagen, Inc.) from the MSCs after adenovirus transduction and
purified with 75% ethanol. RNA concentration was determined by
spectrophotometry. The purified total RNA (200 ng/sample) was
reverse transcribed into cDNA using a transcription kit (cat. no.
RR037A; Takara Bio, Inc.). qPCR reactions were performed in
triplicate using a SYBR® Green Master Mix (Bio-Rad
Laboratories, Inc.) and run on a LightCycler 480 system (Roche
Diagnostics GmbH). The following thermocycling conditions were
used: Initial denaturation at 95°C for 30 sec; followed by 40
cycles of denaturation at 95°C for 5 sec, and annellation and
extension at 60°C for 31 sec. The following primers were used in
the current study: SOD2 forward, 5′-ACGTACTAGACGCGCAATT-3′ and
reverse, 5′-ACTTGTTAGAGTTGCGGTGG-3′; and GAPDH forward,
5′-CAGTTACTTCCCCAGCAA-3′ and reverse, 5′-CACGACTCATACAGCACCT-3′.
The relative gene expression was quantified using the
2−∆∆Cq method (23).
Statistical analysis
SPSS 21.0 (IBM Corp.) was used for statistical
analyses. Data are presented as the mean ± SD of experiments
repeated in triplicate. For multiple comparisons, data were
analyzed using one-way ANOVA followed by a Tukey's post hoc test.
Analysis between two groups was performed using an unpaired
Student's t-test. P<0.05 was considered to indicate a
statistically significant difference.
Results
Expression level of SOD2 in BM-MSCs
with adenovirus vector transduction
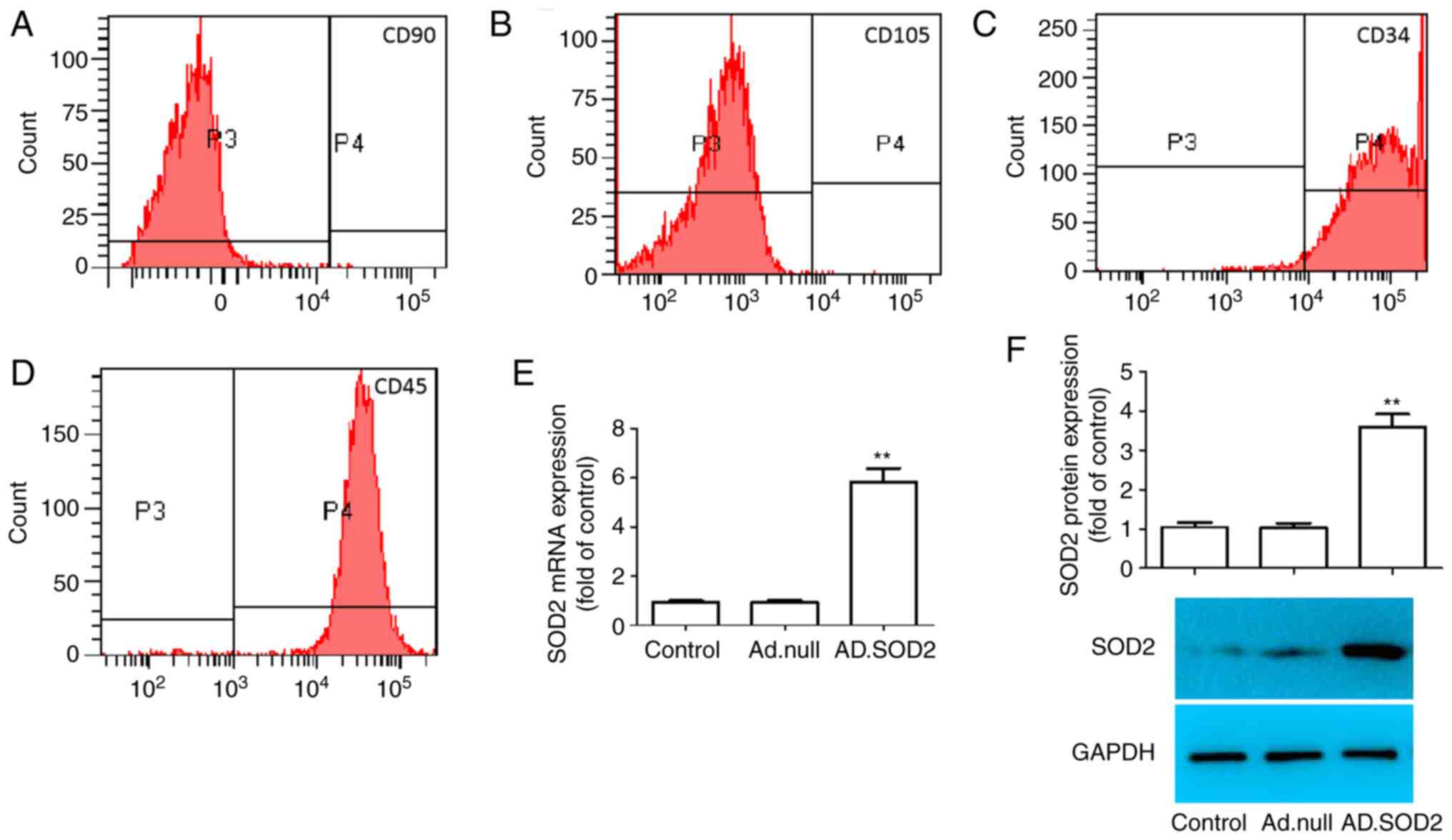
To verify whether the collected cells were BM-MSCs,
MSC-specific cell surface markers were detected via staining. As
shown in Fig. 1A-D, BM-MSCs were
positive for CD90 and CD105, which are specific MSC surface markers
(24). Additionally, these cells
were negative for CD34 and CD45, which are non-MSC markers. The
findings suggested that these BM-MSCs were typical MSCs, and were
used for subsequent experiments.
To verify SOD2 overexpression in BM-MSCs, Ad.SOD2
was transduced into BM-MSCs. After infection for 24 h, the mRNA and
protein expression levels of SOD2 were assessed. The results
demonstrated that SOD2 mRNA and protein expression levels were
significantly increased (Fig. 1E and
F) in Ad.SOD2-transduced BM-MSCs compared with mock group or
Ad.null-transduced BM-MSCs (**P<0.01).
Effects of BM-MSCs overexpressing SOD2
on AST and ALT levels after HIRI
Elevated AST and ALT levels are important signs of
severe liver injury (25). To
determine the degree of I/R-induced hepatic injury in the liver
tissues of the rat receiving BM-MSCs or SOD2-overexpressing
BM-MSCs, ALT and AST levels were assessed after I/R and cell
transplantation. Compared with the sham group, markers of liver
damage, including ALT and AST, were significantly increased in I/R
model rats (**P<0.01; Fig. 2A and
B). However, there were significantly decreased levels of ALT
and AST in the I/R + MSCs and I/R + SOD2-MSCs groups compared with
the I/R group (#P<0.05). These findings suggested
that the transplantation of SOD2-overexpressing BM-MSCs improved
HIRI.
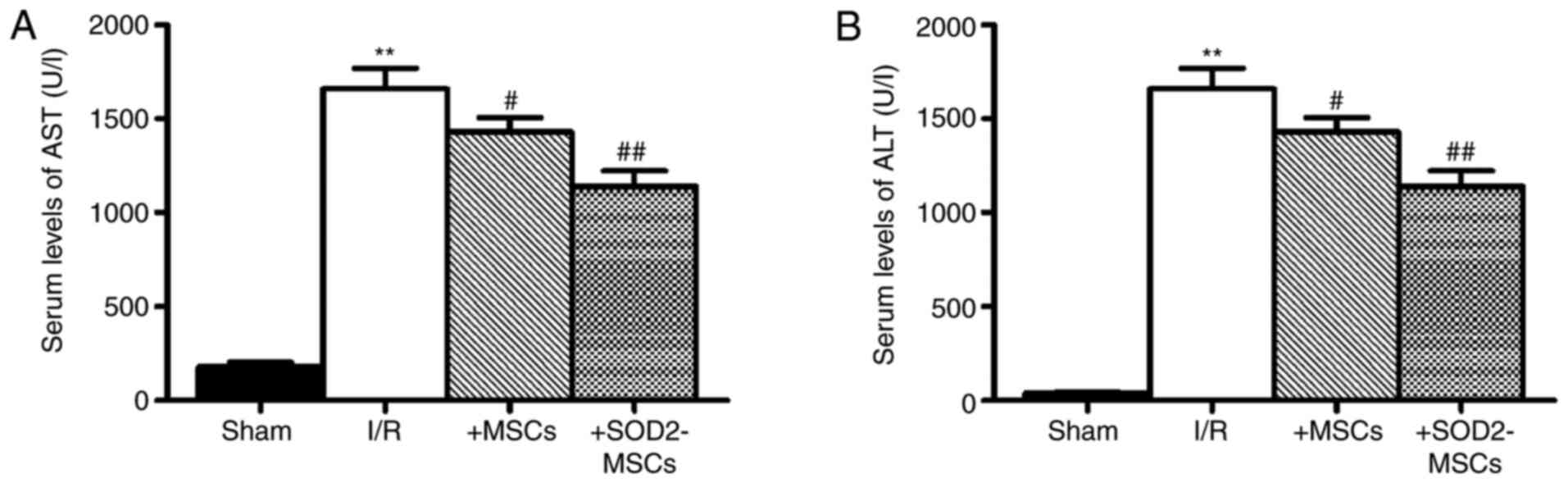 | Figure 2.Effects of SOD2-overexpressing MSC
transplantation on AST and ALT levels in the serum of rats with
hepatic I/R injury. (A) AST and (B) ALT levels in the serum. Blood
samples were collected after I/R and cell transplantation for ALT
and AST analysis. Sham group, rats were subjected to laparotomy
only; I/R group, rats were induced with I/R and received PBS;
+MSCs, rats were induced with I/R and then transplanted with MSCs;
and +SOD2-MSCs, rats were induced with I/R and then transplanted
with SOD2-overexpressing MSCs. n=8 rats in each experimental group.
Data are expressed as the mean ± SD. **P<0.01 vs. sham;
#P<0.05, ##P<0.01 vs. I/R. I/R,
ischemia/reperfusion; SOD2, superoxide dismutase 2; MSCs,
mesenchymal stem cells; ALT, alanine aminotransferase; AST,
aspartate aminotransferase. |
BM-MSCs overexpressing SOD2 improve
the histopathological changes of HIRI
To further confirm the role of SOD2 overexpression
on HIRI in rats, the histopathology of livers harvested after I/R
induction and cell transplantation were examined. As shown in
Fig. 3A-D, the pathological
findings identified sinusoidal congestion, cytoplasmic
vacuolization and necrosis in the I/R group, which are indicative
of severe damage. Moreover, the results of the Suzuki scores
indicated worse histopathology in the I/R group compared with the
sham group (**P<0.01), whereas the scores indicated improved
histopathology in the I/R + MSCs and I/R + SOD2-MSCs groups
compared with the I/R group (#P<0.05; Fig. 3E).
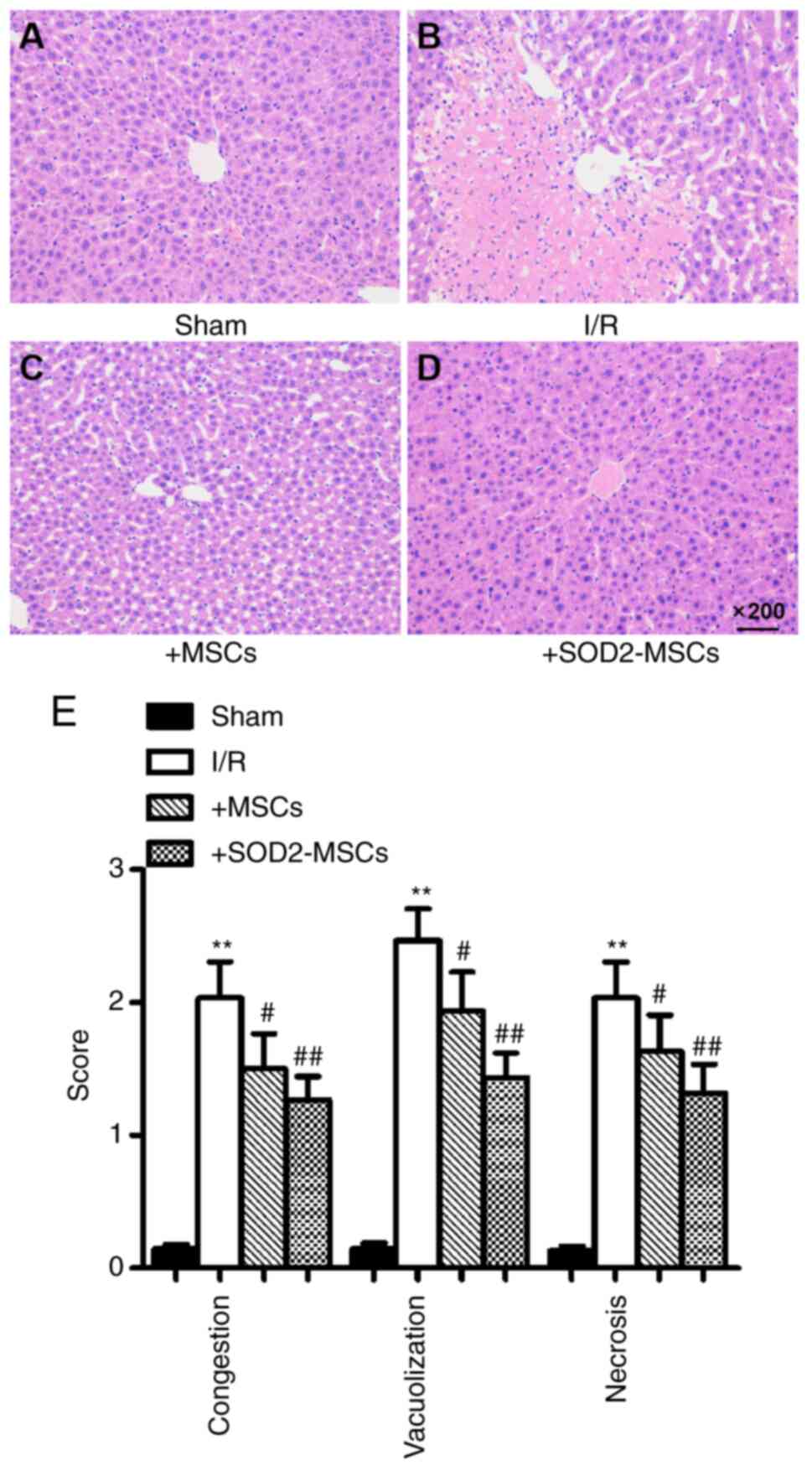 | Figure 3.Histopathological analysis of liver
tissues in rats. Liver tissue sections were stained with
hematoxylin and eosin and scored according to the Suzuki Score
System. Represented micrographs of liver tissue staining.
Representative micrographs of livers from the (A) sham, (B) I/R,
(C) +MSCs and (D) +SOD2-MSCs groups. (E) Suzuki scores from each
group. Sham group, rats were subjected to laparotomy only; I/R
group, rats were induced with I/R and received PBS; +MSCs, rats
were induced with I/R and then transplanted with MSCs; and
+SOD2-MSCs, rats were induced with I/R and then transplanted with
SOD2-overexpressing MSCs. n=8 rats in each experimental group. Data
are expressed as the mean ± SD. **P<0.01 vs. sham;
#P<0.05, ##P<0.01 vs. I/R. I/R,
ischemia/reperfusion; SOD2, superoxide dismutase 2; MSCs,
mesenchymal stem cells. |
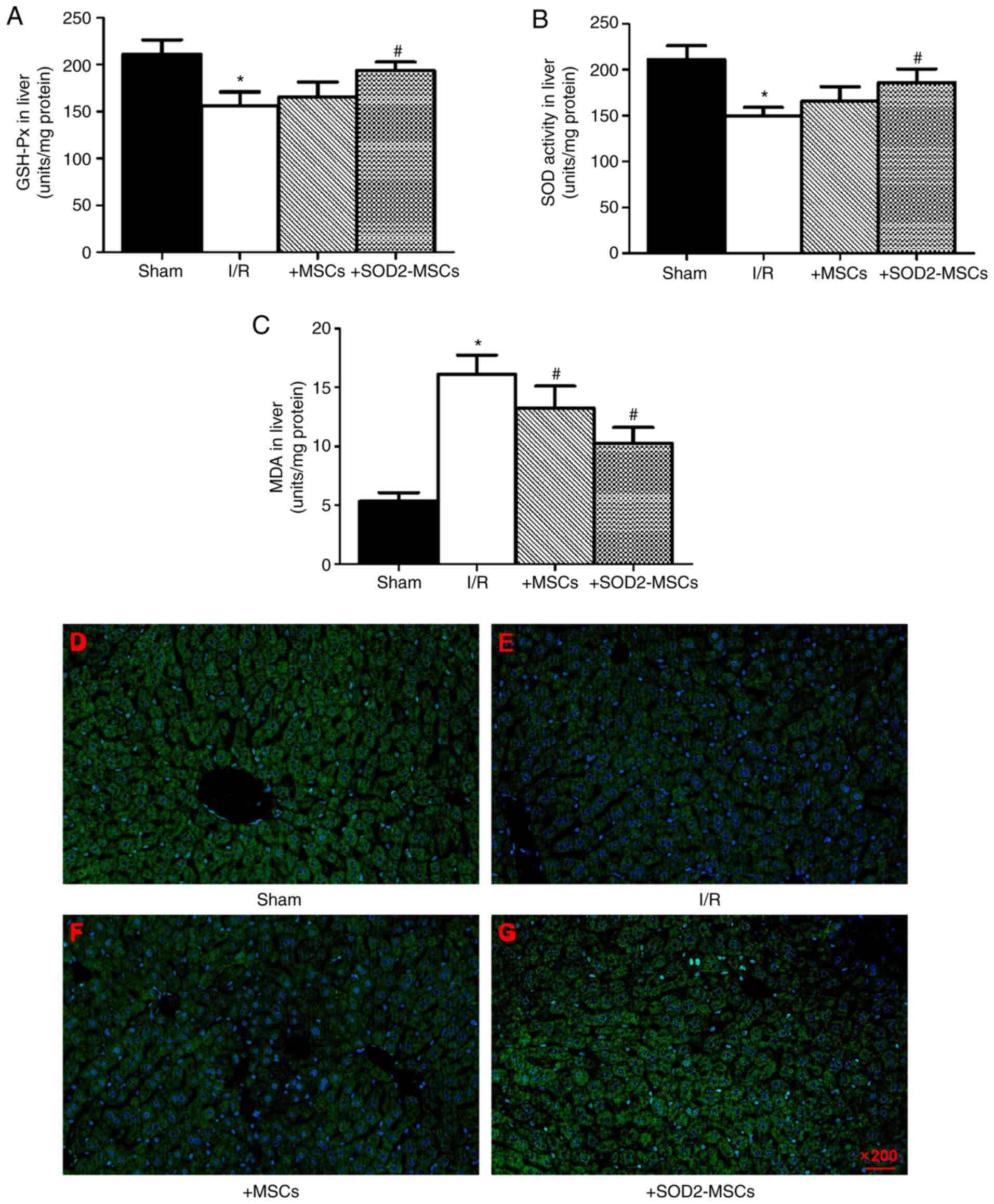
BM-MSCs overexpressing SOD2 attenuate
the oxidative stress response in I/R injury
The oxidative stress response is considered to be an
important contributor to HIRI (26). Therefore, SOD, GSH-Px and MDA levels
were assessed after HIRI induction. It was found that SOD and
GSH-Px levels were markedly decreased, while the MDA level was
notably increased in I/R rats compared with the sham group
(*P<0.05; Fig. 4A-C). These
findings indicated that elevated oxidative stress was an important
contributor in liver injury. Moreover, in the I/R + MSCs and I/R +
SOD2-MSCs groups, increased SOD and GSH-Px activities were
observed, while MDA levels were decreased compared with the I/R
group (#P<0.05; Fig.
4A-C). In line with the results of the biochemical index, the
production of reactive oxygen species (ROS) was detected in the
liver tissues. The results demonstrated that an increased number of
apoptotic cells was observed in the hepatic I/R group compared with
the sham group, whereas the number of apoptotic cells was notably
decreased in the I/R + MSCs and I/R + SOD2-MSCs groups compared
with the I/R group (Fig. 4D-G).
These findings suggested that transplanted SOD2-overexpressing
BM-MSCs attenuated the oxidative stress in HIRI.
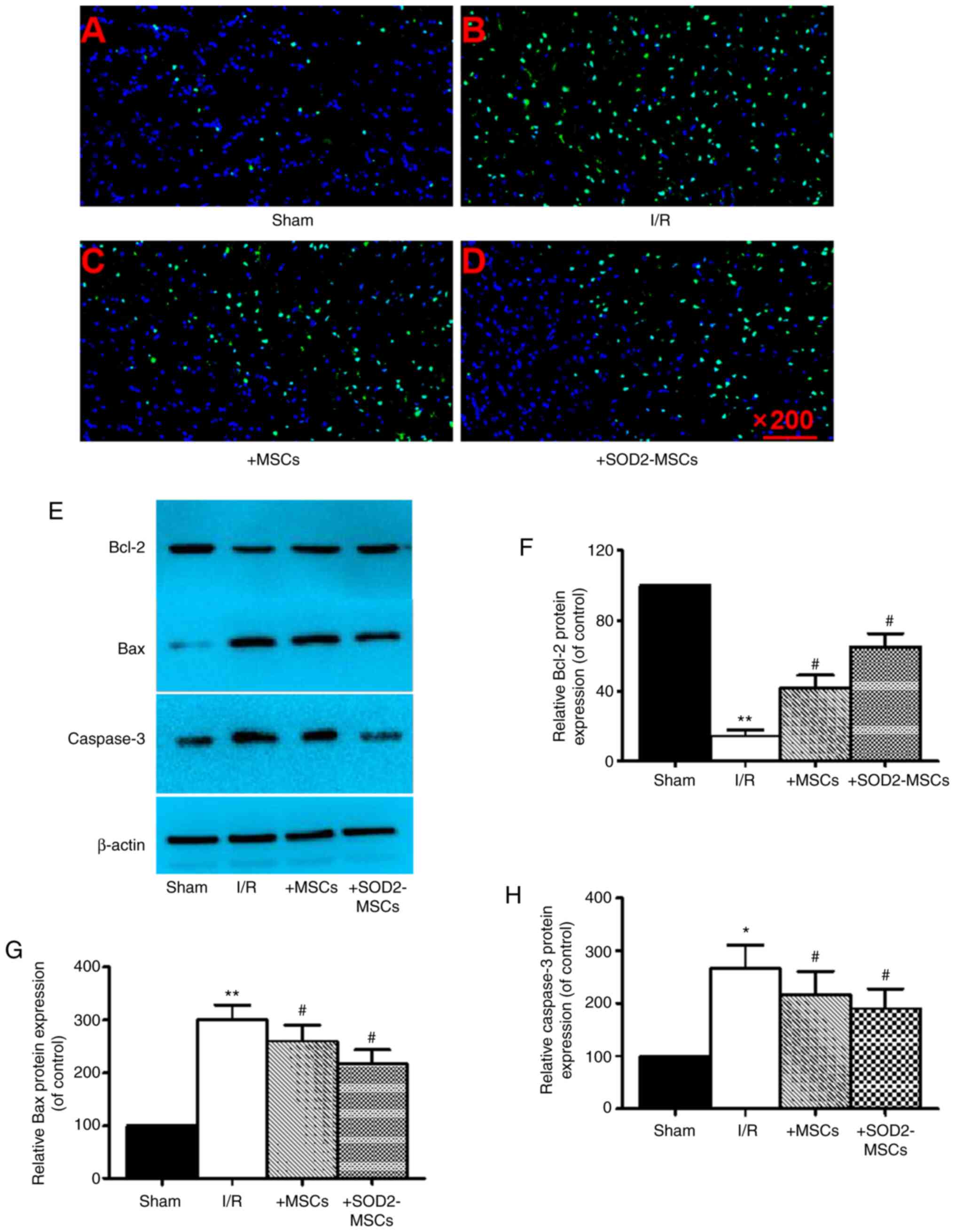
BM-MSCs overexpressing SOD2 inhibit
cell apoptosis after HIRI
To investigate the effect of SOD2-overexpressing
BM-MSCs against HIRI-related apoptosis, TUNEL staining was firstly
utilized to examine the number of apoptotic cells and western
blotting was used to detect the expression levels of Bcl-2, Bax and
caspase-3. It was identified that hepatic I/R caused a markedly
higher AI, suggesting that I/R resulted in hepatocyte apoptosis
(Fig. 5A and B). Furthermore, the
findings indicated a significant downregulation in Bcl-2
expression, and an upregulation in Bax and caspase-3 expression in
the I/R group compared with the sham group (*P<0.05; Fig. 5E-H). It was also found that these
effects were partially reversed in the I/R + MSCs and I/R +
SOD2-MSCs groups compared with the I/R group
(#P<0.05; Fig. 5C-H).
Collectively, these results suggested that SOD2-overexpressing
BM-MSCs exerted an inhibitory effect on cell apoptosis in HIRI.
Discussion
HIRI is a widespread clinical concern, which is
common across multiple pathological conditions and liver surgical
procedures, such as hemorrhagic shock, liver resection and liver
transplantation (27). Hepatic I/R
can lead to hepatic sinusoidal endothelial cell damage, Kupffer
cell and leukocyte activation, leukocyte and platelet adhesion and
microcirculation disorders, which directly affect the prognosis of
the disease, the success rate of surgery and the survival rate of
patients (28). The main mechanisms
of HIRI are associated with intracellular calcium overload and
oxygen free radicals (29). Thus,
it is important for investigators to develop for novel strategies
for hepatic I/R treatment.
BM-MSCs have a strong self-replication ability and
multi-differentiation potential, which are based on their
pluripotency and the secretion of beneficial molecules; therefore,
these cells are widely used in basic and clinical trials (6,13). It
has been reported that BM-MSCs possess potent migratory and
differentiation abilities under both physiological and pathological
conditions, and may be cultured in vitro under specific
conditions to differentiate into cells derived from multiple germ
layers, such as hepatocyte-like and adult cells (30,31).
Accumulating evidence has indicated that nerve
cells, adipocytes, osteoblasts and stem cell transplantation
therapy are conducive to tissue damage repair and functional
recovery (32–34). HIRI is a common pathological event
during liver surgery, and is widely accepted as a common and
inevitable complication during liver resection and liver
transplantation, particularly in patients with end-stage liver
disease (35). Moreover, the
commonly used treatment methods for this include liver resection
and liver transplantation, and thus, a method to reduce the I/R
injury of the liver during surgery will be of great significance to
the clinical treatment of liver diseases (36). Previous studies have confirmed that
BM-MSC transplantation exerts positive effects in treatment and has
a protective function against liver tissue damage, and may be a
potential intervention in the field of medicine and biology
(37,38). It has also been revealed that the
overexpression of cytokines and growth factors targeting certain
diseases may be effective strategies for dysfunction-targeted
therapy (39). Therefore, the
method of gene transfer allows BM-MSCs to express additional genes
that are beneficial to treatment, and can enhance the efficacy of
BM-MSC transplantation.
Intravenous administration of BM-MSCs has become a
widely accepted treatment for decreasing intracellular calcium
overload and oxygen free radicals, as well as for hepatoprotection
following HIRI (13). The present
study used the method of adenovirus transfection to overexpress the
SOD2 gene in MSCs, and RT-qPCR and western blotting demonstrated
that SOD2 expression was upregulated in MSCs after adenovirus
transfection, suggesting that SOD2 overexpression was successfully
established. Moreover, SOD2 overexpression decreased oxidative
stress and increased the expression levels of apoptosis-related
genes.
Oxidative stress caused by hepatic I/R is the main
mechanism contributing to the deterioration of liver function
induced by I/R (40). SODs are
endogenous antioxidants that catalyze the conversion of
O2·− to H2O2, and helps maintain
the redox balance by diffusing the superoxide (41). SOD2, a member of the SOD family, is
a mitochondrial antioxidant enzyme that scavenges superoxide
radicals (42). The continuous
expression of SOD2 is regulated by the cell oxygen concentration,
and once hepatic hypoxic-ischemic injury occurs, SOD2 is activated
(43). By regulating its downstream
target genes, SOD2 participates in regulating anaerobic metabolism,
cytokine production and regulating cell apoptosis (44). Therefore, SOD2 overexpression may be
a promising novel intervention for the treatment of hepatic
ischemia and hypoxic injury.
An increasing number of basic and clinical
experiments have reported that SOD2 may serve a protective role in
hepatocytes during HIRI (45).
Reduction of SOD2 activity has been shown to decrease liver
function induced by transient ischemia (44,46).
Moreover, overexpression of SOD2 can provide direct
hepatic-protection via increasing levels of serum transaminases
(including AST and ALT) and reversing pathological changes
(47,48). In animal models of HIRI, significant
increases in liver SOD2 mRNA and protein expression levels have
been observed (47,48). These findings support the idea that
SOD2-overexpressing BM-MSC transplantation may mitigate HIRI. In
the present study, it was demonstrated that HIRI caused increases
in the levels of AST and ALT, and SOD2-overexpressing BM-MSC
transplantation obviously decreased the abnormal increases of these
indicators, which in line with the reversal of pathological changes
in HIRI.
In the current study, it was observed that
SOD2-overexpressing BM-MSCs exerted the protective effect of SOD2
against oxidative stress in rats subjected to hepatic I/R.
Furthermore, SOD2 overexpression in BM-MSCs exerted protective
effects in promoting liver function recovery and decreasing
oxidative stress in a rat model of hepatic I/R. These effects were
mediated via the amelioration of oxidative stress and apoptosis
inhibition after BM-MSC transplantation. SODs are endogenous
antioxidants, and their activities are associated with SOD2 levels.
The present study demonstrated that SOD and GSH-Px activities were
significantly increased, and MDA content was decreased after
SOD2-overexpressing BM-MSC transplantation.
The concentration of lipid peroxides in the plasma
during HIRI is closely associated with serum transaminase activity,
and is parallel to the severity of liver morphological damage
(49). The present findings
indicated that I/R caused obvious necrosis and congestion in the
injured liver lobules, accompanied by the presence of a large
number of red blood cells in the venules and inflammatory cell
infiltration. However, after SOD2-overexpressing BM-MSC
transplantation, the liver tissue structure was partially restored
to normal, although changes, such as dilated central veins, were
still visible (50). These results
suggested that MSC treatment regenerated damaged liver tissues
subjected to I/R.
SOD2 exerts a hepatoprotective effect in HIRI, and
several studies have reported that the increase in sirtuin (Sirt)3
activity promotes the deacetylation of SOD2, which can decrease
oxidative stress by eliminating ROS (46,51).
Sirt3 can stabilize hypoxia-inducible factor 1α (HIF-1α), and
HIF-1α can activate the Sirt3 gene promoter, which leads to
increased Sirt3 mRNA transcript synthesis and the inactivation of
cyclophilin D, thereby reducing the opening of the mitochondrial
permeability transition pore channel. In turn, the extent of liver
damage can be reduced (52). The
present results demonstrated that SOD2 overexpression enhanced the
therapeutic effect of BM-MSCs in HIRI, improved liver function and
effectively reduced the degree of tissue injury. Compared with the
BM-MSCs group and control group, liver tissue damage in the
SOD2-MSCs group was significantly reduced. These results indicated
that SOD2 overexpression could exert a positive synergistic effect
with BM-MSCs in the treatment of liver injury.
However, the limitations of the current study have
yet to be considered. Despite progress in revealing the
ameliorative potentials of SOD2 overexpression in MSCs, the
complete regulatory molecular targets of SOD2 overexpression
require additional research. Moreover, the application of SOD2
overexpression in MSCs for the treatment of other liver I/R-related
injuries, including liver transplantation, requires more rigorous
and scientific verification, as well as subsequent clinical
trials.
In conclusion, the present study demonstrated that
BM-MSCs could be a potential new method for cell therapy,
particularly in liver injury. At present, the mortality rate of
liver injury is increasing worldwide annually, making the
development of novel strategies for liver injury treatment highly
important. The present study identified that transplantation of
SOD2-overexpressing BM-MSCs may have the potential to ameliorate
HIRI via the inhibition of oxidative stress and apoptosis.
Acknowledgements
The authors would like to thank the Laboratory
Animal of Second Xiangya Hospital of Central South University for
their technical assistance.
Funding
The present study was supported by the National
Natural Science Foundation of China (grant nos. 81571784 and
30870695), the Provincial Natural Science Foundation of Hunan
(grant nos. 2019JJ4044 and 2018JJ2578) and the Scientific Research
Project of Hunan Health and Family Planning Commission (grant no.
B20180048).
Availability of data and materials
All data generated or analyzed during this study are
included in this published article.
Authors' contributions
QL, WZ and EX designed and performed experiments,
and analyzed, interpreted and presented the results for group
discussions. QL, WZ and EX confirm the authenticity of all the raw
data. EX made substantial contributions to the conception of the
study. All authors read and approved the final manuscript.
Ethics approval and consent to
participate
The protocol of the present study was approved by
the Laboratory Animal Center, The Second Xiangya Hospital of
Central South University (Changsha, China).
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Huang Y, Gao F, Zhang Y, Chen Y, Wang B,
Zheng Y and Shi G: N-n-Butyl haloperidol iodide inhibits the
augmented Na+/Ca2+ exchanger currents and
L-type Ca2+ current induced by hypoxia/reoxygenation or
H2O2 in cardiomyocytes. Biochem Biophys Res Commun. 421:86–90.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Cannistrà M, Ruggiero M, Zullo A, Gallelli
G, Serafini S, Maria M, Naso A, Grande R, Serra R and Nardo B:
Hepatic ischemia reperfusion injury: A systematic review of
literature and the role of current drugs and biomarkers. Int J
Surg. 33 (Suppl 1):S57–S70. 2016. View Article : Google Scholar
|
|
3
|
Hirakawa Y, Tsuchishima M, Fukumura A,
Kinoshita K, Hayashi N, Saito T, George J, Toshikuni N, Ueda Y and
Tsutsumi M: Recombinant thrombomodulin prevented hepatic
ischemia-reperfusion injury by inhibiting high-mobility group box 1
in rats. Eur J Pharmacol. 863:1726812019. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Li J, Li RJ, Lv GY and Liu HQ: The
mechanisms and strategies to protect from hepatic
ischemia-reperfusion injury. Eur Rev Med Pharmacol Sci.
19:2036–2047. 2015.PubMed/NCBI
|
|
5
|
Chao YH, Lin CW, Pan HH, Yang SF, Weng TF,
Peng CT and Wu KH: Increased apoptosis and peripheral blood
mononuclear cell suppression of bone marrow mesenchymal stem cells
in severe aplastic anemia. Pediatr Blood Cancer. 65:e272472018.
View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Rong Y, Liu W, Wang J, Fan J, Luo Y, Li L,
Kong F, Chen J, Tang P and Cai W: Neural stem cell-derived small
extracellular vesicles attenuate apoptosis and neuroinflammation
after traumatic spinal cord injury by activating autophagy. Cell
Death Dis. 10:3402019. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Zha JM, Li HS, Lin Q, Kuo WT, Jiang ZH,
Tsai PY, Ding N, Wu J, Xu SF, Wang YT, et al: Interleukin 22
expands transit-amplifying cells while depleting Lgr5+
stem cells via inhibition of wnt and notch signaling. Cell Mol
Gastroenterol Hepatol. 7:255–274. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Sun X, Luo L and Li J: LncRNA MALAT1
facilitates BM-MSCs differentiation into endothelial cells via
targeting miR-206/VEGFA axis. Cell Cycle. 19:3018–3028. 2020.
View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Zuo R, Liu M, Wang Y, Li J, Wang W, Wu J,
Sun C, Li B, Wang Z, Lan W, et al: BM-MSC-derived exosomes
alleviate radiation-induced bone loss by restoring the function of
recipient BM-MSCs and activating Wnt/β-catenin signaling. Stem Cell
Res Ther. 10:302019. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Tan YF, Tang L, OuYang WX, Jiang T, Zhang
H and Li SJ: β-catenin-coordinated lncRNA MALAT1 up-regulation of
ZEB-1 could enhance the telomerase activity in HGF-mediated
differentiation of bone marrow mesenchymal stem cells into
hepatocytes. Pathol Res Pract. 215:546–554. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Shay JW, Homma N, Zhou R, Naseer MI,
Chaudhary AG, Al-Qahtan M, Hirokawa N, Goudarzi M, Fornace AJ Jr,
Baeesa S, et al: Abstracts from the 3rd international genomic
medicine conference (3rd IGMC 2015): Jeddah, kingdom of Saudi
Arabia. 30 November-3 December 2015. BMC Genomics. 17 (Suppl
6):S4872016. View Article : Google Scholar
|
|
12
|
Wang X, Wang S, Zhou Y, Obulkasim H, Zhang
ZH, Dai B, Zhu W and Shi XL: BM-MSCs protect against liver
ischemia/reperfusion injury via HO-1 mediated autophagy. Mol Med
Rep. 18:2253–2262. 2018.PubMed/NCBI
|
|
13
|
Yang B, Duan W, Wei L, Zhao Y, Han Z, Wang
J, Wang M, Dai C, Zhang B, Chen D and Chen Z: Bone Marrow
mesenchymal stem cell-derived hepatocyte-like cell exosomes reduce
hepatic ischemia/reperfusion injury by enhancing autophagy. Stem
Cells Dev. 29:372–379. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Zare MA, Zare A, Azarpira N and Pakbaz S:
The protective effect of bone marrow-derived mesenchymal stem cells
in liver ischemia/reperfusion injury via down-regulation of
miR-370. Iran J Basic Med Sci. 22:683–689. 2019.PubMed/NCBI
|
|
15
|
Zheng J, Chen L, Lu T, Zhang Y, Sui X, Li
Y, Huang X, He L, Cai J, Zhou C, et al: MSCs ameliorate
hepatocellular apoptosis mediated by PINK1-dependent mitophagy in
liver ischemia/reperfusion injury through AMPKα activation. Cell
Death Dis. 11:2562020. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Jin G, Qiu G, Wu D, Hu Y, Qiao P, Fan C
and Gao F: Allogeneic bone marrow-derived mesenchymal stem cells
attenuate hepatic ischemia-reperfusion injury by suppressing
oxidative stress and inhibiting apoptosis in rats. Int J Mol Med.
31:1395–1401. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Chen P, Cui L, Chen G, You T, Li W, Zuo J,
Wang C, Zhang W and Jiang C: The application of
BMP-12-overexpressing mesenchymal stem cells loaded 3D-printed PLGA
scaffolds in rabbit rotator cuff repair. Int J Biol Macromol.
138:79–88. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Shi X, Bai Y, Zhang G, Liu Y, Xiao H, Liu
X and Zhang W: Effects of over-expression of SOD2 in bone
marrow-derived mesenchymal stem cells on traumatic brain injury.
Cell Tissue Res. 372:67–75. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Lee M, Song BR, Kim DH, Ha J, Lee M, Choi
SJ, Oh W, Um S and Jin HJ: Up-regulation of superoxide dismutase 2
in 3D spheroid formation promotes therapeutic potency of human
umbilical cord blood-derived mesenchymal stem cells. Antioxidants
(Basel). 9:662020. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Mohr BJ, Fakoya FA, Hau J, Souilem O and
Anestidou L: The Governance of Animal Care and Use for Scientific
Purposes in Africa and the Middle East. ILAR J. 57:333–346. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Krüger-Haag A, Lehmann C, Schmidt E,
Sonntag F, Hörer M and Kochanek S: Evaluation of life cycle
defective adenovirus mutants for production of adeno-associated
virus vectors. J Gene Med. 21:e30942019. View Article : Google Scholar
|
|
22
|
Kronenburg A, Bulder MMM, Bokkers RPH,
Hartkamp NS, Hendrikse J, Vonken EJ, Kappelle LJ, van der Zwan A,
Klijn CJM and Braun KPJ: Cerebrovascular reactivity measured with
ASL perfusion MRI, Ivy sign, and regional tissue vascularization in
moyamoya. World Neurosurg. 125:e639–e650. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Lv FJ, Tuan RS, Cheung KM and Leung VY:
Concise review: The surface markers and identity of human
mesenchymal stem cells. Stem Cells. 32:1408–1419. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Xu L, Yu Y, Sang R, Li J, Ge B and Zhang
X: Protective effects of taraxasterol against ethanol-induced liver
injury by regulating CYP2E1/Nrf2/HO-1 and NF-kappaB signaling
pathways in mice. Oxid Med Cell Longev. 2018:82841072018.
View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Yuan R, Tao X, Liang S, Pan Y, He L, Sun
J, Wenbo J, Li X, Chen J and Wang C: Protective effect of acidic
polysaccharide from Schisandra chinensis on acute ethanol-induced
liver injury through reducing CYP2E1-dependent oxidative stress.
Biomed Pharmacother. 99:537–542. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Zhang M, Yang D, Gong X, Ge P, Dai J, Lin
L and Zhang L: Protective benefits of AMP-activated protein kinase
in hepatic ischemia-reperfusion injury. Am J Transl Res. 9:823–829.
2017.PubMed/NCBI
|
|
28
|
Peralta C, Rull R, Rimola A, Deulofeu R,
Roselló-Catafau J, Gelpí E and Rodés J: Endogenous nitric oxide and
exogenous nitric oxide supplementation in hepatic
ischemia-reperfusion injury in the rat. Transplantation.
71:529–536. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Hataji K, Watanabe T, Oowada S, Nagaya M,
Kamibayashi M, Murakami E, Kawakami H, Ishiuchi A, Kumai T, Nakano
H, et al: Effects of a calcium-channel blocker (CV159) on hepatic
ischemia/reperfusion injury in rats: Evaluation with selective
NO/pO2 electrodes and an electron paramagnetic resonance
spin-trapping method. Biol Pharm Bull. 33:77–83. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Charbord P: Bone marrow mesenchymal stem
cells: Historical overview and concepts. Hum Gene Ther.
21:1045–1056. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Zhou W, Lin J, Zhao K, Jin K, He Q, Hu Y,
Feng G, Cai Y, Xia C, Liu H, et al: Single-cell profiles and
clinically useful properties of human mesenchymal stem cells of
adipose and bone marrow origin. Am J Sports Med. 47:1722–1733.
2019. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Kassis I, Vaknin-Dembinsky A and Karussis
D: Bone marrow mesenchymal stem cells: Agents of immunomodulation
and neuroprotection. Curr Stem Cell Res Ther. 6:63–68. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Miao C, Lei M, Hu W, Han S and Wang Q: A
brief review: The therapeutic potential of bone marrow mesenchymal
stem cells in myocardial infarction. Stem Cell Res Ther. 8:2422017.
View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Yang W and Ma B: A mini-review: The
therapeutic potential of bone marrow mesenchymal stem cells and
relevant signaling cascades. Curr Stem Cell Res Ther. 14:214–218.
2019. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Yang C, Yang HJ, Deng SP and Zhang Y:
Current status of ex-vivo liver resection and autologous liver
transplantation for end-stage hepatic alveolar echinococcosis. Ann
Palliat Med. 9:2271–2278. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Konishi T and Lentsch AB: Hepatic
Ischemia/reperfusion: Mechanisms of tissue injury, repair, and
regeneration. Gene Expr. 17:277–287. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Mohamed HE, Elswefy SE, Rashed LA, Younis
NN, Shaheen MA and Ghanim AM: Bone marrow-derived mesenchymal stem
cells effectively regenerate fibrotic liver in bile duct ligation
rat model. Exp Biol Med (Maywood). 241:581–591. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Souza MC, Silva JD, Pádua TA, Torres ND,
Antunes MA, Xisto DG, Abreu TP, Capelozzi VL, Morales MM, Sá
Pinheiro AA, et al: Mesenchymal stromal cell therapy attenuated
lung and kidney injury but not brain damage in experimental
cerebral malaria. Stem Cell Res Ther. 6:1022015. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Jain R, Dey B, Khera A, Srivastav P, Gupta
UD, Katoch VM, Ramanathan VD and Tyagi AK: Over-expression of
superoxide dismutase obliterates the protective effect of BCG
against tuberculosis by modulating innate and adaptive immune
responses. Vaccine. 29:8118–8125. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Du Y, Qian B, Gao L, Tan P, Chen H, Wang
A, Zheng T, Pu S, Xia X and Fu W: Aloin preconditioning attenuates
hepatic ischemia/reperfusion injury via inhibiting TLR4/MyD88/NF-κB
signal pathway in vivo and in vitro. Oxid Med Cell Longev.
2019:37658982019. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Marampon F, Codenotti S, Megiorni F, Del
Fattore A, Camero S, Gravina GL, Festuccia C, Musio D, De Felice F,
Nardone V, et al: NRF2 orchestrates the redox regulation induced by
radiation therapy, sustaining embryonal and alveolar
rhabdomyosarcoma cells radioresistance. J Cancer Res Clin Oncol.
145:881–893. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Zelko IN, Mariani TJ and Folz RJ:
Superoxide dismutase multigene family: A comparison of the CuZn-SOD
(SOD1), Mn-SOD (SOD2), and EC-SOD (SOD3) gene structures,
evolution, and expression. Free Radical Biol Med. 33:337–349. 2002.
View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Katwal G, Baral D, Fan X, Weiyang H, Zhang
X, Ling L, Xiong Y, Ye Q and Wang Y: SIRT3 a major player in
attenuation of hepatic ischemia-reperfusion injury by reducing ROS
via its downstream mediators: SOD2, CYP-D, and HIF-1α. Oxid Med
Cell Longev. 2018:29769572018. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Miyauchi T, Uchida Y, Kadono K, Hirao H,
Kawasoe J, Watanabe T, Ueda S, Jobara K, Kaido T, Okajima H, et al:
Preventive effect of antioxidative nutrient-rich enteral diet
against liver ischemia and reperfusion injury. JPEN J Parenter
Enteral Nutr. 43:133–144. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Elias-Miró M, Mendes-Braz M, Cereijo R,
Villarroya F, Jiménez-Castro MB, Gracia-Sancho J, Guixé-Muntet S,
Massip-Salcedo M, Domingo JC, Bermudo R, et al: Resistin and
visfatin in steatotic and non-steatotic livers in the setting of
partial hepatectomy under ischemia-reperfusion. J Hepatol.
60:87–95. 2014. View Article : Google Scholar
|
|
46
|
Katwal G, Baral D, Fan X, Weiyang H, Zhang
X, Ling L, Xiong Y, Ye Q and Wang Y: SIRT3 a major player in
attenuation of hepatic ischemia-reperfusion injury by reducing ROS
via its downstream mediators: SOD2, CYP-D, and HIF-1α. Oxid Med
Cell Longev. 2018:29769572018. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Wheeler MD, Katuna M, Smutney OM, Froh M,
Dikalova A, Mason RP, Samulski RJ and Thurman RG: Comparison of the
effect of adenoviral delivery of three superoxide dismutase genes
against hepatic ischemia-reperfusion injury. Hum Gene Ther.
12:2167–2177. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Wheeler MD, Nakagami M, Bradford BU,
Uesugi T, Mason RP, Connor HD, Dikalova A, Kadiiska M and Thurman
RG: Overexpression of manganese superoxide dismutase prevents
alcohol-induced liver injury in the rat. J Biol Chem.
276:36664–36672. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Rodríguez-Reynoso S, Leal-Cortés C,
Portilla-de Buen E and López-De la Torre SP: Ischemic
preconditioning preserves liver energy charge and function on
hepatic ischemia/reperfusion injury in rats. Arch Med Res.
49:373–380. 2018. View Article : Google Scholar
|
|
50
|
Wang X, Wang S, Zhou Y, Obulkasim H, Zhang
ZH, Dai B, Zhu W and Shi XL: BM-MSCs protect against liver
ischemia/reperfusion injury via HO-1 mediated autophagy. Mol Med
Rep. 18:2253–2262. 2018.PubMed/NCBI
|
|
51
|
Li DP, Chen YL, Jiang HY, Chen Y, Zeng XQ,
Xu LL, Ye Y, Ke CQ, Lin G, Wang JY and Gao H: Phosphocreatine
attenuates Gynura segetum-induced hepatocyte apoptosis via a
SIRT3-SOD2-mitochondrial reactive oxygen species pathway. Drug Des
Devel Ther. 13:2081–2096. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Wang Q, Wei S, Li L, Qiu J, Zhou S, Shi C,
Shi Y, Zhou H and Lu L: TGR5 deficiency aggravates hepatic
ischemic/reperfusion injury via inhibiting SIRT3/FOXO3/HIF-1
pathway. Cell Death Discov. 6:1162020. View Article : Google Scholar : PubMed/NCBI
|