Introduction
Acute liver failure (ALF) is a fatal hepatic disease
associated with rapid loss of liver function, resulting in
multiorgan dysfunction, encephalopathy and coagulopathy in
patients. This critical illness has fatal consequences, and the
only treatment option is an emergency liver transplant (1). Therefore, an effective drug for
treating ALF is urgently required.
Kaempferol is a flavonoid that is primarily
extracted from the root of Kaempferia galanga L., and it is
widely present in various natural plants, fruits, vegetables,
beverages and teas (2,3). Kaempferol possesses several
pharmacological properties, including cardioprotective,
neuroprotective, antioxidative, antidiabetic and anticarcinogenic
effects (4). A growing number of
studies have shown that kaempferol can reduce the risk of
developing various cardiovascular diseases, diabetes and cancer,
amongst other diseases (4–6). It has been reported that kaempferol
may be associated with the treatment of numerous diseases. For
example, in a time- and concentration-dependent manner, kaempferol
can decrease HeLa cervical cancer cell viability and induce
apoptosis by downregulating the activity of the PI3K/AKT signaling
pathway (7). Previous research also
revealed that kaempferol distinctly inhibited the proliferation of
the EJ bladder cancer cell line by inducing S cell cycle arrest and
apoptosis, and increasing the expression levels of phosphorylated
(p)-p53 via regulation of the mitochondrial-mediated apoptotic
signaling pathways (8).
The potential protective effects of kaempferol on
liver injury have been previously demonstrated (9–11),
where it has been shown to exhibit effective anti-inflammatory
properties in liver cells (12). In
addition, kaempferol has been demonstrated to significantly
alleviate acute liver injury, inflammation and early hepatocyte
apoptosis caused by propacetamol (13). Our previous study revealed that
kaempferol pretreatment could alleviate liver damage in
D-galactosamine (D-GalN)/lipopolysaccharide (LPS)-induced mice
(14). However, the underlying
molecular mechanisms via which kaempferol exerts it effects in ALF
remain to be determined.
Autophagy is an intracellular catabolic signaling
pathway, which is a highly conserved evolutionarily process, where
biomolecules and organelles can be degraded by the lysosomes.
Autophagy is crucial for maintaining the homeostasis of cells and
replenishing several types of substances for cell survival under
stressful conditions (15).
Autophagy is closely associated with liver disease. It has
previously been observed that autophagy may suppress the growth of
tumors in chronic liver disease, and impaired autophagy can lead to
a significant increase in glycolysis in liver cancer cells
(16). In addition, autophagy can
protect against the accumulation of fat in hepatocytes during
nonalcoholic fatty liver disease (17). In our previous study, it was
identified that activation of autophagy protected mice from ALF by
inhibiting GSK-3β activity (18).
In current therapeutic research, the problem of drug toxicity
remains a persistent issue. To the best of our knowledge, there are
no previous studies showing the effects of different doses of
kaempferol on ALF. Therefore, the toxicity of kaempferol requires
further study.
Given the aforementioned information, in the present
study, a mouse model of ALF induced by D-GalN/LPS was utilized, as
it has been widely used to examine the underlying mechanisms of
potential therapeutic drugs for the treatment of ALF (19,20).
The functional effects of different doses of kaempferol were
determined, the survival rate, liver function and levels of
inflammatory cytokines were assessed, and the related regulatory
pathways were evaluated in the context of ALF.
Materials and methods
Animals and treatments
A total of 40 male wild-type mice (C57BL/6; age,
8–12 weeks) were purchased from Capital Medical University (CMU;
Beijing, China). The mice were housed in a standard environment
with a controlled temperature of 22±2°C and humidity of 55±5%,
under a 12-h light/dark cycle. Animals were provided with ad
libitum access to water and food. The animal experimental
protocol was approved by the Institutional Animal Care and Use
Committee of CMU (approval no. AEEI-2020-009 on 2019/12/30).
The mice were injected intraperitoneally with D-GalN
(700 mg/kg; Sigma-Aldrich; Merck KGaA) and LPS (10 µg/kg;
InvivoGen) to induce ALF, or were injected with an equivalent
volume of normal saline. A total of 2 h before the administration
of D-GalN/LPS, mice were pretreated with different doses (2.5, 5,
10, 20 or 40 mg/kg) of kaempferol (Sigma-Aldrich; Merck KGaA) via
tail vein injection (n=8/group). Survival analysis was conducted
based on the number of survivors in each group. 3-methyladenine
(3-MA; Sigma-Aldrich; Merck KGaA) can be used to block autophagy,
and chloroquine (CQ; Sigma-Aldrich; Merck KGaA) can be used to
inhibit the fusion of lysosomes and autophagosomes, and are
commonly used to assess autophagic flux (21,22).
3-MA (10 mg/kg) or CQ (60 mg/kg) were administered
intraperitoneally 2 h before the administration of D-GalN/LPS. The
mice were anesthetized by intraperitoneal injection of
pentobarbital sodium (60 mg/kg). Liver tissues were collected for
further analysis. Blood samples (~100 µl) were collected from the
eyeballs. Subsequently, mice were sacrificed by intraperitoneal
injection of 100 mg/kg sodium pentobarbital after 24 h, and death
was confirmed by observing respiration and by using the corneal
reflection method.
Serum aminotransferase activity
Blood samples were collected from the mice 6 h after
D-GalN/LPS administration. Serum levels of alanine aminotransferase
(ALT) and aspartate aminotransferase (AST), which are markers of
hepatic damage, were measured using a multiparametric analyzer (AU
5400; Olympus Corporation) according to the manufacturer's protocol
(23).
Histopathological analysis
Liver samples were fixed in 4% formaldehyde solution
for 12 h at 4°C and embedded in paraffin wax, after which, they
were sectioned (4 µm) and stained using H&E, according to
routine procedure for histopathological evaluation; sections were
stained with hematoxylin for 10 min and with eosin for 10 sec at
room temperature. Tissues were observed using a light microscope
(magnification, ×200).
Reverse transcription-quantitative
(RT-q) PCR
Total RNA was isolated from liver tissues using
TRIzol® reagent (Invitrogen; Thermo Fisher Scientific,
Inc.) according to the manufacturer's protocol. RT of RNA (2.5 µg)
into cDNA was performed using a SuperScript III first-strand
synthesis system according to the manufacturer's protocol
(Invitrogen; Thermo Fisher Scientific, Inc.). A DNA Engine with a
Chromo 4 detector (MJ Research, Inc.; Bio-Rad Laboratories, Inc.)
was used for qPCR. The final reaction volume was 20 µl and
consisted of 1X super mix (Platinum SYBR-Green qPCR kit;
Invitrogen; Thermo Fisher Scientific, Inc.), 2 µl cDNA and 0.5 µl
each primer. The amplification conditions were as follows: Initial
denaturation at 50°C (2 min), 95°C (5 min), followed by 50 cycles
of 95°C (15 sec) and 60°C (30 sec). The mRNA expression levels were
calculated using the 2−ΔΔCq method (24). The sequences of the PCR primers used
are listed in Table I and were
produced by Sangon Biotech, Co., Ltd.
 | Table I.Sequences of the primers used for
reverse transcription-quantitative PCR. |
Table I.
Sequences of the primers used for
reverse transcription-quantitative PCR.
| Gene | Forward primers
(5′-3′) | Reverse primers
(5′-3′) |
|---|
| TNF-α |
GCCTCTTCTCATTCCTGCTTGT |
TTGAGATCCATGCCGTTG |
| CXCL-10 |
AAGTGCTGCCGTCATTTTCT |
GTGGCAATGATCTCAACACG |
| CXCL-2 |
AGTGAACTGCGCTGTCAATG |
TTCAGGGTCAAGGCAAACTT |
| IL-6 |
GCTACCAAACTGGATATAATCAGGA |
CCAGGTAGCTATGGTACTCCAGAA |
| IL-1β |
TTGACGGACCCCAAAAGAT |
GATGATCTGAGTGTGAGGGTCTG |
| IL-12p40 |
CAGCTTCTTCATCAGGGACAT |
CTTGAGGGAGAAGTAGGAATGG |
| IL-10 |
ACTGCACCCACTTCCCAGT |
TGTCCAGCTGGTCCTTTGTT |
| LC3 |
AGCAGCATCCAACCAAAATC |
CTGTGTCCGTTCACCAACAG |
| Atg7 |
ACCCAGAAGAAGCTGAACGA |
CTCATTTGCTGCTTGTTCCA |
| HPRT |
TCAACGGGGGACATAAAAGT |
TGCATTGTTTTACCAGTGTCAA |
Western blotting
Liver tissue samples or cells were lysed in RIPA
lysis buffer (Beyotime Institute of Biotechnology) supplemented
with phosphatase and protease inhibitors, and the concentration was
estimated using a Bradford Protein Assay kit (Beyotime Institute of
Biotechnology). A total of 20 µg protein per sample was loaded onto
a 12% SDS gel, resolved using SDS-PAGE at 80 V for 30 min and 120 V
for 1 h, and then transferred to a PVDF membrane using a Bio-Rad
blotting transfer system (Bio-Rad Laboratories, Inc.). The
membranes were blocked with 5% skimmed milk at room temperature for
1 h, and incubated overnight at 4°C with the following primary
antibodies: β-actin (1:1,000; cat. no. 8457S; Cell Signaling
Technology, Inc.), p-JNK (1:1,000; cat. no. 4668S; Cell Signaling
Technology, Inc.), p-ERK (1:1,000; cat. no. 4348S; Cell Signaling
Technology, Inc.), p-p38 (1:1,000; cat. no. 9215S; Cell Signaling
Technology, Inc.), LC3B (1:1,000; cat. no. 3868S; Cell Signaling
Technology, Inc.), autophagy related 7 (Atg7; 1:1,000; cat. no.
8558S; Cell Signaling Technology, Inc.) and p62 (1:1,000; cat. no.
23214S; Cell Signaling Technology, Inc.). After washing, the
membranes with TBS-0.5% Tween-20 (TBS-T) and the corresponding
HRP-conjugated secondary antibody (1:2,000; cat. no. 7074S; Cell
Signaling Technology, Inc.) in 10 ml blocking buffer was added and
incubated for 1 h at room temperature. Next, the membranes were
washed three times with TBS-T for 30 min, and signals were
visualized using an ECL kit (Thermo Fisher Scientific, Inc.).
Restore Western Blot Stripping Buffer was used to re-probe new
target proteins in a same membrane (Thermo Fisher Scientific,
Inc.). The blot was placed in Restore Western Blot Stripping Buffer
and incubated for 5–15 min at 37°C. Then, the blot was removed from
the Restore Western Blot Stripping Buffer and washed in PBS for
15–20 min. Next, as mentioned previously, the blot was incubated
with new primary and secondary antibodies. Densitometry analysis
was performed using ImageJ (version 1.49p; National Institutes of
Health).
Atg7 small interfering (si)RNA
treatment in vivo
siRNA (3 mg/kg; Shanghai Genepharma Co., Ltd.) and
an Entranster™ in vivo transfection reagent (Engreen
Biosystem Co.) were used to knock down Atg7 expression via the
administration of a hydrodynamic tail vein injection in mice. After
transfection for 24 h, subsequent experimentation was performed.
The sequence of Atg7 siRNA was 5′-GCAUCAUCUUCGAAGUGAATT-3′.
Scrambled siRNA (3 mg/kg) was used as a control and the sequence
was 5′-UUCUCCGAACGUGUCACGUTT-3′. These steps were performed in
accordance with the manufacturer's protocol.
Isolation of primary mouse
hepatocytes
Hanks' solution containing collagenase was used to
perfuse mouse livers when the mice were 7 weeks old, and, as
described previously, live hepatocytes were separated using Percoll
isocratic centrifugation at 4°C for 10 min at 10,000 × g (25). The number of mice used was 3, and
they were sacrificed by intraperitoneal injection of 100 mg/kg
sodium pentobarbital after 24 h; death was confirmed by observing
respiration and by using the corneal reflection method.
Starvation-induced autophagy in
vitro
The most robust method of inducing autophagy is
starvation of cells (26). Primary
hepatocytes were transfected with the GFP-LC3 plasmid (1 µg/ml;
Shanghai Genepharma Co., Ltd.) for 12 h at 37°C, and were treated
with serum-free medium for 4 h at 37°C. The percentage of cells was
calculated based on the number of GFP-LC3 puncta in the different
treatment groups. GFP-positive cells were regarded as cells that
exhibited bright, punctate staining. Per condition, ~50 cells were
counted, and the experiment was repeated at least three times.
Cells were observed using a fluorescence microscope (magnification,
×200).
Statistical analysis
The results from three independent experiments are
presented as the mean ± SD. Survival analysis was conducted using a
Kaplan-Meier plot with the log-rank test, and the P-value was
corrected a by Bonferroni's test. Statistical differences were
determined using one-way ANOVA followed by Bonferroni's post hoc
analysis in GraphPad Prism version 7 (GraphPad Software, Inc.).
P<0.05 was considered to indicate a statistically significant
difference.
Results
Effects of kaempferol on
D-GalN/LPS-induced ALF
First, the effects of different doses of kaempferol
on ALF induced by D-GalN/LPS in mice were examined. Different doses
of kaempferol (2.5, 5, 10, 20 or 40 mg/kg) were intraperitoneally
administered. As shown in the survival analysis (Fig. 1A), the survival rate of the 5 and 10
mg/kg kaempferol group was significantly increased compared with
that in the D-GalN/LPS-treated group, and the 5 mg/kg kaempferol
group showed the highest survival rate. This result suggested that
5 mg/kg kaempferol treatment effectively reduced D-GalN/LPS-induced
liver injury.
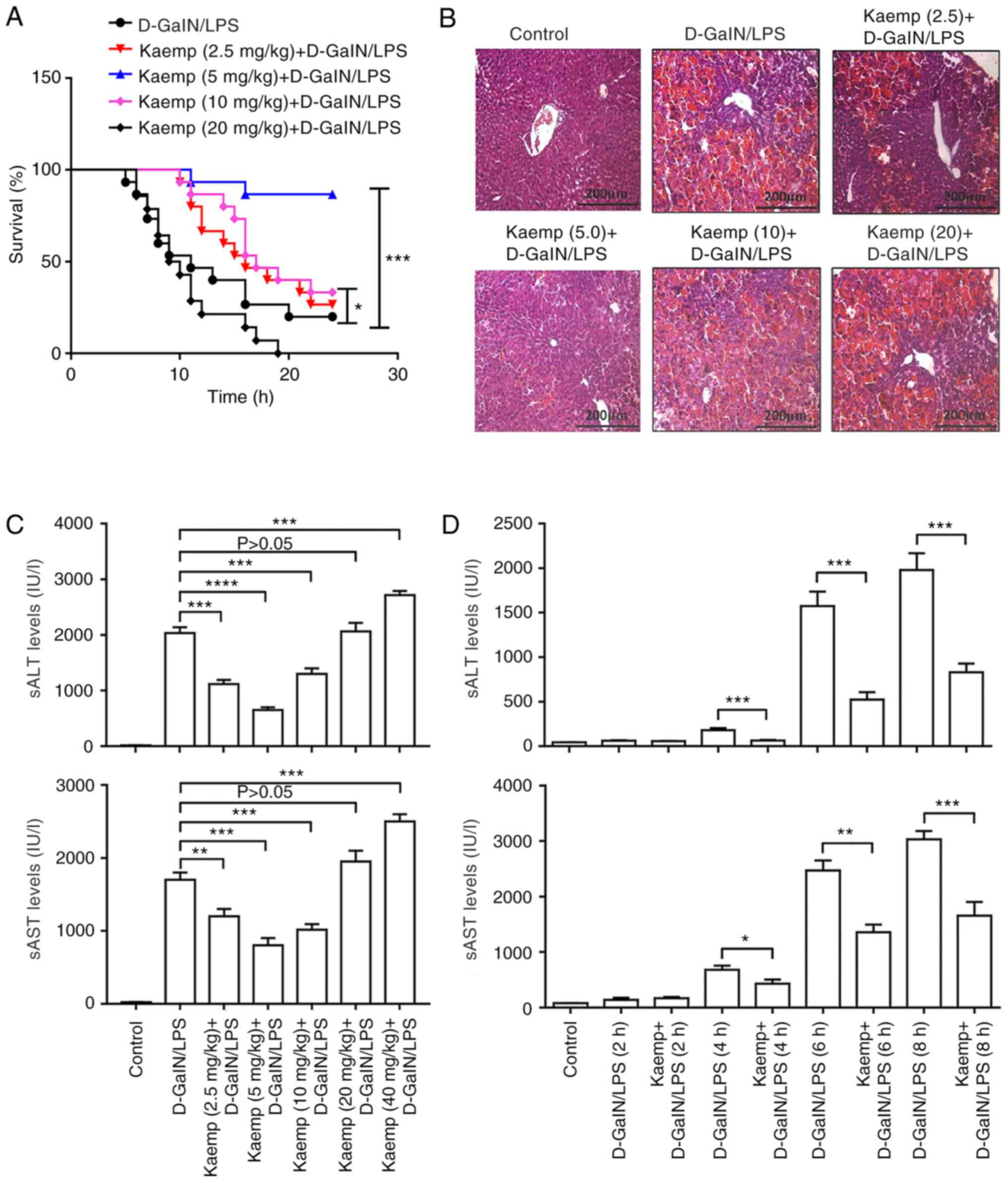 | Figure 1.Effects of different doses of kaemp
on D-GalN/LPS-induced ALF. Mice treated with kaemp + D-GalN/LPS
were administered kaemp (2.5, 5, 10, 20 or 40 mg/kg; intravenously)
prior to or after D-GalN/LPS injection (n=20). Control mice were
pretreated with saline before D-GalN/LPS injection (n=20). (A)
Survival rates of ALF-induced mice treated with different doses of
kaemp and the PBS + D-GalN/LPS-treated group. *P<0.05,
***P<0.001 vs. D-GalN/LPS treated group. (B) H&E-stained
liver sections of ALF induced model mice treated with different
doses of kaemp and the D-GalN/LPS-treated mice. Scale bar, 200 µm.
(C) sAST and sALT enzyme levels in the different groups. (D) sAST
and sALT enzyme levels in the different groups, including mice
pretreated with 5 mg/kg kaemp, were measured to evaluate liver
injury in mice after 2, 4, 6 and 8 h of D-GalN/LPS administration.
*P<0.05, **P<0.01, ***P<0.001, ****P<0.0001. ALF, acute
liver failure; sALT, serum alanine aminotransferase; sAST, serum
aspartate aminotransferase; D-GalN/LPS,
D-galactosamine/lipopolysaccharide; kaemp, kaempferol. |
Pretreatment with and 40 mg/kg kaempferol resulted
in higher serum ALT (sALT) and serum AST (sAST) levels compared
with the D-GalN/LPS induced group; whereas the 2.5, 5 and 10
mg/kg-treated mice exhibited lower levels of sALT and sAST,
particularly the 5 mg/kg kaempferol group (Fig. 1C). Consistent with the ALT and AST
activities, the liver histopathology in the 20 mg/kg kaempferol
group showed increased hepatocyte injury, similar to that of the
D-GalN/LPS group, but mice in the groups treated with 2.5 and 5
mg/kg kaempferol showed decreased hepatocyte injury, and the 5
mg/kg kaempferol group exhibited the lowest degree of injury
(Fig. 1B).
Next, the protective effects of kaempferol (5 mg/kg)
pretreatment on liver injury in mice 2, 4, 6 and 8 h after
D-GalN/LPS administration was determined. The results demonstrated
that the D-GalN/LPS-induced increases in sALT and sAST levels were
significantly decreased by 5 mg/kg kaempferol after 4, 6 and 8 h
(Fig. 1D). These results suggested
that high doses of kaempferol can induce more severe injury,
whereas pretreatment with low doses of kaempferol significantly
increased the survival rates of mice and protected against ALF
induced by D-GalN/LPS.
Effects of kaempferol on liver
inflammation in mice with D-GalN/LPS-induced ALF
Our previous study revealed that the inflammatory
response served an important role in ALF (27). Therefore, whether liver inflammation
was induced by the different doses of kaempferol in the
D-GalN/LPS-induced ALF mice was next determined. As shown in
Fig. 2A, compared with the
D-GalN/LPS group, the mice pretreated with 2.5 and 5 mg/kg
kaempferol exhibited lower gene expression levels of TNF-α, IL-6,
IL-1β, IL-12p40, C-X-C motif chemokine ligand (CXCL)-10, CXCL-2 in
the livers, and the 5 mg/kg kaempferol group exhibited the lowest
levels. By contrast, pretreatment with 10 and 20 mg/kg kaempferol
resulted in notably increased gene expression levels of these
cytokines. Additionally, pretreatment with 2.5, 5 or 10 mg/kg
kaempferol resulted in increased gene expression levels of IL-10
when compared with the D-GalN/LPS-induced mice, and the 5 mg/kg
kaempferol group exhibited the highest levels.
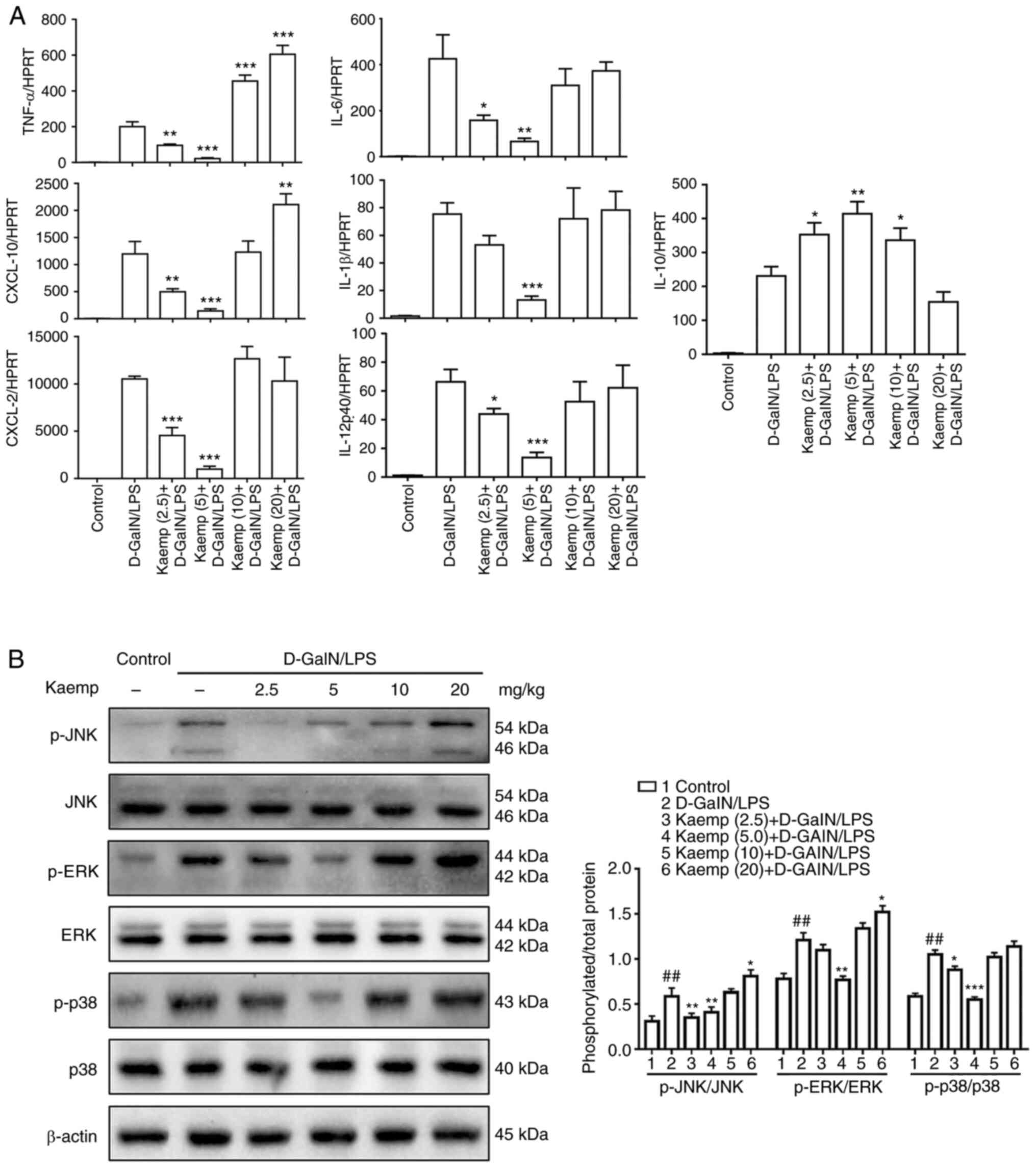 | Figure 2.Effects of different doses of kaemp
on liver inflammation in D-GalN/LPS-induced ALF. (A) Reverse
transcription-quantitative PCR analysis was performed to determine
the gene expression levels of cytokines and chemokines, including
TNF-α, IL-6, IL-12p40, IL-10, IL-1β, CXCL-10 and CXCL-2, in the
livers of mice in the different groups. (B) Western blot analysis
to determine hepatic protein expression levels of p-JNK, JNK,
p-ERK, ERK, p-p38, p38 and β-actin. Data are presented as the mean
± SD. ##P<0.01 vs. control group; *P<0.05,
**P<0.01, ***P<0.001 vs. D-GalN/LPS group. D-GalN/LPS,
D-galactosamine/lipopolysaccharide; kaemp, kaempferol; CXCL, C-X-C
motif chemokine ligand; p-, phosphorylated. |
Next, whether the MAPK signaling pathway was
affected by different doses of kaempferol was assessed. The protein
expression levels of p-JNK, JNK, p-ERK, ERK, p-p38, p38 were
determined using western blotting. Mice treated with 2.5 or 5 mg/kg
kaempferol exhibited reduced levels of these proteins, with the 5
mg/kg kaempferol group exhibiting the lowest levels. By contrast,
pretreatment with 10 and 20 mg/kg kaempferol resulted in markedly
increased expression levels of these proteins (Fig. 2B). Therefore, these results
indicated that pretreatment with high doses of kaempferol can
increase the hepatic inflammatory response, whereas pretreatment
with a low dose of kaempferol can significantly decrease it in
D-GalN/LPS-induced ALF.
Effects of kaempferol on liver
autophagy in mice with D-GalN/LPS-induced ALF
Given that our previous study observed that
autophagy serves a significant role in D-GalN/LPS-induced ALF
(18), whether different doses of
kaempferol affected liver autophagy in the D-GalN/LPS-induced ALF
was next assessed. The results of the RT-qPCR analysis revealed
that pretreatment with 2.5, 5, 10 or 20 mg/kg kaempferol
significantly increased the gene expression levels of LC3 compared
with the untreated D-GalN/LPS-induced ALF in mice (Fig. 3A). Moreover, pretreatment with 2.5,
5 and 20 mg/kg kaempferol increased the gene expression levels of
Atg7, and the 5 mg/kg kaempferol group exhibited the highest
expression levels of these genes. The western blot analysis
produced consistent results. Pretreatment with 2.5, 5, 10 or 20
mg/kg kaempferol resulted in increased LC3 protein expression, and
the 5 mg/kg kaempferol group exhibited the highest expression
levels. In addition, pretreatment with 2.5 and 5 mg/kg kaempferol
significantly decreased the protein expression levels of p62,
whereas 10 and 20 mg/kg kaempferol significantly increased its
protein expression. Furthermore, pretreatment with 2.5, 5, 10 and
20 mg/kg kaempferol significantly increased the protein expression
levels of Atg7, and the 5 mg/kg kaempferol group exhibited the
highest expression levels (Fig.
3B).
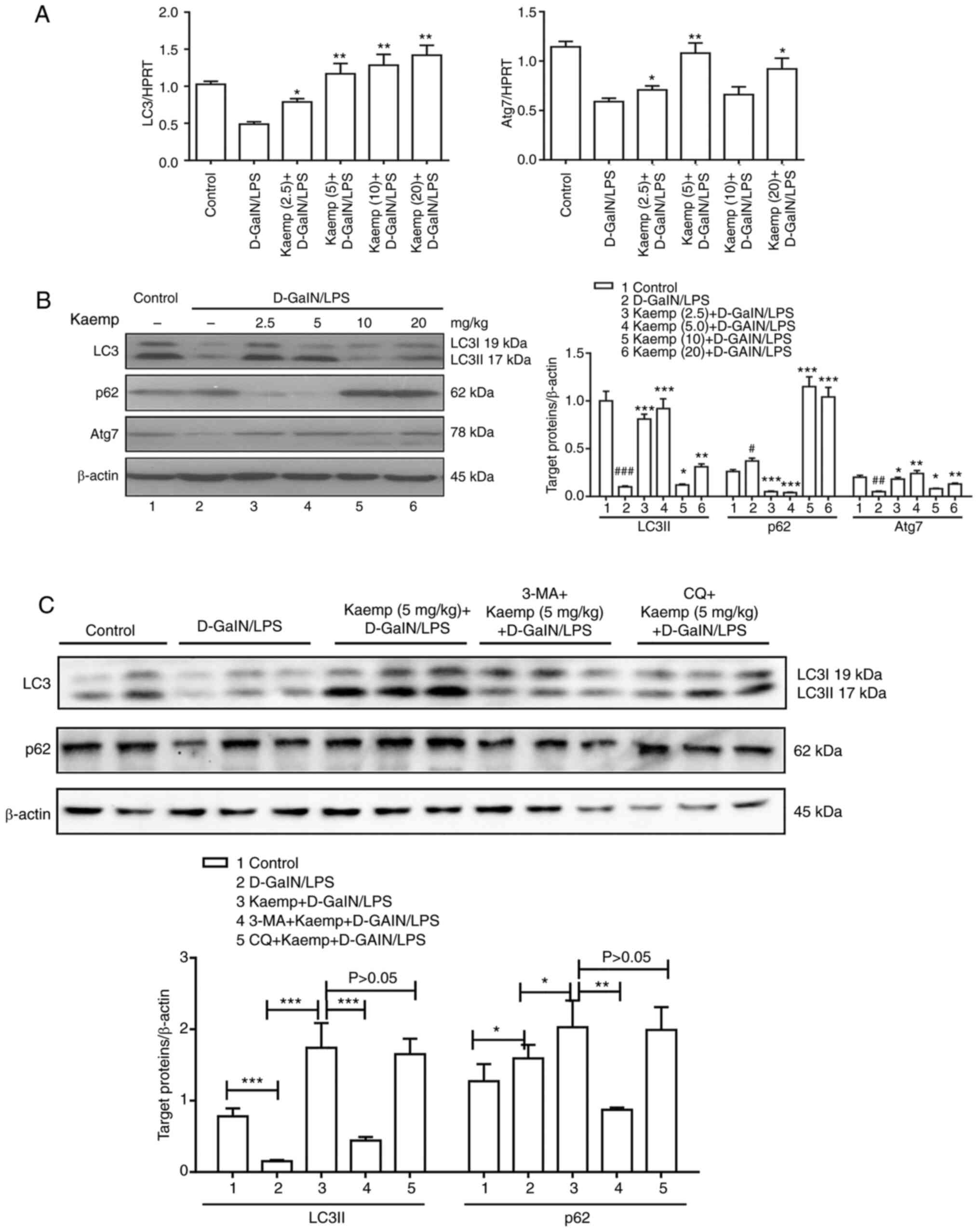 | Figure 3.Effects of different doses of kaemp
on liver autophagy in D-GalN/LPS-induced ALF. (A) Gene expression
levels of LC3 and Atg7 in livers from mice in the different groups.
HPRT was used as the reference gene. *P<0.05, **P<0.01 vs.
D-GalN/LPS group. (B) Western blotting was performed to determine
hepatic protein expression levels of LC3, p62 and Atg7, with
β-actin as the loading control. #P<0.05,
##P<0.01, ###P<0.001 vs. control group;
*P<0.05, **P<0.01, ***P<0.001 vs. D-GalN/LPS group. (C)
Mice were pretreated for 2 h with or without kaemp (5 mg/kg,
intraperitoneally), and then stimulated with D-GalN/LPS for 6 h
(n=12). Mice treated with 3-MA + kaemp + D-GalN/LPS were
co-administered 3-MA (10 mg/kg) and kaemp 2 h before D-GalN/LPS
injection (n=12). Mice treated with CQ + kaemp + D-GalN/LPS were
co-administered CQ (60 mg/kg) and kaemp 2 h before D-GalN/LPS
injection (n=12). A total of 2 h after vehicle injection, control
mice were pretreated with PBS (n=12). Western blot analysis was
performed to determine hepatic protein expression levels of LC3,
p62 and β-actin. *P<0.05, **P<0.01, ***P<0.001. Data are
presented as the mean ± SD. D-GalN/LPS,
D-galactosamine/lipopolysaccharide; 3-MA, 3-Methyladenine; CQ,
chloroquine; kaemp, kaempferol; Atg7, autophagy related 7; HPRT,
hypoxanthine guanine phosphoribosyl transferase. |
To further demonstrate the effect of kaempferol (5
mg/kg) on autophagy flux, the fusion of lysosomes with
autophagosomes was inhibited using CQ pretreatment. It was found
that CQ pretreatment did not further increase LC3II conversion and
did not further decrease p62 levels compared with the 5 mg/kg
kaempferol pretreated D-GalN/LPS induced ALF mice (Fig. 3C). These data suggest that
kaempferol pretreatment may facilitate autophagosome function and
inhibit autophagic flux in D-GalN/LPS-induced ALF.
Effects of kaempferol on
starvation-induced autophagy in vitro
To further support the results of the in vivo
experiments, the effects of kaempferol on starved primary
hepatocytes were assessed in vitro. To observe the formation
of autophagosomes, the GFP-LC3 plasmid was transfected into
hepatocytes. As shown in Fig. 4A,
compared with that of the control group, the GFP-LC3 signal was
weak in the starved group and high-dose kaempferol-treated groups,
but was bright, with a lower number of puncta, in the low-dose
kaempferol-treated groups.
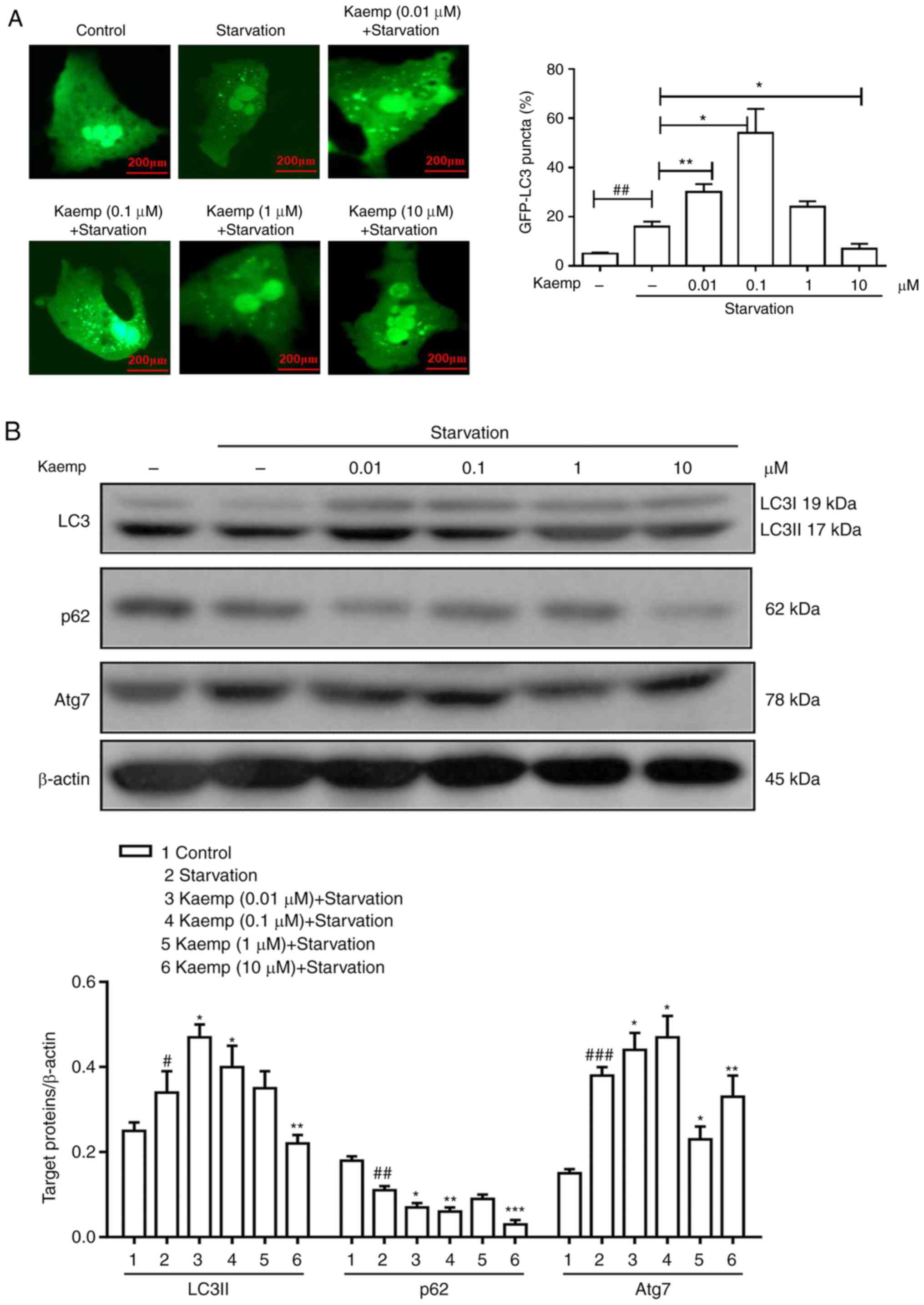 | Figure 4.Effects of different doses of kaemp
on starvation-induced autophagy in vitro. (A) Primary
hepatocytes were transfected with GFP-LC3 plasmids for 12 h and
preincubated with kaemp (0.01, 0.1, 1 or 10 µM) for 12 h, after
which, the formation of autophagosomes was observed. (B) Western
blotting was performed to determine the protein expression levels
of autophagy-related proteins, including LC3, p62 and Atg7, in
primary hepatocytes under conditions of starvation. Data are
presented as the mean ± SD. #P<0.05,
##P<0.01, ###P<0.001 vs. control group;
*P<0.05, **P<0.01, ***P<0.001 vs. starvation group. kaemp,
kaempferol; Atg7, autophagy related 7. |
Next, the protein expression levels of LC3, p62 and
Atg7 were measured via western blotting (Fig. 4B). The starved and low-dose
kaempferol-treated groups exhibited higher LC3 protein expression
levels, and the high-dose kaempferol group exhibited lower
expression levels compared with the control group. Furthermore, the
low-dose kaempferol groups showed reduced protein expression levels
of p62, and the high-dose groups also appeared to show decreased
p62 expression compared with the control. Additionally, low-dose
kaempferol groups showed reduced protein expression levels of Atg7,
and the high-dose kaempferol groups exhibited lower expression
levels compared with the control group. Collectively, the presence
of autophagic flux was demonstrated by increased expression of LC3
and decreased expression of p62. Thus, the results suggest that
pretreatment with low doses of kaempferol promotes the induction of
autophagic flux, but that the pretreatment with high doses of
kaempferol restrains the induction of autophagic flux in
starvation-induced hepatocytes in vitro.
Kaempferol ameliorates injury in the
livers of ALF model mice through autophagic mechanisms
The aforementioned experiments demonstrated that 5
mg/kg kaempferol significantly reduced ALF induced by D-GalN/LPS.
To further determine whether pretreatment with 5 mg/kg kaempferol
contributed to the induction of autophagy to protect against liver
injury, Atg7 knockdown using siRNA was performed in vivo.
The results indicated that 3-MA or Atg7 siRNA partially reversed
kaempferol-mediated hepatoprotection in the ALF model mice, as
shown by the increased levels of sALT and sAST (Fig. 5A), and the histological analysis
showing a relatively less well-preserved liver architecture
(Fig. 5B). Additionally, the
protein expression levels of LC3, p62 and Atg7 were measured using
western blotting (Fig. 5C). The
Atg7 siRNA group had lower protein expression levels of LC3II and
Atg7, and higher protein expression levels of p62. Furthermore, the
expression levels of LC3II, Atg7 and p62 were assessed following
pretreatment with 3-MA, and the expression levels of Atg7 were
reduced with pretreatment of 3-MA. The results demonstrated that,
compared with mice treated with kaempferol (5 mg/kg) and
D-GalN/LPS, 3-MA pretreatment decreased conversion of LC3II and
increased the degradation of p62 (Fig.
3C). These data indicated that pretreatment with 5 mg/kg
kaempferol ameliorated liver injury by regulating autophagy in ALF
model mice.
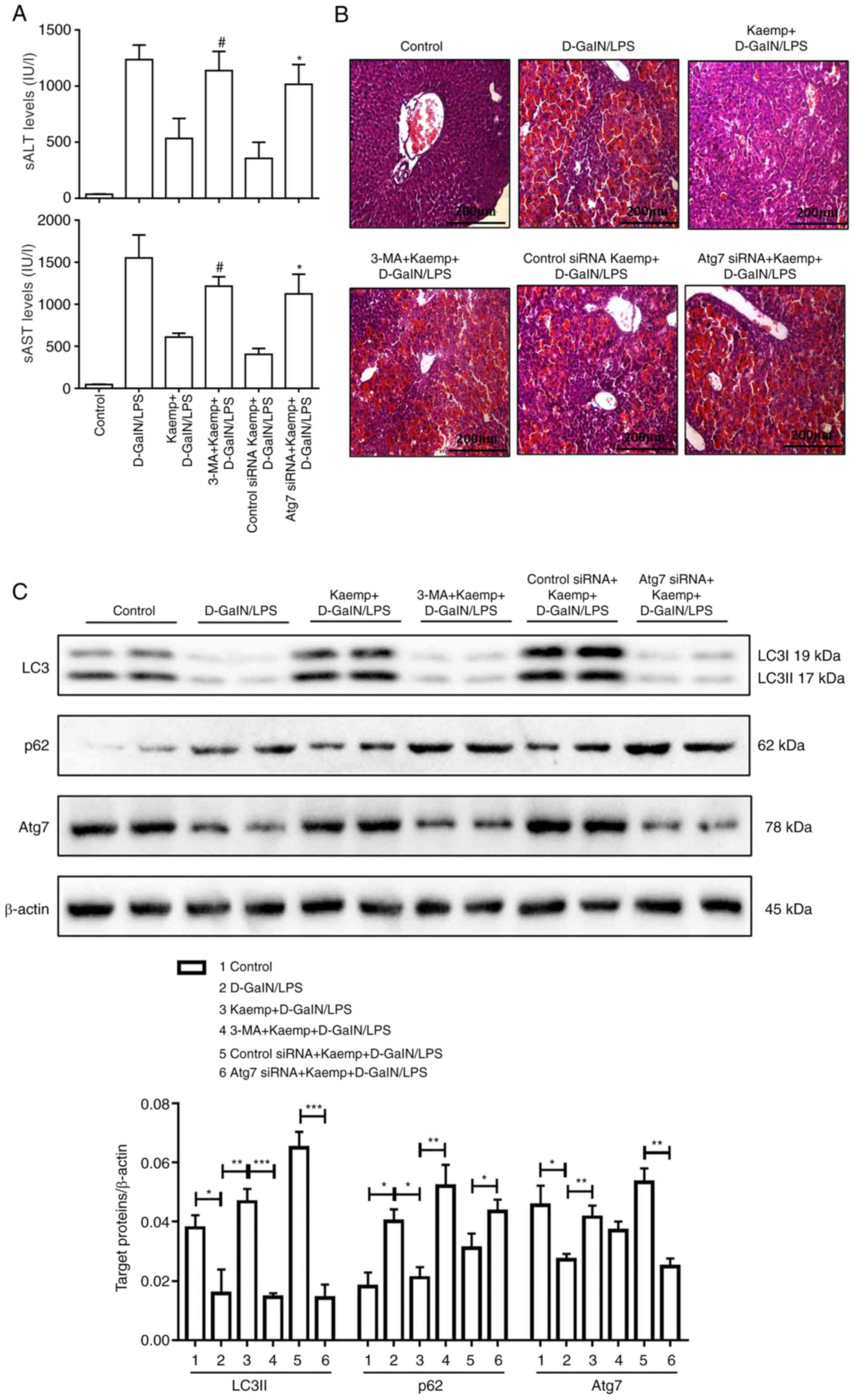 | Figure 5.Kaemp protects mice against ALF by
regulating the autophagy pathway in the liver. Mice were pretreated
for 2 h with or without kaemp (5 mg/kg, intraperitoneally), and
then stimulated with D-GalN/LPS for 6 h (n=20). Mice treated with
3-MA + kaemp + D-GalN/LPS were administered 3-MA (10 mg/kg) and
kaemp 2 h before D-GalN/LPS injection (n=20). Mice treated with
control siRNA or Atg7 siRNA + kaemp + D-GalN/LPS were injected via
tail vein for 48 h with control or Atg7 siRNA (3 mg/kg), and were
administered kaemp 2 h before D-GalN/LPS injection (n=20). A total
of 2 h after vehicle injection, control mice were pretreated with
PBS (n=20). (A) sAST and sALT enzyme levels in the different
groups. #P<0.05 vs. kaemp + D-GalN/LPS group;
*P<0.05 vs. kaemp + D-GalN/LPS + control siRNA group. (B)
Representative H&E-stained liver sections from different
groups. (C) Western blot analysis was performed to determine
hepatic protein expression levels of LC3, p62, Atg7 and β-actin in
the different groups. *P<0.05, **P<0.01, ***P<0.001. Data
are presented as the mean ± SD. ALF, acute liver failure; sALT,
serum alanine aminotransferase; sAST, serum aspartate
aminotransferase; D-GalN/LPS, D-galactosamine/lipopolysaccharide;
Atg7, autophagy related 7; 3-MA, 3-Methyladenine; kaemp,
kaempferol; siRNA, small interfering RNA. |
Discussion
Kaempferol is an ingredient in traditional Chinese
herbs, and is found in various vegetables and fruits, including
tomatoes, citrus fruits, grapefruit, onion, broccoli, cabbage and
apples. Kaempferol is a flavonoid that has been shown to possess a
broad range of pharmacological activities, such as antidiabetic,
antioxidative, cardioprotective, angiogenic and anticancer
properties (28,29). A previous study revealed that
kaempferol can suppress liver gluconeogenesis by reducing the
activity of pyruvate carboxylase and glucose-6 phosphatase, thereby
increasing hepatic glucose metabolism and insulin resistance in
diet-induced obese mice (30). In
the present study, it was demonstrated that pretreatment with
kaempferol at various doses had different functional effects on
D-GalN/LPS-induced ALF. The results indicated that 5 mg/kg
kaempferol pretreatment significantly protected against liver
injury induced by D-GalN/LPS in mice, whereas a high dose of
kaempferol decreased the survival rate of mice and resulted in more
severe injury. Thus, when using kaempferol to treat ALF, it is
important to pay attention to the differential effects caused by
different doses, and choose the appropriate dose for treatment.
ALF is associated with high mortality rates; the
clinical symptoms include coagulopathy, hepatic dysfunction and
abnormal liver biochemical parameters. In addition, ALF is closely
associated with the inflammatory response, and is an injury process
associated with inflammation-mediated hepatocellular carcinoma
(31). There are currently no
effective treatments for ALF. Our previous study showed that
endoplasmic reticulum stress can reduce inflammation by regulating
the immune mechanism in ALF (32).
The present study suggested that 10 and 20 mg/kg kaempferol
pretreatment increased the expression of proinflammatory cytokines,
and the expression levels of proinflammatory cytokines were
significantly decreased in the ALF mice pretreated with 2.5 or 5
mg/kg kaempferol. Therefore, these data suggest that a high-dose of
kaempferol pretreatment can promote the hepatic inflammatory
response, whereas low-dose kaempferol pretreatment can
significantly suppress it in D-GalN/LPS-induced ALF. The levels of
proinflammatory cytokines were downregulated by the low dose of
kaempferol, which may be the result of the increased production of
anti-inflammatory cytokines, such as IL-13, which reduce TNF-α
production in vivo (33). A
previous study revealed that IL-13 significantly reduced the lethal
effects of LPS in a neonatal mouse model of endotoxin shock
(34). In a similar manner to
IL-10, a high dose of kaempferol may also decrease the production
of IL-13 (35).
Autophagy is a process of self-digestion that
attempts to maintain cell homeostasis, supply a variety of
substrates for cellular energy generation and ensures cell survival
under stressful conditions to a certain degree. Autophagy is an
important physiological process, and is tightly associated with
regulation of cell death in specific tissues, such as the liver and
brain (36). Autophagy and
inflammation are also closely related (37). Our previous study reported that
peroxisome proliferator activated receptor (PPAR)α activation
alleviated the inflammatory response by promoting autophagy in ALF
model mice induced by D-GalN/LPS (38). Moreover, it was confirmed that
inhibition of GSK-3β activity increased PPARα expression and
decreased the inflammatory response by further increasing autophagy
(32). The present study
demonstrated that different doses of kaempferol had differential
effects on the induction of autophagy and autophagosome formation
in vivo and in vitro. The presence of autophagic flux
was identified based on the increased LC3 expression combined with
decreased p62 expression (39).
These current results indicated that a low-dose of kaempferol
upregulated the expression levels of genes associated with
autophagy, and increased LC3II conversion and p62 degradation,
whereas a high-dose of kaempferol decreased LC3II conversion and
p62 degradation, and increased autophagosome formation. Moreover,
pretreatment with CQ did not significantly alter the effects of 5
mg/kg kaempferol on the expression levels of LC3 and p62 in the ALF
mouse model. It was thus concluded that the inflammatory response
was attenuated by pretreatment with a low dose of kaempferol, and
that this low dose upregulated autophagic activity. However, it is
possible that the effects of kaempferol on the autophagic flux at
different concentrations is a highly complicated physiological
phenomenon, and therefore requires further study in the context of
ALF.
Previous studies have shown that the excessive
activation of mTOR can result in disordered apoptosis and
autophagy, which leads to the occurrence of human immunodeficiency
virus (HIV)-related diseases (40).
Hence, it could be hypothesized that kaempferol may also be useful
for conditions characterized by dysregulated autophagy, such as
HIV-associated neurodegenerative diseases. Additionally, it also
can be assumed that kaempferol may be effective against diseases
characterized by abnormal activation of the PIK3/Akt/mTOR pathway
that benefit from mTOR inhibitors, including infectious diseases,
such as HIV and severe acute respiratory syndrome coronavirus 2
(41–43), and autoimmune diseases, such as
multiple sclerosis and systemic lupus erythematosus (44–46),
as well as cancer (47–49).
In reviewing the literature, kaempferol has been
shown to antagonize Toll-like receptor (TLR) signaling, and in
particular TLR4 signaling (50). It
has previously been observed that autophagy can be regulated by the
TLR4 signaling pathway (51,52).
TLR4 inhibitors, including VGX-1027 and monoclonal antibodies, have
been reported to possess multiple functions in models of
immune-inflammation and autoimmunity, such as anti-inflammatory and
anti-injury effects, and they are being developed for use in
clinical settings (53–56). Therefore, kaempferol may be used as
an emerging class of TLR4 inhibitor to treat immune inflammation
and autoimmune diseases, which highlights the significant
pharmacological potential of kaempferol.
Although this study revealed the different effects
of different doses of kaempferol on D-GalN/LPS-induced ALF via
regulation of the autophagy pathway, the specific molecular pathway
through which autophagy is regulated remains unclear. Further
research is required to explore how autophagy is regulated.
In conclusion, the results of the present study
suggested that the effects of kaempferol on ALF at various doses
had different functional outcomes, and this was mediated by
differential regulation of the autophagy pathway. It was found that
a low dose of kaempferol can significantly protect mice from liver
injury in ALF. A schematic diagram of the potential mechanisms
identified by the present study is presented in Fig. 6. Overall, the optimal dose of
kaempferol should be further assessed, and this may serve as an
effective strategy for treatment of ALF. For further preclinical
studies of autophagy agonists, it is necessary to develop
clinically applicable therapeutic strategies for ALF.
Acknowledgements
Not applicable.
Funding
This study was supported by the National Natural
Science Foundation of China (grant nos. 81770611 and 82002243); the
Demonstrating Application and Research of Clinical Diagnosis and
Treatment Technology in Beijing (grant nos. Z191100006619096 and
Z191100006619097); Key Projects of the Beijing Municipal Education
Commission's Science and Technology Plan (grant no.
KZ202010025035); Beijing Municipal Administration of Hospitals
Clinical Medicine Development of Special Funding Support (grant no.
XMLX201830); Key Medical Major of Beijing Sailing Plan (grant no.
ZYLX201819); the National Science and Technology Key Project on
‘Major Infectious Diseases such as HIV/AIDS, Viral Hepatitis
Prevention and Treatment’ (grant nos. 2018ZX10301407-005-002 and
2018ZX10302205-004-004); the Beijing Talents foundation (grant no.
2018000021469G289); Beijing Hospitals Authority Youth Programme
(grant no. QML20201702); the Scientific Research Project of Beijing
Youan Hospital, Capital Medical University (grant no.
YNKTQN20180202); and the Key Public Relations Project of Capital
Health Development Scientific Research Project (grant nos.
SF2020-1-1151 and SF2021-1G-2181).
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
YT and FR wrote the manuscript and performed the
experiments. YT and LX collected and analyzed the data. FR and XZ
designed the experiments. FR and XZ confirm the authenticity of all
the raw data. All authors read and approved the final
manuscript.
Ethics approval and consent to
participate
All animal experiments were reviewed and approved by
Institutional Animal Care and Use Committee of Capital Medical
University (approval no. AEEI-2020-009 on 2019/12/30).
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
European Association for the Study of the
Liver, . Electronic address: easloffice@easloffice.eu. European
Association for the Study of the Liver: EASL clinical practical
guidelines on the management of acute (fulminant) liver failure. J
Hepatol. 66:1047–1081. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Rajendran P, Rengarajan T, Nandakumar N,
Palaniswami R, Nishigaki Y and Nishigaki I: Kaempferol, a potential
cytostatic and cure for inflammatory disorders. Eur J Med Chem.
86:103–112. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Sharifi-Rad M, Fokou PVT, Sharopov F,
Martorell M, Ademiluyi AO, Rajkovic J, Salehi B, Martins N, Iriti M
and Sharifi-Rad J: Antiulcer agents: From plant extracts to
phytochemicals in healing promotion. Molecules. 23:17512018.
View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Imran M, Salehi B, Sharifi-Rad J, Aslam
Gondal T, Saeed F, Imran A, Shahbaz M, Tsouh Fokou PV, Umair Arshad
M, Khan H, et al: Kaempferol: A key emphasis to its anticancer
potential. Molecules. 24:22772019. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Mahobiya A, Singh TU, Rungsung S, Kumar T,
Chandrasekaran G, Parida S and Kumar D: Kaempferol-induces
vasorelaxation via endothelium-independent pathways in rat isolated
pulmonary artery. Pharmacol Rep. 70:863–874. 2018. View Article : Google Scholar
|
|
6
|
Torres-Villarreal D, Camacho A, Castro H,
Ortiz-Lopez R and de la Garza AL: Anti-obesity effects of
kaempferol by inhibiting adipogenesis and increasing lipolysis in
3T3-L1 cells. J Physiol Biochem. 75:83–88. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Kashafi E, Moradzadeh M, Mohamadkhani A
and Erfanian S: Kaempferol increases apoptosis in human cervical
cancer HeLa cells via PI3K/AKT and telomerase pathways. Biomed
Pharmacother. 89:573–577. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Wu P, Meng X, Zheng H, Zeng Q, Chen T,
Wang W, Zhang X and Su J: Kaempferol attenuates ROS-Induced
hemolysis and the molecular mechanism of its induction of apoptosis
on bladder cancer. Molecules. 23:25922018. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Murakami T, Kohno K, Ninomiya K, Matsuda H
and Yoshikawa M: Medicinal foodstuffs. XXV. Hepatoprotective
principle and structures of ionone glucoside, phenethyl glycoside,
and flavonol oligoglycosides from young seedpods of garden peas,
Pisum sativum L. Chem Pharm Bull (Tokyo). 49:1003–1008.
2001. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Xiong Q, Fan W, Tezuka Y, Adnyana IK,
Stampoulis P, Hattori M, Namba T and Kadota S: Hepatoprotective
effect of Apocynum venetum and its active constituents. Planta Med.
66:127–133. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Matsuda H, Ninomiya K, Shimoda H and
Yoshikawa M: Hepatoprotective principles from the flowers of Tilia
argentea (linden): Structure requirements of tiliroside and
mechanisms of action. Bioorg Med Chem. 10:707–712. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
García-Mediavilla V, Crespo I, Collado PS,
Esteller A, Sánchez-Campos S, Tuñón MJ and González-Gallego J: The
anti-inflammatory flavones quercetin and kaempferol cause
inhibition of inducible nitric oxide synthase, cyclooxygenase-2 and
reactive C-protein, and down-regulation of the nuclear factor
kappaB pathway in chang liver cells. Eur J Pharmacol. 557:221–229.
2007. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Tsai MS, Wang YH, Lai YY, Tsou HK, Liou
GG, Ko JL and Wang SH: Kaempferol protects against
propacetamol-induced acute liver injury through CYP2E1
inactivation, UGT1A1 activation, and attenuation of oxidative
stress, inflammation and apoptosis in mice. Toxicol Lett.
290:97–109. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Wang H, Chen L, Zhang X, Xu L, Xie B, Shi
H, Duan Z, Zhang H and Ren F: Kaempferol protects mice from
d-GalN/LPS-induced acute liver failure by regulating the ER
stress-Grp78-CHOP signaling pathway. Biomed Pharmacother.
111:468–475. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
McEwan DG: Host-pathogen interactions and
subversion of autophagy. Essays Biochem. 61:687–697. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Jiao L, Zhang HL, Li DD, Yang KL, Tang J,
Li X, Ji J, Yu Y, Wu R, Ravichandran S, et al: Regulation of
glycolytic metabolism by autophagy in liver cancer involves
selective autophagic degradation of HK2 (hexokinase 2). Autophagy.
14:671–684. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Madrigal-Matute J and Cuervo AM:
Regulation of liver metabolism by autophagy. Gastroenterology.
150:328–339. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Ren F, Zhang L, Zhang X, Shi H, Wen T, Bai
L, Zheng S, Chen Y, Chen D, Li L and Duan Z: Inhibition of glycogen
synthase kinase 3β promotes autophagy to protect mice from acute
liver failure mediated by peroxisome proliferator-activated
receptor α. Cell Death Dis. 7:e21512016. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Mignon A, Rouquet N, Fabre M, Martin S,
Pagès JC, Dhainaut JF, Kahn A, Briand P and Joulin V: LPS Challenge
in D-galactosamine-Sensitized mice accounts for caspase-dependent
fulminant hepatitis, not for septic shock. Am J Resp Crit Care Med.
159((4 Pt 1)): 1308–1315. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Nakama T, Hirono S, Moriuchi A, Hasuike S,
Nagata K, Hori T, Ido A, Hayashi K and Tsubouchi H: Etoposide
prevents apoptosis in mouse liver with
D-galactosamine/lipopolysaccharide-induced fulminant hepatic
failure resulting in reduction of lethality. Hepatology.
33:1441–1450. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Miller S, Oleksy A, Perisic O and Williams
RL: Finding a fitting shoe for Cinderella: Searching for an
autophagy inhibitor. Autophagy. 6:805–807. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Ahlberg J, Berkenstam A, Henell F and
Glaumann H: Degradation of short and long lived proteins in
isolated rat liver lysosomes. Effects of pH, temperature, and
proteolytic inhibitors. J Biol Chem. 260:5847–5854. 1985.
View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Karmen A, Wroblewski F and Ladue JS:
Transaminase activity in human blood. J Clin Invest. 34:126–131.
1955. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Klaunig JE, Goldblatt PJ, Hinton DE,
Lipsky MM, Chacko J and Trump BF: Mouse liver cell culture. I.
Hepatocyte isolation. In vitro. 17:913–925. 1981. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Nguyen TB, Louie SM, Daniele JR, Tran Q,
Dillin A, Zoncu R, Nomura DK and Olzmann JA: DGAT1-Dependent lipid
droplet biogenesis protects mitochondrial function during
starvation-induced autophagy. Dev Cell. 42:9–21.e5. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Zhang X, Ding J, Gou C, Wen T, Li L, Wang
X, Yang H, Liu D, Lou J, Chen D, et al: Qingchangligan formula
attenuates the inflammatory response to protect the liver from
acute failure induced by d-galactosamine/lipopolysaccharide in
mice. J Ethnopharmacol. 201:108–116. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Devi KS, Mishra D, Roy B, Ghosh SK and
Maiti TK: Assessing the immunomodulatory role of heteroglycan in a
tumor spheroid and macrophage co-culture model system. Carbohyd
Polym. 127:1–10. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Zang Y, Zhang L, Igarashi K and Yu C: The
anti-obesity and anti-diabetic effects of kaempferol glycosides
from unripe soybean leaves in high-fat-diet mice. Food Funct.
6:834–841. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Alkhalidy H, Moore W, Wang A, Luo J,
McMillan RP, Wang Y, Zhen W, Hulver MW and Liu D: Kaempferol
ameliorates hyperglycemia through suppressing hepatic
gluconeogenesis and enhancing hepatic insulin sensitivity in
diet-induced obese mice. J Nutr Biochem. 58:90–101. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Bernal W and Wendon J: Acute liver
failure. N Engl J Med. 369:2525–2534. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Ren F, Zhou L, Zhang X, Wen T, Shi H, Xie
B, Li Z, Chen D, Wang Z and Duan Z: Endoplasmic reticulum
stress-activated glycogen synthase kinase 3β aggravates liver
inflammation and hepatotoxicity in mice with acute liver failure.
Inflammation. 38:1151–1165. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Zaccone P, Phillips J, Conget I, Gomis R,
Haskins K, Minty A, Bendtzen K, Cooke A and Nicoletti F:
Interleukin-13 prevents autoimmune diabetes in NOD mice. Diabetes.
48:1522–1528. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Nicoletti F, Mancuso G, Cusumano V, Di
Marco R, Zaccone P, Bendtzen K and Teti G: Prevention of
endotoxin-induced lethality in neonatal mice by interleukin-13. Eur
J Immunol. 27:1580–1583. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Chung MJ, Pandey RP, Choi JW, Sohng JK,
Choi DJ and Park YI: Inhibitory effects of
kaempferol-3-O-rhamnoside on ovalbumin-induced lung inflammation in
a mouse model of allergic asthma. Int Immunopharmacol. 25:302–310.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Russo M and Russo GL: Autophagy inducers
in cancer. Biochem Pharmacol. 153:51–61. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Fésüs L, Demény MÁ and Petrovski G:
Autophagy shapes inflammation. Antioxid Redox Sign. 14:2233–2243.
2010. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Jiao M, Ren F, Zhou L, Zhang X, Zhang L,
Wen T, Wei L, Wang X, Shi H, Bai L, et al: Peroxisome
proliferator-activated receptor α activation attenuates the
inflammatory response to protect the liver from acute failure by
promoting the autophagy pathway. Cell Death Dis. 5:e13972014.
View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Klionsky DJ, Abdelmohsen K, Abe A, Abedin
MJ, Abeliovich H, Acevedo Arozena A, Adachi H, Adams CM, Adams PD,
Adeli K, et al: Guidelines for the use and interpretation of assays
for monitoring autophagy (3rd edition). Autophagy. 12:1–222. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Nicoletti F, Fagone P, Meroni P, McCubrey
J and Bendtzen K: mTOR as a multifunctional therapeutic target in
HIV infection. Drug Discov Today. 16:715–721. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Donia M, McCubrey JA, Bendtzen K and
Nicoletti F: Potential use of rapamycin in HIV infection. Br J Clin
Pharmacol. 70:784–793. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Nicoletti F, Lapenta C, Donati S, Spada M,
Ranazzi A, Cacopardo B, Mangano K, Belardelli F, Perno C and Aquaro
S: Inhibition of human immunodeficiency virus (HIV-1) infection in
human peripheral blood leucocytes-SCID reconstituted mice by
rapamycin. Clin Exp Immunol. 155:28–34. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Fagone P, Ciurleo R, Lombardo SD,
Iacobello C, Palermo CI, Shoenfeld Y, Bendtzen K, Bramanti P and
Nicoletti F: Transcriptional landscape of SARS-CoV-2 infection
dismantles pathogenic pathways activated by the virus, proposes
unique sex-specific differences and predicts tailored therapeutic
strategies. Autoimmun Rev. 19:1025712020. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Mammana S, Bramanti P, Mazzon E, Cavalli
E, Basile MS, Fagone P, Petralia MC, McCubrey JA, Nicoletti F and
Mangano K: Preclinical evaluation of the PI3K/Akt/mTOR pathway in
animal models of multiple sclerosis. Oncotarget. 9:8263–8277. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Donia M, Mangano K, Amoroso A, Mazzarino
MC, Imbesi R, Castrogiovanni P, Coco M, Meroni P and Nicoletti F:
Treatment with rapamycin ameliorates clinical and histological
signs of protracted relapsing experimental allergic
encephalomyelitis in Dark Agouti rats and induces expansion of
peripheral CD4+CD25+Foxp3+ regulatory T cells. J Autoimmun.
33:135–140. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Ji L, Xie W and Zhang Z: Efficacy and
safety of sirolimus in patients with systemic lupus erythematosus:
A systematic review and meta-analysis. Semin Arthritis Rheum.
50:1073–1080. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Xiao W, Liu Y, Dai M, Li Y, Peng R, Yu S
and Liu H: Rotenone restrains colon cancer cell viability, motility
and epithelial-mesenchymal transition and tumorigenesis in nude
mice via the PI3K/AKT pathway. Int J Mol Med. 46:700–708. 2020.
View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Zhang Z, Li N, Liu S, Jiang M, Wan J,
Zhang Y, Wan L, Xie C and Le A: Overexpression of IFIT2 inhibits
the proliferation of chronic myeloid leukemia cells by regulating
the BCR-ABL/AKT/mTOR pathway. Int J Mol Med. 45:1187–1194.
2020.PubMed/NCBI
|
|
49
|
Liu Z, Liu H, Li Y, Wang Y, Xing R, Mi F,
Xiang C and Fu R: Adiponectin inhibits the differentiation and
maturation of osteoclasts via the mTOR pathway in multiple myeloma.
Int J Mol Med. 45:1112–1120. 2020.PubMed/NCBI
|
|
50
|
Cheng X, Yang YL, Yang H, Wang YH and Du
GH: Kaempferol alleviates LPS-induced neuroinflammation and BBB
dysfunction in mice via inhibiting HMGB1 release and
down-regulating TLR4/MyD88 pathway. Int Immunopharmacol. 56:29–35.
2018. View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Xu Y, Jagannath C, Liu XD, Sharafkhaneh A,
Kolodziejska KE and Eissa NT: Toll-like receptor 4 is a sensor for
autophagy associated with innate immunity. Immunity. 27:135–144.
2007. View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Delgado MA, Elmaoued RA, Davis AS, Kyei G
and Deretic V: Toll-like receptors control autophagy. Embo J.
27:1110–1121. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
53
|
Stojanovic I, Cuzzocrea S, Mangano K,
Mazzon E, Miljkovic D, Wang M, Donia M, Al Abed Y, Kim J, Nicoletti
F, et al: In vitro, ex vivo and in vivo immunopharmacological
activities of the isoxazoline compound VGX-1027: Modulation of
cytokine synthesis and prevention of both organ-specific and
systemic autoimmune diseases in murine models. Clin Immunol.
123:311–323. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
54
|
Fagone P, Muthumani K, Mangano K, Magro G,
Meroni PL, Kim JJ, Sardesai NY, Weiner DB and Nicoletti F: VGX-1027
modulates genes involved in lipopolysaccharide-induced Toll-like
receptor 4 activation and in a murine model of systemic lupus
erythematosus. Immunology. 142:594–602. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
55
|
Lee JC, Menacherry S, Diehl MC, Giffear
MD, White CJ, Juba R, Bagarazzi ML, Muthumani K, Boyer J, Agarwal
V, et al: Safety, bioavailability, and pharmacokinetics of
VGX-1027-A novel oral anti-inflammatory drug in healthy human
subjects. Clin Pharmacol Drug Dev. 5:91–101. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
56
|
Huang C, Pan L, Lin F, Dai H and Fu R:
Monoclonal antibody against Toll-like receptor 4 attenuates
ventilator-induced lung injury in rats by inhibiting MyD88- and
NF-κB-dependent signaling. Int J Mol Med. 39:693–700. 2017.
View Article : Google Scholar : PubMed/NCBI
|