Introduction
Non-small-cell lung cancer (NSCLC) is the commonest
lung malignant tumor worldwide, with a high incidence and mortality
rate, ranking first among all malignant tumors (1–3).
Therefore, it is necessary to further explore the function of the
new gene and its precise mechanism in the malignant progression of
lung cancer (4–6).
Circular (circ)RNAs are special non-coding RNAs that
connect the 3′ and 5′ ends to form a closed circular structure
(7,8). CircRNAs are abundant, conserved and
stabile in the cytoplasm. Increasing evidence suggests that
circRNAs may serve an important role in the occurrence and
development of cancers, including lung adenocarcinoma, and may be a
novel marker of cancer (9). The
role of circRNA in lung cancer has been the subject of a number of
studies, but as a biomarker and its mechanism of action remains to
be elucidated. Jiang et al (10) demonstrated that hsa_circ_0007385 is
involved in the occurrence and development of NSCLC and is expected
to become a potential marker and diagnosis and treatment target.
Zhang et al (11)
demonstrated that hsa_circ_0014130 is closely related to NSCLC and
can be used as a biomarker of NSCLC. Yao et al (12) confirmed that the expression level of
hsa_circ_100876 is increased in NSCLC tissues and Kaplan-Meier
survival analysis demonstrated that the overall survival rate of
patients with high expression of hsa_circ_100876 was shorter. The
results of Wei et al (13)
showed that circCD151 can promote tumor progression and immune
evasion by regulating the miR-370-3p/CXCL12 axis of melanoma.
However, the role of circCD151 in lung cancer has not been
reported.
MicroRNAs (miRNAs/miRs) are a type of non-coding RNA
with a length of ~22nt (14,15),
which directly degrade and inhibit protein synthesis and regulate
post-transcriptional gene expression level through complete or
incomplete complementary binding with target mRNA 3′UTR (16). miRNAs have been shown to regulate a
variety of physiological and pathological processes, such as cell
differentiation, cell proliferation and tumor formation (17,18).
Some miRNAs can participate directly in the formation of human
tumors, such as lung cancer, craniocerebral tumor, liver cancer,
colorectal cancer and lymphoma. miRNAs can be used as both
oncogenes and tumor suppressor genes to participate in multiple
signaling pathways of human tumor formation (19,20).
Therefore, the study of specific miRNA function provides a new
direction for tumor treatment and prevention (21).
A number of studies have found that abnormal
activation of the Hh signaling pathway is closely related to the
occurrence of liver cancer and the maintenance of malignant
biological characteristics (22–24).
Glioma cancer related gene homologous protein 2 (GLI2) is one of
the signal pathways of terminal transcription factors. GLI2
transfers extracellular Hh to the cell nucleus, which, combined
with the downstream gene promoter regions, starts the transcription
of target genes, including Gli1; the Hh signaling pathway serves a
very important role (25,26).
The purpose of the present study was to investigate
the expression of circCD151 in lung cancer tissues and cells and
its effect on proliferation, migration and invasion of lung cancer
cells. The regulatory effect of circCD151 on miR-30d-5p/GLI2 axis
was also investigated.
Materials and methods
Clinical patient information
Samples of 20 cases of lung adenocarcinoma and
normal tissues around the carcinoma (>5 cm away from the tumor)
were collected from October 2020 to January 2021. All specimens
were taken from hospital thoracic surgery lobectomy and confirmed
by postoperative pathology for the patients with lung
adenocarcinoma (~I–III A period). There were 10 males and 10
females, aged ~51–78 (63.11±6.31) years. Cancer staging was based
on the American Cancer Federation's Cancer Staging Guidelines, 8th
edition (27,28). The specimens were immediately frozen
in liquid nitrogen and stored at −80°C. None of the patients had
received radiation or chemotherapy before surgery and had no
history of malignant tumor. The present study was approved by the
medical Ethics Committee of Tianjin First Central Hospital
(approval no. TJ202008096). All patients signed informed
consent.
Cell culture
Human normal lung epithelial cell BEAS-2B, lung
cancer cell lines A549, NCI-H460, NCI-H292, 95-D were purchased
from the American Type Culture Collection. All cells were
subcultured in RPMI-1640 medium (Gibco; Thermo Fisher Scientific,
Inc.). RPMI-1640 was supplemented with 10% fetal bovine serum
(Gibco; Thermo Fisher Scientific, Inc.), 100 U/ml penicillin and
100 mg/l streptomycin (Gibco; Thermo Fisher Scientific, Inc.).
Human normal bronchial epithelial cells BEAS-2B were subcultured in
DMEM (Gibco; Thermo Fisher Scientific, Inc.) supplemented with 10%
fetal bovine serum, 100 U/ml penicillin and 100 mg/l streptomycin.
Incubation was at 37°C and 5% CO2. The fresh medium was
changed every 2–3 days and the confluence of cells was >75% for
cell passage.
Cell transfection
Plasmid transfection was performed with
Lipofectamine® 2000 according to the manufacturer's
protocol (Invitrogen; Thermo Fisher Scientific, Inc.). Transfection
was initiated when cell confluence reached 80%. A549 and NCI-H292
cells were divided into two groups: One group was transfected with
2.5 µg pcDNA.3.1 empty vector and the other was transfected with
2.5 µg pcDNA.3.1 circCD151-expression vector (Guangzhou Geneseed
Biotech Co., Ltd.). Following 24 h of transfection at 37°C with 5%
CO2, the cells were subcultured at a dilution ratio of
1:5. After transfection for 48 h, the transfection efficiency was
determined by reverse transcription-quantitative PCR (RT-qPCR).
Follow-up experiments were performed 48 h after transfection.
miRNAs and small interfering RNAs (siRNAs/si) were
purchased from Shanghai GenePharma Co. Ltd. RNAiMAX transfection
reagent (Thermo Fisher Scientific, Inc.), and 50 pmol
miR-mimics-negative control (NC; 5′-UUGUACUACACAAAAGUACUG-3′), 50
pmol miR-30d-5p mimics (5′-UGUAAACAUCCCCGACUGGAAG-3′), 50 pmol
miR-inhibitor-NC (5′-CAGUACUUUUGUGUAGUACAA-3′), 50 pmol miR-30d-5p
inhibitor (5′-CUUCCAGUCGGGGAUGUUUACA-3′) were mixed separately and
subsequent operations were carried out according to the kit
instructions to transfect miRNAs into the cells. Following 48 h of
cell transfection, the transfection efficiency was determined using
RT-qPCR and subsequent experiments were performed.
The siRNAs were transfected into cells following the
same method; that is, 50 pmol si-NC (5′-UUCUCCGAACGUGUCACGU-3′), 50
pmol si-circCD151 (5′-CACTTGTAGAGCAGAATTC-3′) or 50 pmol si-GLI2
(5′-GACAUGAGCUCCAUGCUCA-3′) were transfected into cells using
RNAiMAX. Follow-up experiments were performed 48 h after
transfection.
Separating the nucleus and
cytoplasm
The nucleus and cytoplasm were separated using a
nuclear/cytoplasmic separation kit (Thermo Fisher Scientific,
Inc.). Cells (5×106) were taken and centrifuged at 500 ×
g at 4°C for 2–3 min to collect the cells. The cells were washed
twice with cold PBS. Cold extract A (200 µl) was added to the cell
precipitation, mixed well and agitated over ice for 30 min then
centrifuged for 5 min at 1,200 × g at 4°C. The supernatant was
aspirated into another clean precooled centrifuge tube to obtain
the cytoplasmic component. The precipitate (nuclear component) was
washed once with PBS, then centrifuged at 4°C for 5 min at 2,000 ×
g and the supernatant was discarded. The precipitate was
resuspended with 200 µl storage solution B and stored for later use
or directly used for experiments.
CCK 8 experiments
A549 and NCI-H292 cell suspension was inoculated in
96-well plates with 5 multiple wells, ~3,000 cells/well/100 µl in
each group. The culture plates were placed in an incubator for
pre-culture (37°C, 5% CO2). CCK8 detection reagent (10
µl) was added to each well and incubated in an incubator for 4 h at
37°C with 5% CO2. The absorbance at 450 nm was measured
with a microplate reader and the OD value of each well was
determined.
Transwell experiment
Matrigel was diluted with blank medium (1:3). The
diluted Matrigel was added to the superior compartment surface of
the membrane at the bottom of the Transwell compartment. Matrigel
was polymerized into gel at 37°C for 30 min. DMEM complete medium
(0.6 ml) containing 10% FBS was added into the Transwell chamber.
The prepared transfected cell suspension (1.0×106/ml) of
20 µl was absorbed and added to 200 µl serum-free DMEM culture.
After mixing well 200 µl was absorbed into the upper chamber, which
was incubated for 48 h in an incubator at 37°C and 5%
CO2. Any cells that had not been passed through the
upper chamber were wiped off with a cotton swab. The lower chamber
was washed with PBS and fixed with 4% paraformaldehyde for 15 min
at room temperature. After removing the fixing solution, the
chamber was cleaned with PBS once and 0.1% crystal violet was used
for staining at room temperature for 15 min. Then, the staining
solution was removed and the cells in the lower compartment were
washed with PBS for 2–3 times and left to dry naturally at room
temperature. The culture plates were placed under an inverted
microscope and 3 fields of each filter membrane were randomly
selected for examination and imaging.
RT-qPCR
Reverse transcription and RT-qPCR followed the
manufacturer's instructions. Total RNA was extracted from tissues
and cells using TRIzol® reagent (Thermo Fisher
Scientific, Inc.). RNA was reversely transcribed into cDNA using
the Prime-Script one-step RT-PCR kit (Takara Biotechnology Co.,
Ltd.). LightCycler® 480 SYBR Green I Master mix (Roche
Diagnostics) was used to detect the expression of each gene on an
ABI PRISM 7300 detection system (Applied Biosystems; Thermo Fisher
Scientific, Inc.). The primer sequences used for the qPCR were as
follows: circCD151 forward: 5′-CACTTGTAGAGCAGAATTCTC-3′, reverse:
5′-CGTTGAACTCACCCATCCTGG-3′; miR-30d-5p forward:
5′-UGUAAACAUCCCCGACUGGAAG-3′, reverse:
5′-TGTAAACATCCCCGACTGGAAGA-3′; GLI2 forward:
5′-GAGGGATCCGCCCTCACCTCCATCAAT-3′, reverse:
5′-GAGGAATTCCTAGGTCATCATGTTCAGG-3′; SOX2 forward:
5′-GCCGATGTGAAACTTTTGTCG-3′, reverse: 5′-GGCAGCGTGACTTATCCTTCT-3;
CD44 forward: 5′-TTGCAGTCAACAGTCGAAGAAG-3′, reverse:
5′-CCTTGTTCACCAAATGCACCA-3′; Oct4 forward:
5′-CTTGCTGCAGAAGTGGGTGGAGGAA-3′, reverse:
5′-CTGCAGTGTGGGTTTCGGGCA-3′; Nanog forward:
5′-AATACCTCAGCCTCCAGCAGATG-3′, reverse:
5′-TGCGTCACACCATTGCTATTCTTC-3′; U6 forward:
5′-CTCGCTTCGGCAGCACATATACT-3′, reverse:
5′-ACGCTTCACGAATTTGCGTGTC-3′; GAPDH forward:
5′-GGTATGACAACGAATTTGGC-3′, reverse 5′-GAGCACAGGGTACTTTATTG-3′. U6
and GAPDH were used as reference genes. Reverse transcription
conditions were 16°C for 30 min, 50°C for 30 min, 75°C for 15 min
and 4°C for use. PCR conditions were 10 min at 95°C, 15 sec at 95°C
and 60 sec at 60°C for 40 cycles. A total of three parallel samples
were provided for each 20 µl system. U6 RNA was used as the
internal standard for normalization. The relative expression of
gene levels were quantified using the 2−ΔΔCq method and
normalized to the internal reference gene (29).
Clone formation experiment
The low soluble point agarose solution was prepared
with distilled water. The temperature was maintained at 40°C after
autoclave sterilization. The agarose was mixed with 2X medium in a
ratio of 1:1. Then 3 ml of the mixture was added to a plate with a
diameter of 6 cm. After it cooled and solidified, the agar was
placed in the incubator (37°C) for later use. Cells of each group
were inoculated in 6-well plates with 1×103 cells per
well. Visible cell colonies were formed after ~2 weeks of culture.
After washing with PBS three times and fixing with 75% ethanol for
30 min at room temperature, the cells were stained with 1% crystal
violet for 10 min at room temperature. After PBS washing, images
were captured using an optical microscope (Nikon Corporation) in
five randomly selected fields of view Three replicates were used
for each experiment and the average of the three replicates was
calculated.
StarBase and TargetScan analysis
In this study, the StarBase database (http://starbase.sysu.edu.cn) was used to analyze the
binding sites of circCD151 and miR-30d-5p. The TargetScan 7.2
database (http://www.targetscan.org/vert_72) was used to analyze
the binding sites between miR-30d-5p and GLI2.
Double luciferase reporter gene
assay
Binding sites between circCD151 and miR-30d-5p were
predicted using StarBase. Binding sites between GLI2 and miR-30d-5p
were predicted using TargetScan. The luciferase reporter vectors,
psiCHECK2-circCD151-wild-type (WT), psiCHECK2-circCD151-mutant
(MT), psiCHECK2-GLI2-WT and psiCHECK2-GLI2-MT (all Promega
Corporation), were co-transfected with miR-30d-5p mimics and miR-NC
respectively using Lipofectamine. Following 48 h of cell
transfection, the relative luciferase activity was detected using a
Dual-Luciferase Reporter assay system (Promega Corporation). The
ratio of the luminescence intensity of Renilla luciferase to
that of firefly luciferase reflected the binding force.
Statistical analysis
The data were presented as mean ± standard deviation
and analyzed using SPSS 17.0 software (SPSS, Inc.). Each group of
experiment was repeated three times. One-way ANOVA and Tukey's post
hoc test was used to test the data of variables between groups.
Student's t-test test was used to compare the variable data between
the two groups. Paired Student's t-test was used for tumor and
adjacent normal tissues. P<0.05 was considered to indicate a
statistically significant difference.
Results
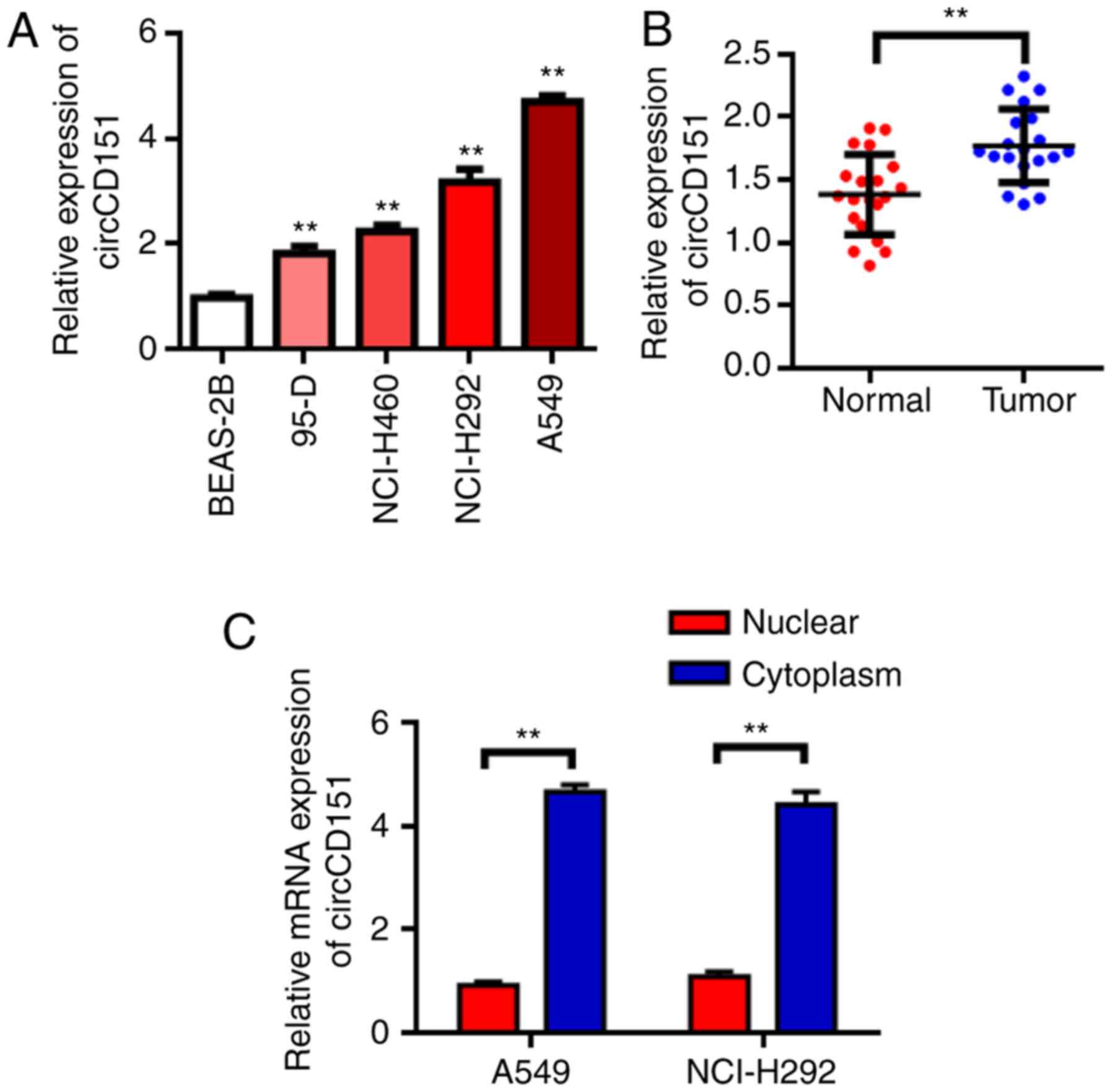
circCD151 is upregulated in NSCLC
RT-qPCR results demonstrated that circCD151 was
upregulated in NSCLC cell lines to different degrees compared with
normal lung epithelial cell BEAS-2B (Fig. 1A, P<0.05). The expression of
circCD151 was then evaluated in NSCLC tissues. The results
demonstrated that the expression of circCD151 was significantly
upregulated in NSCLC tissues compared with that in adjacent tissues
(Fig. 1B, P<0.01). These results
suggest that circCD151 may serve an oncogene role in NSCLC. Further
tests revealed that circCD151 was mainly expressed in the cytoplasm
(Fig. 1C).
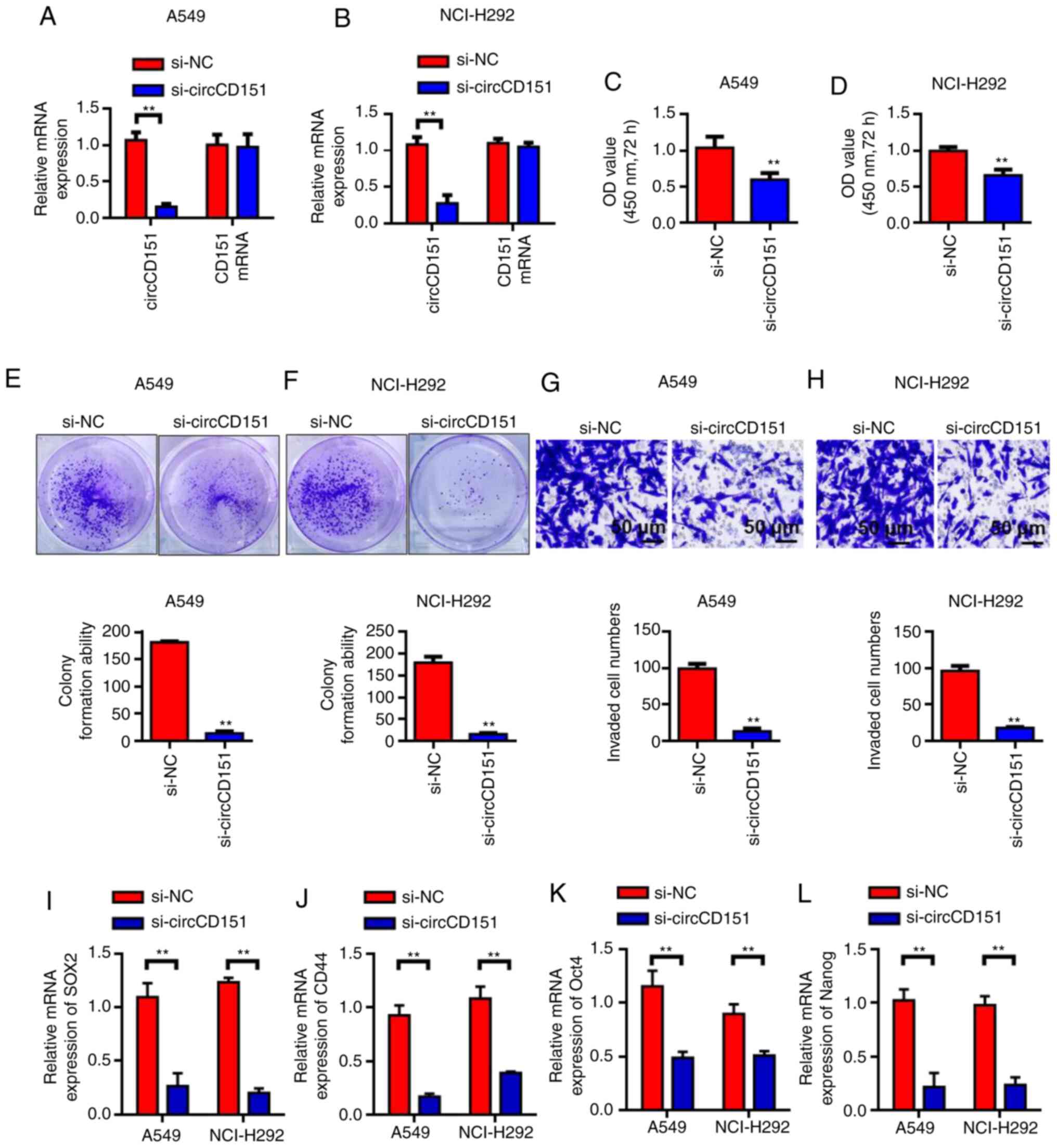
Silencing circCD151 inhibits the
activity, invasion and clonal formation of NSCLC cells
Since circCD151 was upregulated in NSCLC, siRNA weas
used to knock down circCD151 expression to evaluate its biological
function. First, RT-qPCR results demonstrated that circCD151 siRNA
inhibited A549 and NCI-H292 cells significantly (Fig. 2A and B). The results of cck-8
demonstrated that the activity of A549 and nci-h292 cells after
transfection with si-circCD151 for 72 h was significantly lower
than that of the control group (Fig. 2C
and D) (P<0.05). Colony formation test results demonstrated
that the number of colony formation in the si-circCD151
transfection group was significantly lower than that in the control
group (Fig. 2E and F; P<0.01).
Transwell was further used to evaluate the ability of cell
invasion. The results demonstrated that the number of invaded cells
in the si-circCD151 transfection group was significantly lower than
that in the control group (Fig. 2G and
H; P<0.01). RT-qPCR was used to detect the changes in the
expression levels of SOX2, CD44, OCT4 and Nanog in A549 and
NCI-H292 cells after circCD151 knockdown. Results demonstrated that
the expression levels of SOX2, CD44, OCT4 and Nanog were decreased
in A549 and NCI-H292 cells after circCD151 knockdown (Fig. 2I-L).
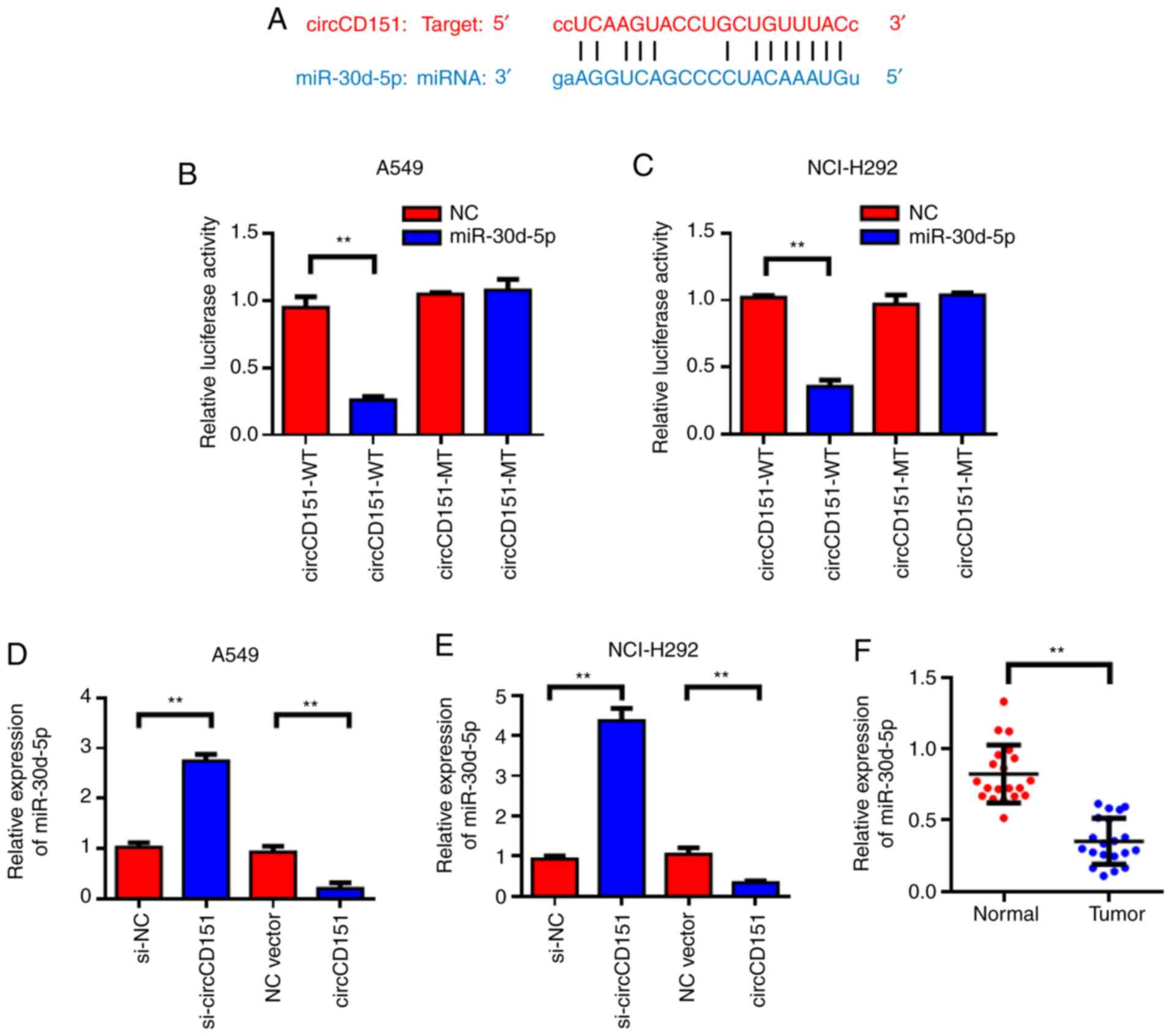
circCD151 targeted regulation of
miR-30d-5p expression
StarBase was used to predict that miR-30d-5p might
be a target gene for circCD151; the forecast sequence is
demonstrated in Fig. 3A. Results of
the dual-luciferase reporter gene demonstrated that overexpression
of miR-30d-5p could reduce the luciferase activity of wild-type
circCD151, while co-transfection of miR-30d-5p mimics and the
circCD151-MUT vector mutated at the targeted site led to loss of
the inhibitory effect of miR-30d-5p on luciferase activity
(Fig. 3B and C). The expression
level of miR-30d-5p in lung cancer cell lines after circCD151
knockdown was detected by qPCR. The results demonstrated that the
knockdown of circCD151 significantly promoted the expression of
miR-30d-5p, while the overexpression of circCD151 inhibited the
expression of miR-30d-5p (Fig. 3D and
E). Detection results of circCD151 overexpression efficiency is
presented in Fig. S1. Clinical
specimens of 20 patients with lung cancer were tested and it was
found that miR-30d-5p was lowly expressed in lung cancer tissues
compared with the para-cancer control group (Fig. 3F). Therefore, miR-30d-5p was the
direct target gene of circCD151 and circCD151 could negatively
regulate the expression of miR-30d-5p.
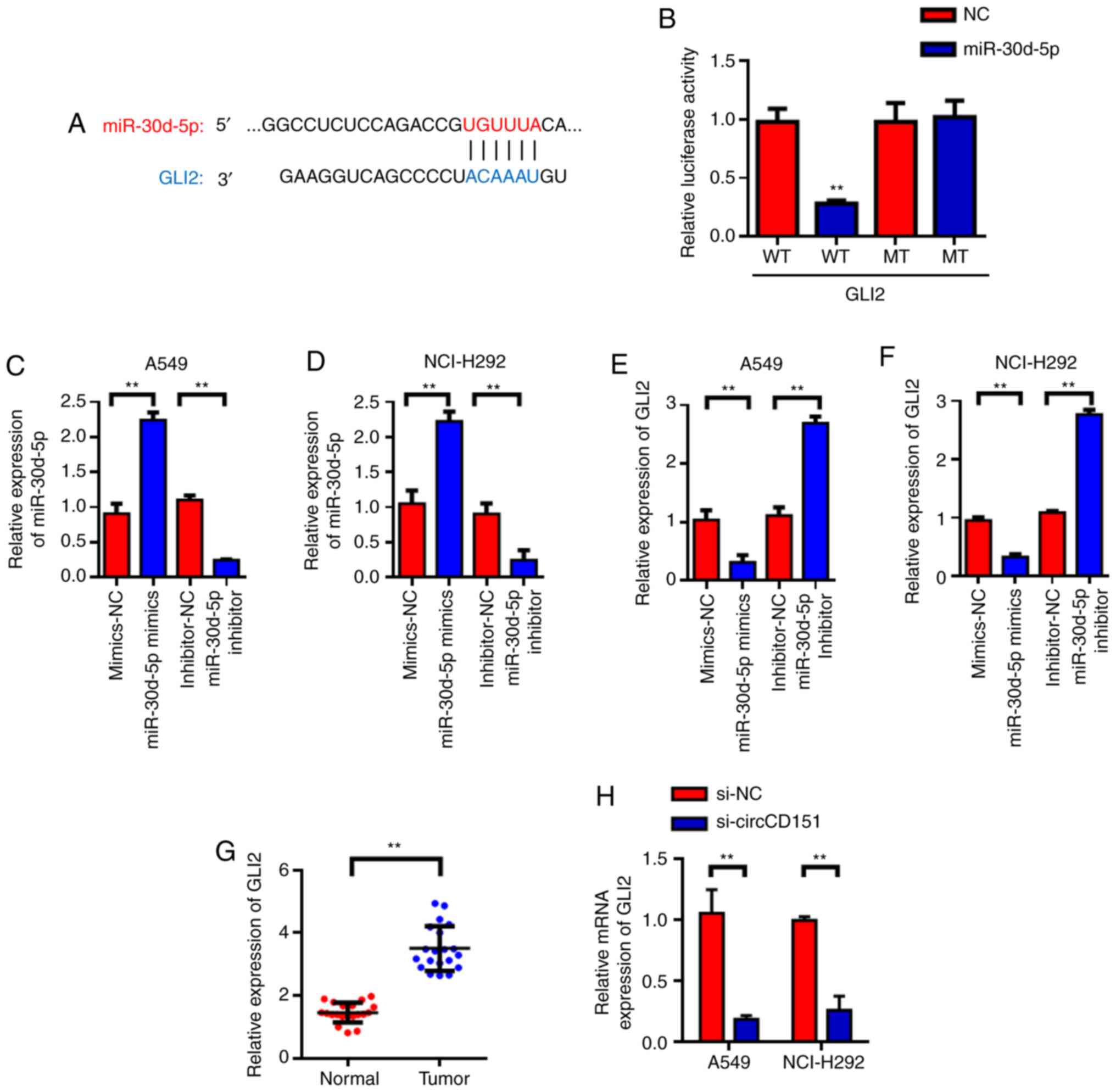
GLI2 is the target gene of
miR-30d-5p
Target genes of miR-30d-5p were predicted with
TargetScan and it was found that GLI2 was the candidate target gene
of miR-30d-5p (Fig. 4A).
Subsequently, luciferase reporter gene validation experiments were
used to find that miR-30d-5p can inhibit luciferase activity of
wild type GLI2, but had no inhibitory effect on mutant GLI2
(Fig. 4B). Further investigation
revealed that miR-30d-5p could negatively regulate the expression
of GLI2. RT-qPCR was used to detect the effects of miR-30d-5p
knockdown on the expression of GLI2 mRNA and the results
demonstrated that miR-30d-5p knockdown could significantly promote
the expression level of GLI2 in lung cancer cells. The
overexpression of miR-30d-5p inhibited the expression of GLI2
(Fig. 4C-F). Then, 20 lung cancer
patients were tested and it was found that GLI2 was highly
expressed in lung cancer tissues (Fig.
4G) compared with the para-cancer control group. The above
experimental results demonstrated that GLI2 was the target gene of
miR-30d-5p and miR-30d-5p could negatively regulate the expression
of GLI2. In addition, RT-qPCR was used to detect the changes in the
expression of GLI2 in A549 and NCI-H292 cells after circCD151
knockdown. The results demonstrated that GLI2 expression was
decreased in A549 and NCI-H292 cells after circCD151 knockdown
(Fig. 4H).
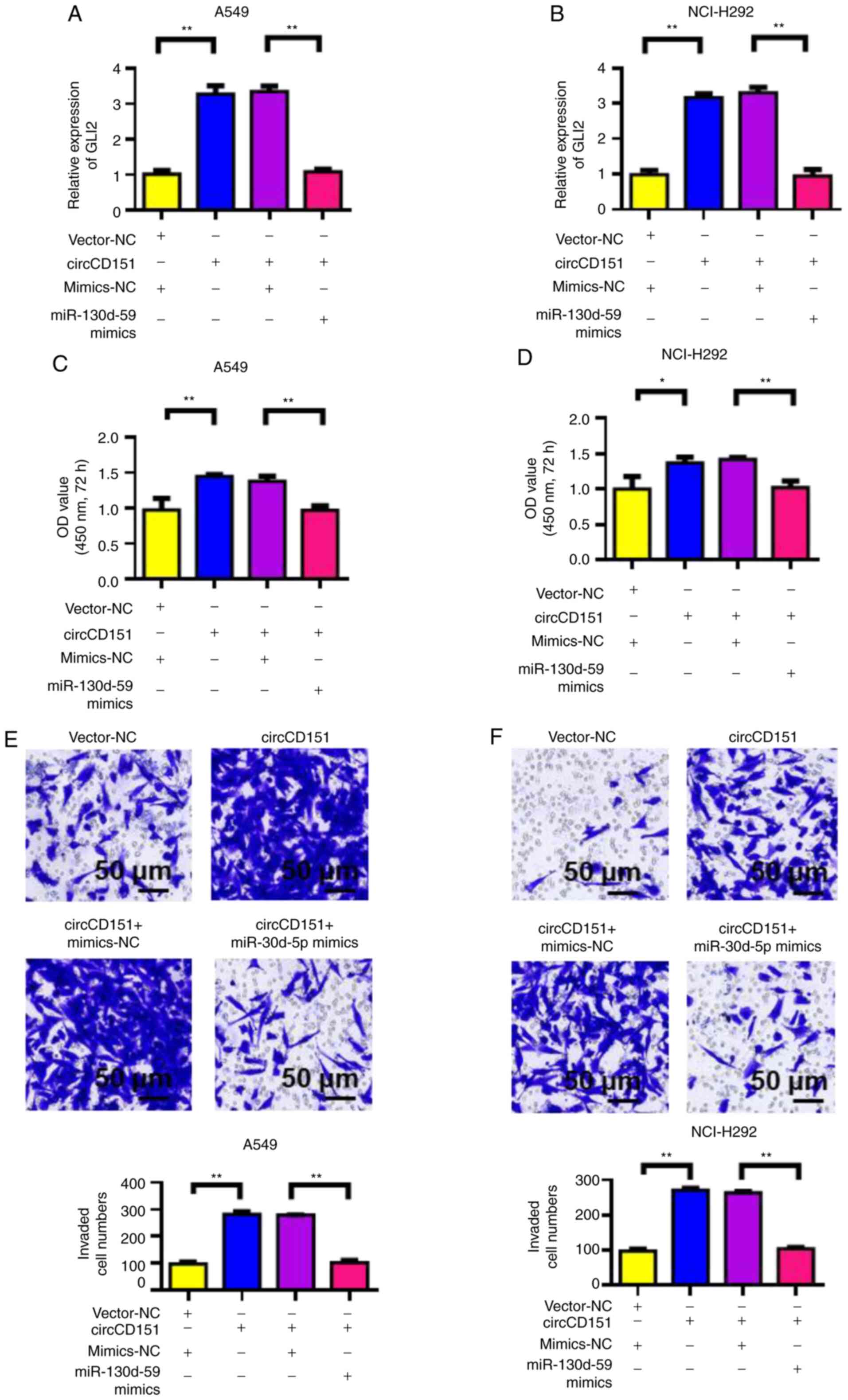
circCD151 promotes malignant
progression of lung cancer through miR-30d-5p/GLI2 molecular
axis
First, RT-qPCR was used to detect the influence of
circCD151 overexpression or circCD151+miR-30d-5p overexpression on
GLI2 expression. The results demonstrated that the overexpression
of circCD151 upregulated GLI2 expression in A549 and NCI-H292 cells
compared with the control group. However, following transfection
with circCD151+miR-30d-5p mimics, the expression of GLI2 was
significantly lower compared with that of circCD151 transfection
only (Fig. 5A and B). At the same
time, CCK-8 was used to detect the proliferation ability of cells.
The results demonstrated that overexpression of circCD151
significantly upregulated the proliferation of A549 and NCI-H292
cells. However, after the simultaneous overexpression of
circCD151+miR-30d-5p, the cell proliferation ability decreased
(Fig. 5C and D). Transwell results
demonstrated that overexpression of circCD151 significantly
upregulated the invasion ability of A549 and NCI-H292 cells.
However, after the simultaneous overexpression of
circCD151+miR-30d-5p, the cell invasion ability was reduced
(Fig. 5E and F). Therefore, the
overexpression of circCD151 promoted the malignant progression of
lung cancer by targeting miR-30d-5p and upregulating GLI2.
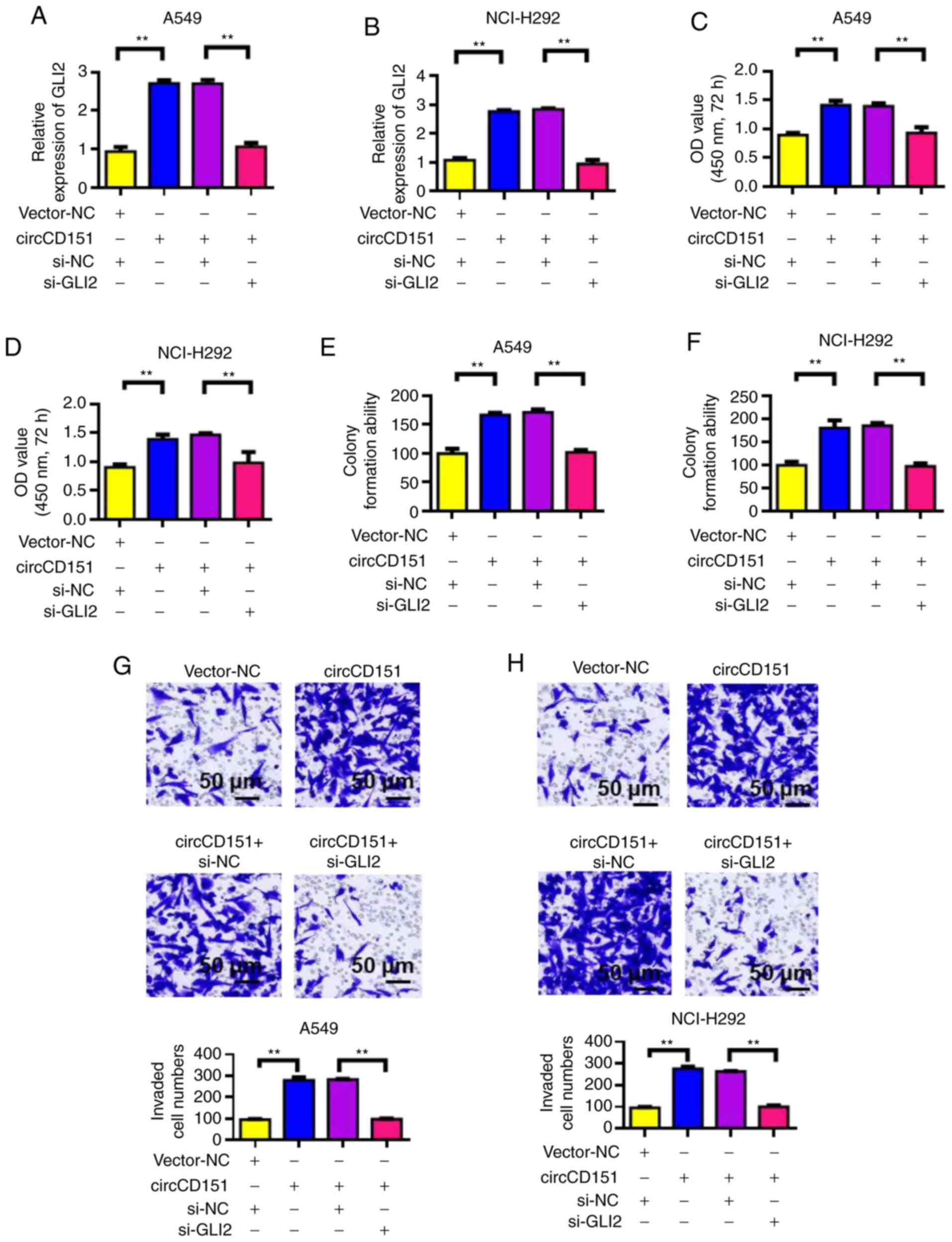
GLI2 knockdown reverses the oncogenic
activity of circCD151
Verification results of siRNA GLI2 knockdown
efficiency is presented in Fig.
S2. After the overexpression of circCD151 or circCD151+siGLI2
was detected by RT-qPCR, the expression level of GLI2 was observed
to change. The results demonstrated that overexpression of
circCD151 could upregulate GLI2 expression. However, following
transfection with circCD151+si-GLI2, the expression of GLI2 was
significantly decreased (Fig. 6A and
B). The proliferation of cells was then tested by CCK-8. The
results demonstrated that overexpression of circCD151 significantly
upregulated the proliferation of A549 and NCI-H292 cells. However,
after circCD151+si-GLI2 was overexpressed at the same time, the
proliferation ability of the cells decreased (Fig. 6C and D). Results of clone formation
experiments demonstrated that overexpression of circCD151 could
increase the clonal formation capacity of A549 and NCI-H292 cells.
However, after the simultaneous overexpression of
circCD151+si-GLI2, the cell clonal formation ability decreased
(Fig. 6E and F). Transwell results
demonstrated that overexpression of circCD151 significantly
upregulated the invasion ability of A549 and NCI-H292 cells.
However, after simultaneous overexpression of circCD151+si-GLI2,
cell invasion ability decreased (Fig.
6G and H). Therefore, GLI2 knockdown reduced the oncogenic
activity of circCD151.
Discussion
CircRNAs, as competitive endogenous RNAs or miRNAs
sponges, regulate variable splicing or transcription and the
expression of parental genes (30).
As miRNAs sponges, circRNAs may serve a major role in the
progression of lung cancer by regulating the expression of
oncogenes and/or tumor suppressor genes (31). However, little is known about the
molecular mechanism of the positive or negative relationship
between circRNAs and miRNAs in lung cancer (32,33).
The role of circRNA in lung cancer has been widely
studied, but as a biomarker its mechanism of action still need to
be further clarified (34,35). Zhu et al (36) first used microarray chip technology
to screen circRNA in lung adenocarcinoma tissue and found that the
expression of nearly 60 circRNA molecules is changed. It was
further confirmed that the upregulation of hsa_circ_0013958
expression is closely related to the proliferation and metastasis
of cancer cells. Zhao et al (37) screened 357 differentially expressed
circRNAs by sequencing and analysis of tumor samples and adjacent
normal tissues of 4 patients with early lung adenocarcinoma.
The present study found that circCD151 was
upregulated to different degrees in lung cancer tissues and cell
lines by RT-qPCR. circCD151 may serve the role of oncogene in lung
cancer and is a potential tumor marker. In this study, we focused
on the biological function of the novel circCD151 in lung cancer.
The reasons for selecting circCD151 in this study are based on a
series of analysis results. The present study used StarBase to
predict that circCD151 could combine with miR-30d-5p. GLI2 was
further predicted to be the target gene of miR-30d-5p by using
TargetScan. Finally, dual luciferase reporter gene assay confirmed
the existence of the ceRNA regulation mechanism of
circCD151/miR-30d-5p/GLI2 in lung cancer.
The present study found that circCD151 was
downregulated following knockdown, suggesting that circCD151 may be
involved in the malignant progression of lung cancer by regulating
GLI2. Furthermore, circCD151 indirectly regulated GLI2 expression
through the adsorption of miR-30d-5p. Studies have shown that
microRNA serves an important role in the occurrence and development
of a variety of tumors, including the invasion and metastasis of
tumors (38–40). One of them, miR-30d, has been shown
to have a significant anticancer effect. In the tumor tissues of
patients with esophageal squamous cell carcinoma, the expression of
miR-30d is significantly lower compared with normal tissues.
Following the overexpression of miR-30d in tumor cells, the growth,
migration and invasion ability of cells were significantly reduced.
In addition, miR-30d can also inhibit PI3K/Akt and MEK/ERK
signaling pathways, resulting in corresponding biological functions
(41–45). In the present study, miR-30d-5p was
found to have a tumor suppressive effect. Overexpression of
miR-30d-5p could reverse the oncogenic effect of circCD151.
GLI2 is a key factor in Hh signaling mediated tumor
proliferation, apoptosis resistance, angiogenesis and invasion and
metastasis (46). Studies have
shown that by interfering GLI2 expression and downregulating
downstream target gene transcription with RNAi technology, the
proliferation and metastasis of liver cancer, prostate cancer and
basal cell carcinoma can be inhibited (47–49).
In the present study, RT-qPCR analysis demonstrated that the
expression levels of GLI2 mRNA in lung cancer tissues were
significantly higher compared with adjacent tissues. Knockdown of
GLI2 reversed the cancer-promoting effect of circCD151. Cyclin D1
is a key protein regulating G1/S phase transformation,
which can cause DNA replication and division (50,51).
Studies have shown that the expression of CyclinD1 and Bax
decreases and the expression of Bcl-2 increases following GLI2 gene
silencing (52,53). It was hypothesized that the
expression of cyclin D1 was decreased after GLI2 silencing and the
tumor cells had cycle arrest, which might be in the G1
phase, leading to prolonged cell doubling time. The cell cycle was
prolonged and the ability to proliferate was decreased. The
increased ratio of anti-apoptotic Bcl-2 to pro-apoptotic Bax
induces downstream apoptotic cascade effect, promotes cell
apoptosis and ultimately leads to weakened cell growth ability
(54).
The findings of the present study and the
investigation of the function of circCD151, may provide further
reference for the treatment of lung cancer. The advancement of gene
therapy vectors and the development of pharmaceutical engineering
are expected to develop a highly effective and feasible
intervention against circCD151. Further research directions include
the development of novel non-viral gene vectors with higher
transfection efficiency.
There are also shortcomings in the present study.
The expression of circRNA is regulated by a variety of factors. The
present study only observed that circCD151 was highly expressed in
lung cancer cells and tissues, but the upstream regulatory factor
of circCD151 remains to be elucidated. As an important pro-cancer
factor, GLI2 serves an important role in the development of lung
cancer and the downstream regulatory signaling pathway of GLI2 also
requires further study.
In summary, circCD151 was significantly increased in
lung cancer tissues and cells. Silencing the circCD151 gene in A549
and NCI-H292 cells inhibited the proliferation, migration and
invasion of A549 and NCI-H292 cells. The upregulated expression of
circCD151 could promote the proliferation, migration and invasion
of A549 and NCI-H292 cells. These results indicated that circCD151
serves an important role in the development and progression of lung
cancer. Mechanism studies showed that circCD151 promoted cancer by
regulating miR-30d-5p/GLI2 axis.
Supplementary Material
Supporting Data
Acknowledgements
Not applicable.
Funding
No funding was received.
Availability of data and materials
The datasets used and/or analyzed in the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
LZ and PJ confirm the authenticity of all the raw
data. PJ designed the experiments. LZ and HZ performed the
experiments and data analysis. LZ wrote the manuscript, with
contributions from all authors. All authors read and approved the
final manuscript.
Ethics approval and consent to
participate
The present study was approved by the medical Ethics
Committee of Tianjin First Central Hospital (approval no.
TJ202008096). All patients signed informed consent.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
Glossary
Abbreviations
Abbreviations:
|
NSCLC
|
Non-small-cell lung cancer
|
|
GLI2
|
Glioma cancer related gene homologous
protein 2
|
References
|
1
|
de Groot PM, Wu CC, Carter BW and Munden
RF: The epidemiology of lung cancer. Transl Lung Cancer Res.
7:220–233. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Barta JA, Powell CA and Wisnivesky JP:
Global epidemiology of lung cancer. Ann Glob Health. 85:82019.
View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Herbst RS, Morgensztern D and Boshoff C:
The biology and management of non-small cell lung cancer. Nature.
553:446–454. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Hirsch FR, Scagliotti GV, Mulshine JL,
Kwon R, Curran WJ Jr, Wu YL and Paz-Ares L: Lung cancer: Current
therapies and new targeted treatments. Lancet. 389:299–311. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Xi X, Liu N, Wang Q, Chu Y, Yin Z, Ding Y
and Lu Y: ACT001, a novel PAI-1 inhibitor, exerts synergistic
effects in combination with cisplatin by inhibiting PI3K/AKT
pathway in glioma. Cell Death Dis. 10:7572019. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Zhong W, Yang W, Qin Y, Gu W, Xue Y, Tang
Y, Xu H, Wang H, Zhang C, Wang C, et al: 6-Gingerol stabilized the
p-VEGFR2/VE-cadherin/β-catenin/actin complex promotes microvessel
normalization and suppresses tumor progression. J Exp Clin Cancer
Res. 38:2852019. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Salzman J, Gawad C, Wang PL, Lacayo N and
Brown PO: Circular RNAs are the predominant transcript isoform from
hundreds of human genes in diverse cell types. PLoS One.
7:e307332012. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Chen D, Ma W, Ke Z and Xie F: CircRNA
hsa_circ_100395 regulates miR-1228/TCF21 pathway to inhibit lung
cancer progression. Cell Cycle. 17:2080–2090. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Zhang HD, Jiang LH, Sun DW, Hou JC and Ji
ZL: CircRNA: A novel type of biomarker for cancer. Breast Cancer.
25:1–7. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Jiang MM, Mai ZT, Wan SZ, Chi YM, Zhang X,
Sun BH and Di QG: Microarray profiles reveal that circular RNA
hsa_circ_0007385 functions as an oncogene in non-small cell lung
cancer tumorigenesis. J Cancer Res Clin Oncol. 144:667–674. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Zhang S, Zeng X, Ding T, Guo L, Li Y, Ou S
and Yuan H: Microarray profile of circular RNAs identifies
hsa_circ_0014130 as a new circular RNA biomarker in non-small cell
lung cancer. Sci Rep. 8:28782018. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Yao JT, Zhao SH, Liu QP, Lv MQ, Zhou DX,
Liao ZJ and Nan KJ: Over-expression of CircRNA_100876 in non-small
cell lung cancer and its prognostic value. Pathol Res Pract.
213:453–456. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Wei CY, Zhu MX, Lu NH, Liu JQ, Yang YW,
Zhang Y, Shi YD, Feng ZH, Li JX, Qi FZ and Gu JY: Circular RNA
circ_0020710 drives tumor progression and immune evasion by
regulating the miR-370-3p/CXCL12 axis in melanoma. Mol Cancer.
19:842020. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Ambros V: MicroRNAs: Tiny regulators with
great potential. Cell. 107:823–826. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Xi X, Chu Y, Liu N, Wang Q, Yin Z, Lu Y
and Chen Y: Joint bioinformatics analysis of underlying potential
functions of hsa-let-7b-5p and core genes in human glioma. J Transl
Med. 17:1292019. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
de Moor CH, Meijer H and Lissenden S:
Mechanisms of translational control by the 3′ UTR in development
and differentiation. Semin Cell Dev Biol. 16:49–58. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Bartel DP: MicroRNAs: Genomics,
biogenesis, mechanism, and function. Cell. 116:281–297. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Manikandan J, Aarthi JJ, Kumar SD and
Pushparaj PN: Oncomirs: The potential role of non-coding microRNAs
in understanding cancer. Bioinformation. 2:330–334. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Esquela-Kerscher A and Slack FJ:
Oncomirs-microRNAs with a role in cancer. Nat Rev Cancer.
6:259–269. 2006. View
Article : Google Scholar : PubMed/NCBI
|
|
20
|
Zhang B, Pan X, Cobb GP and Anderson TA:
MicroRNAs as oncogenes and tumor suppressors. Dev Biol. 302:1–12.
2007. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Waldman SA and Terzic A: MicroRNA
signatures as diagnostic and therapeutic targets. Clin Chem.
54:943–944. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Sicklick JK, Li YX, Jayaraman A, Kannangai
R, Qi Y, Vivekanandan P, Ludlow JW, Owzar K, Chen W, Torbenson MS
and Diehl AM: Dysregulation of the Hedgehog pathway in human
hepatocarcinogenesis. Carcinogenesis. 27:748–757. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Patil MA, Zhang J, Ho C, Cheung ST, Fan ST
and Chen X: Hedgehog signaling in human hepatocellular carcinoma.
Cancer Biol Ther. 5:111–117. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Huang S, He J, Zhang X, Bian Y, Yang L,
Xie G, Zhang K, Tang W, Stelter AA, Wang Q, et al: Activation of
the hedgehog pathway in human hepatocellular carcinomas.
Carcinogenesis. 27:1334–1340. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Tang YA, Chen YF, Bao Y, Mahara S, Yatim
SMJM, Oguz G, Lee PL, Feng M, Cai Y, Tan EY, et al: Hypoxic tumor
microenvironment activates GLI2 via HIF-1α and TGF-β2 to promote
chemoresistance in colorectal cancer. Proc Natl Acad Sci USA.
115:E5990–E5999. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Xia L, Bouamar H, Gu X, Zeballos C, Qin T,
Wang B, Zhou Y, Wang Y, Yang J, Zhu H, et al: Gli2 mediates the
development of castration-resistant prostate cancer. Int J Oncol.
57:100–112. 2020.PubMed/NCBI
|
|
27
|
Detterbeck FC, Boffa DJ, Kim AW and Tanoue
LT: The eighth edition lung cancer stage classification. Chest.
151:193–203. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Koul R, Rathod S, Dubey A, Bashir B and
Chowdhury A: Comparison of 7th and 8th editions of the UICC/AJCC
TNM staging for non-small cell lung cancer in a non-metastatic
North American cohort undergoing primary radiation treatment. Lung
Cancer. 123:116–120. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Hu W, Bi ZY, Chen ZL, Liu C, Li LL, Zhang
F, Zhou Q, Zhu W, Song YY, Zhan BT, et al: Emerging landscape of
circular RNAs in lung cancer. Cancer Lett. 427:18–27. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Huang X, Zhang W and Shao Z: Prognostic
and diagnostic significance of circRNAs expression in lung cancer.
J Cell Physiol. 234:18459–18465. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Ma Y, Zhang X, Wang YZ, Tian H and Xu S:
Research progress of circular RNAs in lung cancer. Cancer Biol
Ther. 20:123–129. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Di X, Jin X, Li R, Zhao M and Wang K:
CircRNAs and lung cancer: Biomarkers and master regulators. Life
Sci. 220:177–185. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Wang C, Tan S, Li J, Liu WR, Peng Y and Li
W: CircRNAs in lung cancer-Biogenesis, function and clinical
implication. Cancer Lett. 492:106–115. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Braicu C, Zimta AA, Harangus A, Iurca I,
Irimie A, Coza O and Berindan-Neagoe I: The function of non-coding
RNAs in lung cancer tumorigenesis. Cancers (Basel). 11:6052019.
View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Zhu X, Wang X, Wei S, Chen Y, Chen Y, Fan
X, Han S and Wu G: hsa_circ_0013958: A circular RNA and potential
novel biomarker for lung adenocarcinoma. FEBS J. 284:2170–2182.
2017. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Zhao J, Li L, Wang Q, Han H, Zhan Q and Xu
M: CircRNA expression profile in early-stage lung adenocarcinoma
patients. Cell Physiol Biochem. 44:2138–2146. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
McGuire A, Brown JA and Kerin MJ:
Metastatic breast cancer: The potential of miRNA for diagnosis and
treatment monitoring. Cancer Metastasis Rev. 34:145–155. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
39
|
van Schooneveld E, Wildiers H, Vergote I,
Vermeulen PB, Dirix LY and Van Laere SJ: Dysregulation of microRNAs
in breast cancer and their potential role as prognostic and
predictive biomarkers in patient management. Breast Cancer Res.
17:212015. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Zhong W, Hou H, Liu T, Su S, Xi X, Liao Y,
Xie R, Jin G, Liu X, Zhu L, et al: Cartilage oligomeric matrix
protein promotes epithelial-mesenchymal transition by interacting
with transgelin in colorectal cancer. Theranostics. 10:8790–8806.
2020. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Chung SJ, Nagaraju GP, Nagalingam A,
Muniraj N, Kuppusamy P, Walker A, Woo J, Győrffy B, Gabrielson E,
Saxena NK and Sharma D: ADIPOQ/adiponectin induces cytotoxic
autophagy in breast cancer cells through STK11/LKB1-mediated
activation of the AMPK-ULK1 axis. Autophagy. 13:1386–1403. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Cheng SM, Chang YC, Liu CY, Lee JY, Chan
HH, Kuo CW, Lin KY, Tsai SL, Chen SH, Li CF, et al: YM155
down-regulates survivin and XIAP, modulates autophagy and induces
autophagy-dependent DNA damage in breast cancer cells. Br J
Pharmacol. 172:214–234. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Zhu B, Chen H, Zhang X, Pan Y, Jing R,
Shen L, Wang X, Ju S, Jin C and Cong H: Serum miR-30d as a novel
biomarker for multiple myeloma and its antitumor role in U266 cells
through the targeting of the MTDH/PI3K/Akt signaling pathway. Int J
Oncol. 53:2131–2144. 2018.PubMed/NCBI
|
|
44
|
Xu X, Zong K, Wang X, Dou D, Lv P, Zhang Z
and Li H: MiR-30d suppresses proliferation and invasiveness of
pancreatic cancer by targeting the SOX4/PI3K-AKT axis and predicts
poor outcome. Cell Death Dis. 12:3502021. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Ye C, Yu X, Liu X, Dai M and Zhang B:
MiR-30d inhibits cell biological progression of Ewing's sarcoma by
suppressing the MEK/ERK and PI3K/Akt pathways in vitro. Oncol Lett.
15:4390–4396. 2018.PubMed/NCBI
|
|
46
|
Bhateja P, Cherian M, Majumder S and
Ramaswamy B: The Hedgehog signaling pathway: A viable target in
breast cancer? Cancers (Basel). 11:11262019. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
He Y, Huang H, Jin L, Zhang F, Zeng M, Wei
L, Tang S, Chen D and Wang W: CircZNF609 enhances hepatocellular
carcinoma cell proliferation, metastasis, and stemness by
activating the Hedgehog pathway through the regulation of
miR-15a-5p/15b-5p and GLI2 expressions. Cell Death Dis. 11:3582020.
View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Thiyagarajan S, Bhatia N, Reagan-Shaw S,
Cozma D, Thomas-Tikhonenko A, Ahmad N and Spiegelman VS: Role of
GLI2 transcription factor in growth and tumorigenicity of prostate
cells. Cancer Res. 67:10642–10646. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Regl G, Neill GW, Eichberger T, Kasper M,
Ikram MS, Koller J, Hintner H, Quinn AG, Frischauf AM and Aberger
F: Human GLI2 and GLI1 are part of a positive feedback mechanism in
basal cell carcinoma. Oncogene. 21:5529–5539. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Hinz M, Krappmann D, Eichten A, Heder A,
Scheidereit C and Strauss M: NF-kappaB function in growth control:
Regulation of cyclin D1 expression and G0/G1-to-S-phase transition.
Mol Cell Biol. 19:2690–2698. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Saha A, Halder S, Upadhyay SK, Lu J, Kumar
P, Murakami M, Cai Q and Robertson ES: Epstein-Barr virus nuclear
antigen 3C facilitates G1-S transition by stabilizing and enhancing
the function of cyclin D1. PLoS Pathog. 7:e10012752011. View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Rutter M, Wang J, Huang Z, Kuliszewski M
and Post M: Gli2 influences proliferation in the developing lung
through regulation of cyclin expression. Am J Respir Cell Mol Biol.
42:615–625. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
53
|
Zhang D, Liu J, Wang Y, Chen J and Chen T:
shRNA-mediated silencing of Gli2 gene inhibits proliferation and
sensitizes human hepatocellular carcinoma cells towards
TRAIL-induced apoptosis. J Cell Biochem. 112:3140–3150. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
54
|
Singh R, Letai A and Sarosiek K:
Regulation of apoptosis in health and disease: The balancing act of
BCL-2 family proteins. Nat Rev Mol Cell Biol. 20:175–193. 2019.
View Article : Google Scholar : PubMed/NCBI
|