Introduction
Ferroptosis is an iron-dependent,
caspase-independent and non-apoptotic regulated cell death, which
is induced by excessive accumulation of lipid peroxide (1). Cell death is believed to be the result
of one of two distinct processes; programmed cell death (apoptosis,
pyroptosis, autophagy and ferroptosis) or uncontrolled cell death
(necrosis and oncosis) (2).
Ferroptosis has been distinguished from other types of cell death,
such as apoptosis, autophagic cell death and necroptosis, at the
cell morphology, genetic and biochemistry levels (1). Iron accumulation and an increase in
lipid peroxidation, determined by high levels of malondialdehyde
(MDA) content, are typical characteristics of ferroptosis (3). The upregulation of
prostaglandin-endoperoxide synthase 2 (Ptgs2), a peroxidase, is the
only downstream marker of ferroptosis (3). Ferroptosis has been reported to
participate in various pathological processes, including cancer
cell death (4), neurotoxicity
(5), acute kidneys failure
(6), hepatotoxicity (7) and intestinal ischemia/reperfusion
injury (8).
Ferroptosis can be elicited by pharmacologically
inhibiting the cysteine/glutamate antiporter, system
Xc−, in cancer cells (3). Erastin was first identified in 2003 as
a ferroptosis inducer for screening a cancer drug to be selectively
lethal to oncogenic RAS mutant cell lines (9). This compound could inhibit solute
carrier family 7 member 11 (SLC7A11), part of a heterodimer named
system Xc−. System Xc− in the cell membrane
is responsible for cellular cystine uptake to synthesize
glutathione (10). Since cysteine
is necessary for the synthesis of the antioxidant glutathione
(GSH), erastin leads to a depletion of intracellular GSH and
inactivation of glutathione peroxidase 4 (GPX4), increased lipid
reactive oxygen species (ROS) formation and peroxidation, thus
triggering ferroptosis (11).
Erastin-induced ferroptosis shows great potential for cancer
therapy (12). However, the
physiological role and pathological effect of erastin-induced
ferroptosis on healthy tissues has not been well characterized.
Erastin is already widely used to induce ferroptosis
in cancer cells in vitro (13). The aim of the present study was to
identify the side effects of erastin-induced ferroptosis on healthy
tissues in vivo. A mouse model of erastin-induced
ferroptosis was established and it was found that erastin-induced
ferroptosis caused mild cerebral infarction of the brain, duodenal
epithelium hyperplasia and glomeruli enlargement. The present study
provided some evidence of pathological changes induced by erastin
treatment and it is hoped that further studies will look at the
prevention of such side effects of erastin when used for cancer
therapy.
Materials and methods
Animals
A total of 12 male C57BL/6 mice (weight, 20–21 g;
age, 8 weeks) were purchased from SLAC Laboratory Animal Co., Ltd.
All mice were housed in standard plastic cages and contained food
and water ad libitum at Zhejiang University at 24°C with a
12-h light/dark cycle. The experimental procedures for the mice
were approved by Animal Ethics Committee of Zhejiang University
(approval no. 20077). In accordance with a previous study by Huo
et al (14), mice were
intraperitoneal injected with 25 mg/kg body weight of erastin
(Selleck Chemicals; n=6) or solvent (5% DMSO + 40% PEG400 + 5%
Tween-80 + 50% physiological saline; n=6) for 2 days at 12-h
intervals. Mice were anesthetized via an intraperitoneal injection
of avertin (500 mg/kg) 6 h after the last injection. Following
anesthesia, peripheral blood (~0.5 ml per mouse) was drawn from
retroorbital plexus once by inserting a micro-hematocrit capillary
tube (cat. no. HCH-42A2502; Kimble Chase Life Science and Research
Products LLC) into the venous sinus behind the eyeball and was used
to determine erythrocyte parameters and serum iron level by
automatic SYSMEX F820 analyzer (Sysmex Corporation). Total iron
binding capacity was calculated by measuring serum iron and serum
unsaturated iron-binding capacity and summing these values. Animals
were sacrificed following blood collection by cervical dislocation.
Tissue samples from duodenum, kidney, liver, spleen, testis and
brain were frozen in liquid nitrogen and held at −80°C for
analysis.
Reverse transcription-quantitative
(RT-q) PCR
Total RNA was isolated from intestine, kidney,
liver, spleen and testis tissues, respectively, using
TRIzol® (Invitrogen; Thermo Fisher Scientific, Inc.).
RNA quantity and purity were determined using a NanoDrop 2000
spectrophotometer (Thermo Fisher Scientific, Inc.). For cDNA
synthesis, 2,000 ng RNA was reverse transcribed using
Hifair® III 1st Strand cDNA Synthesis SuperMix for qPCR
(Shanghai Yeasen Biotechnology Co., Ltd.). qPCR was conducted on a
StepOne Real-Time PCR System (ABI StepOnePlus, Applied Biosystems;
Thermo Fisher Scientific, Inc.) using Hief UNICON® qPCR
SYBR Green Master Mix (Shanghai Yeasen Biotechnology Co., Ltd.).
The following thermocycling conditions were used for the qPCR:
Initial denaturation at 95°C for 1 min; followed by 40 cycles at
95°C for 15 sec and 60°C for 1 min. The primers were synthesized by
BioSune Biotechnology. The following primers were used: Ptgs2
forward 5′-ATACGCTGAGTGTGGTTTGC-3′ and reverse
5′-CTCACTAAACCATCCAATCGG-3′; GAPDH forward
5′-TGCGACTTCAACAGCAACTC-3′ and reverse 5′-GCCTCTCTTGCTCAGTGTCC-3′.
Ptgs2 fold changes were calculated after normalizing the change in
expression of GAPDH using 2−ΔΔCq method (15). The experiments were repeated in
triplicate.
Western blotting
Total protein was isolated from tissues by
homogenizing in RIPA lysis buffer (Beyotime Institute of
Biotechnology) containing protease inhibitor cocktail (Thermo
Fisher Scientific, Inc.). The total protein content was determined
using a BCA Protein Assay kit (Nanjing KeyGen Biotech Co., Ltd.).
Protein samples (20 µg/lane) were separated by 15% SDS-PAGE and
then transferred onto a PVDF membrane (MilliporeSigma). The
membranes were blocked with skimmed milk for 1 h at room
temperature, after which the membrane was incubated with the
following primary antibodies overnight at 4°C: GAPDH (cat. no.
EM1901-57; 1:10,000; HuaBio), GPX4 (cat. no. ET1706-45; 1:1,000;
HuaBio) and SLC7A11 (cat. no. HA720001; 1:1,000; HuaBio).
Subsequently, the membrane was washed three times with PBS and
incubated with a HRP-conjugated secondary antibody (cat. no. 7074;
1:2,000; Biosharp Life Sciences) at room temperature for 1 h.
Signals were detected using chemiluminescence (ECL Plus detection
system; Clinx Science Instruments Co., Ltd.) and quantified using
ImageJ software (version 2.0; National Institutes of Health).
Measurement of MDA and GSH in
tissues
MDA and GSH were measured using MDA and GSH Assay
kit (Beijing Solarbio Science & Technology Co., Ltd.) according
to the manufacturer's instructions. The production of MDA serves as
an index of lipid peroxidation and gives a pink color once reacted
with thiobarbituric acid with a maximum absorption at 535 nm. GSH
assay is based on the GSH recycling system by GSH substrate (DTNB)
and GSH reductase. DTNB and GSH react to generate
2-nitro-5-thiobenzoic acid which has a yellow color. Therefore, GSH
concentration can be determined by measuring absorbance at 412 nm.
Optical density was measured at 532 nm for MDA or 412 nm for GSH by
a microplate reader (Molecular Devices, LLC).
2,3,5-Triphenyltetrazolium chloride
monohydrate (TTC) staining
The brain was sectioned coronally into 5 slices (2
mm thick) and then incubated in TTC for 10 min at 37°C and fixed in
10% buffered formalin. Survival area was defined as the ratio of
TTC-stained tissue (non-ischemic) area to the entire coronal
section area.
Histological examination of
tissues
Cerebral, duodenal, renal, hepatic, splenic and
testicular tissues were fixed in 4% paraformaldehyde overnight at
4°C and embedded in paraffin blocks. Sections of 5-µm were
deparaffinized, rehydrated and stained with H&E for 1 min at
room temperature. Determination of villus height, crypt depth was
performed with ≥6 villi or crypts per slide. Glomerular volume and
mesangial area in each kidney section were quantified by measuring
10 non-overlapping glomeruli. For Prussian blue staining, cerebral,
duodenal, renal, hepatic, splenic and testicular sections were
deparaffinized and rehydrated in serial alcohols. The tissue was
then incubated for 30 min at room temperature in an equal mixture
of 2% potassium ferrocyanide and 2% hydrochloric acid. After
washing, the slides were counterstained with eosin for 20 sec,
dehydrated and sealed. For Masson staining, duodenal, renal and
splenic sections were stained by Wiegert's iron hematoxylin for 10
min at room temperature, and 1% hydrochloric acid in alcohol used
for differentiation at room temperature. Subsequently, sections
were counterstained in Masson Ponceau acid solution for 10 min and
sealed at room temperature. For Periodic acid-Schiff (PAS)
staining, renal sections were deparaffinized and rehydrated in
serial alcohols (70–100%). Sections were treated for 15 min with 1%
periodic acid followed by Schiff's reagent treatment for 10 min,
then stained with Gills hematoxylin for 3 min at room temperature.
Images were captured using an Olympus NP-26 inverted light
microscope (magnification, ×200; Olympus Corporation). The
mesangial matrix index was defined as the ratio of mesangial matrix
area divided by the tuft area.
Statistical analysis
All assays were performed at least in triplicate and
the values are expressed as mean ± standard error of the mean.
Statistical analysis was performed using GraphPad software (version
8.0; GraphPad Prism, Inc.) and analyzed by unpaired two-tailed
Student's t-tests. P<0.05 was considered to indicate a
statistically significant difference.
Results
Erastin injection induces ferroptosis
in mice
Erastin-treated mice were inactive and liked to
gather together, but exhibited no significant difference in body
weight compared with control mice (Fig. S1). Following erastin injection,
hemoglobin, hematocrit, red blood cell count and red blood cell
distribution width were all significantly (P<0.05) decreased in
the peripheral blood of mice (Table
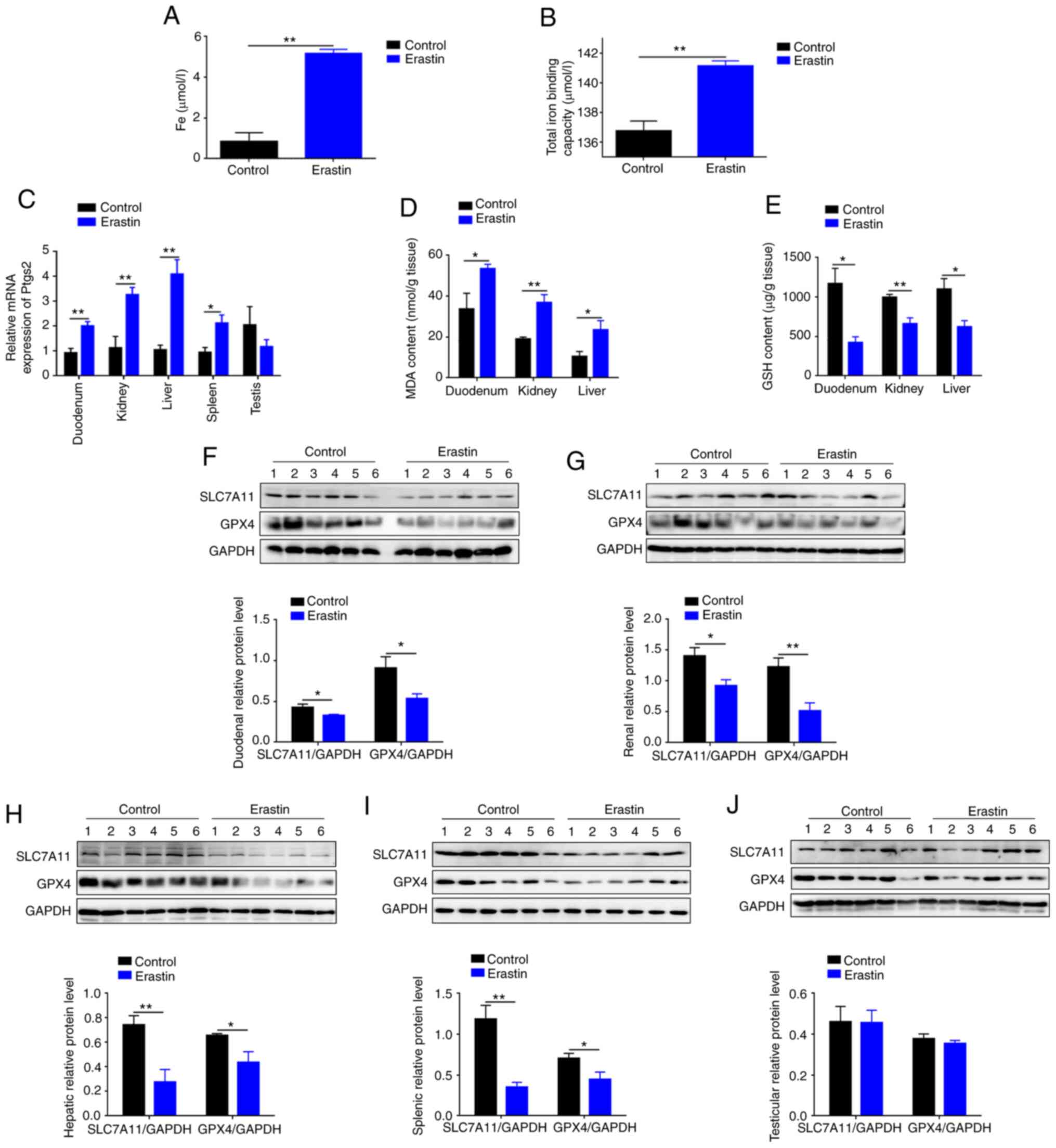
I). Serum iron of erastin-treated mice was 6.16-fold higher
(P<0.01) compared with control mice (Fig. 1A), as well as total iron binding
capacity (Fig. 1B). Erastin
injection induced a robust increase (P<0.05) in mRNA level of
Ptgs2, a putative molecular marker of ferroptosis, in
duodenum, kidney, liver and spleen (Fig. 1C). Compare with the control group,
MDA was increased by 58% in duodenum (P<0.05), 93% in kidney
(P<0.01) and 2.25-fold in liver (P<0.05) of erastin-treated
mice (Fig. 1D). Erastin injection
led to a decrease of GSH by 64% in duodenum (P<0.05), 34% in
kidneys (P<0.01) and 43% in liver (P<0.05) (Fig. 1E). Erastin injection also inhibited
SLC7A11 and GPX4 protein expression in duodenum, kidneys, liver and
spleen (Fig. 1F-I). However, both
proteins and mRNA level of Ptgs2 in testis were not significantly
(P>0.05) changed (Fig. 1C and
J). Taken together, above data indicated that erastin injection
successfully induced ferroptosis.
 | Table I.Erastin-induced ferroptosis altered
blood index values of mice. |
Table I.
Erastin-induced ferroptosis altered
blood index values of mice.
| Parameter | Control | Erastin | P-value |
|---|
| Hemoglobin
(g/l) | 164.30±2.73 | 155.50±1.56 | 0.04 |
| Hematocrit (%) | 52.85±0.80 | 49.55±0.52 | 0.02 |
| Red blood cell
count (1012/l) | 10.43±0.15 | 9.76±0.13 | 0.01 |
| Red blood cell
distribution width (%) | 16.34±0.21 | 15.30±0.15 | 0.01 |
| Mean corpuscular
hemoglobin (pg) | 15.77±0.08 | 15.95±0.05 | 0.14 |
| Mean corpuscular
hemoglobin concentration (g/l) | 311.00±0.73 | 313.80±1.11 | 0.06 |
| Mean corpuscular
volume (fl) | 50.67±0.21 | 50.78±0.23 | 0.74 |
| Platelet count
(109/l) | 959.70±56.82 | 830.50±75.40 | 0.20 |
| Mean platelet
volume (fl) | 7.13±0.07 | 7.00±0.13 | 0.34 |
| Plateletcrit
(%) | 0.68±0.04 | 0.58±0.05 | 0.16 |
| Platelet
distribution width (%) | 14.73±0.02 | 14.68±0.03 | 0.11 |
| ALT (U/l) | 37.40±2.58 | 33.80±3.54 | 0.44 |
| AST (U/l) | 135.00±9.26 | 158.00±13.94 | 0.19 |
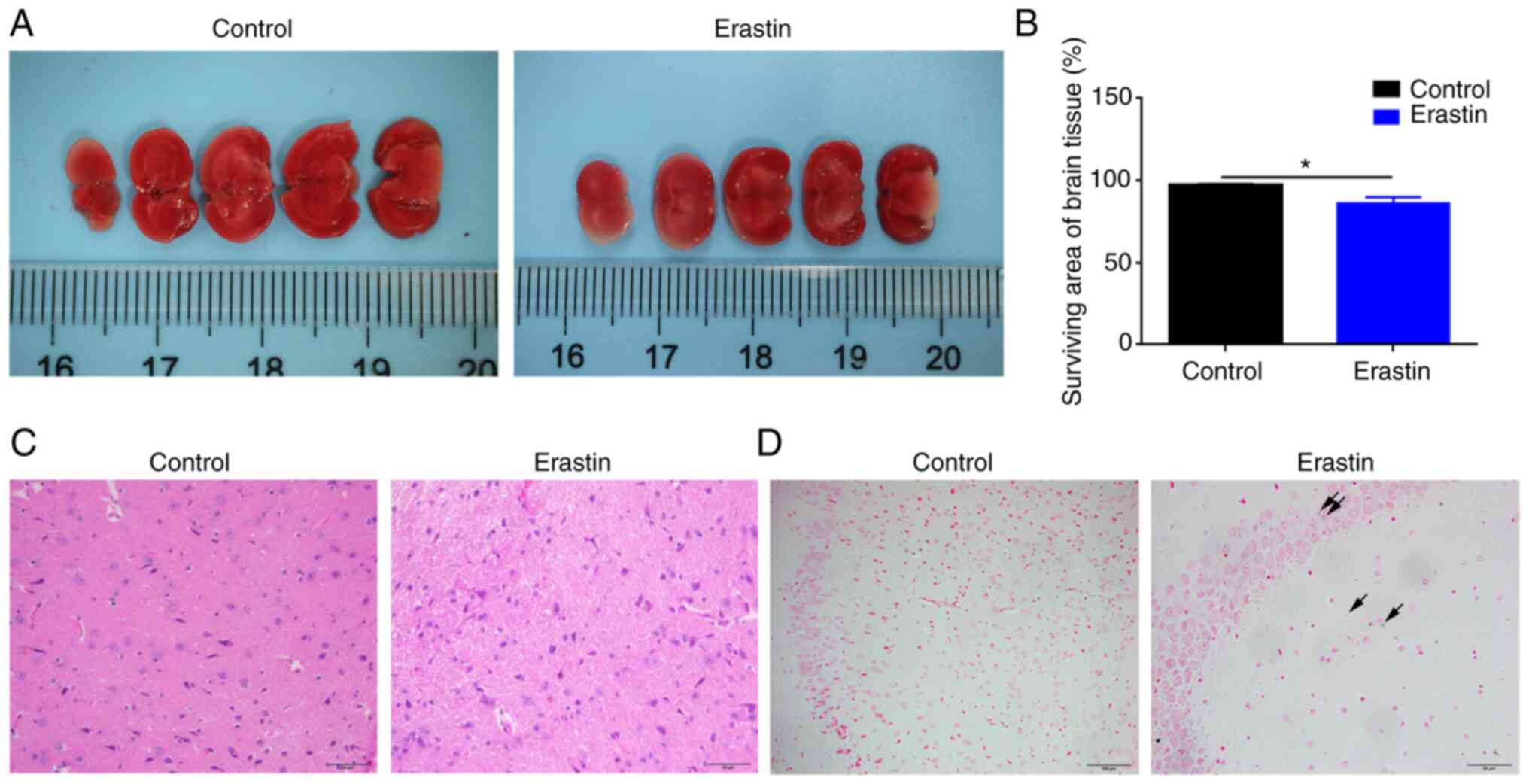
Erastin injection causes mild infarct
and iron deposition in the brain
TTC staining demonstrated that erastin injection
caused a mild cerebral infarction in brain, where living tissue is
red while infarcted area remains white (Fig. 2A). Erastin-treated mice demonstrated
11% lower (P<0.05) surviving area of brain tissue compared with
control mice (Fig. 2B). H&E
staining demonstrated that there was no difference between two
groups in brain white matter (Fig.
2C). Prussian blue staining revealed a slight iron deposition
in the brain of erastin-treated mice (Fig. 2D).
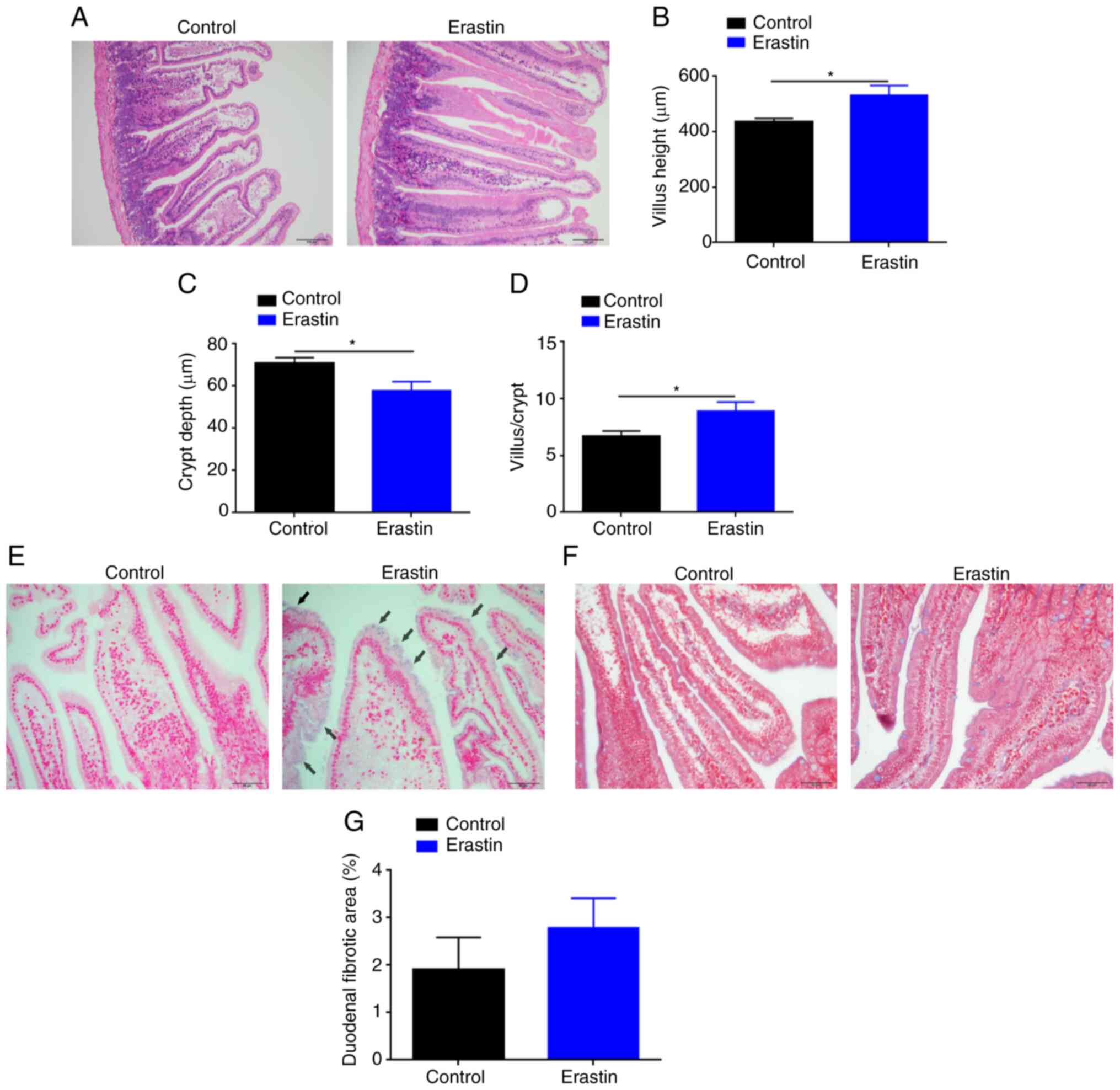
Erastin injection promotes the growth
and iron deposition of duodenum
H&E staining demonstrated that erastin-treated
mice had thicker, longer and denser mucosal villi compared with the
control mice (Fig. 3A). Image
analysis revealed a significant increment (P<0.05) in the villus
height (531.9±35.48 µm) of erastin-treated mice, as compared with
that (435.70±12.43 µm) of untreated mice (Fig. 3B). Erastin injection also induced a
marked reduction (P<0.05) in the crypt depth (57.86±4.32 µm) of
erastin-treated mice, as compared with that (70.84±2.74 µm) of
untreated mice (Fig. 3C).
Therefore, the ratio of villus height to crypt depth, an indicator
of absorptive function of duodenum, was upregulated by 33% in
erastin-treated mice (Fig. 3D).
Prussian blue staining revealed that the blue granules of
hemosiderin mainly appeared on the tip of villi in erastin-treated
mice (Fig. 3E). Masson staining
demonstrated that there was no significant difference (P>0.05)
in tissue fibrosis of duodenum between two groups (Fig. 3F and G).
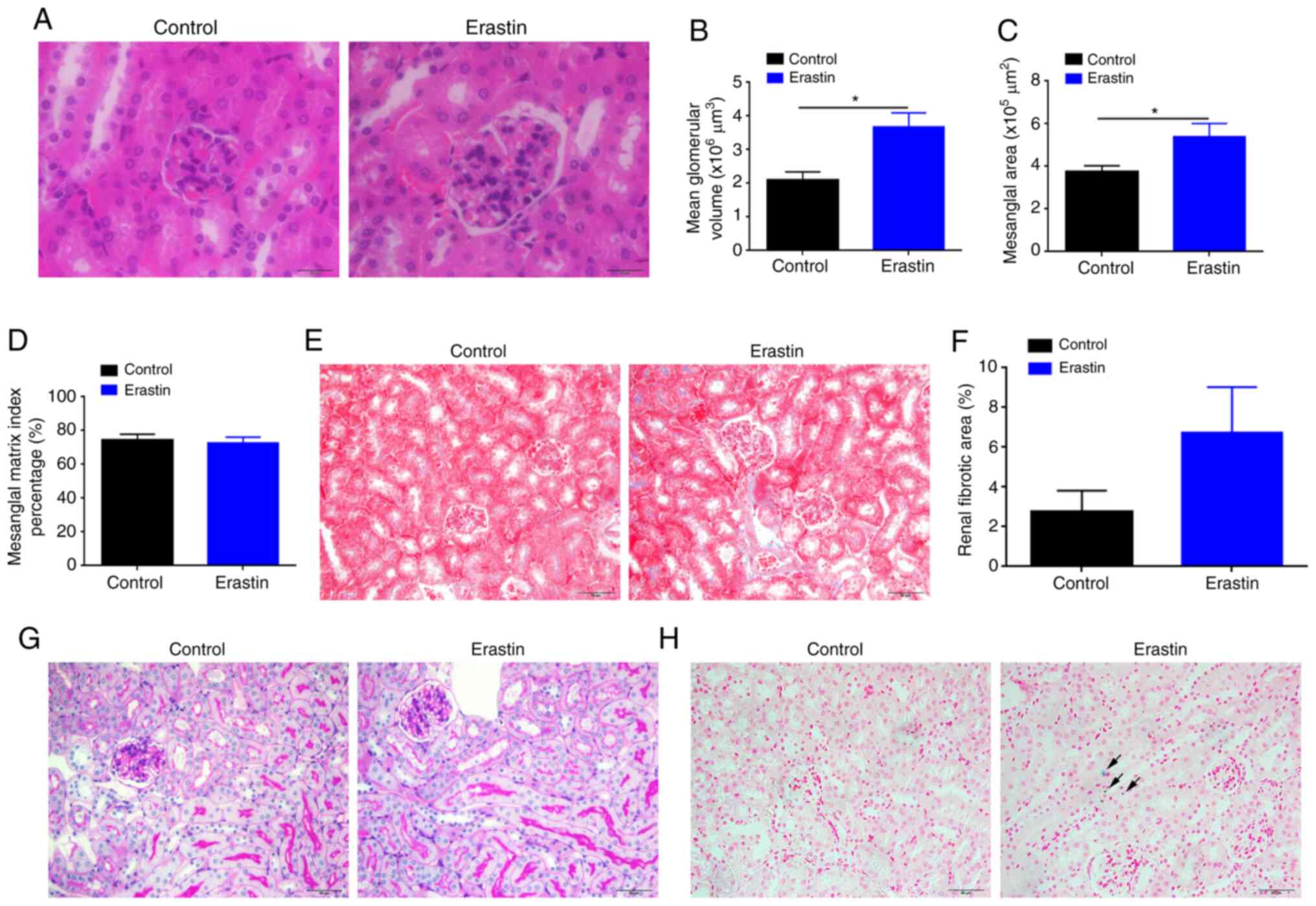
Erastin injection increases glomerular
volume of kidney
Erastin-treated mice demonstrated increased
glomerular volume with dilated glomerular capillaries (Fig. 4A). The mean glomerular volume and
mesangial area of erastin-treated mice were significantly
(P<0.05) increased 1.76- and 1.44-fold, respectively (Fig. 4B and C), compared with those of
control mice. However, the index of mesangial matrix did not change
significantly (P>0.05; Fig. 4D).
Furthermore, Masson staining demonstrated that erastin treatment
did not significantly affect the degree of renal fibrosis (Fig. 4E and F). PAS staining demonstrated
no significant raise of PAS-positive cells in glomerulus and no
abnormality in renal tubules in erastin-treated mice (Fig. 4G). Prussian blue staining indicated
that erastin-induced ferroptosis also caused mild iron deposition
in the kidneys (Fig. 4H).
Erastin injection affects iron
deposition in the spleen, but no liver
In consideration of significant changes in the
indicators of ferroptosis in the liver (Fig. 1C-E and H), the present study
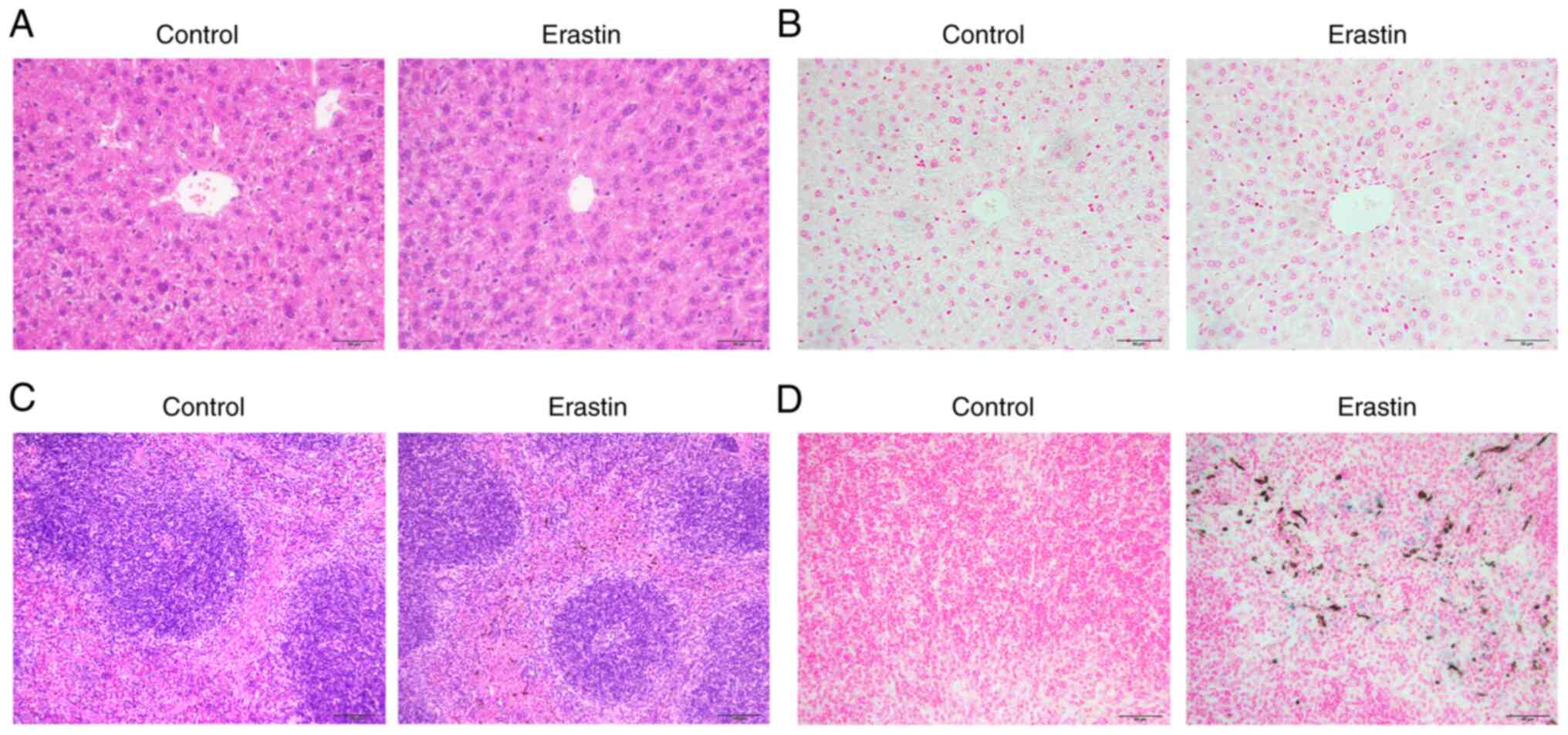
examined hepatic pathological changes by H&E staining. Notably,
no significant difference was observed between the two groups.
There was no necrosis, hemorrhage and inflammatory infiltration,
hepatocyte apoptosis and vacuolar degeneration in the liver of
erastin-treated mice (Fig. 5A).
Prussian blue staining also demonstrated no apparent iron
deposition in the liver of erastin-treated mice (Fig. 5B). The spleen consisted of the red
pulp and white pulp and the structure of splenic corpuscles was
clear in both groups (Fig. 5C).
Prussian blue staining revealed the appearance of a brown substance
and distinguished it from hemosiderin, which would be stained as
blue granules (Fig. 5D). In
addition, erastin injection also caused severe iron deposition in
the spleen (Fig. 5D).
Discussion
Ferroptosis is a recently defined programmed cell
death, which has been revealed in various organ diseases, such as
neurological disorders (16), liver
injury (17) and cardiomyopathy
(18). Ferroptosis is an
iron-dependent non-apoptotic cell death that can produced by
pharmacologically inhibiting the cysteine/glutamate antiporter,
system Xc− or directly binding and loss of activity of
GPX4 in cancer cells with high level RAS-RAF-MEK pathway activity
or p53 expression, but it does not occur in normal cells (19). A series of small molecules including
erastin have been found to induce ferroptosis in various cancer
cells. They offer the possibility of cancer therapies through
pharmacological interference with ferroptotic cell death, which has
aroused great interest in scientific and medical researches
(13). However, few studies focused
on the effects of erastin-induced ferroptosis on healthy tissues
and organs in vivo. The present study provided some
evidences of metabolic and physiologic changes of mice injected
with erastin.
According to the pharmacokinetics of erastin,
erastin has low solubility and a previous study demonstrated a poor
metabolic stability in a mouse liver microsome assay (4). The once-per-day-dosage frequency might
be too low to see a tumor shrinking effect. Consequently, the
present study conducted a preliminary in vivo study and
selected a 2-day erastin administration protocol. Following
intraperitoneal injection of erastin for 2 days, ferroptosis was
induced, as expected, with accumulation of iron and lipid peroxides
(Fig. 1). Erastin-induced
ferroptosis caused the reduction of typical red-blood-cell-related
hematological parameters, including red blood cell count,
hematocrit, hemoglobin and red blood cell distribution width
(Table I). This observation leads
to the hypothesis that erastin tended to lead to anemia in mice.
Alternation of blood indexes caused by erastin-induced ferroptosis
was specific to red blood cells, but not platelets. In addition,
increased serum iron and tissue iron were observed in the
erastin-treated group. This suggested that erastin-caused anemia
was not related to iron deficiency. This result may also hint at a
targeted role of erastin to induce ferroptosis of erythrocytes. It
has been reported that acute ingestion of large amounts of red
blood cells could direct splenic macrophages to ferroptosis
(20). Heme could mediate the
activation and death of human platelets through ferroptosis
(21). Iron constitutes the crucial
part of heme and ~70% of the body's iron is existed in red blood
cells (22). It might be the reason
that red blood cells, macrophages after robust erythrophagocytosis
and platelets toxified by free heme are more sensitive to
ferroptosis, which is an iron-dependent form of regulated cell
death (1).
In its mouse model of erastin-induced ferroptosis,
the present study observed mild infraction and iron deposition in
the brain (Fig. 2), which was
consistent with a previous study on cerebral ischemic/reperfusion
(23). Due to the presence of the
blood-brain barrier, the brain can adjust its iron status somewhat
independently of other organs (24). For the low iron content in mouse
brains, the classic Prussian blue staining had weak sensitivity for
assessing iron content (24). The
tiny blue granules observed in the cerebral sections may reflect a
change of iron level in the brain of erastin-treated mice.
Ferroptosis is observed in the intestine of
ischemia/reperfusion (8),
ulcerative colitis (25) and cystic
fibrosis (26). These studies
explore the relationship between ferroptosis and the intestine in a
pathological environment, but not in a healthy physiological state.
The present study is a clear demonstration of the role of
ferroptosis in duodenum growth as the villi were much stronger
following erastin treatment (Fig.
3), which may be attributed to improved absorption of nutrients
and shortened division cycle of intestinal stem cells. This result
may once again suggest that ferroptosis was a double-edged sword in
promoting and damaging organ growth. It has been reported that
ferroptosis might be involved in limb development by helping remove
interdigital webbing (27).
Prussian blue staining also revealed that the blue granules of
hemosiderin mainly appeared on the tip of villi in the duodenum of
erastin-treated mice (Fig. 3E). It
indicated that iron levels were increased in the epithelial cells
of duodenum. Another possible explanation for ferroptosis leading
to duodenum growth is that adequate iron supplementation may
promote intestine growth, as iron supplementation increases villus
height and depth, which is beneficial to intestinal mucosa in
piglets (28).
Unexpectedly, the present study found that
ferroptosis did not affect hepatic morphology or iron deposition in
the mouse model of ferroptosis (Fig.
5). This result was inconsistent with previous report that
ferroptosis was associated with liver injury (29), liver fibrosis (30) and nonalcoholic steatohepatitis
(31). These differences might be
attributed to the short-term injection of erastin. Notably, severe
iron deposition and unknown substance accumulation were seen in the
spleen of erastin-treated mice (Fig.
5). Ferroptosis can act on splenic macrophages (20) and cause DNA damage (32), but there was no deposition of
unknown matter. Therefore, it was hypothesized that this substance
might be secreted by the splenocytes, macrophages or other cells in
the spleen, which needs further investigation.
In summary, the present study confirmed that erastin
could induce ferroptosis in mice and revealed that erastin-induced
ferroptosis altered the blood index values, causing mild cerebral
infarction of brain and enlarged glomerular volume of kidney.
Erastin also promoted the growth of duodenal epithelium with
thicker, longer and denser villi in treated mice, but had no effect
on liver or testis. These findings provided a new perspective for
studying the role of erastin under healthy physiological
conditions.
Supplementary Material
Supporting Data
Acknowledgements
Not applicable.
Funding
This work was supported by Natural Science
Foundation of Zhejiang province of China (no. LZ20C170004,
LY19HD70003) and National Natural Science Foundation of China (no.
31872363).
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
JZ and HD were responsible for the conception and
design of the study. JZ, BX and QX performed the experiments. JZ
and BX confirm the authenticity of all the raw data. JZ, YF and HD
performed the statistical analysis. YF and HD were responsible for
funding acquisition. HD was responsible for project administration.
JZ wrote the original draft. YF and HD wrote, reviewed and edited
the manuscript. All authors read and approved the final
manuscript.
Ethics approval and consent to
participate
The experimental procedures for the mice were
approved by Animal Ethics Committee of Zhejiang University
(approval no. 20077).
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Dixon SJ, Lemberg KM, Lamprecht MR, Skouta
R, Zaitsev EM, Gleason CE, Patel DN, Bauer AJ, Cantley AM, Yang WS,
et al: Ferroptosis: An iron-dependent form of nonapoptotic cell
death. Cell. 149:1060–1072. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Tang D, Kang R, Berghe TV, Vandenabeele P
and Kroemer G: The molecular machinery of regulated cell death.
Cell Res. 29:347–364. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Xie Y, Hou W, Song X, Yu Y, Huang J, Sun
X, Kang R and Tang D: Ferroptosis: Process and function. Cell Death
Differ. 23:369–379. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Yang WS, SriRamaratnam R, Welsch ME,
Shimada K, Skouta R, Viswanathan VS, Cheah JH, Clemons PA, Shamji
AF, Clish CB, et al: Regulation of ferroptotic cancer cell death by
GPX4. Cell. 156:317–331. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Xia Y, Sun X, Luo Y and Stary CM:
Ferroptosis contributes to isoflurane neurotoxicity. Front Mol
Neurosci. 11:4862019. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Belavgeni A, Meyer C, Stumpf J, Hugo C and
Linkermann A: Ferroptosis and necroptosis in the kidneys. Cell Chem
Biol. 27:448–462. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Yamada N, Karasawa T, Kimura H, Watanabe
S, Komada T, Kamata R, Sampilvanjil A, Ito J, Nakagawa K, Kuwata H,
et al: Ferroptosis driven by radical oxidation of n-6
polyunsaturated fatty acids mediates acetaminophen-induced acute
liver failure. Cell Death Dis. 11:1442020. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Li Y, Feng D, Wang Z, Zhao Y, Sun R, Tian
D, Liu D, Zhang F, Ning S, Yao J and Tian X: Ischemia-induced ACSL4
activation contributes to ferroptosis-mediated tissue injury in
intestinal ischemia/reperfusion. Cell Death Differ. 26:2284–2299.
2019. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Dolma S, Lessnick SL, Hahn WC and
Stockwell BR: Identification of genotype-selective antitumor agents
using synthetic lethal chemical screening in engineered human tumor
cells. Cancer Cell. 3:285–296. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Bridges RJ, Natale NR and Patel SA: System
xc− cystine/glutamate antiporter: An update on molecular
pharmacology and roles within the CNS. Br J Pharmacol. 165:20–34.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Forcina GC and Dixon SJ: GPX4 at the
crossroads of lipid homeostasis and ferroptosis. Proteomics.
19:e18003112019. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Xu T, Ding W, Ji X, Ao X, Liu Y, Yu W and
Wang J: Molecular mechanisms of ferroptosis and its role in cancer
therapy. J Cell Mol Med. 23:4900–4912. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Zhao Y, Li Y, Zhang R, Wang F, Wang T and
Jiao Y: The role of erastin in ferroptosis and its prospects in
cancer therapy. Onco Targets Ther. 13:5429–5441. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Huo H, Zhou Z, Qin J, Liu W, Wang B and Gu
Y: Erastin disrupts mitochondrial permeability transition pore
(mPTP) and induces apoptotic death of colorectal cancer cells. PLoS
One. 11:e01546052016. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Hambright WS, Fonseca RS, Chen L, Na R and
Ran Q: Ablation of ferroptosis regulator glutathione peroxidase 4
in forebrain neurons promotes cognitive impairment and
neurodegeneration. Redox Biol. 12:8–17. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Sui M, Jiang X, Chen J, Yang H and Zhu Y:
Magnesium isoglycyrrhizinate ameliorates liver fibrosis and hepatic
stellate cell activation by regulating ferroptosis signaling
pathway. Biomed Pharmacother. 106:125–133. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Fang X, Wang H, Han D, Xie E, Yang X, Wei
J, Gu S, Gao F, Zhu N, Yin X, et al: Ferroptosis as a target for
protection against cardiomyopathy. Proc Natl Acad Sci USA.
116:2672–2680. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Imai H, Matsuoka M, Kumagai T, Sakamoto T
and Koumura T: Lipid peroxidation-dependent cell death regulated by
GPx4 and ferroptosis. Curr Top Microbiol Immunol. 403:143–170.
2017.PubMed/NCBI
|
|
20
|
Youssef LA, Rebbaa A, Pampou S, Weisberg
SP, Stockwell BR, Hod EA and Spitalnik SL: Increased
erythrophagocytosis induces ferroptosis in red pulp macrophages in
a mouse model of transfusion. Blood. 131:2581–2593. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
21
|
NaveenKumar SK, SharathBabu BN, Hemshekhar
M, Kemparaju K, Girish KS and Mugesh G: The role of reactive oxygen
species and ferroptosis in heme-mediated activation of human
platelets. ACS Chem Biol. 13:1996–2002. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Li X, He T, Yu K, Lu Q, Alkasir R, Guo G
and Xue Y: Markers of iron status are associated with risk of
hyperuricemia among Chinese adults: Nationwide population-based
study. Nutrients. 10:1912018. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Tuo QZ, Lei P, Jackman KA, Li XL, Xiong H,
Li XL, Liuyang ZY, Roisman L, Zhang ST, Ayton S, et al:
Tau-mediated iron export prevents ferroptotic damage after ischemic
stroke. Mol Psychiatry. 22:1520–1530. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Hanninen MM, Haapasalo J, Haapasalo H,
Fleming RE, Britton RS, Bacon BR and Parkkila S: Expression of
iron-related genes in human brain and brain tumors. BMC Neurosci.
10:362009. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Xu M, Tao J, Yang Y, Tan S, Liu H, Jiang
J, Zheng F and Wu B: Ferroptosis involves in intestinal epithelial
cell death in ulcerative colitis. Cell Death Dis. 11:862020.
View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Simões F, Ousingsawat J, Wanitchakool P,
Fonseca A, Cabrita I, Benedetto R, Schreiber R and Kunzelmann K:
CFTR supports cell death through ROS-dependent activation of
TMEM16F (anoctamin 6). Pflugers Arch. 470:305–314. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Schnabel D, Salas-Vidal E, Narváez V,
Sánchez-Carbente Mdel R, Hernández-García D, Cuervo R and
Covarrubias L: Expression and regulation of antioxidant enzymes in
the developing limb support a function of ROS in interdigital cell
death. Dev Biol. 291:291–299. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Pu Y, Li S, Xiong H, Zhang X, Wang Y and
Du H: Iron promotes intestinal development in neonatal piglets.
Nutrients. 10:7262018. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Park SJ, Cho SS, Kim KM, Yang JH, Kim JH,
Jeong EH, Yang JW, Han CY, Ku SK, Cho IJ and Ki SH: Protective
effect of sestrin2 against iron overload and ferroptosis-induced
liver injury. Toxicol Appl Pharmacol. 379:1146652019. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Kong Z, Liu R and Cheng Y: Artesunate
alleviates liver fibrosis by regulating ferroptosis signaling
pathway. Biomed Pharmacother. 109:2043–2053. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Tsurusaki S, Tsuchiya Y, Koumura T,
Nakasone M, Sakamoto T, Matsuoka M, Imai H, Yuet-Yin Kok C, Okochi
H, Nakano H, et al: Hepatic ferroptosis plays an important role as
the trigger for initiating inflammation in nonalcoholic
steatohepatitis. Cell Death Dis. 10:4492019. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Li T, Liu X, Jiang L, Manfredi J, Zha S
and Gu W: Loss of p53-mediated cell-cycle arrest, senescence and
apoptosis promotes genomic instability and premature aging.
Oncotarget. 7:11838–11849. 2016. View Article : Google Scholar : PubMed/NCBI
|