Introduction
Chronic hepatitis B virus (HBV) infection is the
main cause of different degrees of hepatic inflammation. Moreover,
chronic hepatitis can lead to the development of liver cirrhosis
and hepatocellular carcinoma (HCC) (1,2).
According to a report from the World Health Organization, ~257
million individuals are chronically infected with HBV worldwide,
and 0.74 million individuals die of HBV-related liver cirrhosis
annually (2).
Chemokines and their receptors serve essential roles
in the development of HCC (3).
C-X-C motif chemokine receptor 3 (CXCR3) belongs to the G-protein
coupled receptor 1 family, and it is expressed in a variety of
cells, such as T1 helper T cells, CD8 T cells, natural killer (NK)
T cells, NK cells, dendritic cells and some cancer cells, such as
colon and breast cancer cells (4,5). CXCR3
specifically binds with C-X-C motif chemokine ligand 9 (CXCL9) to
promote the migration and invasion of cancer cells (6). In addition, it activates the PI3K/Akt
signaling pathway to regulate cell proliferation, apoptosis and
angiogenesis (7,8). Furthermore, CXCR3 is rapidly increased
after interferon stimulation, and it promotes the migration of
CD4+ T and CD8+ T cells to the inflamed
tissues (9,10). It has been shown that the expression
of CXCR3 on T cells mainly exerts anti-tumor effects, while its
expression on tumor cells mainly promotes cancer metastasis
(11,12).
HBV causes hepatitis and HCC mainly via the host's
immune and inflammatory response (13). Innate immunity and adaptive immunity
serve important roles in HBV infection (14). Toll-like receptors (TLRs) are innate
immune recognition receptors that can resist infection by
recognizing microbial cell wall components and triggering immune
response (15,16). Myeloid differentiation primary
response protein 88 (MyD88) is a cytosolic adapter protein that is
involved in the TLR signaling pathway (17). TLR4 can activate the TIR domain
containing adaptor protein/MyD88 signal transduction pathway, and
induce the production of proinflammatory cytokines, such as IL-6,
IL-10, IL-12 and TNF-α, via NF-κB and activator protein 1 (18). In hepatocytes, the TLR/MyD88
signaling pathway is activated upon contact with HBV (19,20).
Moreover, the enhanced production of IL-6 and TNF-α could inhibit
HBV replication and prevent the death of HBV-infected hepatocytes
(21,22). Host recognition of HBV also leads to
the activation of NLR family pyrin domain containing 3 (NALP3), and
subsequently to the release of IL-1β. Abnormal secretion of IL-1β
can aggravate the inflammatory reaction, and mature IL-1β binds to
its receptor, IL-1βR, and reactivates MyD88 (23,24).
Previous studies have mainly focused on the role of
CXCR3 or TLRs in cancer, and have reported their respective roles
in the development of hepatitis B into HCC (25–27).
However, the regulatory relationship between CXCR3 and the
TLRs/MyD88 signaling pathway has been rarely reported. Thus, the
present study aimed to investigate whether CXCR3 regulated the
genes in the TLRs/MyD88 signaling pathway, and to examine the
effect of CXCR3 on the proliferation, migration and apoptosis of
hepatic stellate cells infected with HBV. The current study may
reveal the regulatory relationship between CXCR3 and the TLRs/MyD88
signaling pathway in the carcinogenesis of liver cirrhosis.
Materials and methods
Materials
The human hepatic stellate cell line LX-2 (cat. no.
BNCC341586) was obtained from BeNa Culture Collection (Neijing
Beina Chunglian Institute of Biotechnology). Rabbit anti-CXCR3
(cat. no. ab71864), rabbit anti-TLR4 (cat. no. ab13556), rabbit
anti-MyD88 (cat. no. ab133739), rabbit anti-NALP3 (cat. no.
ab214185), rabbit anti-COL1A1 (cat. no. ab34710), rabbit
anti-baculoviral IAP repeat containing 5 (BIRC5; cat. no. ab469)
and rabbit anti-GAPDH (cat. no. ab181602) antibodies were purchased
from Abcam. The goat anti-rabbit IgG (H+L) (HRP) antibody (cat. no.
ZB-2301) was from Beijing Zhongshan Goldenbridge Biotechnology Co.,
Ltd. FBS (cat. no. 10099141) and Opti-MEM™ (cat. no. 31985-062)
were purchased from Gibco (Thermo Fisher Scientific, Inc.). RIPA
buffer (cat. no. 1053) was from Applygen Technologies, Inc. DMEM
(cat. no. KGM12800S) was purchased from Nanjing KeyGen Biotech Co.,
Ltd. pCDH-CMV-MCS-EF1-copGFP-T2A-Puro vector (cat. no. P0268) was
provided by Wuhan Miaoling Biotechnology Co., Ltd.
SuperSignal® West Pico Chemiluminescent Substrate (cat.
no. 34077) was from Thermo Fisher Scientific, Inc.
Masson's Trichrome stain kit (cat. no. G1340) and
BSA (cat. no. A8020) were purchased from Beijing Solarbio Science
& Technology Co., Ltd. IL-6 (cat. no. m1058097-1) and IL-1β
(cat. no. m105859-1) ELISA kits were from Shanghai Enzyme-linked
Biotechnology Co., Ltd. The Annexin V-APC/7-AAD apoptosis kit (cat.
no. AP105-100-kit) was from Multisciences (Lianke) Biotech Co.,
Ltd. Ultrapure RNA kit (cat. no. CW0581M), HiFiScript cDNA
Synthesis kit (cat. no. CW2569M), UltraSYBR Mixture (cat. no.
CW0957M) and BCA Protein Assay kit (cat. no. CW0014S) were
purchased from CoW in Biosciences. The Cell Counting Kit (CCK)-8
(cat. no. KGA317) was from Nanjing KeyGen Biotech Co., Ltd.
Sample collection
The study was approved by The Ethics Committee of
Hwa Mei Hospital, University of Chinese Academy of Sciences
(Ningbo, China; approval no. PJ-NBEY-KY-2018-016-01), and written
informed consent was obtained from all participants. In total, 20
subjects were recruited into this study, including 10 healthy
controls, 4 patients with liver cancer and 6 patients with liver
cancer with cirrhosis. The clinical and pathological features of
the patients with cancer were summarized in Table I. Liver cancer tissues and the
paracancerous tissues were collected from 6 patients with liver
cancer who had a history of liver cirrhosis. A total of 10 ml serum
samples were collected from 10 patients with liver cancer and 10
healthy controls. The controls were healthy subjects who received
physical examinations. The patients with cancer were recruited at
Hwa Mei Hospital between January 2019 and May 2019.
 | Table I.Clinical and pathological features of
the patients with cancer. |
Table I.
Clinical and pathological features of
the patients with cancer.
| No. | Sex | Age, years | Clinical
diagnosis | Histological
type | TNM stage | Metastasis |
|---|
| 1 | Male | 63 | Malignant tumor of
liver | Moderately
differentiated HCC | I | No |
| 2 | Male | 58 | Liver tumor,
HB | Moderately
differentiated HCC | II | No |
| 3 | Male | 62 | Liver tumor, active
CHB | Poorly
differentiated HCC | II | No |
| 4 | Female | 77 | Liver tumor | Moderately
differentiated HCC | II | No |
| 5 | Male | 69 | Malignant tumor of
liver | Well-moderately
differentiated HCC | II | No |
| 6 | Male | 56 | Malignant tumor of
liver | Moderately-poorly
differentiated HCC | II | No |
| 7 | Female | 58 | Liver tumor | Well-moderately
differentiated HCC | I | No |
| 8 | Male | 60 | Liver tumor | Moderately
differentiated HCC | III | Yes (blood) |
| 9 | Male | 53 | Liver tumor | Poorly
differentiated HCC | II | No |
| 10 | Female | 77 | Liver tumor | Moderately-poorly
differentiated HCC | II | No |
The inclusion criteria were as follows: Subjects
join the study voluntarily and sign informed consent; aged 18–75
years; any sex; patients with liver cancer diagnosed by
histopathology/cytology or clinical diagnosis; ≥1 lesion that could
be measured according to the Response Evaluation Criteria In Solid
Tumors v1.1 (28); liver function
status as Child-Pugh Class A or B (score ≤7); Eastern Cooperative
Oncology Group physical status score of 0–1; life expectancy ≥12
weeks; laboratory examination findings of: i) Blood system
function, absolute neutrophil count ≥1.5×109/l, platelet
count ≥75×109/l, hemoglobin ≥90 g/l; ii) liver function,
total bilirubin ≤1.5× the upper limit of normal value (ULN), serum
albumin ≥28 g/l, alanine aminotransferase and aspartate
transaminase ≤5× ULN; and iii) renal function, serum creatinine
≤1.5× ULN, endogenous creatinine clearance rate ≥60 ml/min; normal
blood pressure or use of one antihypertensive drug can that can
make the blood pressure ≤150/90 mmHg; and the patients are willing
and able to follow up. The exclusion criteria were as follows:
Brain metastasis; tumor thrombus in the main portal vein or
inferior vena cava; presence of double or multiple cancer types;
moderate or severe ascites; history of bleeding tendency or
thrombosis; history of severe cardiovascular disease; history of
unhealed wound or ulcer, or fracture within 3 months; laboratory
abnormalities: i) Hyponatremia (sodium <130 mmol/l); baseline
serum potassium <3.5 mmol/l; ii) abnormal thyroid function, and
thyroid function that cannot be maintained in the normal range with
drugs; iii) HIV positive; pregnant or lactating women; and patients
considered unsuitable for the trial by the researchers.
Generation of stable cell line
The human hepatic stellate cell line LX-2 was
cultured in DMEM supplemented with 10% FBS. The cells were
maintained at 37°C in the presence of 5% CO2. The
sequences of the HBV (gene ID, 944566) gene were synthesized by
General Biosystems (Anhui) Co., Ltd., and inserted into a
lentiviral expression vector pCDH-CMV-MCS-EF1-copGFP-T2A-Puro. LX-2
cells were transfected with HBV lentivirus vector using 8 µg/ml
polyamine at 37°C. Cells were observed under a fluorescence
microscope (MF53; Guangzhou Mingmei Photoelectric Technology Co.,
Ltd.), and the images were taken under white and green fluorescent
light. At 72 h following transfection, cells were incubated with 5
µg/ml puromycin at 37°C for 72 h, followed by selection with 2
µg/ml puromycin. The puromycin resistant clones were tested for HBV
expression via PCR and western blotting. The positive stable clones
were maintained in the presence of puromycin.
Transient transfection
The sequences of all small interfering (si)RNAs used
in this study were: CXCR3-siRNA-F1 forward,
5′-GCUAAAUGACGCCGAGGUUTT-3′ and reverse,
5′-AACCUCGGCGUCAUUUAGCTT-3′; CXCR3-siRNA-F2 forward,
5′-AGAGAGGGCUCCAGAGGCATT-3′ and reverse,
5′-UGCCUCUGGAGCCCUCUCUTT-3′; CXCR3-siRNA-F3 forward,
5′-CUGGAGAACUUCAGCUCUUTT-3′ and reverse,
5′-AAGAGCUGAAGUUCUCCAGTT-3′; and siRNA-negative control (NC)
forward, 5′-UUCUCCGAACGUGUCACGUTT-3′ and reverse,
5′-ACGUGACACGUUCGGAGAATT-3′. For transient CXCR3 knockdown, LX-2
cells with stable HBV overexpression were transiently transfected
with CXCR3 siRNAs using Lipofectamine® 3000 (Invitrogen;
Thermo Fisher Scientific, Inc.), according to the manufacturer's
instructions. In brief, cell culture medium was replaced with
serum-free medium when cells were cultured to 70% confluence. Then,
5 µl Lipofectamine 3000 and 12.5 µl siRNA were diluted in 125 µl
Opti-MEM, and then incubated at room temperature for 5 min. The
diluted Lipofectamine was mixed with the diluted siRNA and
incubated at room temperature for 15 min. The complexes were added
to the culture medium. After 6 h, the transfection medium was
replaced with culture medium. Subsequent experiments were performed
48 h following transfection.
Reverse transcription (RT)-PCR and
RT-quantitative (q)PCR
Total RNA from the LX-2 cells was extracted with the
Ultrapure RNA kit according to the manufacturer's instructions. RT
reactions were performed with the HiFiScript cDNA Synthesis kit in
a total reaction volume of 20 µl containing 50 pg-5 µg RNA
template, 4 µl dNTP mix, 4 µl 5X RT buffer, 2 µl DTT, 2 µl primer
mix and 1 µl HiFiScript. The reaction was incubated at 42°C for 15
min, 85°C for 5 min and finally held at 4°C. The primers for HBV
were 5′-TCTCAGCAATGTCAACGACC-3′ (forward) and
5′-AATTTATGCCTACAGCCTCCT-3′ (reverse). The cDNA of HBV was
subjected to PCR, and the RT-PCR products were analyzed by
electrophoresis on a 2% agarose gel. The primers used were as
follows: CXCR3 forward, 5′-AATGACGCCGAGGTTGC-3′ and reverse,
5′-CCAGAGCCAAAGACCCACT-3′; and GAPDH forward,
5′-TGACTTCAACAGCGACACCCA-3′ and reverse,
5′-CACCCTGTTGCTGTAGCCAAA-3′. qPCR was performed using the UltraSYBR
Mixture on a CFX Connect™ Real-time PCR Detection system (Bio-Rad
Laboratories, Inc.), under the following thermocycling conditions:
Initial denaturation at 95°C for 10 min, followed by 40 cycles of
95°C for 10 sec, 58°C for 30 sec and 72°C for 30 sec, and final
extension at 72°C for 10 min. All samples were run in duplicate,
and the mRNA expression level of CXCR3 was analyzed using the
2−ΔΔCq method (29).
Western blot analysis
Protein in cells was extracted with ice-cold RIPA
buffer. The samples were then centrifuged at 13,780 × g at 4°C for
10 min, and the supernatant was transferred to a new tube. The
protein concentration was detected with a BCA Protein assay kit. A
total of 40 µg protein per lane was loaded and subsequently
separated by 10% SDS-PAGE and transferred to PVDF membranes.
Membranes were blocked in 3% non-fat milk at 37°C for 1 h before
incubation with the primary antibodies at 4°C overnight, including
rabbit anti-CXCR3 (1:1,000), rabbit anti-TLR4 (1:1,000), rabbit
anti-MyD88 (1:1,000), rabbit anti-NALP3 (1:500), rabbit anti-COL1A1
(1:1,000), rabbit anti-BIRC5 (1:5,000) and rabbit anti-GAPDH
(1:10,000). After washing with 1X TBS-Tween-20 (0.1%) three times,
each for 10 min, the membranes were incubated with the goat
anti-rabbit IgG (H+L) (HRP) secondary antibody (1:2,000) at 37°C
for 2 h. Protein bands were detected using the
SuperSignal® West Pico Chemiluminescent Substrate and
analyzed via Image-Pro Plus software (version 6.0; Media
Cybernetics, Inc.).
CCK-8 assay
The CCK-8 kit was used according to the
manufacturer's instructions. Following transfection, 10 µl CCK-8
solution was added to the 96-well plates and incubated for 2 h at
37°C. The optical density (OD) value of each well was detected at
48 and 72 h at 450 nm wavelength using an enzyme-labeled
instrument, and the cell viability was calculated.
Cell apoptosis assay
In total, 1–3×106 cells were collected
and centrifuged with 1 ml PBS twice at 210 × g for 3 min at room
temperature. Then, 5X Binding Buffer was diluted to 1X Binding
Buffer with double distilled water, and the cells were then
resuspended in 1X Binding Buffer. Subsequently, 3 µl annexin V-APC
and 5 µl 7-AAD were added to incubate the cells in the dark at room
temperature for 10 min. Then, 200 µl pre-cold 1X Binding Buffer was
added into each tube and mixed well. Cell apoptotic rates were
calculated by detecting the percentage of early and late apoptotic
cells using a flow cytometer (NovoCyte® 2060R; ACEA
Bioscience, Inc.) with NovoExpress software (version 1.3.4; Agilent
Technologies, Inc.).
Cell migration assay
At the start of the assay, cells on either side of
the would were at 95–100% confluence. A sterile 10-µl pipette tip
was used to make a single scratch, then the culture medium was
discarded and the HBV-LX-2 cells were washed three times with PBS.
Serum-free medium was added to each well, and the scratch of each
well was imaged at 0, 24 and 48 h under an inverted fluorescence
microscope (MF53; Guangzhou Mingmei Photoelectric Technology Co.,
Ltd.). The cell migration rate was calculated according to the
width of the scratch using Image-Pro Plus software (version
5.1.0.20; Media Cybernetics, Inc.).
ELISA
IL-6 and IL-1β levels in serum were assessed using
the ELISA kits according to the manufacturer's instructions.
Briefly, 50 µl Standard Substance with different concentrations at
48, 24, 12, 6, 3 and 0 µg/ml were added into the standard wells.
The samples were incubated with 100 µl enzyme labeled reagent at
37°C for 60 min. Following washing with PBS, 50 µl Developer A and
50 µl Developer B were added into each well, shaken and mixed
gently, and then the color was developed at 37°C for 15 min.
Subsequently, 50 µl termination solution was added to stop the
reaction. The OD of each well was measured at 450 nm on an
automatic microplate reader (WD-2102B; Beijing Liuyi Biotechnology
Co., Ltd.).
Masson staining
Tissues were fixed in 10% formaldehyde at 4°C for 48
h. Liver sections (4 µm) were deparaffinized, rehydrated, stained
in Weigert's iron hematoxylin solution at room temperature for 10
min and then differentiated with acid ethanol for 5–15 sec. After
washing in running water for 10 min, the slides were stained in
Masson blue solution for at 37°C for 5 min and rinsed in distilled
water, which was followed by staining in ponceau-acid fuchsin
solution at 37°C for 8 min and differentiation in phosphomolybdic
acid solution for 2 min. The slides were then transferred directly
to aniline blue solution and stained at 37°C for 2 min, washed with
weak acid working solution for 1 min and quickly dehydrated three
times in 95% ethanol and anhydrous ethanol. Finally, the slides
were sealed and observed under an optical microscope
(magnification, ×200 and ×400).
Statistical analysis
All statistical analyses were performed using SPSS
19.0 software (IBM Corp.). Data are presented as the mean ±
standard deviation of three independent experiments. One-way ANOVA
was used to compare the differences between multiple groups,
followed by the post hoc Student-Newman-Keuls test and Tukey's
test. P<0.05 was considered to indicate a statistically
significant difference.
Results
Establishment of LX-2 cell line stably
overexpressing HBV
The HBV lentivirus vector was transfected into LX-2
cells, and the cells were observed using a fluorescence microscope.
Representative images of the green fluorescent protein fluorescence
from LX-2 cells following transfection of the empty lentiviral
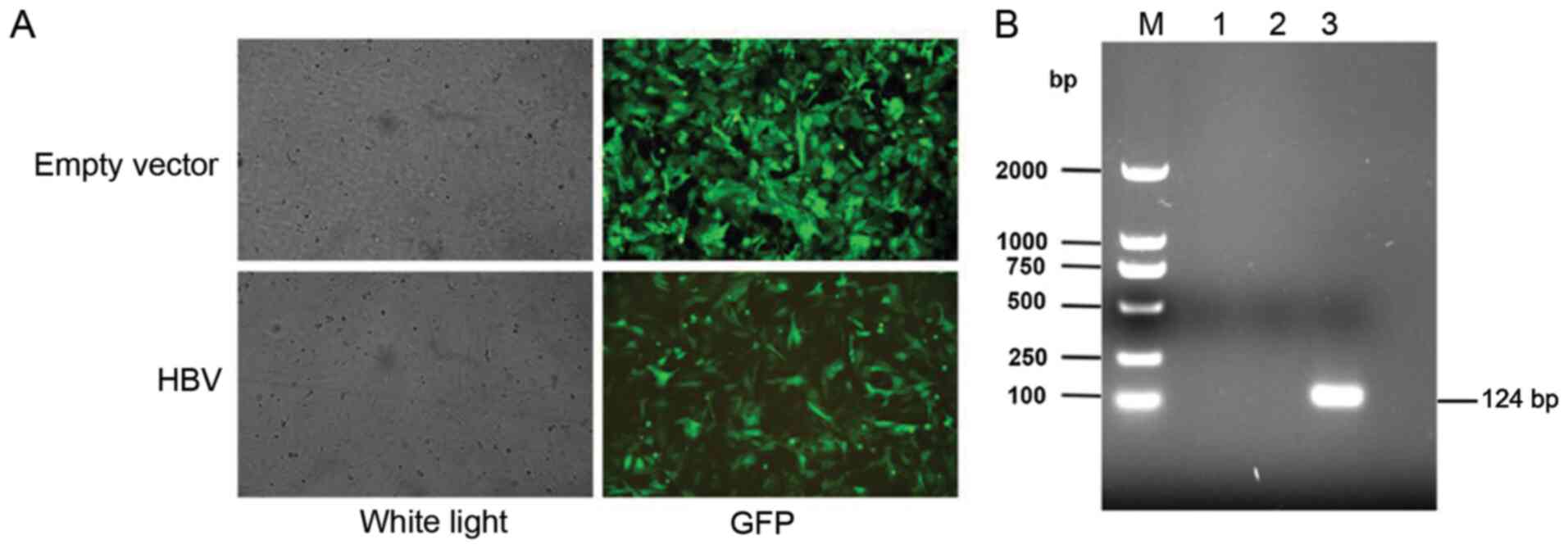
vector and the HBV lentivirus vector are shown in Fig. 1A. The HBV-overexpressing stable cell
line was selected by puromycin, and the RT-PCR results demonstrated
that HBV mRNA expression was notably increased in the
HBV-transfected cells compared with the empty lentiviral
vector-transfected cells and the control cells (Fig. 1B).
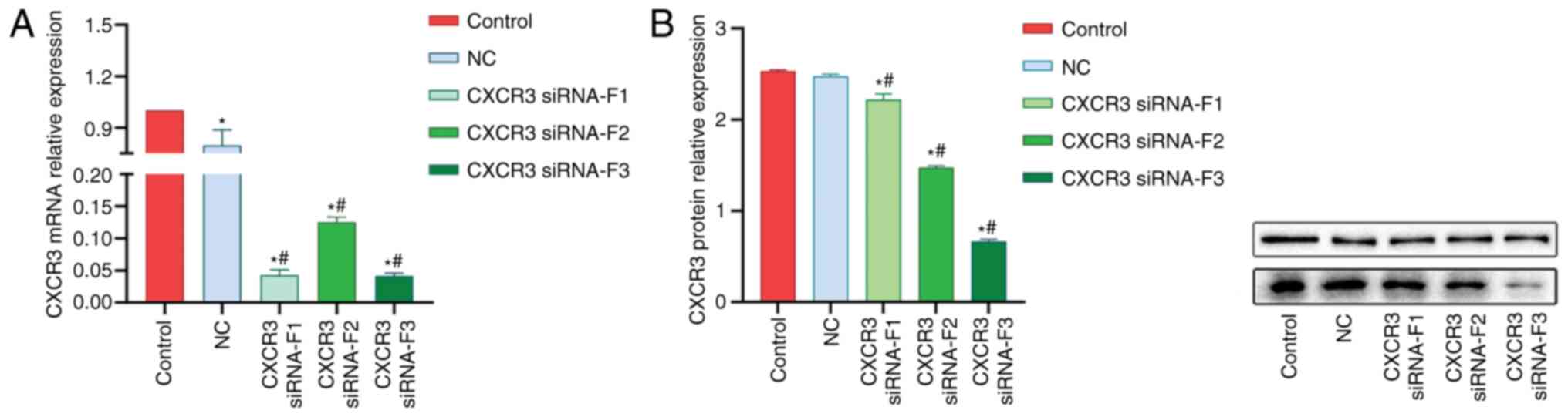
Interference of CXCR3 gene
expression
A total of three CXCR3 siRNAs were synthesized and
transfected into LX-2 cells stably expressing HBV (named HBV-LX-2
cells). The interference efficiency of siRNAs was detected via
RT-qPCR and western blot analyses. The RT-qPCR results demonstrated
that the interference efficiencies of all the three siRNAs were
>50% (Fig. 2A). The results from
western blot analysis revealed that siRNA-F3 showed the strongest
interference efficiency on CXCR3 protein expression, which was
>50% (Fig. 2B).
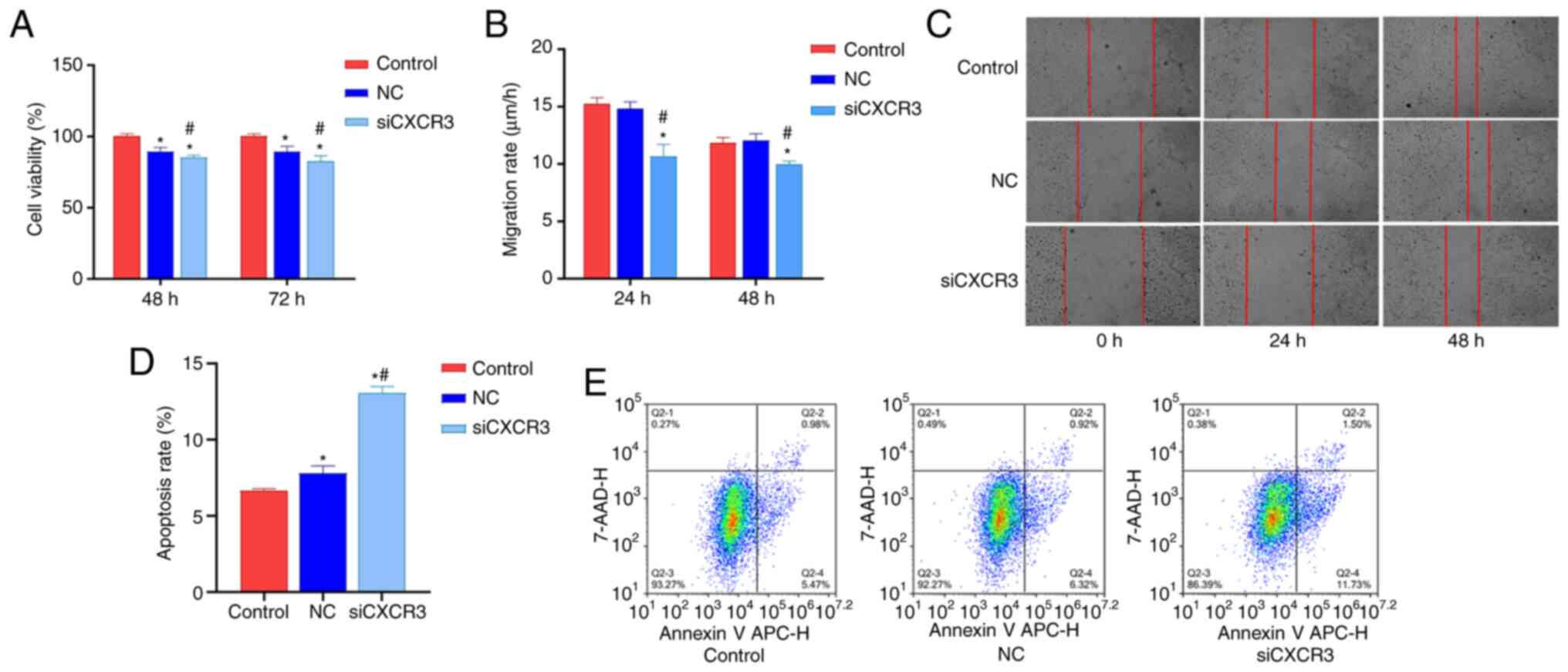
Interference with CXCR3 inhibits
HBV-LX-2 cell viability and migration and promotes cell
apoptosis
In order to examine the effect of CXCR3 on the
viability, migration and apoptosis of HBV-LX-2 cells, CXCR3
siRNA-F3 was transfected into HBV-LX-2 cells. The cells were then
subjected to CCK-8 assays, and cell migration and cell apoptosis
analyses. Compared with the control group and NC group, the
viability of the siCXCR3 group was significantly decreased at 48
and 72 h (Fig. 3A), and the cell
migratory rate significantly decreased at 24 and 48 h (Fig. 3B and C). Moreover, the cell
apoptotic rate was significantly increased in the siCXCR3 group
compared with the control group and the NC group (Fig. 3D and E).
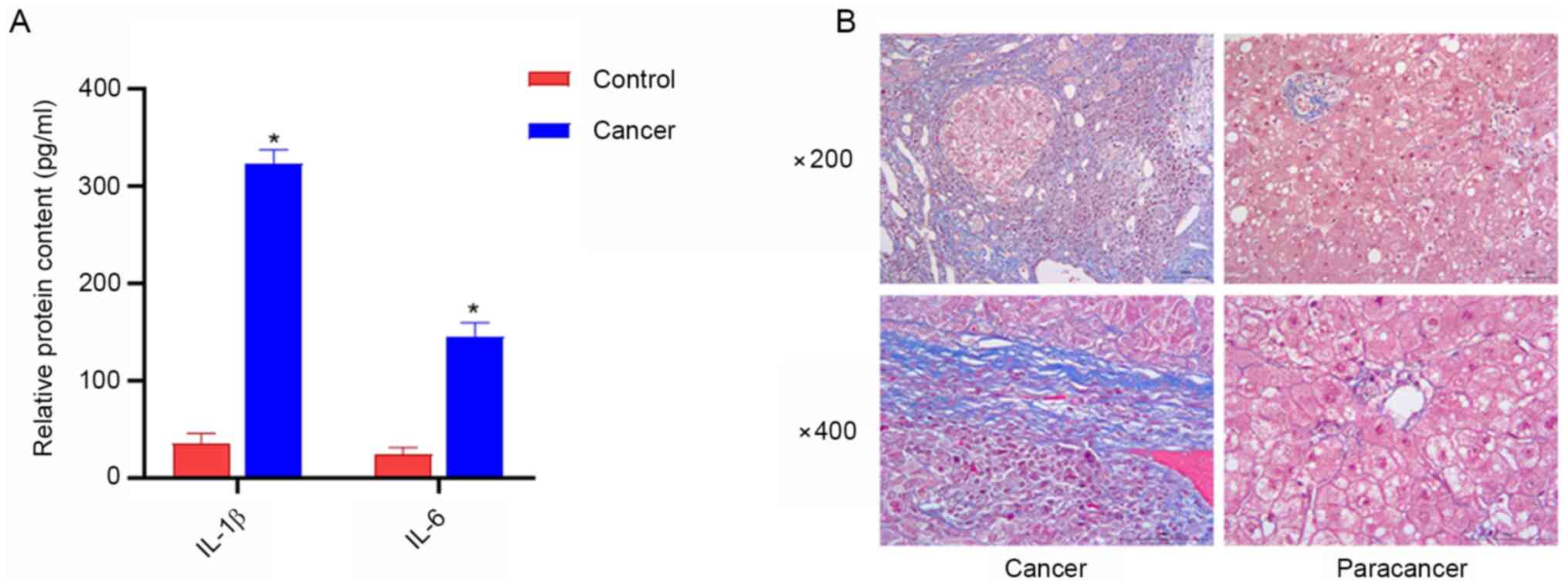
Serum levels of IL-1β and IL-6 are
increased in patients with liver cancer, and a high amount of
collagen fibers exists in liver cancer tissues
ELISA was used to detect serum levels of IL-β and
IL-6 in patients with liver cancer, and the degree of liver
fibrosis was examined using Masson staining. The results of ELISA
demonstrated that the serum levels of IL-1β and IL-6 in patients
with liver cancer were higher compared with those in the healthy
controls (Fig. 4A). Furthermore,
Masson staining revealed that liver cancer tissues contained a
large number of collagen fibers, while the content of collagen
fibers in paracancerous tissues was reduced (Fig. 4B).
Interference with CXCR3 inhibits the
expression levels of COL1A1 and the proteins in the TLRs/MyD88
pathway in HBV-LX-2 cells
CXCR3 siRNA was transfected into LX-2 cells
overexpressing HBV, and the cells were subjected to western blot
analysis. The results demonstrated that the protein expression
levels of TLR4, MyD88, COL1A1 and BIRC5 were significantly lower in
the siCXCR3 group compared with those in the control group and the
NC group (Fig. 5B). Compared with
the control group, the expression level of NALP3 protein was
significantly decreased in the siCXCR3 group. However, there was no
significant difference in NALP3 protein expression between siCXCR3
group and NC group.
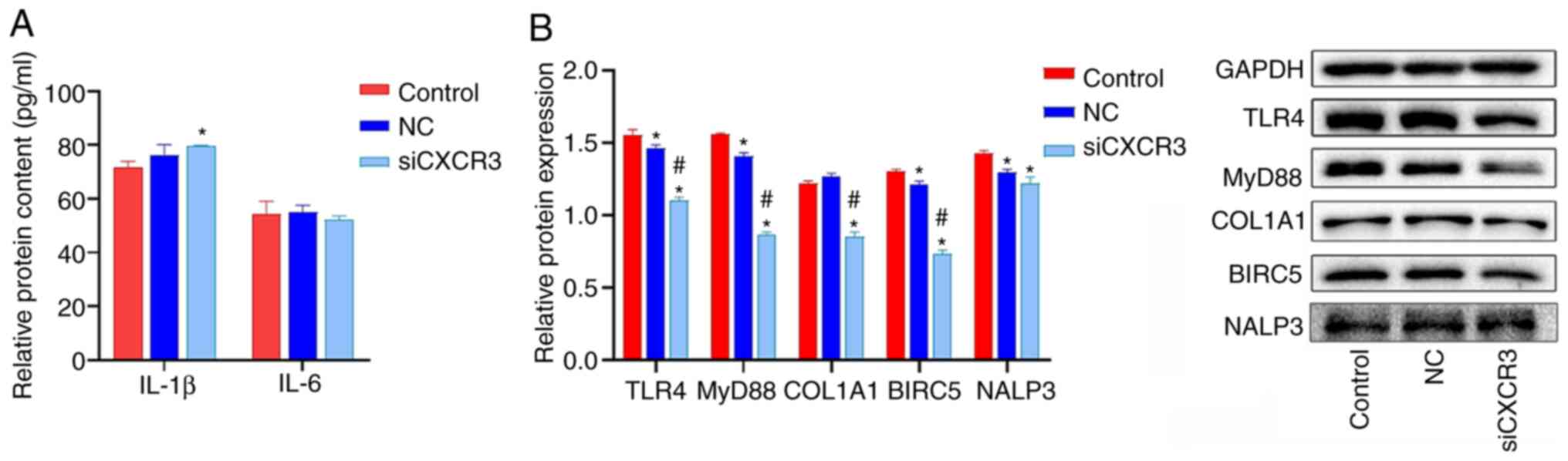 | Figure 5.Effect of siCXCR3 on the expression
levels of COL1A1 and the proteins in the TLRs/MyD88 pathway in
HBV-LX-2 cells. (A) Interference with CXCR3 increases the
expression of IL-1β in HBV-LX-2 cells, as determined by ELISA. (B)
Interference with CXCR3 inhibits the protein levels of TLR4, MyD88,
COL1A1 and BIRC5 in HBV-LX-2 cells, as determined via western
blotting. *P<0.05 vs. Control; #P<0.05 vs. NC.
CXCR3, C-X-C motif chemokine receptor 3; siRNA/si, small
interfering RNA; NC, siRNA-negative control; TLR, Toll-like
receptor; MyD88, myeloid differentiation primary response protein
88; COL1A1, collagen type I α 1 chain; BIRC5, baculoviral IAP
repeat containing 5; NALP3, NLR family pyrin domain containing
3. |
The levels of IL-6 and IL-1β in HBV-LX-2 cells were
detected via ELISA. IL-1β and IL-6 levels in LX-2 cells were lower
than those in the serum of patients with liver cancer (Figs. 4A and 5A). IL-6 levels were significantly
increased in the siCXCR3 group compared with the control group, but
there was no significant difference in the level of IL-6 between
siCXCR3 group and NC group. Compared with the control group and NC
group, the level of IL-1β was not significantly changed in the
siCXCR3 group (Fig. 5A).
Discussion
In the process of hepatic fibrosis, hepatic stellate
cells differentiate into proliferative and contractile
myofibroblasts, as well as secrete COL1A1 and other proteins that
constitute pathological fibrous tissues (30). Fibrosis can lead to liver cancer,
and CXCR3 serves an important role in this process. In previous
studies, CXCR3 has been reported to promote the proliferation and
migration of cancer cells, including gastric, colorectal and tongue
squamous cell carcinoma cells (6–8). The
present study established a HBV-overexpressing stable cell line
using hepatic stellate cells LX-2. RNA interference-mediated
knockdown of CXCR3 was performed in HBV-LX-2 cells, and the results
demonstrated that CXCR3 was significantly decreased at both the
mRNA and protein levels in cells following transfection with CXCR3
siRNA. Furthermore, it was found that interference with the
expression level of CXCR3 could inhibit cell proliferation and
migration and promote cell apoptosis. Previous studies have shown
that CXCR3 was upregulated in primary and metastatic cancer cells
(8,31,32).
In glioma cells, CXCR3 activates PI3K/Akt via the G protein subunit
to increase intracellular calcium level, thereby promoting the
proliferation, migration and invasion of cancer cells (31). Moreover, CXCR3 is highly expressed
in gastric cancer tissues with lymph node metastasis, suggesting
that CXCR3 can promote lymph node metastasis (8). In tongue squamous cell carcinoma,
CXCL9/CXCR3 promotes the invasion and proliferation of cancer cells
by activating the Akt signaling pathway (32). All these reports suggest that CXCR3
can promote cell proliferation, migration and invasion. In the
present study, the results demonstrated that interfering with the
expression level of CXCR3 inhibited cell proliferation and
migration. This finding was consistent with the role of CXCR3 in
other cancer cells (8,31,32),
indicating that interference with CXCR3 can inhibit the
carcinogenesis of hepatic stellate cells overexpressing HBV.
Chronic HBV infection may progress to liver
cirrhosis and hepatocellular carcinoma (33). In the present study, Masson staining
was performed on the liver cancer tissues of liver cancer cases
with HBV infection. A large number of collagen fibers were observed
in the liver cancer tissues, but the amount of collagen fibers in
adjacent tissues was small, indicating that the excessive
accumulation of collagen fibers in liver cancer tissues may lead to
liver cirrhosis. Moreover, it was found the serum levels of IL-1β
and IL-6 were increased in the patients with liver cancer. These
findings suggested that the liver cancer induced by hepatitis B can
promote the expression of inflammatory factors. Excessive IL-1β and
IL-6 will aggravate the inflammatory reaction and promote the
development of liver cirrhosis (24). In the current study, LX-2 cells
stably overexpressing HBV were cultured in vitro. It was
found that the levels of IL-1β and IL-6 in LX-2 cells were much
lower than those in the serum of patients with liver cancer.
Moreover, CXCR3 inhibition did not significantly downregulate the
levels of IL-1β and IL-6 in HBV-LX-2 cells.
Previous studies have reported that CXCR3 is
involved in the activity of cancer cells, such as proliferation,
migration and invasion (6–8). In addition, CXCR3 can induce the
chemotaxis of immune cells, especially T cells (34,35).
Upregulated CXCR3 can promote the migration of T cells to specific
organs (9,36), induce the secretion of cytokines
such as IL-6 and IFN-γ, and cause inflammation. In the current
experiments, only one cell line was cultured in vitro.
Therefore, CXCR3 could not exert its effect on T cell chemotaxis to
induce inflammatory response. It has been reported that IL-1β is
expressed not only in immune cells, but also in other cells
(37). The expression and
activation of IL-1β is controlled by two different signaling
pathways (38,39). The first signaling pathway is
NF-κB-mediated IL-1β section, and this pathway is induced by TLR
ligands. Another signaling pathway is the cleavage of the precursor
IL-1β to mature IL-1β via caspase-1, which is mediated by NALP3
(38,39). In the present study, it was found
that TLR4 and MyD88 were significantly downregulated in cells
following CXCR3 interference, but the expression level of NLRP3, as
well as the levels of IL-1β and IL-6 did not change significantly.
This finding suggested that in HBV-LX-2 cells, the expression of
IL-1β and IL-6 was mediated by NLRP3, but not by TLR4/MyD88.
Moreover, it was identified that CXCR3 could not directly regulate
the expression level of NLRP3, so IL-1β and IL-6 were almost
unchanged in vitro.
Although there was no significant change in
inflammatory factors, COL1A1 protein expression was significantly
downregulated in HBV-LX-2 cells following transfection with CXCR3
siRNA, indicating that CXCR3 knockdown could decrease the synthesis
of collagen and reduce the degree of liver fibrosis. Previous
studies have shown that COL1A1 expression was upregulated in
gastric cancer cells and promoted cell invasion and metastasis
(40,41). In cervical cancer, COL1A1 can
inhibit the apoptosis of cancer cells via the PI3K pathway, while
interference with COL1A1 can significantly promote the apoptosis of
cancer cells (42). The present
results were consistent with those of previous studies in that
interference with CXCR3 downregulates COL1A1 protein expression,
promotes apoptosis and inhibits the migration of HBV-LX-2 cells
(8,32,40,42).
Similar results were observed with regards to BIRC5 protein
expression. BIRC5 is an anti-apoptotic protein, which is widely
expressed in malignant tumors, such as gastric cancer, lung cancer,
colon cancer and ovarian cancer (43,44).
BIRC5 can affect cell division and proliferation, as well as
inhibit apoptosis of cancer cells (45). Moreover, interference with BIRC5
triggers the apoptosis of cancer cells (46). The present study demonstrated that
CXCR3 siRNA could downregulate BIRC5 protein expression, inhibit
cell proliferation and promote cell apoptosis, suggesting that
interference with CXCR3 could inhibit the differentiation of
HBV-LX-2 cells to cancer cells.
The present study only detected the expression level
of CXCR3 in HBV-LX-2 cells and solely investigated the role of
CXCR3 at the cellular level. As a limitation of this study, protein
expression studies were not performed on patient tissues. Further
studies may be conducted in clinical tissues and animal model
tissues to perform correlation analysis between CXCR3 and the
inflammatory factors, genes in the TLR4/MyD88 signaling pathway and
other factors involved in the process of liver cirrhosis to liver
cancer.
In conclusion, the present study demonstrated that
CXCR3 knockdown can inhibit the expression levels of TLR4/MyD88,
BIRC and COL1A1, as well as inhibit cell proliferation and
migration and promote cell apoptosis, so as to inhibit the
development of liver cirrhosis to liver cancer. It was also found
that knockdown of CXCR3 had little effect on the expression levels
of NALP3 and the related inflammatory factors IL-1β and IL-6. Thus,
the role of CXCR3 in regulating the inflammatory factors IL-1β and
IL-6 during the process of liver cirrhosis to liver cancer should
be further studied.
Acknowledgements
Not applicable.
Funding
This study was funded by Medical and Health Project
of Zhejiang Province (grant no. 2019KY179) and Joint Construction
of Key Medical Disciplines in Zhejiang Province (grant no.
2016-7).
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
DW and GY conceived and designed the experiments, as
well as prepared the manuscript. BC, XS, YM and JL conducted the
experiments. AH and YH contributed to data collection and analyzed
the data. DW and GY are confirm the authenticity of all the raw
data. All authors read and approved the final manuscript.
Ethics approval and consent to
participate
This study was approved by the Ethics Committee of
Hwa Mei Hospital, University of Chinese Academy of Sciences
(Ningbo, China). All patients provided written informed consent in
compliance with the code of ethics of the World Medical Association
(Declaration of Helsinki).
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Bray F, Ferlay J, Soerjomataram I, Siegel
RL, Torre LA and Jemal A: Global cancer statistics 2018: GLOBOCAN
estimates of incidence and mortality worldwide for 36 cancers in
185 countries. CA Cancer J Clin. 68:394–424. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
McMahon BJ: Chronic hepatitis B virus
infection. Med Clin North Am. 98:39–54. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Liang CM, Chen L, Hu H, Ma HY, Gao LL, Qin
J and Zhong CP: Chemokines and their receptors play important roles
in the development of hepatocellular carcinoma. World J Hepatol.
7:1390–1402. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Tokunaga R, Zhang W, Naseem M, Puccini A,
Berger MD, Soni S, McSkane M, Baba H and Lenz HJ: CXCL9, CXCL10,
CXCL11/CXCR3 axis for immune activation-A target for novel cancer
therapy. Cancer Treat Rev. 63:40–47. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Karin N: CXCR3 ligands in cancer and
autoimmunity, chemoattraction of effector T cells, and beyond.
Front Immunol. 11:9762020. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Humblin E and Kamphorst AO: CXCR3-CXCL9:
It's all in the tumor. Immunity. 50:1347–1349. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Friedl P and Alexander S: Cancer invasion
and the microenvironment: Plasticity and reciprocity. Cell.
147:992–1009. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Zhou H, Wu J, Wang T, Zhang X and Liu D:
CXCL10/CXCR3 axis promotes the invasion of gastric cancer via
PI3K/AKT pathway-dependent MMPs production. Biomed Pharmacother.
82:479–488. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Kohlmeier JE, Cookenham T, Miller SC,
Roberts AD, Christensen JP, Thomsen AR and Woodland DL: CXCR3
directs antigen-specific effector CD4+ T cell migration
to the lung during parainfluenza virus infection. J Immunol.
183:4378–4384. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Marshall A, Celentano A, Cirillo N,
McCullough M and Porter S: Tissue-specific regulation of
CXCL9/10/11 chemokines in keratinocytes: Implications for oral
inflammatory disease. PLoS One. 12:e01728212017. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Abron JD, Singh NP, Murphy AE, Mishra MK,
Price RL, Nagarkatti M, Nagarkatti PS and Singh UP: Differential
role of CXCR3 in inflammation and colorectal cancer. Oncotarget.
9:17928–17936. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Zhang Y, Xu L and Peng M: CXCR3 is a
prognostic marker and a potential target for patients with solid
tumors: A meta-analysis. Onco Targets Ther. 11:1045–1054. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Villarrubia VG, Alvarez-Mon M, Chirigos MA
and Herrerías JM: Hepatitis B virus (HBV) and the
inflammatory/immune response. I. The natural environment of the
antigen presentation and immunologic chaos induced by the virus.
Rev Esp Enferm Dig. 89:919–928. 1997.(In Spanish). PubMed/NCBI
|
|
14
|
Fisicaro P, Valdatta C, Boni C, Massari M,
Mori C, Zerbini A, Orlandini A, Sacchelli L, Missale G and Ferrari
C: Early kinetics of innate and adaptive immune responses during
hepatitis B virus infection. Gut. 58:974–982. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Nosratabadi R, Alavian SM, Zare-Bidaki M,
Shahrokhi VM and Arababadi MK: Innate immunity related pathogen
recognition receptors and chronic hepatitis B infection. Mol
Immunol. 90:64–73. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Gao D and Li W: Structures and recognition
modes of toll-like receptors. Proteins. 85:3–9. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Takeda K and Akira S: TLR signaling
pathways. Semin Immunol. 16:3–9. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Firmal P, Shah VK and Chattopadhyay S:
Insight Into TLR4-mediated immunomodulation in normal pregnancy and
related disorders. Front Immunol. 11:8072020. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Mylona EE, Mouktaroudi M, Crisan TO, Makri
S, Pistiki A, Georgitsi M, Savva A, Netea MG, van der Meer JW,
Giamarellos-Bourboulis EJ and Joosten LA: Enhanced interleukin-1β
production of PBMCs from patients with gout after stimulation with
toll-like receptor-2 ligands and urate crystals. Arthritis Res
Ther. 14:R1582012. View
Article : Google Scholar : PubMed/NCBI
|
|
20
|
Guo H, Jiang D, Ma D, Chang J, Dougherty
AM, Cuconati A, Block TM and Guo JT: Activation of pattern
recognition receptor-mediated innate immunity inhibits the
replication of hepatitis B virus in human hepatocyte-derived cells.
J Virol. 83:847–858. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Hösel M, Quasdorff M, Wiegmann K, Webb D,
Zedler U, Broxtermann M, Tedjokusumo R, Esser K, Arzberger S,
Kirschning CJ, et al: Not interferon, but interleukin-6 controls
early gene expression in hepatitis B virus infection. Hepatology.
50:1773–1782. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Xu Y, Köck J, Lu Y, Yang D, Lu M and Zhao
X: Suppression of hepatitis B virus replication in Tupaia
hepatocytes by tumor necrosis factor alpha of Tupaia belangeri.
Comp Immunol Microbiol Infect Dis. 34:361–368. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Gupta P and Barthwal MK: IL-1 β genesis:
The art of regulating the regulator. Cell Mol Immunol. 15:998–1000.
2018. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Mitroulis I, Kambas K and Ritis K:
Neutrophils, IL-1β, and gout: Is there a link? Semin Immunopathol.
35:501–512. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Elia G and Fallahi P: Hepatocellular
carcinoma and CXCR3 chemokines: A narrative review. Clin Ter.
168:e37–e41. 2017.PubMed/NCBI
|
|
26
|
Xu HZ, Liu YP, Guleng B and Ren JL:
Hepatitis B virus-related hepatocellular carcinoma: Pathogenic
mechanisms and novel therapeutic interventions. Gastrointest
Tumors. 1:135–145. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Zare-Bidaki M, Tsukiyama-Kohara K and
Arababadi MK: Toll-like receptor 4 and hepatitis B infection:
Molecular mechanisms and pathogenesis. Viral Immunol. 27:321–326.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Eisenhauer EA, Therasse P, Bogaerts J,
Schwartz LH, Sargent D, Ford R, Dancey J, Arbuck S, Gwyther S,
Mooney M, et al: New response evaluation criteria in solid tumours:
Revised RECIST guideline (version 1.1). Eur J Cancer. 45:228–247.
2009. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Brenner DA, Waterboer T, Choi SK,
Lindquist JN, Stefanovic B, Burchardt E, Yamauchi M, Gillan A and
Rippe RA: New aspects of hepatic fibrosis. J Hepatol. 32 (Suppl
1):S32–S38. 2000. View Article : Google Scholar
|
|
31
|
Zhou YQ, Liu DQ, Chen SP, Sun J, Zhou XR,
Xing C, Ye DW and Tian YK: The role of CXCR3 in neurological
diseases. Curr Neuropharmacol. 17:142–150. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Li Z, Liu J, Li L, Shao S, Wu J, Bian L
and He Y: Epithelial mesenchymal transition induced by the
CXCL9/CXCR3 axis through AKT activation promotes invasion and
metastasis in tongue squamous cell carcinoma. Oncol Rep.
39:1356–1368. 2018.PubMed/NCBI
|
|
33
|
Papatheodoridis GV, Chan HL, Hansen BE,
Janssen HL and Lampertico P: Risk of hepatocellular carcinoma in
chronic hepatitis B: Assessment and modification with current
antiviral therapy. J Hepatol. 62:956–967. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Zeng Z, Li L, Chen Y, Wei H, Sun R and
Tian Z: Interferon-γ facilitates hepatic antiviral T cell retention
for the maintenance of liver-induced systemic tolerance. J Exp Med.
213:1079–1093. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Benigni G, Dimitrova P, Antonangeli F,
Sanseviero E, Milanova V, Blom A, van Lent P, Morrone S, Santoni A
and Bernardin G: CXCR3/CXCL10 axis regulates neutrophil-NK cell
cross-talk determining the severity of experimental osteoarthritis.
J Immunol. 198:2115–2124. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Masopust D, Vezys V, Marzo AL and
Lefrançois L: Preferential localization of effector memory cells in
nonlymphoid tissue. Science. 291:2413–2417. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Rébé C and Ghiringhelli F: Interleukin-1β
and cancer. Cancers (Basel). 12:17912020. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Walsh JG, Muruve DA and Power C:
Inflammasomes in the CNS. Nat Rev Neurosci. 15:84–97. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Dinarello CA: Immunological and
inflammatory functions of the interleukin-1 family. Ann Rev
Immunol. 27:519–550. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Li AQ, Si JM, Shang Y, Gan LH, Guo L and
Zhou TH: Construction of COL1A1 short hairpin RNA vector and its
effect on cell proliferation and migration of gastric cancer cells.
Zhejiang Da Xue Xue Bao Yi Xue Ban. 39:257–263. 2010.(In Chinese).
PubMed/NCBI
|
|
41
|
Sun H: Identification of key genes
associated with gastric cancer based on DNA microarray data. Oncol
Lett. 11:525–530. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Liu S, Liao G and Li G: Regulatory effects
of COL1A1 on apoptosis induced by radiation in cervical cancer
cells. Cancer Cell Int. 17:732017. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Ambrosini G, Adida C and Altieri DC: A
novel anti-apoptosis gene, survivin, expressed in cancer and
lymphoma. Nat Med. 3:917–921. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Gunaldi M, Isiksacan N, Kocoglu H,
Okuturlar Y, Gunaldi O, Topcu TO and Karabulut M: The value of
serum survivin level in early diagnosis of cancer. J Cancer Res
Ther. 14:570–573. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Wheatley SP and Altieri DC: Survivin at a
glance. J Cell Sci. 132:jcs2238262019. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Li F, Aljahdali I and Ling X: Cancer
therapeutics using survivin BIRC5 as a target: What can we do after
over two decades of study? J Exp Clin Cancer Res. 38:3682019.
View Article : Google Scholar : PubMed/NCBI
|